
The Axes of Divergence for the Evolutionary Radiation of Notothenioid Fishes in Antarctica


Department of Biomedical Sciences, Ohio University, Athens, OH 45701-2979, USA
Diversity 2024, 16(4), 214; https://doi.org/10.3390/d16040214
Submission received: 25 January 2024
/
Revised: 20 March 2024
/
Accepted: 21 March 2024
/
Published: 30 March 2024
(This article belongs to the Special Issue Diversity in 2024)
Abstract
:Notothenioid fishes, a perciform group, radiated in the cold shelf waters around the Antarctic continent and the 110 species dominate fish diversity, abundance, and biomass at levels of ≈77%, 92%, and 91%, respectively. This occurred in a locality with frequent glaciomarine cycles that fragmented and obliterated habitats, disrupted ecosystems, and made parts of the high latitude shelves periodically uninhabitable. The notothenioid radiation encompasses three stages and 10 axes: for the habitat stage, divergence in (1) depth and (2) biotope, meaning subdivisions within the pelagic and benthic realms; for the morphology stage, divergence in (3) body size, (4) body density based on proportions of skeletal and adipose tissues, (5) body shape, (6) trophic morphology, specifically head morphology related to feeding (jaws, teeth, head size, and pharyngeal gape), and (7) neuromorphology (brain and sensory systems); and for the communication stage, divergence in (8) fecundity and egg size, (9) sexual dichromatism and dimorphism in body and fins, and (10) parental care behavior associated with nesting. There was an antecedent Eocene fossil fauna that did not include notothenioids and that has minimal taxonomic representation in the modern fauna. Liparids (snailfishes) and zoarcids (eelpouts) also diversified in Antarctic waters but with minimal divergence. Hypotheses are offered as to why these radiations played out the way they did and why notothenioids are dominant.
1. Introduction
The interaction between geophysical and biological processes underpins the nature and composition of the modern fauna of the world. This interplay has given rise to distinctive faunas in areas that became tectonically isolated during geologic history. Antarctica is such a place. It is still uncertain why the modern Antarctic fish fauna is the way it is—vastly different than the only known antecedent fauna and now overwhelmingly dominated by a group of perciforms, the notothenioids (Figure 1). Faunal changes over time are among the most interesting but least understood natural events on Earth and, in the case of notothenioids obscured by a host of background factors, including a dubious fossil record, periodic fluctuations in climate and habitat availability, and the abundant historical presence of other cold-tolerant groups with antifreeze, gadiforms, for example, that could have radiated instead of notothenioids. That this played out on the Antarctic shelf, one of the most naturally disturbed regions on Earth (Barnes & Conlan 2007, p. 11) [1], makes it all the more intriguing.
The continental fragmentation driven by the plate tectonic regime is positively correlated with increasing global marine biodiversity throughout the Phanerozoic (Zaffosa et al., 2017) [4]. These historical or abiotic factors initially create new eco-space and opportunities for radiation to occur (Benton 2009) [5]. Notothenioids originated in waters of the western portion of the Gondwanan coast during the Late Cretaceous, ≈96 million years (Ma) ago (Near et al., 2015) [2]. During the Paleocene (66–56 Ma) and Eocene (56–33.9 Ma), sea floor spreading and the associated oceanographic changes resulted in the isolation of Antarctica in a south polar position in the Oligocene at 30 Ma (Anderson 1999) [6]. During the course of these events, a taxonomically diverse Eocene fish fauna became extinct but, in tectonic isolation and with reduced competition, notothenioids diversified and diverged as the climate became colder (Near et al., 2012) [7]. The terrestrial East Antarctic Ice Sheet is grounded above sea level (Anderson 1999, p. 6) [6] and has historically been more stable than the marine-based West Antarctic Ice Sheet. The West Antarctic Ice Sheet has been especially dynamic in the past 5 Ma, with advances as far as the shelf break in the Ross Sea at the Last Glacial Maximum (Pollard and DeConto, 2009; Anderson et al., 2014) [8,9]. At least some marine-based sectors of the East Antarctic Ice Sheet were also subject to growth and collapse during the Miocene (23.3–5.3 Ma) and Pliocene (5.3–2.6 Ma) (Pekar and DeConto, 2006; McKay, 2014; Scherer et al., 2016) [10,11,12]. These frequent glaciomarine cycles fragmented and obliterated habitats, disrupted ecosystems, and made parts of the high-latitude shelves periodically uninhabitable (Barnes and Conlan 2006) [1]. Therefore, when considering available habitat and the radiation of notothenioids, it should be recognized that historically, the Antarctic climate has not been invariant. Under such conditions, organismal and body system adaptations to a steady state environment and specific trophic resources would likely have prevented notothenioids from capitalizing on new ecological opportunities, likely the most important factor in determining how radiation proceeds (Schluter, 2000, pp. 69, 239) [13].
Notothenioids dominate diversity, abundance and biomass (Figure 2) in both the benthic and pelagic realms of the high-latitude shelf and upper slope around Antarctica (DeWitt, 1970; Hubold and Ekau, 1987; Ekau, 1990; Eastman and Hubold, 1999; Donnelly et al., 2004; Eastman, 2005; Koubbi et al., 2010; Causse et al., 2011; Hanchet et al., 2013) [14,15,16,17,18,19,20,21,22]. Although the polar oceans lack the species richness of coral-dominated tropical marine regions like the Indo-Pacific, they are “present-day hotspots of species formation” with high-latitude taxa having a more rapid rate of speciation, exemplified by notothenioids, zoarcids, and liparids (Rabosky et al., 2018) [23]. There was skepticism when it was proposed that the adaptive radiation and species flock concepts were applicable to a marine fish group that, instead of being confined to a lake, occupied a large and seemingly unrestricted environment like the Antarctic shelf and slope (Eastman and McCune, 2000; Eastman, 2005) [19,24]. A species flock is a monophyletic group of ecologically diverse, closely related species that have evolved within a single macrohabitat, frequently a lake basin. Doubts about the validity of this concept in Antarctic waters have now waned as there has been repeated confirmation, employing multiple lines of evidence and approaches, that bolster the rationale for concluding that notothenioids, or components thereof, are paradigms of adaptive radiation and species flock formation (Janko et al., 2011; Matschiner et al., 2011, 2015; Lautrédou et al., 2012; Lecointre et al., 2013; Wilson et al., 2013; Colombo et al., 2015; Chenuil et al., 2018; Bowen et al., 2020) [25,26,27,28,29,30,31,32,33]. The current hypothesis (Near et al., 2012, 2015) [2,7] holds that during the past 25 million years, and especially from 10–5 Ma, the Antarctic continental shelf and slope have hosted irregular bursts of diversification and divergence that led to modern notothenioids occupying inshore to upper slope depths and most of the energetically feasible niches on the substrate and in the water column. Recent research also suggests that much of the high-latitude avian and mammalian megafauna, including some penguins, lobodontine seals, and Minke and blue whales, appeared between 10–4 Ma (Árnason et al., 2006, 2018; Fulton and Strobeck, 2010; Crame, 2018; Vianna et al., 2020) [34,35,36,37,38].
Until recently there had been little effort devoted to the important step of identifying the stages and axes of the notothenioid radiation. Vertebrate radiations sometimes progress through similar stages or phases, diverging sequentially in habitat utilization, trophic morphology, and sensory communication as represented by sexual selection (Streelman et al., 2002; Streelman and Danley, 2003; Gavrilets and Losos, 2009) [39,40,41]. The axes of the habitat stage are abiotic; those of the morphology and communication stages are biotic. In this chapter I am proposing that the notothenioid radiation, meaning the five Antarctic families or «cryonotothenioids» shown in Figure 1, involves these axes: for the habitat stage, divergence in (1) depth and (2) biotope, meaning subdivisions within the pelagic and benthic realms; for the morphology stage, divergence in (3) body size, (4) body density based on proportions of various skeletal and adipose tissues, (5) body shape, (6) trophic morphology, specifically oral and pharyngeal morphology related to feeding behavior (jaws, teeth, head size, and pharyngeal gape) and (7) neuromorphology (brain and sensory systems); and for the communication stage, divergence in (8) fecundity and egg size, (9) sexual dichromatism and dimorphism in body and fins and (10) parental care behavior associated with nesting. The depth, biotope, size, and body density axes have been documented (Eastman et al., 2014; Eastman 2017, 2019, 2020) [42,43,44,45]. My emphasis here is to briefly review axes (1–5). I will add data to the trophic morphology axis (6) and provide an overview of the neuromorphology axis (7). I will highlight morphological aspects of organismal and organ system divergence. There is considerable information on aspects of reproduction for individual notothenioid species, and axes (8–10) of the communication stage are now known (La Mesa et al., 2020 [46]. Given their recent thorough treatment, they are not described again here. Future work could be directed toward contributing data to axes with limited information, such as the body shape and trophic morphology axes, or towards identifying new axes. I conclude the chapter with an overview of the Eocene fossil fish fauna because a historical perspective is beneficial in evaluating the emergence and dominance of notothenioids instead of another cold-tolerant group.
Taxonomic note. I recognize 140 species of notothenioids, with 110 Antarctic in distribution and 30 non-Antarctic (Eastman and Eakin 2021) [47]. Some aspects of the classification and taxonomy follow Near et al. (2012, 2013, 2015, and 2018) [2,3,7,48] in (1) recognizing a reduced and newly constituted Perciformes that includes notothenioids, liparids, and zoarcids among ≈2000 other species; (2) recognizing the Brazilian flathead, Percophis brasiliensis, as the phylogenetically basal notothenioid; and (3) employing a colloquial rank-free name «cryonotothenioids» for the five primarily Antarctic families formerly known as “the Antarctic clade”—the group that radiated and composes 91% (128/140) of notothenioid diversity. Use of left («) and right (») guillemets or double chevrons indicate that this name, and that of the rank-free fossil group «mesetaichthys», are not compliant with the International Code of Zoological Nomenclature (Sheiko 2019, pp. 48–49) [49]. These names are also not capitalized.
2. The Stages and Axes of the «Cryonotothenioid» Radiation
2.1. Habitat Stage—Axis 1: Depth, Represented by Maximum Depth
Background. In spite of the absence of primary production, the ocean depths are the largest living space on earth and therefore provide an axis for divergence and diversification in the vertical dimension (Haedrich, 1996) [50]. Divergence along a depth gradient has been documented even in relatively shallow ocean habitats, for example at 0–600 m in the case of species of the northeast Pacific scorpaenid (rockfish) genus Sebastes (Ingram, 2011) [51]. Antarctic ichthyologists recognized that many «cryonotothenioids» possessed greater than expected maximum depths and depth ranges (Nybelin, 1947; Andriashev, 1965, 1987) [52,53,54] and that some species could be considered eurybathic, although not approaching the depths inhabited by some phylogenetically older deep-sea taxa that reach ≈8000 m (Linley et al., 2016) [55]. In addition, many near-shore «cryonotothenioids» also have ranges encompassing outer shelf and upper slope depths (DeWitt, 1971) [56].
Eighty-six percent of the Southern Ocean seafloor is >1000 m deep (Griffiths et al., 2014) [57]. Closer to the continent the mass of the ice sheet and glacial erosion deepened the High Antarctic continental shelves to a mean of 500 m (Anderson, 1999) [6]. Near the coast, outlet glaciers have gouged inner shelf depressions that reach depths of 1200 m. In West Antarctic and sub-Antarctic areas, however, there is less glacial ice and hydrostatic depression is not as great because the West Antarctic Ice Sheet is marine-based rather than terrestrial like that of East Antarctica. In non-Antarctic locations where some «cryonotothenioids» are found, for example, the Falklands/Malvinas Islands, southern South America, and Tasmania, shelf depths are the same as those in most other shelf areas in the world.
Maximum depths. Data are available for 128 of 140 notothenioid species that collectively range from 0 to 2941 m (Figure 3), the intertidal to the upper continental slope (Eastman, 2017) [43]. Eighty percent have maximum depths of <1000 m, 15% are 1000–2000 m, and 5% are 2000–2941 m. If a maximum depth or depth range of ≥1500 m is used to distinguish deeper-living species, thereby eliminating most outer-shelf species with ranges extending onto the upper slope at ≈1200 m, 13 species, or 10%, fall into this category—four nototheniids, four bathydraconids (Bathydraco spp.), three artedidraconids of the genus Pogonophryne, and two channichthyids. In the case of shallower-living species, many are phylogenetically basal species of the non-Antarctic genera Pseudaphritis, Halaphritis, and Bovichtus. Among «cryonotothenioids», the sub-Antarctic genus Harpagifer is the most shallow-dwelling taxon. The five species of Bathydraco range at a maximum depth between 1250 m to 2941 m, with a mean of 2098 m. Bathydraco scotiae is the deepest living species at 2941 m. The deep sea is usually demarcated by depths ≥1000 m (Angel, 1997) [58] and deep-sea fishes have mean maximum depths of occurrence at or below this depth (Priede and Froese, 2013) [59]. Employing this definition, about 25% of Antarctic and sub-Antarctic «cryonotothenioids» could be considered deep sea.
Depth ranges. Depth ranges are maximum depth minus minimum depth. Among «cyronotothenioids» the shallowest-living species are members of the genera Patagonotothen (all non-Antarctic with one exception) and Harpagifer, a primarily intertidal/inshore group with a sub-Antarctic distribution. Trematomus is also of interest with respect to the habitation of both shallow and deep waters. Trematomus has a circum-Antarctic distribution in both the seasonal and permanent pack ice zones. Most of the 15 species live at 400–700 m, although T. loennbergii (65–1243 m) and T. lepidorhinus (99–1658 m) have more extensive depth ranges and exhibit neuromorphological disparity (Eastman & La Mesa 2021) [60]. Depth ranges for artedidraconids, bathydraconids, and channichthyids also include one to a few deep-living (≥1500 m) genera in each family.
Depth and species diversity. Cumulative species versus depth plots (Figure 4) show that diversity for non-Antarctic notothenioids is greatest at 50 m and then falls off steeply while that for sub-Antarctic species peaks at 100 m with a range of maximum diversity at 100–300 m. For Antarctic species diversity peaks at 500 m, with the 300–600 m increment encompassing the maximum. This unimodal curve is evident only for Antarctic species and likely reflects the evolutionary divergence of these species into the deeper waters of the outer shelf and upper slope. However, it should be noted that, based on data from a photographic survey of the outer shelf and upper slope of the western Antarctic Peninsula, at 700–999 m the diversity and abundance shift from being dominated by «cryonotothenioids» to domination by macrourids (rattails) of the genus Macrourus and zoarcids (Amsler et al., 2016) [61].
2.2. Habitat Stage—Axis 2: Biotopes, with Percentage Buoyancy Values (%B) Used as Proxies for Subdivisions within the Pelagic and Benthic Realms
Background. Although notothenioids lack a swim bladder, their divergence from the ancestral benthic biotope and into the water-column habitats is a fundamental attribute of the radiation (Eastman, 1985a, 1993) [62,63]. The four defined biotopes are based on the mean percentage buoyancy (%B) measurements for each of the 59 species where these values are known (Near et al., 2012; Eastman, 2020) [7,45]. The percentage buoyancy measurement, abbreviated hereafter as %B, is expressed as:
where Wwater is the weight of the fish in local seawater and Wair is the weight of the fish in the air (DeVries and Eastman 1978; Near et al., 2003) [64,65]. A smaller %B value indicates a fish is more buoyant than one with a larger value. It is important to realize that a measurement of %B is not a determination of buoyancy or density per se, but rather the relative weight of a fish in water compared to its weight in air. The means of density reduction are considered later in the density axis of the morphology stage. Here, the %B value is used as a proxy for habitat in the sense that fish with the least weight to support in water (greater buoyancy) will be occupants of the pelagic biotope, with the converse applicable to heaviest fish and the benthic biotope.
%B = (Wwater/Wair) × 102,
Biotopes are inferred from %B values. A traditional definition of a biotope is used here: a geographical unit of a habitat that can be delimited by boundaries and is characterized by its biota (Lincoln et al., 1998, p. 42) [66]. Because depth is a separate axis of divergence in the habitat stage, no specific depths are implied for the biotopes, and definitions are also irrespective of the standard depth zones for the ocean. Notothenioids occupy these biotopes, subdivisions of habitat, defined as follows (Eastman, 2020) [45]:
- Pelagic biotope (%B = 0–0.5), 5% of species. The water column is irrespective of depth. In some species, neutral buoyancy may not be permanent and is contingent on the availability of lipid-rich prey.
- Semipelagic biotope (%B = 1.3–2.0), 10% of species. The water column and the substrate. Some species previously considered cryopelagic or epibenthic, based on ecomorphological measurements, are here included as occupying the semipelagic or demersal biotopes.
- Demersal biotope (%B = 2.4–4.6), 73% of species. The substrate or water column near the substrate, irrespective of depth. The term benthopelagic is not used because it limits the implied habitat depth to within 100 m of the substrate (Lincoln et al., 1998, p. 37) [66]. Demersal more accurately reflects the ecological plasticity of the species that occupy this biotope.
- Benthic biotope (%B = 5.0–7.0), 12% of species. The substrate, including the intertidal zone in areas where this exists. Species occupying this biotope are frequently in contact with the substrate.
The scatterplot in Figure 5 shows the delimitation of the four biotopes using mean %B values as proxies for habitats that range from pelagic to benthic. Species with the lowest values have reduced density and neutral buoyancy (%B = 0) and are inhabitants of the pelagic biotope, and those with the highest values (%B = 5–7) inhabit the benthic biotope. Seventy-three percent of %B values for species are within the range of the demersal biotope (2.4–4.6%). It is important to note that the biotope of a species is not contingent on foraging localities or signatures obtained from stable isotopes of carbon and nitrogen. Most «cryonotothenioids», regardless of the biotope, will opportunistically feed on pelagic resources. Cryopelagic and epibenthic are accurate descriptors when inferring the body shape and feeding mode of species based on ecomorphological measurements (Klingenberg and Ekau, 1996) [67], but they do not pertain to the habitat stage of the radiation.
Ontogenetic change in buoyancy is typical. Ontogenetic variation in the percentage buoyancy reflects the growth processes over the life history of a species. This is attributable to the differential growth of organ systems with differing densities, a general characteristic of vertebrate development. Fishes have indeterminate growth and, while growth never ceases, musculoskeletal growth slows after sexual maturity and is surpassed by the growth of viscera, especially the lower-density tissues of the reproductive and gastrointestinal systems. Therefore, as body size increases from the juvenile through the adult stage, there is an ontogenetic decrease in the mean density of the fish that is reflected in an increase in buoyancy (a lower %B value). The demersal sister species Notothenia coriiceps and N. rossii provide an example (Eastman et al., 2011) [68]. Figure 6 is a scatter plot for percentage buoyancy relative to standard length (SL) in a sample composed of small juveniles to adults of both species. There is considerable dispersion in the data, especially for N. rossii, because of the measurement error associated with the difficulty of weighing these small specimens (<10 cm SL) under water. A quadratic model fitted to the data suggests a curvilinear relationship with minimum buoyancy (greater %B) at 12–20 cm SL in both species and buoyancy increasing (lower %B) with growth. Larvae and early juvenile stages of both species are pelagic and presumably close to neutral buoyancy. However, in adults, the mean %B of N. coriiceps (4.3%) is significantly greater (t = 12.242, p < 0.0001) than that of N. rossii (3.8%). This is consistent with what is known about their morphology and ecology and reflects the differing activity levels of the two species. Notothenia coriiceps is relatively sedentary and benthic in its feeding preferences (Barrera-Oro and Casaux, 1990; North, 1996) [69,70], while N. rossii is semipelagic, migratory, and regularly feeds on pelagic prey (Casaux et al., 1990; Barrera-Oro et al., 2019) [71].
The external morphology and biotopes that are inferred from measurements of %B are sometimes discordant. Nototheniops larseni has a streamlined compressed body, laterally located eyes, a terminal mouth, and silvery coloration, all characteristics of a species that dwells and possibly feeds in the water column (Targett, 1981; DeWitt et al., 1990; Eastman, 1993; Bushula et al., 2005) [63,72,73,74]. Gobionotothen gibberifrons, on the other hand, is a benthic browser with a depressed body shape and notable ventral flattening, a subterminal mouth, dorsally located eyes, and a mottled pigmentation pattern. It is frequently found on mud bottoms where it is a “slurp-feeder” on infauna and its intestine contains mud and small fragments of rock that are inadvertently swallowed as it grubs through the sediment (Targett, 1981; Daniels, 1982) [72,75]. Based on the external morphology of these two species, L. larseni would be expected to have a relatively low %B value given that its appearance suggests that it dwells in the water column. However, the bottom-grubbing G. gibberifrons is more buoyant (%B = 4.54, demersal biotope) than L. larseni (%B = 5.62, benthic biotope). The percentages of skeletal mass to body mass are similar at 1.93% and 2.04%, respectively (Eastman et al., 2014) [42], and the explanation for the difference in %B values is obscure. Diets indicate that Nototheniops larseni feeds primarily on krill and other water-column organisms (Permitin, 1970; Permitin and Tarverdiyeva, 1972; Barrera-Oro and Tomo, 1987; Takahashi and Iwami, 1997) [76,77,78,79], but other research shows that it also feeds on benthos (Shust and Pinskaya, 1978; Daniels, 1982; Gröhsler, 1994) [75,80,81]. While G. gibberifrons does browse infauna, it also feeds on active organisms like krill when available (Kock, 1985; DeWitt et al., 1990; Takahashi and Iwami, 1997) [73,79,82]. Therefore, in spite of their distinctly different external morphologies and percentage buoyancies, both species are ecologically plastic and opportunistic in their feeding localities and behaviors and, as proxies for subdivisions of the habitat, measurements of %B do capture this.
2.3. Morphology Stage—Axis 3: Body Size, as Represented by Maximum Total Length
Background. Evolutionary radiations frequently have an axis involving organismal size, including the classic example of Darwin’s finches in the Galápagos Islands (Grant and Grant, 2008) [83]. Body size, as represented by length, has a “pervasive” effect on a number of aspects of life history (Losos, 2009, p. 259) [84]. Size influences diet, especially the ability to consume larger prey items, a crucial capability in maximizing net energy return (Wainwright & Richard 1995) [85]. It also has a substantial influence on the potential for dispersal and migratory capability (Knouft and Page, 2003) [86] and, to a disproportionately large extent, on reproductive output (Barneche et al., 2018) [87]. An especially large species in the Antarctic (Dissostichus mawsoni) plays an essential role as both predator and prey in a local food web (Ainley et al., 2021) [88].
Andriashev (1965) [53] noted differences in average lengths among four Antarctic families and found that harpagiferids were the smallest and channichthyids the largest. Balushkin (1984) [89] measured 23 species of nototheniids, grouped them into 15 cm bins and noted that >50% of species were 20–35 cm in total length. Kock (1992) [90], also employed 15-cm bins and observed that most nototheniids are <45 cm whereas about two-thirds of channichthyids are >45 cm. A recent comprehensive analysis of total length data, employing 10-cm bins, shows that size, represented by length, is an axis of divergence among «cryonotothenioids» (Eastman, 2019) [44].
Total lengths. The following serves as a frame of reference for considering the size of notothenioids relative to other actinopterygians. Miniature and small fishes are <1 cm and <10 cm, respectively (Moyle and Cech, 2004; Helfman et al., 2009) [91,92], and there are only 52 species of actinopterygians considered large at >2 m (Paxton, 1998) [93], including swordfishes (Xiphiidae) at 4.5 m and, the largest of all, the oarfishes (Regalecidae) at 8 m (Roberts, 2012; McClain et al., 2015) [94,95].
Maximum total lengths are available for 138 notothenioid species (Eastman, 2019) [44]. Median and mean total lengths are 26.6 cm and 33.5 cm. Figure 7 is a frequency distribution for total length in 10-cm bins, and also delineates the four size categories recognized here. The distribution peaks at 20–30 cm with 32% (44/138) of species, a feature that is less apparent with the 15-cm bins used previously. The inclusion of one bin on either side of the peak encompasses 71% (98/138) of species that are medium-sized at 10–39 cm. Twenty-one percent (29/138) are in the medium-large category, and only 7% (9/138) and 2% (2/138) are in the small and large categories, respectively.
Figure 8 shows that species in the five «cryonotothenioid» families encompass the entire range of divergence in size among notothenioids—a distinguishing feature of adaptive radiation. Most «cryonotothenioids» are of medium size, and species of the genera Harpagifer (5.7 cm for H. nybelini) and Dissostichus (225 cm for D. eleginoides) are the small and large outliers (Figure 9). The phylogenetically basal Percophis brasiliensis (74 cm) and Eleginops maclovinus (80 cm), the sister group of the «cryonotothenioids» are medium–large.
Perspective on body size. The magnitude of the 39-fold difference between the smallest and largest «cryonotothenioids» is not remarkable when viewed with respect to actinopterygians in general. A > 50-fold difference is seen in labriforms (wrasses), ≈100-fold in serranids (sea basses), and >200-fold in characiforms (characins) and cyprinids (minnows) (Nelson, 2006; Nelson et al., 2016) [96,97]. What is unusual, however, is that the «cryonotothenioid» size range occurs in a relatively small clade of 128 species, primarily Antarctic in distribution. The non-notothenioid taxa used for comparison are more speciose (500–3000 species) and have near-global marine or multi-continental freshwater distributions. An additional factor that may have constrained the divergence of small «cryonotothenioids» is that many of the prime habitats where small fishes are frequently found do not exist in shallow high-latitude Antarctic waters. For example, there are no coral and rocky reefs with interstices for hiding, nor are there salt marshes, eelgrass beds, or tide pools.
2.4. Morphology Stage—Axis 4: Body Density, Based on Relative Proportions of Skeletal and Adipose Tissues
Background. Notothenioids do not have a swim bladder, the organ of buoyancy in actinopterygian fishes. Under conditions where its presence is not adaptive, it has been independently lost in numerous clades of primarily benthic teleosts including notothenioids (McCune and Carlson 2004) [98]. Based on the higher-level taxa in McCune and Carlson (2004) [98], and using the count of current species in Nelson et al. (2016) [97], about 20% of teleost species (5617/29,585) lack a swim bladder. This count includes 407 species of liparids that were apparently not included by McCune and Carlson (2004) [98].
Densities of tissues. As evidenced by the different percentage buoyancy values, there has been a divergence in body density among «cryonotothenioids». Without a swim bladder, alteration in density is accomplished through the static lift provided by body tissues with a combined average density that approaches that of seawater, at least in the case of the neutrally buoyant species. Table 1 provides the densities of the major body constituents. With the exception of skeletal tissues, the densities of fish tissues are reasonably close to the density of seawater and the addition of lipids to the body reduces overall density. However, reduction in the proportion and therefore mass of bone, with its content of dense hydroxyapatite, amounting to 60–70% of dry bone mass in actinopterygians (Meunier, 2002, p. 71) [99], is also essential in altering organismal density. Hydrodynamic lift produced by swimming is of little consequence in «cryonotothenioids» because of the sedentary nature of many species, where inactivity can encompass as much as 97% of a 24-h period (North 1996; Zimmermann & Hubold 1998) [70,100]. Furthermore, most routine swimming does not produce lift because it is a drag-based labriform locomotion directed horizontally, meaning an anterior-posterior rowing stroke propels the fish forward rather than a dorsal-ventral flapping or lift-producing stroke similar to that of a bird in flight.
Skeletal masses of notothenioids. Relative dry skeletal masses are available for a sample of 54 individuals representing 20 species and six of the nine families (Eastman et al., 2014) [42]. These are shown in Figure 10 along with relative skeletal masses for three species of non-notothenioid perciforms. The skeletal mass is not available for the phylogenetically basal Percophis brasiliensis; however, its skeleton is well-ossified and adults do not possess substantial amounts of cartilage or a partially persistent notochordal canal (Odani et al., 2006 [107]; Eastman, personal observations). Another phylogenetically basal species, Bovichtus diacanthus, also has a well-ossified skeleton (Figure 11 top) and a percentage skeletal mass similar to that of some percids and serranids (3–5%). A value of ≈3.5% separates the non-notothenioids and B. variegatus from all other notothenioids (Figure 10). The non-Antarctic Eleginops maclovinus (Figure 11 middle), the sister species of the «cryonotothenioids», also has a relatively low percentage of skeletal mass and clusters among these Antarctic species at ≤2%. A light skeleton is, therefore, a phylogenetically persistent trait in «cryonotothenioids». The most important organismal manifestation of this is that it lowers the average body density and this contributes to an increase in buoyancy (a lower %B value).
Bone mineral density is not reduced. It has been assumed that bone mineral density, meaning the volumetric density of hydroxyapatite, is reduced in the bones of «cryonotothenioids», especially in channichthyids. However, bone mineral density, as inferred from various techniques (Meunier, 2002; Meunier et al., 2018; Ashique et al., 2022) [99,108,109], is similar to that in other teleosts. They are not osteopenic. Instead the entire skeletal system of «cryonotothenioids» has become less dense through neotenic retention that results in ontogenetic displacements of the relative proportions of bone, cartilage, and notochordal tissue, a relatively common occurrence among fishes (Schaeffer, 1961; Gosline, 1971, pp. 11, 89–90; Witten and Hall 2015) [110,111,112]. The pedomorphic «cryonotothenioid» larvae, especially those of channichthyids, begin life with minimal bone and there is no loss of the small amount of existing bone during ontogeny. All «cryonotothenioid» larvae are pelagic (Loeb et al., 1993; Andrew et al., 1995; Rodrigues et al., 2013) [113,114,115], and the influence and significance of pedomorphy are now recognized at levels ranging from the macroevolutionary (Balushkin, 1984, pp. 127–128) [89], to the relative proportions of skeletal tissues, especially the persistence of cartilage in lieu of bone (Voskoboinikova, 2001, 2007, 2010; Voskoboinikova et al., 2017) [116,117,118,119] and finally to the expression of bone and cartilage collagen genes (Albertson et al., 2010; Postlethwait et al., 2016) [120,121].
The body density axis. The body density axis reflects a reduction in bone mass and the addition of adipose tissue, specifically, the alteration of (1) the type and amount of bone present, (2) the amount of persisting cartilage, (3) the amount of persisting notochordal tissue in lieu of vertebral bone and (4) the capacity for storing lipid in expanded adipose tissue or larger lipid sacs with cellular perimeters (DeVries and Eastman, 1978; Eastman and DeVries, 1981a, 1982, 1989; Eastman et al., 2014; Chen et al., 2019) [42,64,122,123,124,125].
- Reduction in bone mass—predominance of various forms of cancellous rather than compact bone. The type of bone, whether cancellous (light and spongy) or compact (heavy and dense), influences its density by as much as 4-fold (Wainwright et al., 1976, p. 167) [126]. Cavities in cancellous bone decrease the bone mass but also provide space for low-density lipid that further decreases overall body density (Pelster, 1998) [127]. Examples of this are seen in the vertebral centra of many «cryonotothenioids». In Notothenia coriiceps and N. rossii the relative skeletal masses are 2.45% and 1.65%, respectively (Eastman et al., 2014) [42]. In both species compact bone in the vertebral centra is confined to the periphery and adjacent to the notochordal vacuoles (Figure 12e). The majority of the centrum is composed of cancellous bone with adipocytes occupying cavities of various sizes. However, there are different degrees of bone sponginess, and this can decrease or increase bone mass and overall body density. In N. coriiceps (Figure 12a,c,f), the volume of the bone space is less than the volume of cavity space whereas the situation is reversed in N. rossii (Figure 12b,d). Surface area measurements from histological sections (Figure 12c,d) show that the bone-space: cavity-space ratios are 44:56 for N. coriiceps and 63:37 for N. rossii. Thus, N. rossii has more bone and less lipid-containing cavity space, and this accounts for the significantly greater relative mass of the vertebral column in this species—28% of skeletal mass versus 24% in N. coriiceps (t = –3.766, p < 0.02) (Eastman et al., 2011) [42]. A more substantial vertebral bone mass may be a response to the increased stress on the column experienced during subcarangiform locomotion by the relatively more active N. rossii. This is paradoxical in the sense that the overall percentage skeletal mass and percentage buoyancy values are lower in N. rossii, but regional specialization does exist in the skeletal system.Figure 12. Bone of the vertebral centra of the sister species N. coriiceps (a,c,e) and N. rossii (b,d). (a,b) Left lateral views of the centra of the last abdominal and first caudal vertebrae of similarly sized specimens. Both species have predominantly cancellous bone, but the mesh is finer (smaller cavities and larger bony trabeculae) in N. rossii. Original magnifications, ×7.0 and ×7.5. (c,d) Histological sections of centra show the distribution of red-staining bony trabeculae and lipid-filled spaces in the centrally located cancellous bone. Compact bone (asterisks) of the amphicelous centra is located at the periphery adjacent to notochordal cavities containing notochordal vacuoles (NV). Long axes of spaces marked by daggers are 420 µm and 225 µm, respectively. ×27 and ×27. (e) A midsagittal section of ananterior caudal vertebra of N. coriiceps, sanded to show the limited extent of compact bone (asterisks) and its location around the notochordal cavities with the remainder of the centrum consisting of cancellous bone. This species has a small notochordal canal (NC) in the middle of each centrum. ×12. (f) Parasagittal histological section of an anterior caudal vertebra of N. coriiceps showing spongy bone and smooth-walled lipid-filled cavities. Arrows indicate accumulation of stain at growth checks in bone. The long axis of space marked by the dagger is 420 µm. ×83. From Eastman et al. (2011 [68], Figure 4), © Inter-Research, 2011, with permission.Figure 12. Bone of the vertebral centra of the sister species N. coriiceps (a,c,e) and N. rossii (b,d). (a,b) Left lateral views of the centra of the last abdominal and first caudal vertebrae of similarly sized specimens. Both species have predominantly cancellous bone, but the mesh is finer (smaller cavities and larger bony trabeculae) in N. rossii. Original magnifications, ×7.0 and ×7.5. (c,d) Histological sections of centra show the distribution of red-staining bony trabeculae and lipid-filled spaces in the centrally located cancellous bone. Compact bone (asterisks) of the amphicelous centra is located at the periphery adjacent to notochordal cavities containing notochordal vacuoles (NV). Long axes of spaces marked by daggers are 420 µm and 225 µm, respectively. ×27 and ×27. (e) A midsagittal section of ananterior caudal vertebra of N. coriiceps, sanded to show the limited extent of compact bone (asterisks) and its location around the notochordal cavities with the remainder of the centrum consisting of cancellous bone. This species has a small notochordal canal (NC) in the middle of each centrum. ×12. (f) Parasagittal histological section of an anterior caudal vertebra of N. coriiceps showing spongy bone and smooth-walled lipid-filled cavities. Arrows indicate accumulation of stain at growth checks in bone. The long axis of space marked by the dagger is 420 µm. ×83. From Eastman et al. (2011 [68], Figure 4), © Inter-Research, 2011, with permission.
![Diversity 16 00214 g012]()

- Neotenic retention of cartilage reduces bone mass in the adult skeleton. The delay in bone development and persistence of cartilage in the adult skeleton is a pedomorphic aspect of skeletal development (Voskoboinikova, 2001, 2010) [116,118] that reduces overall skeletal mass. This is characteristic of the neurocranium, branchial skeleton, and pectoral and pelvic girdles in many «cryonotothenioids». Much of the skull of specimens of Dissostichus mawsoni and channichthyids is cartilage overlain by a thin veneer of dermal bone. A radiograph of the endochondral pectoral girdle of D. mawsoni shows a range of skeletal tissues: a small amount of peripheral compact bone, cancellous bone, and a core of persisting cartilage (Figure 13). This is even more evident in channichthyids, a clade characterized by the late appearance and minimal development of bony elements. This group has more persistent cartilage in adults than in any other «cryonotothenioid» family (Voskoboinikova, 1997, 2010) [118,128]. Figure 11 bottom demonstrates the relatively small amount of bone in the neurocranium and branchiopharyngeal and pectoral regions. Figure 14a,b shows the large extent of persisting cartilage in the adult neurocranium and the thin superficial layer of dermal bone.Figure 13. Radiograph of a transverse section of the pectoral fin of Dissostichus mawsoni showing that the pectoral girdle is composed of a core of cartilage (C), surrounded by cancellous bone (B) of differing degrees of porosity (arrows), although the bone mineral content is typical of that for teleosts. Abbreviations: D, dermis; M, muscle; S, scales.Figure 13. Radiograph of a transverse section of the pectoral fin of Dissostichus mawsoni showing that the pectoral girdle is composed of a core of cartilage (C), surrounded by cancellous bone (B) of differing degrees of porosity (arrows), although the bone mineral content is typical of that for teleosts. Abbreviations: D, dermis; M, muscle; S, scales.
![Diversity 16 00214 g013]() Figure 14. (a) Dorsal view of the neurocranium of the skull of a fresh adult specimen of the channichthyid Chaenocephalus aceratus (29.4 cm TL). Most of the neurocranium, exemplified by the ethmoid region (E), consists of cartilage. The thin sheathing parasphenoid (PS) bone, of dermal origin, detached from the ventral surface of the neurocranium during maceration and is shown to the right. (b) A histological cross-section shows that the dorsolateral posterior neurocranium of C. aceratus consists primarily of cartilage (C), covered by a thin lamina of red staining bone (B, white arrows) beneath the dermis (D) of the skin.Figure 14. (a) Dorsal view of the neurocranium of the skull of a fresh adult specimen of the channichthyid Chaenocephalus aceratus (29.4 cm TL). Most of the neurocranium, exemplified by the ethmoid region (E), consists of cartilage. The thin sheathing parasphenoid (PS) bone, of dermal origin, detached from the ventral surface of the neurocranium during maceration and is shown to the right. (b) A histological cross-section shows that the dorsolateral posterior neurocranium of C. aceratus consists primarily of cartilage (C), covered by a thin lamina of red staining bone (B, white arrows) beneath the dermis (D) of the skin.
Figure 14. (a) Dorsal view of the neurocranium of the skull of a fresh adult specimen of the channichthyid Chaenocephalus aceratus (29.4 cm TL). Most of the neurocranium, exemplified by the ethmoid region (E), consists of cartilage. The thin sheathing parasphenoid (PS) bone, of dermal origin, detached from the ventral surface of the neurocranium during maceration and is shown to the right. (b) A histological cross-section shows that the dorsolateral posterior neurocranium of C. aceratus consists primarily of cartilage (C), covered by a thin lamina of red staining bone (B, white arrows) beneath the dermis (D) of the skin.Figure 14. (a) Dorsal view of the neurocranium of the skull of a fresh adult specimen of the channichthyid Chaenocephalus aceratus (29.4 cm TL). Most of the neurocranium, exemplified by the ethmoid region (E), consists of cartilage. The thin sheathing parasphenoid (PS) bone, of dermal origin, detached from the ventral surface of the neurocranium during maceration and is shown to the right. (b) A histological cross-section shows that the dorsolateral posterior neurocranium of C. aceratus consists primarily of cartilage (C), covered by a thin lamina of red staining bone (B, white arrows) beneath the dermis (D) of the skin.![Diversity 16 00214 g014]()


The genetic basis for the reduction of bone in «cryonotothenioids» is an alteration in the expression of certain genes during larval life, specifically delayed or absence of expression of type I and type X bone collagen genes, col1a1 and col10a1, and prolonged expression of the type II cartilage collagen gene col2a1 (Albertson et al., 2010) [120]. Changes in the expression patterns of these genes, possibly due to unidentified mutations in regulatory genes, result in a reduction in the amount of bone in the adult skeleton.
- Neotenic retention of notochordal tissue displaces and reduces some of the bone mass of vertebral centra. There is extensive interspecific variation in the size of the persisting notochord and vertebral canal in the centra of adult «cryonotothenioids» (Eastman et al., 2014) [42]. This results in the reduction in the mass of vertebral bone, a substantial component of overall skeletal mass. The centra of actinopterygians do not have a cartilaginous stage and develop by progressive mineralization of the notochordal sheath, with a subsequent contribution of bone from the somites (Arratia et al., 2001; Bensimon-Brito et al., 2012) [129,130]. During this process, the vertebral canal becomes progressively diminished in size or eliminated by bone during ontogeny. However, in many «cryonotothenioids», the canal remains partially patent, and the centra exhibit various degrees of “hollowness”, never becoming completely amphicelous (hourglass-shaped). This means that the biconid-shaped layer of compact bone is reduced and that this space is instead occupied by persisting notochordal vacuoles containing fluid that is only slightly denser than seawater (Table 1).
Based on a sample of 38 species from eight of nine families, non-Antarctic clades have relatively small vertebral canals, ≤11% of the diameter of the centra, whereas the canals are larger in the «cryonotothenioids» (Eastman et al., 2014) [42]. The species with the largest canals (65% and 48%, respectively) are the neutrally buoyant Pleuragramma antarcticum and Aethotaxis mitopteryx, and they are distinct among «cryonotothenioids» in the extent of this trait. Figure 15 shows the ontogenetic change in the appearance and relative size of the notochord and vertebrae of P. antarcticum. In the initial years of life (Figure 15a) the notochord is large in diameter and there is minimal vertebral bone—a gel-based body axis that is analogous to the hydrostatic skeleton of some soft-bodied invertebrates. As Pleuragramma increases in length (Figure 15b,c), the centra becomes more constricted as bone develops and the size of the vertebral canal is reduced but still evident (Figure 15c). The bone of the centra is cancellous with compact bone confined to a thin peripheral layer (Figure 16).
Relatively large vertebral canals (32–46%) are also seen in the bathydraconid genera Vomeridens, Racovitzia, Akarotaxis, and Bathydraco, and in the channichthyid genera Pagetopsis, Chaenodraco, Chaenocephalus, and Dacodraco. Modest-sized canals (25–30%) are present in species of the genus Trematomus (22–30%). Members of the demersal genus Notothenia and the primarily benthic families Harpagiferidae and Artedidraconidae have smaller canals (≤20%, and frequently <10% in larger specimens). Interestingly, the demersal Eleginops maclovinus (%B = 3.64) and adults of Dissostichus mawsoni (%B = 0) and possibly D. eleginoides have canals as small as 4% of the diameter of the centra. These three species undertake migrations as part of their life cycles and have well-developed white axial musculature, constituting about 60% of the body mass in D. mawsoni. Although composed predominantly of cancellous bone, completely ossified centra in these species are likely needed to support the vertebral axis during the stress produced by bouts of subcarangiform locomotion powered by the axial musculature.
Accumulated lipids are essential for neutral buoyancy. Only ≈5% of «cryonotothenioid» species are neutrally buoyant at some period in their adult lives, with %B of values 0–0.5. These are Pleuragramma antarcticum, Aethotaxis mitopteryx, Dissostichus mawsoni, and possibly D. eleginoides, and Gvozdarus svetovidovi. The locations of the triacylglycerol lipid stores that lower the overall density of neutrally buoyant species have been extensively documented (DeVries and Eastman, 1978; Eastman and DeVries, 1981, 1982, 1989; Clarke et al., 1984; Eastman, 1988a, 1993, 1997; Fenaughty et al., 2008; La Mesa and Eastman, 2012) [63,64,122,123,124,131,132,133,134,135]. As far as the density axis is concerned, the key point is that neutral buoyancy cannot be achieved solely by skeletal reduction; lipids are also necessary to reduce overall body density, and this is contingent on the availability of lipids in the diet. For example, in D. mawsoni living in the southwestern Ross Sea total body lipid content is about 10% of body mass in neutrally buoyant individuals. The lipid accumulates in typical white adipose cells in and around the axial musculature and beneath the skin. This lipid is provided by a diet consisting of P. antarcticum at levels of 71% by occurrence and 89% by weight (Eastman, 1985a,b) [62]. The lipid content of P. antarcticum is ≈40–50% of dry mass at a size of 16 cm (Bock et al., 2017; Hagen & Kattner 2017) [136,137] and continues to increase with length.
Stored lipids in Dissostichus mawsoni accumulate with growth in size (Near et al., 2002) [65], and may be labile in adults. The lipid stored in the neutrally buoyant species must also be maintained during the course of their life history—significant biological consequences ensue if the lipid is accessible for other purposes. In most fishes, including D. mawsoni, a lipid is stored in typical adipocytes and is therefore not sequestered from the reproductive, nutritional, and general metabolic needs of the fish (Pelster 2009, p. 81) [104]. Evidence of the non-protected status of the buoyancy lipid in D. mawsoni came to light in specimens captured by the commercial longline fishery in the northern Ross Sea. Some of the catch consisted of emaciated, low-condition “axe handle” individuals, likely in a post-spawning state after having exhausted their lipid stores for migration, gametogenesis, and nutritional needs in waters of the spawning grounds devoid of Pleuragramma (Fenaughty et al., 2008) [134]. On the slope, non-fatty fish like macrourids (rattails) of the genus Macrourus and the channichthyid Chionobathyscus dewitti are the primary prey for D. mawsoni (Fenaughty et al., 2003; Petrov and Tatarnikov, 2011; Roberts et al., 2011) [138,139,140]. With this in mind, the lipid-based neutral buoyancy in the population of D. mawsoni in the southwestern Ross Sea could be viewed as a contingency in the sense that its long-term permanence in any individual is dependent on geographic location, adequate diet, and amount of energy expended on migration and reproduction. A diet dominated by lipid-rich Pleuragramma has conferred neutral buoyancy on fish in this locality—the only population documented to date as having neutrally buoyant adults (Eastman, 2020) [45].
Impermanent lipid-based neutral buoyancy is neither unique to notothenioids nor is it maladaptive (Eastman, 2020) [45]. Given the disruptive climatic and trophic conditions on the Antarctic shelf during the last 10 million years, it may have been advantageous for the largest predatory fish in the ecosystem to have a wide niche breadth and the ability to occupy biotopes in the pelagic and benthic realms. Figure 17a,b shows a ≈65 cm TL D. mawsoni that was captured on video living under a rock ledge at 400 m in Gerlache Strait (≈64° S) off the Antarctic Peninsula. The fish emerged from under the ledge in response to the disturbance caused by the propulsion system and lights of the ROV and after a few seconds returned under the ledge. While the fish was not the size of a mature adult and not neutrally buoyant, it was obviously in good condition. In the absence of Pleuragramma antarcticum, D. mawsoni in this area may not become neutrally buoyant as adults.
Stored lipids in Pleuragramma are more stable. Although Pleuragramma antarcticum does have some typical adipose tissue, most lipids are contained in grossly visible 1–2 mm-diameter “sacs” (Figure 16), rather than 100–200 µm adipocytes (Eastman, 1997) [133]. The composition of the sac wall was unknown at this time as it could not be resolved with light microscopy, and the sac was thus viewed as a means of sequestering the lipid for buoyancy. However, electron microscopy subsequently revealed that the sac wall is composed of an array of adipocytes arranged circumferentially around a single large lipid droplet and that there are capillaries external to the adipocytes (Eastman and DeVries, 1989) [124]. This is expected because a cellular mechanism is needed to effect the transport of lipids from blood into the sac lumen—only intracellular lipids can be made available for metabolism (Pelster, 1998, p. 28) [127]. A sphere has a minimal surface area for its volume, and the advantage of a large “sac” compared to single small adipocytes is that there is relatively less cell membrane surface for transport to occur, and the lipid store is probably more stable and less subject to metabolic demand. However, the store is not fully uncoupled from metabolism (Maes et al., 2006) [141] and is available under unusual conditions (Eastman, 2020, pp. 1226–1227) [45].
2.5. Morphology Stage—Axis 5: Body Shape
There are nine body shapes among fishes (Barton, 2007, pp. 35–39) [142] with the spindle-shaped or fusiform body being the most common. Body shape provides insight into aspects of life history including habitat, locomotion, and feeding mode. In an analysis of more than 3300 species of marine teleosts, Friedman et al. (2020) [143] found that there has been divergence along the benthic to pelagic axis and that within lineages most disparity in body shape is attributable to either widening or elongation. Furthermore, the fastest rate of widening and elongation in a sample of more than 6100 teleosts is seen in a clade formed by notothenioids and scorpaeniforms (Price et al., 2019) [144]. While there is a disparity in body shape among «cryonotothenioids», the subzero water temperatures of the Southern Ocean shelf have imposed metabolic constraints on activity levels. There are not any persistently active «cryonotothenioids»—most species are sedentary, and either demersal (73%) or benthic (12%) (Eastman 2020) [45]. This lack of activity may occupy 97% of a 24-h period (North, 1996; Zimmermann and Hubold, 1998) [70,100] and restrict the divergence of body shapes associated with maneuvering and sustained activity. The physical properties of cold seawater may limit the divergence of body shapes associated with swimming because the dynamic viscosity of seawater is nearly 2-fold greater at −2 °C than at 20 °C (Macdonald and Wells, 1991) [145]. Therefore «cryonotothenioids» exhibit little continuous swimming or maneuvering and only two species, (Pleuragramma antarcticum and Aethotaxis mitopteryx) have a life history free of the substrate.
There are some morphometric data for the body shape axis in «cryonotothenioids» (Klingenberg and Ekau, 1996; Colombo et al., 2015) [31,67]. As indicated above, most disparity in body shape does involve differing degrees of widening or elongation of the fusiform phenotype (Figure 9a,b,f,i). The mouth is usually terminal and the lateral profile of the head is frequently angular, but may be blunt in species that live or forage on the substrate (Figure 9e,i,k). The head is dorsoventrally flattened in species of Pogonophryne (Figure 9g). Some channichthyids have notably long (35–40% SL) and wide heads (Figure 9c). Elongation of the body (Neat and Campbell, 2013) [146] is characteristic of some species of Gymnodraconinae, especially Parachaenichthys spp. (Figure 9d), and also of the Bathydraconinae (Figure 9h). Lateral compression is evident in the pelagic nototheniids Pleuragramma antarcticum and Aethotaxis mitopteryx. There are no deep-bodied «cryonotothenioids» as this latter body shape is usually associated with fishes that require stability and maneuverability when swimming. However, ecomorphological measurements and statistical analyses do reveal divergence of body and fin shapes and proportions associated with locomotion for foraging in the water column, even among closely related species that are demersal based on measurements of their %B. For example, Trematomus lepidorhinus (Figure 9f) is more streamlined and presumably more active than its sister species T. loennbergii (Klingenberg and Ekau, 1996) [67], and this is also reflected in their neuromorphology (Eastman and La Mesa, 2021) [60].
Fin positions are relatively stable among «cryonotothenioids» and the only loss is of the first dorsal fin in bathydraconids (Figure 9d,h). The pectoral fins are large, fan-shaped, and the primary means of locomotion in most species. They are especially long, 27–30% SL, in Vomeridens infuscipinnis), a species that hover above the substrate. The pelvic fins of many demersal and benthic species have thickened fin rays and skin where they are subject to contact with the substrate. Channichthyids with a bipod or tripod stance (Pagetodes antarcticus and P. atkinsoni) have long strut-like pelvic fins. The semipelagic channichthyids Neopagetopsis ionah and Dacodraco hunteri have pelvic fins that are long and wide, with thin rays and skin. When extended into a fixed position, they may be used as hydrodynamic lifting surfaces to ride and maintain position in currents.
2.6. Morphology Stage—Axis 6: Trophic Morphology, Specifically Oral and Pharyngeal Morphology (Jaws, Teeth, Head Size, and Oral and Pharyngeal Gapes)
There is extensive information on the diets of «cryonotothenioids», but minimal data on trophic morphology. Below is information on jaw and tooth morphology and aspects of the divergence in head morphology with examples drawn from channichthyids and bathydraconids. This is prefaced by remarks on the food web, diets, and the ecomorphology of feeding.
Food may not be a limiting resource for many «cryonotothenioids». Antarctic benthic communities have high levels of biomass—at depths <1000 m it is higher than in non-Antarctic regions (Brey and Gerdes, 1997) [147]. The Antarctic benthos is also among the most long-lived and ecologically stable in the world (Clarke and Johnston, 2003) [148]. Although there is marked seasonality in primary production in the water column, the benthic fauna on the Antarctic shelf may not be food-limited even in winter because detrital sediments serve as a food bank and contain labile organic matter all year (Mincks et al., 2005; Smith et al., 2006) [149,150]. «Cryonotothenioids» do not stop feeding during winter (Targett et al., 1987; Gröhsler 1994) [81,151], and some benthic species like Harpagifer antarcticus show seasonal stability in the elements (C, H, N, C:N ratio) composing their tissues (Obermüller et al., 2013) [152].
«Cryonotothenioid» diets include a diverse array of mobile and sessile organisms including krill, amphipods, copepods, isopods, polychaetes, salps, echinoderms, bivalves, gastropods, chitons, fish and macroalgae (Targett, 1981; Daniels, 1982; Casaux et al., 1990, 2003; McKenna, 1991; Kiest, 1993; Montgomery et al., 1993; Vacchi et al., 1994, 2000; Barrera-Oro, 2003; La Mesa et al., 2004a,b) [72,75,153,154,155,156,157,158,159,160,161,162]. Many species are dietary generalists and feed opportunistically on seasonally and locally abundant items, although certain prey items are especially important. For example, given their speciosity, great abundance, and high energy density (Brey, 2001) [163], amphipods are probably the most important invertebrate benthic organisms consumed by «cryonotothenioids», and a primary food source in many areas throughout the year (Barrera-Oro, 2002, p. 298) [164].
Krill and calanoid copepods have similar status as important pelagic prey for «cryonotothenioids». Many demersal and benthic species also depend on water-column resources, especially krill (Permitin, 1970; Barrera-Oro and Casaux, 1990; Foster and Montgomery, 1993; Pakhomov, 1997; Barrera-Oro, 2002; Stowasser et al., 2012; Casaux and Barrera-Oro, 2013) [69,76,164,165,166,167,168]. In high-latitude shelf waters, the dominant krill species is Euphausia crystallorophias, but this is replaced by E. superba in off-shelf and lower-latitude waters. Most «cryonotothenioid» species will opportunistically consume larval and adult krill as seen in Figure 17c. This screenshot from an ROV video sequence documents a demersal species, Trematomus scotti, capturing a single krill by means of a quick upward lunge from the substrate. The presence of large numbers of krill close to the substrate in a particular location and at a particular time can be a confounding factor in analyzing diets by masking selective feeding. However, if krill are absent from an area, the diets of sympatric species do show more selectivity that reflects known differences in their morphology and buoyancy. For example, the demersal sister species Notothenia coriiceps and N. rossii are sympatric in Potter Cove, King George Island, are morphologically distinct, and have different activity levels and percentage buoyancies (Eastman et al., 2011) [68]. In some summers when krill are absent or sparse in Potter Cove, the diets of both species when summarised at a high taxonomic level, are similar and consist mainly of benthic amphipods and macroalgae. However, when prey is identified and analyzed at the level of the species, the less buoyant N. coriiceps is more herbivorous and feeds more intensively than N. rossii on benthos such as algae and other (non-amphipod) groups like gastropods and bivalves. Notothenia rossii, a more buoyant and active species that spends more time in the water column, feeds on a greater proportion of epibenthic amphipods and other prey (Barrera-Oro et al., 2019) [71].
On high-latitude shelves, shoals of juvenile, small, and adult Pleuragramma antarcticum may be the piscine equivalent of a krill swarm. Although their presence in a given area is a random occurrence, they will likely be preyed on by most other «cryonotothenioids» in the vicinity, irrespective of their biotope, as well as being subject to cannibalism within the shoal (Eastman, 1985b) [169]. During the Nathaniel B. Palmer Cruise 97-9, trawling through a sponge community near Beaufort Island captured 68 specimens of the demersal Trematomus eulepidotus (%B = 3.41) that had been feeding in the water column on small P. antarcticum. Dissostichus mawsoni also feeds on Pleuragramma, not because of specialization to do so but rather because of the abundance of the latter in some areas. The high lipid content of Pleuragramma (Friedrich & Hagen 1994; VNIRO 2000) [170,171] is also essential for both the attainment and persistence of neutral buoyancy in D. mawsoni.
The ecomorphology of feeding. There are three basic feeding modes in fishes: suction, ram, and manipulation or biting (Liem, 1980) [172]. While inertial suction feeding is most common (Lauder, 1982; Liem, 1993; Westneat, 2006) [173,174,175], many teleosts use a combination of modes depending on the nature of the prey, usually involving ram and suction feeding (Norton and Brainerd, 1993) [176]. The morphology associated with each of these modes is as follows (Liem, 1993) [174]. Suction feeders have deep heads, a relatively small oral gape, and a cone-shaped branchiopharyngeal cavity that widens posteriorly. Ram feeders have a large oral gape, a cylindrical branchiopharyngeal cavity, and longer jaws. Biters have variable head shapes, usually a small oral gape and robust jaws. Biomechanical measurements of the jaws and calculation of the mechanical advantage ratios and suction indices are available for only three «cryonotothenioids»: Trematomus bernacchii (Bansode et al., 2014; Carlig et al., 2018) [177,178], Dissostichus mawsoni and Pleuragramma antarcticum (Carlig et al., 2018) [178]. Hopefully, more data on feeding ecomorphology will be added to the trophic morphology axis in the future. The following are some general observations on morphological divergence and feeding.
The morphology of jaw bones and teeth. The phylogenetically basal Percophis, Pseudaphritis, and Bovichtus have jaw morphology and mobility typical of many percoids (Iwami, 1985; Odani et al., 2006) [107,179]. The premaxilla of Eleginops maclovinus (Figure 18a), the sister group of the «cryonotothenioids», also has a long ascending process and a well-developed articular process. The postmaxillary process is small. The long ascending process implies that there is a considerable protrusion of the premaxillae during biting and browsing. The dentary has a large coronoid process and considerable surface area on the ventral limb for the insertion of the A2 subdivision of the adductor mandibulae. This muscle is responsible for the jaw closure associated with seizing, holding, and biting prey. The jaws of Eleginops are substantial bones that are likely subject to stress during browsing. Eleginops have a subterminal mouth and are opportunistic omnivores (Pequeño, 1989; Licandeo et al., 2006) [180,181] capable of feeding on infaunal and epifaunal invertebrates. It also browses among bryozoans that provide refuge for crustacean prey (Brickle et al., 2005a,b; Pavés et al., 2005) [182,183,184].
The jaws of «cryonotothenioids» depart from the morphology in Eleginops in that the bone mass is reduced although features associated with benthic browsing involving suction and manipulation are retained. For example, the premaxillae of Notothenia coriiceps (Figure 18b) and Trematomus bernacchii (Figure 18c) have long vertical ascending processes. Dissostichus eleginoides (Figure 18e) has a shorter more oblique ascending process, and this species may have less capability for protrusion, but more jaw stability for ram strike feeding on primary fishes (Troccoli et al., 2020) [185]. Both species of Dissostichus are capable of ram-strike and ram-suction feeding (Collins et al., 2010 [186]; Eastman, personal observation).
The head length and jaw morphology of bathydraconids are interspecifically variable. Psilodraco, Prionodraco, and Racovitzia jaws have well-developed processes on the premaxillae, morphology associated with browsing. However, the jaws of Gerlachea and especially the species of Parachaenichthys (Figure 19a) are more similar to those of channichthyids as they are elongated with reduced surface area, mass, and rudimentary processes. Unlike channichthyids, however, the postmaxillary processes of the premaxillae are well developed in predatory bathydraconids such as Parachaenichthys (Figure 19a) that grasp and maintain prey in the jaws rather than relying on a large oral gape for swallowing prey. This dorsally projecting crest may prevent lateral displacement of the premaxillae relative to the maxilla (Gregory, 1933, p. 241) [187].
Channichthyids have widened and elongated heads and correspondingly larger jaws and oral gapes than bathydraconids. The jaw bones are reduced in mass and gracile, the limbs of the dentary are short, especially the ventral limb, and the ascending and articular processes of the premaxilla are reduced to small knobs (Figure 19b,c), with the loss of premaxillary protrusion in most species (Iwami, 1985; Voskoboinikova, 1993) [179,188]. Pseudochaenichthys georgianus has a rudimentary ascending process (Figure 19b) but it has been lost in Chaenocephalus aceratus (Figure 19c). Elongated and fixed jaws are characteristic of ram-strike feeders because the stability they provide is advantageous in capturing large or struggling prey (Liem, 1993) [174].
Simple conical teeth are the norm among bony fishes (Peyer, 1968, p. 84) [189], including «cryonotothenioids», and the primary function of the jaw teeth is seizing prey. The jaw teeth in the phylogenetically basal Percophis brasiliensis are conical with sharp tips; those situated anteriorly are larger and fang-like, but single large teeth are also situated among smaller teeth at intervals along the length of the jaws (Eastman, personal observation). All are firmly ankylosed to the premaxillae and dentaries, the “Type 1” mode of attachment (Fink, 1981) [190]. Eleginops maclovinus, the sister species of the «cryonotothenioids» has numerous small conical teeth and the mode of attachment is uncertain. In a sample of water-macerated skeletons from five specimens, ranging in total length from 26.1 cm to 74.5 cm, some or all of the teeth remained ankylosed to the jaws in three specimens, but most became detached in two others including the largest (Figure 18a). With the exception of enlarged fang-like symphyseal teeth, tooth-crowns are usually not ankylosed to the pedicles on the jaws but, instead, are attached by a collagenous ligament. This “Type 2” mode of tooth attachment is considered a pedomorphic trait (Fink 1981) [190] and, with the exceptions noted below, the teeth of most «cryonotothenioids» are slightly moveable in life. There is also a phylogenetic reduction in the number of rows of teeth among «cryonotothenioids» (Voskoboinikova 1993) [188].
While the dentition of «cryonotothenioids» appears to be homodont, it is functionally heterodont meaning that there are regional specializations in the teeth (Cohen et al., 2020) [191]. The conical teeth differ in size, sharpness, curvature, and position along the jaws (Figure 18 and Figure 19). They are suited for biting, grasping, and manipulating prey but not for shredding, grinding, or substantial crushing. Some in Trematomus bernacchii (Figure 18c) are unusual in showing evidence of wear and are used in breaking the thin shells of a scallop (Vacchi et al., 2000) [159]. The teeth of Notothenia rossii (Figure 18d) are sharp and vary in size along the dentary. The large symphyseal teeth on the premaxillae and dentaries of D. eleginoides (Figure 18e) are sharp, slightly recurved (bent posteriorly), and are firmly ankylosed to the jaws. They are suited for impaling and holding prey during the strike and subsequent engulfment. The bathydraconid Gymnodraco acuticeps is a sedentary, lurking predator with a pointed snout and a projecting lower jaw. It has several large, sharp, posteriorly projecting dagger-like symphyseal teeth; those on the dentaries are prominent and externally visible. The bodies of the premaxillae and dentaries also have a single row of smaller dagger-like teeth (Andriashev et al., 1989; Voskoboinikova, 1991; Balushkin and Voskoboinikova, 2011) [192,193,194]. The teeth of the piscivorous bathydraconid Parachaenichthys charcoti (Figure 19a) are recurved and relatively homogeneous in size. The teeth on the upper jaws of the channichthyid Pseudochaenichthys georgianus (Figure 19b) have a lingually-directed curvature; the anterior teeth of the lower jaw are recurved. This species eats fish and krill. The channichthyid Chaenocephalus aceratus is capable of seizing and swallowing large piscine prey. The teeth on the upper jaw have a medial curvature (Figure 19c) whereas most of those on the lower jaw are straight. They probably retain the prey while it is being swallowed, a process that can take 24 h in cases where the prey fish is large.
The alimentary canal morphology of «cryonotothenioids» exhibits minor divergence and little relationship to diet. The heads of some «cryonotothenioids» display specialized morphology for the acquisition of prey, but the alimentary canal from the esophagus to the rectum is generally similar in its gross morphology and histology in the species studied to date. The interspecific differences include: (1) a phylogenetic decrease in mean number of pyloric caeca from 6–7 in most nototheniids to 2 in channichthyids, although Dacodraco has 4; (2) a maximum ≈1.5-fold difference in average relative stomach lengths; and (3) a maximum ≈2-fold difference in average relative intestine lengths (Ojeda, 1986; Eastman and DeVries, 1997; Eastman 1999; Moreira et al., 2020) [195,196,197,198]. The absence of specialization may be attributable to the opportunistic feeding and dietary plasticity of many species. For example, depending on the time of year and location, the alimentary canal of Notothenia coriiceps processes food on a scale ranging from carnivory to omnivory to herbivory (Barrera-Oro and Casaux, 1990; Casaux et al., 1990) [69,153], although perhaps less efficiently that if it was more specialized for only carnivory or herbivory. Hence there is no morphology that reflects intermittent herbivory on kelp, and attempts to correlate diet with stomach length, intestine length, or number of pyloric caeca are inconclusive (Moreira et al., 2020) [198]. There may also be non-dietary influences on gut morphology. During benthic browsing, for example, the ingested inorganic detritus occupies space in the intestinal lumen and may reduce the efficiency of digestion and absorption. This could necessitate a longer intestine, with Gobionotothen gibberifrons the exemplar among «cryonotothenioids» (Moreira et al., 2020) [198]. The absence of specialization in the alimentary canal is neither a new observation nor unique to the «cryonotothenioids»—“the ecological situation of fish may often favor a generalized rather than a highly specialized digestive system” (Barrington, 1957, p. 112) [199].
Head-related trophic morphology is divergent in «cryonotothenioids». Species with elongated heads and jaws occur in numerous, unrelated, predatory percoid clades including some channichthyids and, to a lesser extent, the bathydraconids Parachaenichthys spp., Cygnodraco, and Gymnodraco. In addition to head length, head width and size of oral gape are also increased. This allows larger prey such as other fish to be struck and held in the mouth before being manipulated and then supported while being swallowed head-first. There is also a disparity in the shape of the snout, whether U-shaped (rounded) or V-shaped (pointed), that also reflects the size of the prey consumed. The snout is U-shaped (Figure 20a, Figure 21b and Figure 22a) in species that engulf large prey fishes head-first, and V-shaped (Figure 20b and Figure 23a,b) in species that dwell in the water column and consume the narrow-bodied Pleuragramma antarcticum and krill by striking them laterally. In channichthyids, the phylogenetic trend toward reduction in head ossification decreases the rigidity of the branchial and pharyngeal regions (Eastman et al., 2014) [42], and this allows the swallowing of large piscine prey (Iwami, 1985) [179]. Below are examples of disparate head and trophic morphology in channichthyids and bathydraconids.
- Channichthyids. The channichthyids are distinctive among «cryonotothenioids» in having the largest mean total length, the longest snouts, the largest oral and pharyngeal gapes, and fixed premaxillae. The functional implications of this morphology are that, along with Dissostichus spp., they are the only «cryonotothenioids» capable of ram-strike feeding on relatively large prey in the water column or on the substrate. There are no channichthyids that are obligate benthivores (Voronina & Neelov 2001) [201]. Channichthyids exemplify the divergence of head morphology away from that suited for benthic browsing, involving biting and suction of relatively small prey to consuming large prey such as fish. This transition has involved both the lengthening of the neurocranial base, especially in the ethmoid region (Iwami, 1985; Voskoboinikova, 1997, 2010) [118,128,179] and the widening of the head. The heads are distinctively large, one-third or more SL, with the snout length accounting for ≈50% of head length (HL) in species with the largest heads (Iwami and Kock, 1990) [202].
The retention of large amounts of cartilage and the small amounts of bone are more pronounced in channichthyids than in other «cryonotothenioids» (Voskoboinikova, 1997, 2001) [116,128]. This is relevant to trophic morphology because the pharyngeal gape determines what can actually be swallowed (Wainwright and Richard, 1995) [85], especially when large and rigid prey is consumed. The pharyngeal gape (Figure 21b) is the diameter of the pharyngeal-oesophageal junction when expanded, and this is smaller than the oral gape (Figure 21a). The distance between the cleithra of the pectoral girdles is the limiting dimension (Wainwright and Richard, 1995) [85] or “choke point” where boniness and rigidity become obstacles to swallowing large prey. In channichthyids, however, the cleithra are relatively thin and cartilaginous, and the critical intercleithral space is wide with little bone in the vicinity (Iwami, 1985) [179]. In addition, the fifth ceratobranchials (pharyngeal bones) are thin, and planar, bear only small teeth, and do not impinge on the pharyngeal lumen (Figure 21b). Collectively this morphology, also indicated by the absence of bone in the CT scan in Figure 11 bottom, allows greater than expected lateral and dorsoventral expansion of the pharyngeal gape during swallowing of large prey (Figure 21a and Figure 22a). The following three examples highlight the divergence in trophic morphology in species occupying different biotopes.
- Large-headed (≈40% SL) channichthyids. There are three large-headed species of medium-large body size: Chaenocephalus aceratus (TL = 76 cm, HL = 33–42% SL; %B = 3.46), Pseudochaenichthys georgianus (TL = 60 cm, HL = 36–43% SL; %B = 1.96) and Channichthys rhinoceratus (TL = 58 cm, HL = 37–41% SL; %B unknown). These three species are ram-strike predators on fish, and also krill in the case of P. georgianus (Duhamel et al., 2005; Kock 2005) [203,204]. Pseudochaenichthys georgianus differs from the other two species in being semipelagic (%B = 1.96) and it likely feeds in the water column as well as on the substrate. Chaenocephalus aceratus (Figure 21a) has a diet consisting almost exclusively of fish (Figure 22a). They are capable of engulfing large-headed fish including other channichthyids that are 40–50% of their own length (Kock 2005; Reid et al., 2007; Kock et al., 2013) [204,205,206]. Remarkably, a Chaenocephalus aceratus swallowed a Dissostichus mawsoni that was 76% of its length and 41% of its weight (Kock et al., 2013) [206]. In swallowing large prey fishes head-first, the jaws of channichthyids remain partially open for periods of hours as the head of the prey distends and then slowly passes through the pharyngeal gape and into the stomach causing the ventral body wall to become and large pendulous. The caudal trunk of the prey may sometimes protrude from the mouth for 1–2 days in this type of headfirst ram-strike feeding, and complete digestion may take at least five days (Kock et al., 2013 [206]; Eastman, personal observation on aquarium specimens).
- Substrate-perching channichthyid with an elevated bipodal stance. The semipelagic Pagetodes antarcticus (Figure 22b) (TL = 65 cm, HL = 36% SL; %B = 2.53) has pelvic fins that are 23–70% SL, with a mean of 42% (La Mesa et al., 2002) [207]. There is an ontogenetic decrease in the relative size of these fins (La Mesa and Vacchi, 1997) [208]. Perching has been previously reported in this species, but as a “tripod” stance involving the caudal fin with the head down and body angled toward the substrate (Ekau & Gutt 1991, p. 45) [209]. However, based on videos from an ROV, P. antarcticus also balances itself using only the pelvic fins (Figure 22b) that are, in this instance, ≈45% SL. It is not known if the perching behavior encompasses all sizes or is confined primarily to smaller specimens with relatively longer pelvis’. Perching provides elevation and an enhanced view of the substrate and, with no locomotion involved, is energetically efficient. Given the relatively large size of the pelvis’ throughout the range of body lengths, anything other than a bipodal stance may be impossible when the pelvis’ are fully extended toward the substrate. Pagetodes antarcticus preys primarily on fish (Kock, 2005; Kock et al., 2013) [204,206]. In the southern Scotia Arc, this species is able to consume specimens of Chionodraco rastrospinosus that are 60–66% of its own length and 20–26% of its own weight (Kock & Jones 2002) [210]. They also consume krill (Iwami & Kock 1990) [202].
- Semipelagic channichthyids that feed primarily in the water column. This was first recognized by Nybelin (1947, pp. 55, 58) [52]. Chen et al. (1998) [211] confirmed the existence of ecological disparity at some terminal nodes in their cladogram for channichthyids, with sister species occupying demersal and semipelagic biotopes. Below are examples of two species that possess disparate morphology associated with feeding in and habitation of the water column, and a third species that spends time in the water column and on the substrate.
Dacodraco hunteri (TL = 29 cm, HL = 36–39% SL; %B = 1.41) is the smallest channichthyid, a slim-bodied, streamlined water-column predator, with pelagic coloring—dark dorsally and light ventrally and with a silvery iris. Dacodraco has a relatively narrow head and a V-shaped snout and exhibits little of the opercular flare characteristic of most other channichthyids. Its total head length is within the typical range for most channichthyids, but its snout length (49–55% HL) is among the longest. Dacodraco is semipelagic but does not have a significant amount of lipid for buoyancy. Its large pelvic and pectoral fins, when abducted, probably produce drag and slow its rate of sinking when it is in the water column. They also may function as hydrodynamic lifting surfaces that allow orientation into and riding of currents, an energy-efficient alternative to swimming. The pelvic fins show no evidence of substrate contact adaptations such as thickened rays and skin. Dacodraco has the most distinctively bowed jaws among «cryonotothenioids» (Figure 23a). Large, sharp, conical teeth are spaced regularly along the lengths of the jaws, with a few smaller teeth in between. Dacodraco feeds in the water column on Pleuragramma (Eastman 1999) [197], probably by seizing them in a lateral strike. The bow of the jaws accommodates the laterally held prey and the large teeth maintain the prey firmly in the jaws while is it being repositioned to be swallowed head-first. A wide gape is not necessary as Pleuragramma are narrow-bodied. Krill are another component of the diet (Kock 2005) [204].
Neopagetopsis ionah (TL = 57.5 cm, HL = 32–39% SL; %B = 1.28) is also streamlined with a narrow head, V-shaped snout, slightly bowed jaws and the coloring of a water-column species (Figure 20b and Figure 23b). The holotype was taken from the stomach of a krill-feeding whale (Nybelin, 1947) [52]. Recent collections in the Ross Sea suggest that N. ionah also occurs in the upper reaches of the water column (Hanchet et al., 2013, p. 634) [22]. Its body is fusiform with a more robust trunk musculature than Dacodraco. The pelvic fins are enlarged and fan-shaped (Permitin, 1970, p. 180) [76], with relatively thin rays and thin skin on the distal margins. Similar to Dacodraco, the pelvic fins may function, along with the pectorals, as hydrodynamic lifting surfaces for orienting into currents. Neopagetopsis feeds on krill (Kock 2005) [204] and to a lesser extent on Pleuragramma (Permitin 1970; Abe and Suzuki, 1978, 1981; O’Driscoll et al., 2011) [76,212,213,214]. Neopagetopsis is also a relatively active species. During the Nathaniel B. Palmer Cruise 97-9 in the Ross Sea, a large specimen of N. ionah was maintained in an aquarium for several days. It exhibited considerably more spontaneous swimming activity than specimens of Chionodraco hamatus and Pagetodes antarcticus in the same tank.
Chionobathyscus dewitti (TL = 60 cm, HL = 33–37% SL; %B = 1.22). This species is distinguished from other channichthyids by its wide, blunt, squarish snout, evident in a dorsal view (Figure 23c). It is the most buoyant (%B = 1.22) and, at 2025 m, the deepest living channichthyid. Photographic surveys (Eastman et al., 2013) [200], the bycatch in the commercial longline fishery for Dissostichus mawsoni (Balushkin and Prut’ko, 2006; Sutton et al., 2008) [215,216], and the stomach contents of D. mawsoni from the fishery (Petrov, 2011) [217] indicate that C. dewitti is the most common channichthyid at depths of 600–2000 m. Another photographic survey of the outer shelf/upper slope of the western Antarctic Peninsula found that the maximum abundance of C. dewitti was 900–1499 m (Amsler et al., 2016) [61]. C. dewitti has been documented hovering in the water column, above the substrate at 1360 m (Figure 23c), with the pectoral fins extended away from the body (Eastman et al., 2013) [200]. This position is probably maintained by propagating an undulating wave through the pectoral fin rays. It is possible that the wide snout acts as a horizontal stabilizer. This elevated position above the substrate may provide an enhanced view and ambush position for striking prey. Although it has the lowest %B value of all sampled channichthyids, it also spends time on the substrate when guarding eggs (La Mesa et al., 2021) [46]. Its pelvic fins are covered by thick skin indicating contact with the substrate. Given its greater habitat depth, its diet differs from other channichthyids in that it includes non-«cryonotothenioids» like paralepidids (barracudinas), especially Notolepis, myctophids (lanternfishes), macrourids (rattails) of the genus Macrourus and squid (Sutton et al., 2008; Petrov 2011) [216,217].
- 2.
- Bathydraconids also exhibit divergent head morphology. Bathydraconids are medium-sized and narrow-bodied. The snout ranges from relatively long and pointed to short and blunt. Longer-headed species include the two species of Parachaenichthys (HL = 41% SL), Cygnodraco mawsoni (HL = 41% SL) and Gymnodraco acuticeps (HL = 37% SL) (Figure 24). Figure 24 also shows that the eyes in some species are dorsolateral and “look up”, possibly attributable to the narrowing of the head, although in some species this might also indicate the primary feeding vector. There is also considerable interspecific variation in the premaxillae and dentaries (Figure 25). As feeders on benthos (Gutt and Ekau, 1996) [218], some of the shorter-headed species like Psilodraco, Prionodraco, and Racovitzia have relatively heavy protrusible jaws with well-developed ascending and articular processes on the premaxillae (Andriashev et al., 1989) [192]. These shorter and more rugged jaws are suited for biting and manipulation of prey during feeding. The ascending and articular processes are reduced or vestigial in Parachaenichthys, Cygnodraco, and Gerlachea (Andriashev et al., 1989; Balushkin and Voskoboinikova, 2011) [192,194]. The elongated snouts and less rugged jaws suggest that these species are ram-strike feeders. The oral and pharyngeal gapes, however, do not approach the size of those in channichthyids. As evident in Akarotaxis and Prionodraco (Figure 25b), the body of the dentary is also larger to accommodate the pores, neuromasts, and branches of the cephalic lateral-line nerve. The following examples highlight some of the disparity in trophic and general morphology.
- Cygnodraco mawsoni and Parachaenichthys spp. Have long narrow heads and snouts, well-muscled trunks (Figure 9d and Figure 24), and are more active predators than most other bathydraconids. Parachaenichthys charcoti is benthic (%B = 5.05); the buoyancy of Cygnodraco is unknown. These species may be short-pursuit predators on and near the substrate. In the case of C. mawsoni, a 49 cm TL specimen held in an aquarium exhibited considerable spontaneous swimming employing both subcarangiform and labriform locomotion. The jaws of Parachaenichthys charcoti (Figure 19a) resemble those of channichthyids (Figure 19b,c) except that they are heavier and the inferior limb of the dentary is larger. Parachaenichthys feeds on fishes and mysids (Targett 1981; Gon, 1990) [72,219], and the diet of Cygnodraco includes fishes, decapods, mysids, amphipods, and polychaetes (Gon, 1990) [219]. Based on their head and jaw morphology, both species are probably ram or ram-suction feeders.
- Vomeridens infuscipinnis (Figure 26) hovers above the substrate using its pectoral fins that are the longest among bathydraconids at 27–30% SL. Those of Parachaenichthys and Cygnodraco are only 16–17% SL. Vomeridens and the five species of Bathydraco have delicate spindle-like trunks. Vomeridens is semipelagic with a %B of 1.61. The stomachs of individuals 168–216 mm SL contained adult krill 46–50 mm in length (Kuhn et al., 2011) [220]. During the Nathaniel B. Palmer Cruise 97-9 in the Ross Sea a towed video camera captured a sequence at 1191 m showing a bathydraconid headed into the current, with body axis held straight, simultaneously using its large fan-shaped pectoral fins, and possibly the smaller pelvic fins, to hover a short distance off the substrate (JM Barry and JT Eastman, personal observation). Although it was not possible to identify the species then, in the light of subsequent research, Vomeridens is a possibility. Vomeridens has considerable persistent cartilage in the skull and pectoral girdle, a partially persistent notochord, and extensive subcutaneous lipid deposits, as well as a deep lipid deposit around the most posterior caudal vertebrae (Kuhn et al., 2011) [220]. Thin skin covers all fins, suggesting minimal contact with the substrate.Figure 24. Divergence in head shape among bathydraconids. Note the greater head lengths in Gymnodraco, Gerlachea, and Parachaenichthys, and the enlarged symphyseal teeth on the dentary of Gymnodraco. Psilodraco, Gymnodraco, Geralchea, and Parachaenichthys are gymnodraconines; others are bathydraconins. Modified from Balushkin and Voskoboinikova (2011, p. 19, Figure 3) [194], with permission of the authors.Figure 24. Divergence in head shape among bathydraconids. Note the greater head lengths in Gymnodraco, Gerlachea, and Parachaenichthys, and the enlarged symphyseal teeth on the dentary of Gymnodraco. Psilodraco, Gymnodraco, Geralchea, and Parachaenichthys are gymnodraconines; others are bathydraconins. Modified from Balushkin and Voskoboinikova (2011, p. 19, Figure 3) [194], with permission of the authors.
![Diversity 16 00214 g024]() Figure 25. Divergence in the shape of the premaxillae (a), dentaries (b), and teeth of bathydraconids. Note variation in sizes of the ascending, articular, and postmaxillary processes of the premaxillae, the enlarged teeth in Psilodraco and Gymnodraco, and the enlarged mandibular lateral line pores (arrows) in the dentaries of the bathydraconins Akarotaxis and Prionodraco. Modified from Balushkin and Voskoboinikova (2011, p. 58, Figures 21 and 22) [194], with permission of the authors.Figure 25. Divergence in the shape of the premaxillae (a), dentaries (b), and teeth of bathydraconids. Note variation in sizes of the ascending, articular, and postmaxillary processes of the premaxillae, the enlarged teeth in Psilodraco and Gymnodraco, and the enlarged mandibular lateral line pores (arrows) in the dentaries of the bathydraconins Akarotaxis and Prionodraco. Modified from Balushkin and Voskoboinikova (2011, p. 58, Figures 21 and 22) [194], with permission of the authors.
Figure 25. Divergence in the shape of the premaxillae (a), dentaries (b), and teeth of bathydraconids. Note variation in sizes of the ascending, articular, and postmaxillary processes of the premaxillae, the enlarged teeth in Psilodraco and Gymnodraco, and the enlarged mandibular lateral line pores (arrows) in the dentaries of the bathydraconins Akarotaxis and Prionodraco. Modified from Balushkin and Voskoboinikova (2011, p. 58, Figures 21 and 22) [194], with permission of the authors.Figure 25. Divergence in the shape of the premaxillae (a), dentaries (b), and teeth of bathydraconids. Note variation in sizes of the ascending, articular, and postmaxillary processes of the premaxillae, the enlarged teeth in Psilodraco and Gymnodraco, and the enlarged mandibular lateral line pores (arrows) in the dentaries of the bathydraconins Akarotaxis and Prionodraco. Modified from Balushkin and Voskoboinikova (2011, p. 58, Figures 21 and 22) [194], with permission of the authors.![Diversity 16 00214 g025]()


2.7. Morphology Stage—Axis 7: Neuromorphology (Brain and Sensory Systems, including Intraocular Vasculature)
Background. As systems that collect and process information crucial for the interpretation of their environment, the brains and sense organs of fishes frequently display habitat-specific morphology that is reflected in the relative sizes of areas of the brain, especially sensory regions (Huber and Rylander, 1992; Kotrschal and Palzenberger, 1992; Wagner, 2001, 2003; Deng et al., 2013) [221,222,223,224,225]. Cold, deep, dark Antarctic waters are sometimes likened to the deep sea where there are numerous examples of brain and sense organ specialization including, for example, liparids (Munk, 1964; Stein, 2016) [226,227] and ophidiids (Fine et al., 1987) [228]. The brain, and its developmental derivative the retina, are the most metabolically demanding tissues in vertebrates (Country, 2017) [229], and therefore are subject to evolutionary reduction in mass among species living in energy-limited environments. Increasing depth, a proxy for decreasing basal metabolic rate in fishes, is positively correlated with a decreased brain mass in diverse epipelagic, mesopelagic, and bathypelagic marine teleosts (Iglesias et al., 2015) [230]. This “depth effect” is evident in some «cryonotothenioids» and manifest by a reduction in the neuropil and therefore in the size of brain lobes that leads to a “stalked” appearance, meaning that the neural axis (brain stem) is visible between the reduced lobes of the brain. This depth-related morphology is not unique to «cryonototheniods» and is not an “Antarctic” attribute.
Detailed gross morphological and histological descriptions are available for the brains of 15 species representing six families, including all «cryonotothenioid» families with the exception of the Harpagiferidae. There is also information on sense organ morphology and intraocular vasculature for many additional species (Montgomery and Macdonald, 1987; Eastman, 1988b; Montgomery et al., 1988, 1994; Pankhurst and Montgomery, 1989, 1990; Janssen et al., 1993; Eastman and Lannoo, 1995, 2003a,b, 2004, 2007, 2008, 2011; Janssen, 1996; Montgomery, 1997; Lannoo and Eastman, 2000; Wujcik et al., 2007; Eastman and La Mesa, 2021) [60,231,232,233,234,235,236,237,238,239,240,241,242,243,244,245,246,247,248]. This section provides an overview of neuromorphological disparity.
- The brains of the bovichtids Bovichtus diacanthus (Figure 27a,c) and Cottoperca trigloides (Figure 27b,d). Bovichtus diacanthus inhabits tidepools and subtidal areas to depths of 20 m at Tristan da Cunha (37° S) while C. trigloides live at 5–310 m around the Falklands/Malvinas Islands and on the Patagonian shelf. Among the percoid families with known brain morphology, bovichtid brains resemble those of butterflyfishes (chaetodontids), angelfishes (pomacanthids), and cichlids (Bauchot et al., 1989; Huber et al., 1997) [249,250]. The brains of both bovichtids have a well-developed telencephalon and tectum. The telencephalon of C. trigloides is prominently lobed, more so than in any other notothenioid. The corpus cerebelli is relatively small and upright and, unlike other notothenioids, has prominent transverse sulci on the dorsal and caudal surfaces. Areas for lateral-line mechanoreception (eminentia granularis and crista cerebellaris) are also conspicuous but olfactory, gustatory, and somatosensory areas are not prominent. The olfactory nerves are also small. The brain of B. diacanthus is similar to that of C. trigloides but the anterior lateral-line nerve complex is larger, likely because of its innervation of the neuromasts in the extensively branched cephalic lateral canals. Both species also have large optic nerves and duplex (cone and rod) retinae that are 1.25-fold thicker and have nearly 5-fold more photoreceptors than those of most of the «cryonotothenioids».Figure 27. Lateral and dorsal views of brains and cranial nerves of the bovichtids Bovichtus diacanthus (a,c) and Cottoperca trigloides (b,d). From Eastman and Lannoo (2007) [242]. Abbreviations for this and subsequent brain illustrations: ADLL, anterodorsal lateral line nerve complex; AVLL, anteroventral lateral line nerve complex; CC, crista cerebellaris of the rhombencephalon; CCb, corpus division of the cerebellum; Dd, dorsodorsal subdivision of the telencephalon; Dl, dorsal lateral subdivision of the telencephalon; Dm, dorsal medial subdivision of the telencephalon; EG, eminentia granularis division of the cerebellum; Ha, habenula; IL, inferior lobe of the diencephalon; OB, olfactory bulb; Pit, pituitary gland; PLL, posterior lateral line nerve complex; PreO, preoptic area; SN1, first spinal nerve; SN2, second spinal nerve; SN3, third spinal nerve; SV, saccus vasculosus; Tec, tectum of the mesencephalon; Tel, telencephalon; I, olfactory nerve; II, optic nerve; III, oculomotor nerve; IV, trochlear nerve; V, trigeminal nerve; VII, facial nerve; VIII, auditory/vestibular nerve; IX, glossopharyngeal nerve; X, vagus nerve.Figure 27. Lateral and dorsal views of brains and cranial nerves of the bovichtids Bovichtus diacanthus (a,c) and Cottoperca trigloides (b,d). From Eastman and Lannoo (2007) [242]. Abbreviations for this and subsequent brain illustrations: ADLL, anterodorsal lateral line nerve complex; AVLL, anteroventral lateral line nerve complex; CC, crista cerebellaris of the rhombencephalon; CCb, corpus division of the cerebellum; Dd, dorsodorsal subdivision of the telencephalon; Dl, dorsal lateral subdivision of the telencephalon; Dm, dorsal medial subdivision of the telencephalon; EG, eminentia granularis division of the cerebellum; Ha, habenula; IL, inferior lobe of the diencephalon; OB, olfactory bulb; Pit, pituitary gland; PLL, posterior lateral line nerve complex; PreO, preoptic area; SN1, first spinal nerve; SN2, second spinal nerve; SN3, third spinal nerve; SV, saccus vasculosus; Tec, tectum of the mesencephalon; Tel, telencephalon; I, olfactory nerve; II, optic nerve; III, oculomotor nerve; IV, trochlear nerve; V, trigeminal nerve; VII, facial nerve; VIII, auditory/vestibular nerve; IX, glossopharyngeal nerve; X, vagus nerve.
![Diversity 16 00214 g027]()

- The brain of Eleginops maclovinus, the sister taxon of the «cryonotothenioids». The euryhaline E. maclovinus is migratory and catadromous in the Falklands/Malvinas Islands where it lives at water depths of 0–250 m and temperatures of 0–15 °C (Brickle et al., 2005a,b) [182,183]. As with bovichtids, the brain of E. maclovinus is similar to those of visually oriented temperate and tropical perciforms. The tectum is smaller than in bovichtids but E. maclovinus has well-developed olfactory bulbs and nerves, the latter nearly as large as the optic, and the mechanoreceptive lateral-line areas are also well-developed. There is a large caudally projecting corpus cerebelli (Figure 28a,b). Eye diameter is about twofold smaller in E. maclovinus than in many «cryonotothenioids»; the duplex retina has single and occasional twin cones. The olfactory rosette has 50–55 lamellae, a large number for any notothenioid and, given the large size of the olfactory nerve, olfaction is likely an important modality in this species. Eleginops maclovinus, and some species of «cryonotothenioids», also share an expansion of the proximal olfactory nerve immediately rostral to the olfactory bulb. The expansion, of unknown functional significance, consists of fascicles of olfactory nerve fibers that have an irregular rather than parallel orientation.

- The brains of nototheniids Dissostichus mawsoni and Pleuragramma antarcticum exhibit the most disparate morphology among «cryonotothenioids». Both species occupy the pelagic biotope in high-latitude shelf waters and have relatively wide bathymetric ranges. However, D. mawsoni is a large, migratory, primarily piscivorous predator while P. antarcticum is a medium-sized zooplanktivore. The brain of D. mawsoni (Figure 29a,b) displays some phylogenetically persistent morphology in its overall resemblance to that of Eleginops maclovinus, especially in the large olfactory nerves and bulbs, the proximal swellings of the olfactory nerves, the lobation of the telencephalon, and the relatively large caudally projecting corpus cerebelli. The brain of Pleuragramma (Figure 29c,d) does not retain phylogenetically persistent morphology and is reduced in mass. The corpus cerebelli is small, consistent with the inactivity of this species. Unlike other «cryonotothenioids», the olfactory bulbs are separated, or stalked, away from the telencephalon to reveal the medial and lateral olfactory tracts (arrows in Figure 29c). Stalking between the telencephalon and tectum is also evident (Figure 29d). Mechanosensation is well developed in Pleuragramma, more so than any other «cryonotothenioid» examined to date, as evident by the relatively large eminentia granularis of the cerebellum and crista cerebellaris.

The retinal histology of the two species encompasses the extremes among «cryonotothenioids». Dissostichus mawsoni (Figure 30a,c) has a rod-dominated retina with a high degree of summation. It is suited for vision under conditions of low light at depths > 1000 m. The retina of Pleuragramma (Figure 30b,d) is more cellular with many small single cones and rods, and resembles that of Eleginops maclovinus. It appears suitable for vision under various degrees of illumination as encountered in diel vertical migration, a documented behavior in parts of its range, and for detecting small prey such as calanoid copepods.
Pleuragramma antarcticum exemplifies an interesting evolutionary phenomenon at play in the neural divergence of «cryonotothenioids». In this species, the pedomorphic retention of neuromasts on the skin surface has resulted in detrimental changes to the essential role played by neuromasts in lateral-line mechanosensation. The incomplete development of the bony canals that normally contain the neuromasts was a disaptation (evolutionary loss of function) that resulted in the loss of the mechanical filtering properties provided by the canals. However, the eminentia granulares and cristae cerebellaris of the medulla are enlarged and functionally substitute for the canals by canceling the detrimental effect of the self-generated noise associated with the trunk movement necessary for pelagic feeding (Montgomery and Clements, 2000) [251].
- The brains of nototheniids of the genus Trematomus are similar. The brains of six species have been studied and, as represented by T. bernacchii (Figure 31), are generally comparable in proportions to that of Dissostichus mawsoni, although the corpus cerebelli are smaller. However, there is interspecific variation among the six species that reflects habitat depths and activity levels. The sister species T. lepidorhinus and T. loennbergii occupy the deep shelf-upper slope niche and exhibit differences in the size of brain regions, and divergence in all sensory systems. The morphology is unequivocally differentiating for habitat depths and conclusively documents a shift toward reliance on non-visual senses in T. loennbergii, consistent with collection data indicating that this species has a greater depth of peak abundance than T. lepidorhinus (Eastman and La Mesa, 2021) [60].

- The brain of the artedidraconid Dolloidraco longedorsalis. This species is sedentary, living on the substrate at 99–1243 m. It has a relatively long mental barbel and its brain is reduced in size. The telencephalon is small and the brain exhibits “stalking” (arrows in Figure 32a,b) with a visible neural axis. The corpus cerebelli is also relatively small but mechanosensation is well-developed based on the appearance of the eminentia granularis and crista cerebellaris (Eastman & Lannoo 2003a) [239]. However, the most distinctive features of the brain, apparent in histological sections, are the enlarged chief sensory nucleus of the trigeminal nerve and the well-developed thalamic and telencephalic nuclei of the ascending pathway, features that are correlated across artedidraconids with barbel length. Artedidraconids have single a mental barbel and its function is not understood (Eakin & Eastman, this volume) [252]. The barbels are unusual in lacking taste buds. Branches of the trigeminal nerve do extend into the skin to supply general sensation. Silver staining reveals that the barbel of D. longedorsalis has small fibers, possibly somatosensory and proprioceptive, in both the dermis and epidermis. The barbel, at least in long-barrelled species, may serve as a tactile organ and extend this modality anterior to the mouth to assist in locating benthic prey. Somatosensation is not a dominant sense in long-barbelled artedidraconids, but supplemental to lateral-line mechanosensation.

- Disparity is evident in the brains of species representing the two bathydraconid subfamilies. The brain of the relatively shallow-living Gymnodraco acuticeps (Gymnodraconinae, Figure 33a,b) is similar to that of Trematomus bernacchii (Figure 31). However, the brain of Akarotaxis nudiceps of the deeper-living Bathydraconinae exhibits an overall reduction in the neuropil and in the size of lobes that results in stalking of the brain, but the eminentia granulares and crista cerebellares remain prominent. (Figure 33b,d). The bathydraconins have rod-dominated retinae, dorsolaterally oriented eyes (Figure 24) and an expanded cephalic lateral-line system consisting of large cephalic lateral-line pores, wide head canals (Figure 34d), hypertrophied canal neuromasts and, in some species, an enlarged dorsal branch of the anterior line nerve (Figure 33b,d) (Eastman and Lannoo, 2003b) [240]. The wide canals probably enhance the sensitivity of the neuromasts to near-field particle motion (Coombs et al., 1988; Montgomery & Pankhurst 1997) [253,254]. In addition, the sagittal otoliths of the inner ear are elongated and several-fold larger in the Bathydraconinae, specifically Prionodraco evansii, Bathydraco marri, and A. nudiceps (Figure 34b,d) compared to the small round sagittae of G. acuticeps (Figure 34a). Some bathydraconines hover above the substrate and are more active than gymnodraconines. Otoliths respond to the particle displacement, velocity, and acceleration components of underwater sound, but it is not known how the size and shape of the sagittae are related to the behavior and habitat of any species of fish (Popper and Hawkins, 2020) [255].Figure 33. Lateral and dorsal views of the brains and cranial nerves of the bathydraconids Gymnodraco acuticeps (a,c) and Akarotaxis nudiceps (b,d). Arrows indicate areas of stalking of the brain where lobes are reduced and the brain axis is visible. From Eastman and Lannoo (2003b) [240]. Abbreviations as in Figure 27.Figure 33. Lateral and dorsal views of the brains and cranial nerves of the bathydraconids Gymnodraco acuticeps (a,c) and Akarotaxis nudiceps (b,d). Arrows indicate areas of stalking of the brain where lobes are reduced and the brain axis is visible. From Eastman and Lannoo (2003b) [240]. Abbreviations as in Figure 27.
![Diversity 16 00214 g033]() Figure 34. Slightly oblique left lateral radiographs of the bathydraconids Gymnodraco acuticeps (a), Prionodraco evansii (b), Bathydraco marri (c), and Akarotaxis nudiceps (d). Radiographs were overexposed to emphasize the deeply located and differently sized saccular otoliths in these specimens of similar lengths (105–114 mm SL). The bony canals of the supraorbital lateral line are also visible (arrows)—small in Gymnodraco but larger in the three bathydraconines, especially Akarotaxis. From Eastman and Lannoo (2003b) [240].Figure 34. Slightly oblique left lateral radiographs of the bathydraconids Gymnodraco acuticeps (a), Prionodraco evansii (b), Bathydraco marri (c), and Akarotaxis nudiceps (d). Radiographs were overexposed to emphasize the deeply located and differently sized saccular otoliths in these specimens of similar lengths (105–114 mm SL). The bony canals of the supraorbital lateral line are also visible (arrows)—small in Gymnodraco but larger in the three bathydraconines, especially Akarotaxis. From Eastman and Lannoo (2003b) [240].
Figure 34. Slightly oblique left lateral radiographs of the bathydraconids Gymnodraco acuticeps (a), Prionodraco evansii (b), Bathydraco marri (c), and Akarotaxis nudiceps (d). Radiographs were overexposed to emphasize the deeply located and differently sized saccular otoliths in these specimens of similar lengths (105–114 mm SL). The bony canals of the supraorbital lateral line are also visible (arrows)—small in Gymnodraco but larger in the three bathydraconines, especially Akarotaxis. From Eastman and Lannoo (2003b) [240].Figure 34. Slightly oblique left lateral radiographs of the bathydraconids Gymnodraco acuticeps (a), Prionodraco evansii (b), Bathydraco marri (c), and Akarotaxis nudiceps (d). Radiographs were overexposed to emphasize the deeply located and differently sized saccular otoliths in these specimens of similar lengths (105–114 mm SL). The bony canals of the supraorbital lateral line are also visible (arrows)—small in Gymnodraco but larger in the three bathydraconines, especially Akarotaxis. From Eastman and Lannoo (2003b) [240].![Diversity 16 00214 g034]()


- The brain of the channichthyid Chionodraco hamatus (Figure 35a,b). There is relatively little interspecific variation among the brains of the seven channichthyid species that have been examined (Eastman and Lannoo, 2004) [241], and their brains resemble that of Gymnodraco acuticeps. The brain of Chionodraco hamatus differs from that of G. acuticeps in that it has a slightly regressed telencephalon resulting in stalking in this region (Figure 35b, arrow). It also has a larger tectum and a more dorsally-projecting corpus cerebelli than G. acuticeps. The mechanoreceptive areas are well-developed. Although the olfactory nerves are relatively small, olfactory lamellae are more numerous than in other «cryonotothenioids», but this may simply reflect their larger body lengths. The eyes are large and laterally oriented, the optic nerves are relatively large and the retinae have cones and rods in all eight species where this has been examined. The most unusual morphological feature is not in the nervous tissue per se but in the blood supply of their eyes considered below.

- The neuromorphological divergence of «cryonotothenioids» is accompanied by a reduction, transposition, and subsequent expansion of ocular vascular structures. The teleost retina lacks intrinsic blood vessels and is usually supplied by vessels located on the scleral (outer) surface and, less frequently, by vessels on the vitrad (inner or pre-retinal) surface. Teleosts have three ocular vascular structures that are not present in mammals: the choroid rete, the lentiform body (a small rete), and the falciform process that supplies the retractor lentis (Walls, 1942; Nicol, 1989) [256,257]. The choroid rete is especially labile and has been frequently lost in unrelated teleost taxa (Wittenberg and Haedrich, 1974; Berenbrink et al., 2005) [258,259]. Although ocular vascular structures (Table 2 and Table 3) are well developed in phylogenetically basal notothenioids (Eastman, 1988b, 2006) [232,260], there has been a phylogenetic reduction in these structures among most «cryonotothenioids». Some bathydraconids and all channichthyids have lost the choroid rete and the choriocapillaris, the capillary bed supplying the retina (Eastman and Lannoo, 2004) [241]. The ophthalmic artery may be vestigial or lost as well. However, the optic artery branches into a densely arranged series of hyaloid arteries at the vitreoretinal interface, and these supply the retina. Channichthyids possess the most extensive system of hyaloid arteries documented to date among teleosts (Eastman 1988b; Eastman and Lannoo, 2004; Wujcik et al., 2007) [232,241,248]. These vessels are ≈60 μm, twice the diameter of those in red-blooded nototheniids, and have an extremely dense, anastomosing branching pattern (Figure 36g,h) unlike that in basal notothenioids (Figure 36a,b) and other «cryonotothenioids» (Figure 36c–f). With the loss of the primary vasculature from the scleral surface, there has been a transposition and expansion of the hyaloid vessels on the vitrad surface of the retina. However, as a compensatory response, a dense vascular array in a pre-retinal position may be maladaptive in that: (1) the vessels are situated farther from the photoreceptors, hence the diffusion distance to these oxygen-demanding elements is longer, and (2) the acuity of the visual image is likely compromised by the interposition of vessels containing whitish, non-transparent blood. The extent of development of the hyaloid vasculature is also inversely correlated with levels of circulating hemoglobin among «cryonotothenioids» (Wujcik et al., 2007) [248].Figure 36. (a–h) Eyes of eight species of notothenioids showing varying degrees of development of the hyaloid (preretinal) arteries on the vitrad surface of retinae as visualized by arterial perfusions of Microfil®. The cornea, iris, lens, and vitreous body have been removed; the retractor lentis (RL) muscle is situated ventrally; hyaloid arteries drain to the annular vein (AV). With the exception of (c), all are left eyes. Phylogenetically basal Bovichtus diacanthus (a) has a sparse pattern of hyaloid branches and an open choroid fissure (arrow) is also evident. Eleginops maclovinus (b) lacks branches to the retina and the hyaloid artery continues anteriorly to supply the retractor lentis. Among cryonotothenioids, Notothenia coriiceps (c) and Trematomus borchgrevinki (d) have a widely spaced radial branching pattern emanating from the hyaloid artery at its entry point at the optic disk. Red-blooded bathydraconids Gymnodraco acuticeps (e) and Parachaenichthys georgianus (f) have a denser radial pattern. White-blooded channichthyids Pseudochaenichthys georgianus (g) and Chaenocephalus aceratus (h) have extremely dense networks of anastomosing vessels that are unusual among fishes, with radial (g) or linear (h) branching patterns. Bovichtus diacanthus, E. maclovinus, and N. coriiceps have a choroid rete; the other five species do not.Figure 36. (a–h) Eyes of eight species of notothenioids showing varying degrees of development of the hyaloid (preretinal) arteries on the vitrad surface of retinae as visualized by arterial perfusions of Microfil®. The cornea, iris, lens, and vitreous body have been removed; the retractor lentis (RL) muscle is situated ventrally; hyaloid arteries drain to the annular vein (AV). With the exception of (c), all are left eyes. Phylogenetically basal Bovichtus diacanthus (a) has a sparse pattern of hyaloid branches and an open choroid fissure (arrow) is also evident. Eleginops maclovinus (b) lacks branches to the retina and the hyaloid artery continues anteriorly to supply the retractor lentis. Among cryonotothenioids, Notothenia coriiceps (c) and Trematomus borchgrevinki (d) have a widely spaced radial branching pattern emanating from the hyaloid artery at its entry point at the optic disk. Red-blooded bathydraconids Gymnodraco acuticeps (e) and Parachaenichthys georgianus (f) have a denser radial pattern. White-blooded channichthyids Pseudochaenichthys georgianus (g) and Chaenocephalus aceratus (h) have extremely dense networks of anastomosing vessels that are unusual among fishes, with radial (g) or linear (h) branching patterns. Bovichtus diacanthus, E. maclovinus, and N. coriiceps have a choroid rete; the other five species do not.
![Diversity 16 00214 g036]()

- Overview and summary of brain and sense organ disparity among «cryonotothenioids». The Antarctic environment does not require novel neural and sense organ morphology—nothing beyond that provided by the percoid phenotype with physiological adaptations for low temperatures. «Cryonotothenioid» brains and sense organs resemble those of temperate percoids more than those of phylogenetically older deep-sea species. Habitat depth does influence neuromorphology among «cryonotothenioids», but there are no sensory specialists—species possessing marked hypertrophy of and reliance on primarily one modality, as seen in some deep-sea fishes. Most «cryonotothenioids» retain a relatively balanced array among their sensory systems. Compared with bovichtids (Figure 27) and Eleginops maclovinus (Figure 28), these are the most conspicuous features of neuromorphological disparity:
- There is a reduction in overall brain cellularity in the «cryonotothenioids» compared to Eleginops maclovinus. This is especially evident in taxa with visible “stalking” of the base of the brain: Pleuragramma, Dolloidraco, the Bathydraconinae, and some channichthyids (Figure 29c,d, Figure 32, Figure 33b,d and Figure 35b). It is also evident in the sister species Trematomus with different depth preferences (Eastman & La Mesa 2021) [60]. This is typical depth-related morphology and not unique to the Antarctic environment.
- Sensory regions of the brain are the most variable, especially the mechanosensory and olfactory regions.
- Multimodal integration and association areas of the brain, the corpus cerebelli, tectum, and telencephalon, exhibit moderate variation in size, shape, and in the case of the telencephalon, lobation patterns also vary. The size of the corpora cerebelli ranges from large and caudally projecting in the migratory Dissostichus (Figure 29a), to small and round in the relatively inactive Pleuragramma, Dolloidraco, and Akarotaxis (Figure 29c, Figure 32a, and Figure 33b).
- Sensory cranial nerves exhibit interspecific size differences, especially the olfactory, optic, and anterior lateral-line nerves. There are no species with extreme reduction in any of these nerves. However, compared to the large size of the optic nerves in bovichtids, Eleginops, Dissostichus, Pleuragramma, and Trematomus spp., those of most other «cryonotothenioids» are relatively smaller.
- All species have well-developed eyes and duplex (cone and rod) retinae (Figure 30). This is not surprising as most acanthomorphs are visually oriented (Demski, 2003) [261]. There are, however, large differences in the cone-to-rod ratios, with fewer cones in most deeper-living species. There are also a phylogenetic reductions in the vascularity, thickness and cellularity of the retina (Table 2 and Table 3).
- In artedidraconids with long mental barbels, branches of the trigeminal nerves likely convey somatosensory (tactile) and proprioceptive (position) information from the barbel to the central nervous system.
- Neither oral nor cutaneous taste is notably developed in «cryonotothenioids». Most species have a few taste buds in and around the oral cavity. The barbels of artedidraconids lack taste buds. It is possible that there are solitary chemosensory cells in the skin.
- Chemosensation is understudied but potentially important in «cryonotothenioids». Nothing is known about pheromonal detection and communication in «cryonotothenioids»; however, based on findings in other fishes, it could be involved in activities such as migration to spawning sites and finding of mates (Vrieze et al., 2011; Lim and Sorensen, 2012) [263,264]. Given its well-developed olfactory system and spawning migrations, Dissostichus mawsoni would be a promising subject for this type of research. Trematomus spp. are attracted to fishery waste (Pakhomov, 1998) [265] and can detect and track an odor plume emanating from fish used as bait in experiments (Montgomery et al., 1999) [266]. Scavenging is also known through experimentation designed to test its prevalence (Smale et al., 2007) [267], and by the success of the odor plumes from bait used in the industrial longline fishery for Dissostichus spp.
3. The Fossil Record as Context for the Radiation of «Cryonotothenioids»
The Eocene La Meseta fossil fishes from Seymour Island. Following the Cretaceous–Palaeogene mass extinction event, there was a rapid diversification of marine acanthomorph fish clades (Alfaro et al., 2018) [268]. Therefore it is not surprising that fossil fishes from late Paleocene and Eocene fossil deposits are taxonomically diverse and include some phylogenetically derived taxa. An example is the Eocene La Meseta Fauna from Seymour Island located then at a palaeolatitude of ≈60° S, similar to its current position. Annual sea surface temperatures in this area during the Middle and Late Eocene are estimated to have been 10–17 °C (Douglas et al., 2014) [269]. The units of the La Meseta Formation span most of the Eocene from 56.0–33.9 Ma (Ivany et al., 2008) [270]. Therefore the fossil fish assemblage does not represent a snapshot of a natural community, ecosystem, or habitat at a point in time, but rather material amalgamated from cool-temperate marine associations that existed over much of the Eocene (Přikryl & Vodrážka 2012) [271]. Recent fieldwork on Seymour Island has not only increased the number of known species but also highlighted the family-level taxonomic disparity and the discontinuity between this assemblage and the modern fauna (Kriwet 2005; Kriwet & Hecht 2008; Engelbrecht et al., 2016a,b, 2017a,b, 2018; Kriwet et al., 2016; Schwarzhans et al., 2016) [272,273,274,275,276,277,278,279,280]. The total described material, summarised by family in Table 4, consists of 63 species including 39 chondrichthyans and 24 actinopterygians. Although chondrichthyan diversity in the La Meseta Fauna was high and, in the case of sharks, is comparable to modern tropical faunas (Reguero et al., 2013) [281], the apparent dominance by chondrichthyans is probably an artifact attributable to the well-preserved enamel-like teeth that, in many cases, are sufficiently distinctive that they can be identified to genus or even species. Actinopterygian fossils, on the other hand, are usually isolated bones, fin spines, and otoliths that are more difficult to diagnose at low taxonomic levels. Nevertheless, there is reasonable taxonomic diversity at the family level among actinopterygians, and gadiforms are prominent and represented by major lineages including morids, macrourids, merlucciids, and gadids (Kriwet and Hecht, 2008; Schwarzhans et al., 2016) [273,280]. Although many of the chondrichthyan and actinopterygian species from the La Meseta Fauna were probably endemic to Eocene Antarctic or Southern Hemisphere waters, at higher taxonomic levels the La Meseta Fauna was cosmopolitan.
Uncertainty surrounds some gadiform/notothenioid fossils. There are no fossils from the La Meseta Formation that have been convincingly identified as notothenioid. The skeletal tissues of «cryonotothenioids» are not ideal for fossilization. A relatively low skeletal mass is also characteristic of the non-Antarctic Eleginops maclovinus (Eastman et al., 2014) [42]. The bone is cancellous and the skeleton contains a considerable component of cartilage. Furthermore, the identification of fossils as notothenioid is hampered by the absence of osteological synapomorphies for the group. Notothenioids are diagnosed, in part, by an unremarkable suite of shared characters that may be unique in combination, with the osteological elements consisting of three pectoral radials, posterior pleural ribs that are poorly developed or absent, and non-pungent fin spines (Near et al., 2015) [2].
A dorsal neurocranium from the fifth unit of the Tertiary Eocene La Meseta Formation (TELM 5) now dated at 52.8–49.5 Ma (Ivany et al., 2008; Schwarzhans et al., 2016) [270,280], was originally identified as a gadiform by Eastman & Grande (1991) [282] and subsequently redescribed as a notothenioid (Proeleginops grandeastmanorum) by Balushkin (1994) [283]. Recent discoveries of fossil gadiforms include an incomplete skull and otoliths of a macrourid (Kriwet and Hecht, 2008) [273], a merlucciid jaw (Claeson et al., 2012) [284], and six otolith-based gadiform taxa, findings that increase the probability that Proeleginops is a gadiform (Schwarzhans et al., 2017) [280].
There is another La Meseta fossil fish taxon with a contested identity—«mesetaichthys». Partial premaxillary bones with large canine teeth, considered as possibly gadiform, but ultimately “indeterminate” (Grande and Eastman, 1986, Figure 5a–i) [285], have been supplemented with additional material from larger (90–150 cm) specimens (Jerzmańska & Świdnicki, 1992) [286]. Based on similarities to merlucciid jaws, this material has been collectively identified as gadiform and given the informal, and presumably rank-free, name «mesetaichthys» (Jerzmańska and Świdnicki, 1992) [286]. With an additional partial dentary and teeth similar to «mesetaichthys», and based on shared similarities with the dentaries of «mesetaichthys» and the modern Merluccius, all these specimens have been collectively referred to as the Merlucciidae (hakes) (Claeson et al., 2012) [284]. However, Bieńkowska-Wasiluk et al. (2013) [287], based on their evaluation of large jaw fragments with teeth (from TELM 7, 34.2 Ma), identified their new material and most previous «mesetaichthys» material as the new species Mesetaichthys jermanskae, a notothenioid. They indicate that the key diagnostic feature is in the dentition, specifically “the external ridges and fissures in the enamaloid at the base of the teeth proper” (p. 571). This feature is also present in the teeth of the nototheniid Dissostichus mawsoni, but it is not diagnostic of notothenioids. These axial striae on the external surface indicate the presence of plicidentine internally. Plicidentine is radially folded dentine that borders the pulp cavity near the base of the tooth where it serves to increase the surface area for attachment of the tooth to the underlying bony pedicle, an advantage in resisting dental strain during feeding (Germain et al., 2016, p. 76) [288]. Plicidentine is present in many actinopterygians, and in some species, it is also manifest externally as folds or striae in the enameloid. These characteristics are “ubiquitous” among the enlarged teeth of diverse predatory teleosts (Germain et al., 2016, p. 81) [288]. Therefore this character is too widely distributed to be diagnostic and there remain no fossils from the La Meseta Formation that can be definitively identified as notothenioid.
An otolith described as Harpagifer? morgansi, a putative harpagiferid from the Early Miocene (23–16 Ma), is known from deposits of the South Island of New Zealand (Schwarzhans, 2019, pp. 225–227) [289]. Although the overall outline and the shape of the sulcus closely resemble those of the modern species of Harpagifer, its designation is considered “tentative” by Schwarzhans. The age of this otolith stands in contrast to the age of ≈9 Ma for Harpagifer as inferred from nucleotide sequence data (Near et al., 2012) [7].
The Eocene fauna was replaced. The rise, decline, and replacement of taxa and faunas has occurred frequently over the evolutionary history of fishes. In the case of the La Meseta fishes, the timing and abruptness of their disappearance are unknown. Furthermore, there is no evidence in the fossil record of transitional fauna, and there has been minimal taxonomic carryover from the La Meseta Fauna into the modern fauna. It appears that there was nearly total replacement by «cryonotothenioids». Eighty-seven percent (230/264) of the modern shelf and upper slope species diversity is composed of «cryonotothenioids», liparids, and zoarcids (Figure 37a). Rajiids and gadiforms are the only taxa held in common with the La Meseta Fauna. As waters cooled through and after the Eocene, and as glacial and sea ice increased, there were reductions in available habitats and disruptions of food webs. Ecological opportunity is an essential precondition or facilitator for adaptive radiation (Schluter, 2000, p. 239; Grant and Grant 2008, p. 140) [13,83], and this was created by the local extinction of most components of the La Meseta Fauna. For example, without surviving clupeids (herrings) and large predatory sharks and billfishes, there were niche gaps in the water column for «cryonotothenioid» zooplanktivores (Pleuragramma, Aethotaxis) and large piscivorous predators (Dissostichus spp.).
In searching for notothenioid fossils, it might be productive to concentrate on younger strata such as the Early Miocene (≈22.6 Ma) Cape Melville Formation on King George Island (Whittle et al., 2014) [291] and the Early Pliocene (4.5–4.1 Ma) Sørsdal Formation, Marine Plain, Vestfold Hills, near Prydz Bay in East Antarctica (Quilty et al., 2000) [292]. The rationale is that the western Weddellian Province, centered on South America, was the initial site of «cryonotothenioid» diversification, and inferences from nucleotide sequence data suggest that most species appeared during the last 10–5 Ma or less (Near et al., 2012) [7], including species of the non-Antarctic genus Bovichtus (Near et al., 2019) [293].
Gadiform diversity and abundance were not maintained in the modern fauna. When considering faunal replacement, gadiforms are relevant because, if more had become established in the modern fauna, the outcome may have been different in the sense that ecological opportunities for «cryonotothenioids» might have been reduced. Although gadiforms were a dominant actinopterygian component of the La Meseta Fauna (Kriwet and Hecht, 2008; Schwarzhans et al., 2016) [273,280], they compose only 6% (15 species) of the modern shelf and slope fauna of Antarctica (Figure 37a). This minor representation is surprising given their historic presence in southern latitudes and their modern worldwide distribution. It is also puzzling that muraenolepidids, an early diverging linage endemic to the Southern Hemisphere (Roa-Varón & Ortí, 2009) [294] and the only gadiforms present in the modern subzero shelf waters, are not represented in the La Meseta Fauna. In the Arctic fauna, the 23 gadiform species are third in diversity to zoarcids and cottids (Figure 37b), but they compose the majority of the fish abundance (Mecklenburg et al., 2018, pp. 114, 146, 150) [290]. In the Arctic, they also diverged morphologically and ecologically, ranging in size from 25 cm to 200 cm in Gadus morhua, a species that attains a weight of 96 kg. Depth ranges are 0–3500 m and they occupy the benthic and pelagic realms (Mecklenburg et al., 2018) [290]. Some gadids also evolved an antifreeze glycopeptide de novo from non-coding DNA in the 18–13 Ma time frame (Baalsrud et al., 2018; Zhuang et al., 2019) [295,296]. Boreogadus saida is the most northerly-occurring species of fish, reaching about 88° N under sea ice in −1.9 °C waters (Mecklenburg et al., 2018) [290].
4. Overview of the Radiation
The evolutionary radiation of «cryonotothenioids» involved both lineage diversification and phenotypic divergence that encompassed the morphology and physiology of many body systems as well as organismal behavior. The three stages of the radiation involve these axes. For habitat, divergence was along a depth axis, ranging from tide pools to 2941 m, and along a biotope axis from benthic to pelagic. For morphology, the body size axis spans a 39-fold difference in length, and a body density axis reflects varying proportions of bony, cartilaginous, notochordal, and adipose tissues. A body shape axis is also evident—most species are fusiform or derivations thereof, and a few others are widened or elongated. The trophic morphology axis involves cephalic enlargement affecting the lengths, shapes, and masses of the jaw bones and the sizes of the oral and pharyngeal gapes. Along the neuromorphology axis, the brain and sense organs reflect the influence of depth, light intensity and activity levels, with no regression, extraordinary hypertrophy, or dominance by any single modality. For communication, the axes relate to reproduction and were not covered here given the recent summary by La Mesa et al. (2021) [46]. The axes involve egg size, spawning (whether pelagic or benthic), and the degree of parental care provided.
«Cryonotothenioids» dominate the modern fish fauna of the high-latitude shelf waters where trawls in the water column yield primarily Pleuragramma antarcticum (DeWitt, 1970; Donnelly et al., 2004) [14,18] and benthic trawls consist of other «cryonotothenioids» at levels of diversity, abundance, and biomass of, respectively, 77%, 92%, and 91%. This level of dominance is unusual in the marine realm. «Cryonotothenioids» were resilient during historical climate change—they possessed the physiological and ecological plasticity necessary for accommodation to the after-effects of episodes of Pliocene and post-Pliocene glacial advance and retreat. These encroachments, extreme but potentially opportunistic events (Coleman and Wernberg, 2020) [297], periodically limited the availability of large areas of the Antarctic shelf and probably created a patchwork of altered habitats with greater depths and different trophic conditions.
Antifreeze compounds are essential for the survival of fish in subzero oceanic waters and have evolved independently in many taxa (DeVries, 2020) [298]. The antifreeze glycopeptides of «cryonotothenioids» are a “key adaptation”. They are continuously synthesized, and active throughout life in limiting the size of ice crystals in the body, and therefore they enable survival in and access to biotopes and resources in subzero waters. They are estimated to have originated from a functionally unrelated pancreatic trypsinogen at 14–5 Ma (Chen et al., 1997) [299], and most «cryonotothenioid» diversity is thought to have appeared at 10–5 Ma (Near et al., 2012) [7]. Although the appearance of antifreeze glycopeptides preceded and was uncoupled from most «cryonotothenioid» diversification, they nevertheless constitute a key adaptation and a species-generating trait. A radiation does not necessarily have to emerge immediately after the appearance of the key innovation—it can be “long-delayed” because ecological opportunity is probably a more important factor in determining when the key innovation comes into play, and then allows diversification and divergence to proceed (Schluter, 2000, p. 181) [13].
Why «cryonotothenioids» and not some other taxon? Like many complex historical events, faunal replacements may leave us with loose ends, open questions, and no obvious explanation for a bewildering transformation presented by, in this case, a modern Antarctic fish fauna that is overwhelmingly dominated by a single taxon. How did this come about? Liparids and zoarcids (Figure 37a) are also found in Antarctic waters and, like «cryonotothenioids», exhibit high rates of speciation (Rabosky et al., 2018) [23]. Both taxa have the proven capacity to diversify in deep, cold, oligotrophic waters worldwide and to evolve antifreeze compounds (DeVries, 1983, 2020; Jung et al., 1995; Evans and Fletcher, 2001; DeVries and Cheng, 2005; Zhaung et al., 2019) [1,298,300,301,302,303]. Although liparids are especially speciose, neither they nor zoarcids rival the morphological and ecological diversity of the «cryonotothenioids». Their radiations, other than the divergence of many deep-living species (Anderson and Fedorov, 2004; Stein, 2012a,b, 2016; Linley et al., 2016) [55,227,304,305,306] and some gelatinous pelagic species (Eastman et al., 1994; Balushkin and Moganova, 2017; Gerringer et al., 2017) [307,308,309], did not replicate the other axes seen in «cryonotothenioids». For example, their ranges in body sizes do not include medium-large and large species and the liparid phenotype is gracile and may be unsuited for some habitats and feeding modes. The zoarcids with antifreeze proteins acquired these at 18 Ma, and the component of this lineage that migrated from the Arctic to the Southern Ocean arrived in the last few million years (Hobbs et al., 2020) [310], after most «cryonotothenioid» lineages were established.
Evolvability is “the capacity to rapidly produce a variety of different phenotypes exploiting the environment in novel ways” (Schluter 2000, p. 186) [13]. Perhaps the success of «cryonotothenioids» is attributable to their early colonization of shelf waters and to their robust habitat-bridging phenotype that, without morphological novelties but with the potential for heterochronic change, provided optimal “phenotypic evolvability” (Rabosky et al., 2013, p. 6) [311]. «Cryonotothenioids» are also ecologically and trophically plastic. This probably facilitated their resilience through the multiple stressors associated with the recurring episodes of climatic upheaval caused by advances and retreats of the ice sheet on the continental shelf, events that altered nearly everything about the existing habitat. «Cryonotothenioids» were never in such delicate balance with historic climates and habitats that they were unable to tolerate perturbations. Climate has oscillated over the millennia and, as it always has, will incessantly confront «cryonotothenioids».
Funding
Field and lab work was funded by US National Science Foundation grants ANT 94-16870 and ANT 04-36190 to JTE.
Institutional Review Board Statement
The author has followed all applicable national and institutional guidelines for the collection, care, and ethical use of research organisms and material in the conduct of the research, specifically those of the Ohio University Institutional Animal Care and Use Committees. No experiments were conducted on fishes during the field work and live fishes were not transported from Antarctica to Ohio University.
Data Availability Statement
All fish specimens, skeletons, histological slides, radiographs and laboratory notebooks associated with this work are catalogued into the Ichthyology Collection of the Yale University Peabody Museum of Natural History, New Haven, Connecticut, USA.
Acknowledgments
For providing advice, comments and permission to use photos and figures, I thank David G. Ainley (H.T. Harvey & Associates, Ecological Consultants), Esteban Barrera-Oro (Instituto Antárctico Argentino), Lisa Crockett (Ohio University), Richard R. Eakin (Guys Mills, Pennsylvania), Komoda Masazumi (NHK Japan), Thomas J. Near (Yale University), Werner Schwarzhans (Natural History Museum of Denmark, University of Copenhagen) and Olga S. Voskoboinikova and the late Arcady V. Balushkin (Zoological Institute, Russian Academy of Sciences). I am also grateful to Clive W. Evans and John Montgomery, both of the University of Auckland in New Zealand, for helpful comments on the manuscript. I also thank Jenny Gon for her final check of the manuscript.
Conflicts of Interest
The author declares that he has no conflicts of interest.
References
- Barnes, D.K.; Conlan, K.E. Disturbance, colonization and development of Antarctic benthic communities. Philos. Trans. R. Soc. B Biol. Sci. 2006, 362, 11–38. [Google Scholar] [CrossRef] [PubMed]
- Near, T.J.; Dornburg, A.; Harrington, R.C.; Oliveira, C.; Pietsch, T.W.; Thacker, C.E.; Satoh, T.P.; Katayama, E.; Wainwright, P.C.; Eastman, J.T.; et al. Identification of the notothenioid sister lineage illuminates the biogeographic history of an Antarctic adaptive radiation. BMC Evol. Biol. 2015, 15, 109. [Google Scholar] [CrossRef] [PubMed]
- Near, T.J.; MacGuigan, D.J.; Parker, E.; Struthers, C.D.; Jones, C.D.; Dornburg, A. Phylogenetic analysis of Antarctic notothenioids illuminates the utility of RADseq for resolving Cenozoic adaptive radiations. Mol. Phylogenetics Evol. 2018, 129, 268–279. [Google Scholar] [CrossRef] [PubMed]
- Zaffos, A.; Finnegan, S.; Peters, S.E. Plate tectonic regulation of global marine animal diversity. Proc. Natl. Acad. Sci. USA 2017, 114, 5653–5658. [Google Scholar] [CrossRef] [PubMed]
- Benton, M.J. The Red Queen and the Court Jester: Species Diversity and the Role of Biotic and Abiotic Factors through Time. Science 2009, 323, 728–732. [Google Scholar] [CrossRef] [PubMed]
- Anderson, J.B. Antarctic Marine Geology; Cambridge University Press: Cambridge, UK, 1999; 289p. [Google Scholar]
- Near, T.J.; Dornburg, A.; Kuhn, K.L.; Eastman, J.T.; Pennington, J.N.; Patarnello, T.; Zane, L.; Fernández, D.A.; Jones, C.D. Ancient climate change, antifreeze, and the evolutionary diversification of Antarctic fishes. Proc. Natl. Acad. Sci. USA 2012, 109, 3434–3439. [Google Scholar] [CrossRef] [PubMed]
- Pollard, D.; DeConto, R.M. Modelling West Antarctic ice sheet growth and collapse through the past five million years. Nature 2009, 458, 329–332. [Google Scholar] [CrossRef] [PubMed]
- Anderson, J.B.; Conway, H.; Bart, P.J.; Witus, A.E.; Greenwood, S.L.; McKay, R.M.; Hall, B.L.; Ackert, R.P.; Licht, K.; Jakobsson, M.; et al. Ross Sea paleo-ice sheet drainage and deglacial history during and since the LGM. Quat. Sci. Rev. 2014, 100, 31–54. [Google Scholar] [CrossRef]
- Pekar, S.F.; DeConto, R.M. High-resolution ice-volume estimates for the early Miocene: Evidence for a dynamic ice sheet in Antarctica. Palaeogeogr. Palaeoclim. Palaeoecol. 2006, 231, 101–109. [Google Scholar] [CrossRef]
- McKay, R. Did Antarctica initiate the ice age cycles? Science 2014, 346, 812–813. [Google Scholar] [CrossRef]
- Scherer, R.P.; DeConto, R.M.; Pollard, D.; Alley, R.B. Windblown Pliocene diatoms and East Antarctic Ice Sheet retreat. Nat. Commun. 2016, 7, 12957. [Google Scholar] [CrossRef] [PubMed]
- Schluter, D. The Ecology of Adaptive Radiation; Oxford University Press: Oxford, UK, 2000; 288p. [Google Scholar]
- Dewitt, H.H. The character of the midwater fish fauna of the Ross Sea, Antarctica. In Antarctic Ecology; Holdgate, M.W., Ed.; Academic Press: London, UK, 1970; Volume 1, pp. 305–314. [Google Scholar]
- Hubold, G.; Ekau, W. Midwater fish fauna of the Weddell Sea, Antarctica. In Fifth Congress of European Ichthyologists, Proceedings; Kullander, S.O., Fernholm, B., Eds.; Swedish Museum of Natural History: Stockholm, Sweden, 1987; pp. 391–396. [Google Scholar]
- Ekau, W. Demersal fish fauna of the Weddell Sea, Antarctica. Antarct. Sci. 1990, 2, 129–137. [Google Scholar] [CrossRef]
- Eastman, J.T.; Hubold, G. The fish fauna of the Ross Sea, Antarctica. Antarct. Sci. 1999, 11, 293–304. [Google Scholar] [CrossRef]
- Donnelly, J.; Torres, J.J.; Sutton, T.T.; Simoniello, C. Fishes of the eastern Ross Sea, Antarctica. Polar Biol. 2004, 27, 637–650. [Google Scholar] [CrossRef]
- Eastman, J.T. The nature of the diversity of Antarctic fishes. Polar Biol. 2005, 28, 93–107. [Google Scholar] [CrossRef]
- Koubbi, P.; Ozouf-Costaz, C.; Goarant, A.; Moteki, M.; Hulley, P.-A.; Causse, R.; Dettai, A.; Duhamel, G.; Pruvost, P.; Tavernier, E.; et al. Estimating the biodiversity of the East Antarctic shelf and oceanic zone for ecoregionalisation: Example of the ichthyofauna of the CEAMARC (Collaborative East Antarctic Marine Census) CAML surveys. Polar Sci. 2010, 4, 115–133. [Google Scholar] [CrossRef]
- Causse, R.; Ozouf-Costaz, C.; Koubbi, P.; Lamy, D.; Eléaume, M.; Dettaï, A.; Duhamel, G.; Busson, F.; Pruvost, P.; Post, A.; et al. Demersal Ichthyofaunal Shelf Communities from the Dumont D’urville Sea (East Antarctica). Polar Sci. 2011, 5, 272–285. [Google Scholar] [CrossRef]
- Hanchet, S.M.; Stewart, A.L.; Mcmillan, P.J.; Clark, M.R.; O’driscoll, R.L.; Stevenson, M.L. Diversity, relative abundance, new locality records, and updated fish fauna of the Ross Sea region. Antarct. Sci. 2013, 25, 619–636. [Google Scholar] [CrossRef]
- Rabosky, D.L.; Chang, J.; Title, P.O.; Cowman, P.F.; Sallan, L.; Friedman, M.; Kaschner, K.; Garilao, C.; Near, T.J.; Coll, M.; et al. An inverse latitudinal gradient in speciation rate for marine fishes. Nature 2018, 559, 392–395. [Google Scholar] [CrossRef]
- Eastman, J.T.; Mccune, A.R. Fishes on the Antarctic continental shelf: Evolution of a marine species flock? J. Fish Biol. 2000, 57 (Suppl. A), 84–102. [Google Scholar] [CrossRef]
- Janko, K.; Marshall, C.; Musilová, Z.; Van Houdt, J.; Couloux, A.; Cruaud, C.; Lecointre, G. Multilocus analyses of an Antarctic fish species flock (Teleostei, Notothenioidei, Trematominae): Phylogenetic approach and test of the early-radiation event. Mol. Phylogen. Evol. 2011, 60, 305–316. [Google Scholar] [CrossRef] [PubMed]
- Matschiner, M.; Hanel, R.; Salzburger, W. On the Origin and Trigger of the Notothenioid Adaptive Radiation. PLoS ONE 2011, 6, e18911. [Google Scholar] [CrossRef] [PubMed]
- Matschiner, M.; Colombo, M.; Damerau, M.; Ceballos, S.; Hanel, R.; Salzburger, W. The adaptive radiation of notothenioid fishes in the waters of Antarctica. In Extremeophile Fishes; Riesch, R., Tobler, M., Plath, M., Eds.; Springer: Cham, Switzerland, 2015; pp. 35–57. [Google Scholar]
- Lautrédou, A.-C.; Hinsinger, D.; Gallut, C.; Cheng, C.-H.; Berkani, M.; Ozouf-Costaz, C.; Cruaud, C.; Lecointre, G.; Dettai, A. Phylogenetic footprints of an Antarctic radiation: The Trematominae (Notothenioidei, Teleostei). Mol. Phylogenetics Evol. 2012, 65, 87–101. [Google Scholar] [CrossRef] [PubMed]
- Lecointre, G.; Améziane, N.; Boisselier, M.-C.; Bonillo, C.; Busson, F.; Causse, R.; Chenuil, A.; Couloux, A.; Coutanceau, J.P.; Cruaud, C.; et al. Is the species flock concept operational? The Antarctic Shelf case. PLoS ONE 2013, 8, e68787. [Google Scholar] [CrossRef] [PubMed]
- Wilson, L.A.B.; Colombo, M.; Hanel, R.; Salzburger, W.; Sánchez-Vilagra, M.R. Ecomorphological disparity in an adaptive radiation: Opercular bone shape and stable isotopes in Antarctic icefishes. Ecol. Evol. 2013, 3, 166–3182. [Google Scholar] [CrossRef] [PubMed]
- Colombo, M.; Damerau, M.; Hanel, R.; Salzburger, W.; Matschiner, M. Diversity and disparity through time in the adaptive radiation of Antarctic notothenioid fishes. J. Evol. Biol. 2015, 28, 376–394. [Google Scholar] [CrossRef] [PubMed]
- Chenuil, A.; Saucède, T.; Hemery, L.G.; Eléaume, M.; Féral, J.; Améziane, N.; David, B.; Lecointre, G.; Havermans, C. Understanding processes at the origin of species flocks with a focus on the marine Antarctic fauna. Biol. Rev. 2017, 93, 481–504. [Google Scholar] [CrossRef] [PubMed]
- Bowen, B.W.; Forsman, Z.H.; Whitney, J.L.; Faucci, A.; Hoban, M.; Canfield, S.J.; Johnston, E.C.; Coleman, R.R.; Copus, J.M.; Vicente, J.; et al. Species radiations in the sea: What the flock? J. Hered. 2020, 111, 70–83. [Google Scholar] [CrossRef] [PubMed]
- Arnason, U.; Gullberg, A.; Janke, A.; Kullberg, M.; Lehman, N.; Petrov, E.A.; Väinölä, R. Pinniped phylogeny and a new hypothesis for their origin and dispersal. Mol. Phylogenet. Evol. 2006, 41, 345–354. [Google Scholar] [CrossRef]
- Árnason, Ú.; Lammers, F.; Kumar, V.; Nilsson, M.A.; Janke, A. Whole-genome sequencing of the blue whale and other rorquals finds signatures for introgressive gene flow. Sci. Adv. 2018, 4, eaap9873. [Google Scholar] [CrossRef]
- Fulton, T.L.; Strobeck, C. Multiple fossil calibrations, nuclear loci and mitochondrial genomes provide new insight into biogeography and divergence timing for true seals (Phocidae, Pinnipedia). J. Biogeogr. 2010, 37, 814–829. [Google Scholar] [CrossRef]
- Crame, J.A. Key stages in the evolution of the Antarctic marine fauna. J. Biogeogr. 2018, 45, 986–994. [Google Scholar] [CrossRef]
- Vianna, J.A.; Fernandes, F.A.N.; Frugone, M.J.; Figueiró, H.V.; Pertierra, L.R.; Noll, D.; Bi, K.; Wang-Claypool, C.Y.; Lowther, A.; Parker, P.; et al. Genome-wide analyses reveal drivers of penguin diversification. Proc. Natl. Acad. Sci. USA 2020, 117, 22303–22310. [Google Scholar] [CrossRef] [PubMed]
- Streelman, J.T.; Alfaro, M.; Westneat, M.W.; Bellwood, D.R.; Karl, S.A. Evolutionary history of the parrotfishes: Biogeography, ecomorphology, and comparative diversity. Evolution 2002, 56, 961–971. [Google Scholar] [PubMed]
- Streelman, J.T.; Danley, P.D. The stages of vertebrate evolutionary radiation. Trends Ecol. Evol. 2003, 18, 126–131. [Google Scholar] [CrossRef]
- Gavrilets, S.; Losos, J.B.; Rowland, J.M.; Emlen, D.J. Adaptive Radiation: Contrasting Theory with Data. Science 2009, 323, 732–737. [Google Scholar] [CrossRef] [PubMed]
- Eastman, J.T.; Witmer, L.M.; Ridgely, R.C.; Kuhn, K.L. Divergence in skeletal mass and bone morphology in Antarctic notothenioid fishes. J. Morphol. 2014, 275, 841–861. [Google Scholar] [CrossRef] [PubMed]
- Eastman, J.T. Bathymetric distributions of notothenioid fishes. Polar Biol. 2017, 40, 2077–2095. [Google Scholar] [CrossRef]
- Eastman, J.T. An analysis of maximum body size and designation of size categories for notothenioid fishes. Polar Biol. 2019, 42, 1131–1145. [Google Scholar] [CrossRef]
- Eastman, J.T. The buoyancy-based biotope axis of the evolutionary radiation of Antarctic cryonotothenioid fishes. Polar Biol. 2020, 43, 1217–1231. [Google Scholar] [CrossRef]
- La Mesa, M.; Llompart, F.; Riginella, E.; Eastman, J.T. Parental care and reproductive strategies in notothenioid fishes. Fish Fish. 2020, 22, 356–376. [Google Scholar] [CrossRef]
- Eastman, J.T.; Eakin, R.R. Checklist of the species of notothenioid fishes. Antarct. Sci. 2021, 33, 273–280. [Google Scholar] [CrossRef]
- Near, T.J.; Dornburg, A.; Eytan, R.I.; Keck, B.P.; Smith, W.L.; Kuhn, K.L.; Moore, J.A.; Price, S.A.; Burbrink, F.T.; Friedman, M.; et al. Phylogeny and tempo of diversification in the superradiation of spiny-rayed fishes. Proc. Natl. Acad. Sci. USA 2013, 110, 12738–12743. [Google Scholar] [CrossRef] [PubMed]
- Sheiko, B.A. Comments on the nomenclature of genus- and family-series taxa of notothenioid fishes (Perciformes, Notothenioidei). Bionomina 2019, 16, 46–82. [Google Scholar] [CrossRef]
- Haedrich, R.L. Deep-water fishes: Evolution and adaptation in the earth’s largest living spaces*. J. Fish Biol. 1996, 49, 40–53. [Google Scholar] [CrossRef]
- Ingram, T. Speciation along a depth gradient in a marine adaptive radiation. Proc. R. Soc. B Biol. Sci. 2010, 278, 613–618. [Google Scholar] [CrossRef] [PubMed]
- Nybelin, O. Antarctic fishes. In Scientific Results Norwegian Antarctic expeditions 1927–1928 et sqq.; No. 26; I Kommisjon hos J. Dybwad: Oslo, Norway, 1947; pp. 1–76. [Google Scholar]
- Andriashev, A.P. A general review of the Antarctic fish fauna. In Biogeography and Ecology in Antarctica, Monographiae Biologicae; Van Oye, P., Van Mieghem, J., Eds.; Junk: The Hague, The Netherlands, 1965; Volume XV, pp. 491–550. [Google Scholar]
- Andriashev, A.P. A general review of the Antarctic bottom fish fauna. In Proceedings of the Fifth Congress of European Ichthyologists, Stockholm, Sweden, 12 September 1985; Kullander, S.O., Fernholm, B., Eds.; Swedish Museum of Natural History: Stockholm, Sweden, 1987; pp. 357–372. [Google Scholar]
- Linley, T.D.; Gerringer, M.E.; Yancey, P.H.; Drazen, J.C.; Weinstock, C.L.; Jamieson, A.J. Fishes of the hadal zone including new species, in situ observations and depth records of Liparidae. Deep Sea Res. Part I Oceanogr. Res. Pap. 2016, 114, 99–110. [Google Scholar] [CrossRef]
- Dewitt, H.H. Coastal and deep-water benthic fishes of the Antarctic. In Antarctic Map Folio Series, Folio 15; Bushnell, V.C., Ed.; American Geographical Society: New York, NY, USA, 1971; pp. 1–10. [Google Scholar]
- Griffiths, H.J.; Van de Putte, A.; Danis, B. Data distribution: Patterns and implications. In Biogeographic Atlas of the Southern Ocean; De Broyer, C., Koubbi, P., Griffiths, H.J., Raymond, B., Udekem D’acoz, C.D., Eds.; Scientific Committee on Antarctic Research: Cambridge, UK, 2014; pp. 16–26. [Google Scholar]
- Angel, M.V. What is the deep-sea? In Fish Physiology: Deep-Sea Fishes; Randall, D.J., Farrell, A.P., Eds.; Academic Press: San Diego, CA, USA, 1997; Volume 16, pp. 1–41. [Google Scholar]
- Priede, I.G.; Froese, R. Colonization of the deep sea by fishes. J. Fish Biol. 2013, 83, 1528–1550. [Google Scholar] [CrossRef] [PubMed]
- Eastman, J.T.; La Mesa, M. Neuromorphological disparity in deep-living sister species of the Antarctic fish genus Trematomus. Polar Biol. 2021, 44, 315–334. [Google Scholar] [CrossRef]
- Amsler, M.O.; Eastman, J.T.; Smith, K.E.; Mcclintock, J.B.; Singh, H.; Thatje, S.; Aronson, R.B. Zonation of demersal fishes off Anvers Island, western Antarctic Peninsula. Antarct. Sci. 2015, 28, 44–50. [Google Scholar] [CrossRef]
- Eastman, J.T. The evolution of neutrally buoyant notothenioid fishes: Their specializations and potential interactions in the Antarctic marine food web. In Antarctic Nutrient Cycles and Food Webs; Siegfried, W.R., Condy, P.R., Laws, R.M., Eds.; Springer: Berlin/Heidelberg, Germany, 1985; pp. 430–436. [Google Scholar]
- Eastman, J.T. Antarctic Fish Biology: Evolution in a Unique Environment; Academic Press: San Diego, CA, USA, 1993; 322p. [Google Scholar]
- Devries, A.L.; Eastman, J.T. Lipid sacs as a buoyancy adaptation in an Antarctic fish. Nature 1978, 271, 352–353. [Google Scholar] [CrossRef]
- Near, T.J.; Russo, S.E.; Jones, C.D.; DeVries, A.L. Ontogenetic shift in buoyancy and habitat in the Antarctic toothfish, Dissostichus mawsoni (Perciformes: Nototheniidae). Polar Biol. 2002, 26, 124–128. [Google Scholar] [CrossRef]
- Lincoln, R.; Boxshall, G.; Clark, P. A Dictionary of Ecology, Evolution and Systematics, 2nd ed.; Cambridge University Press: Cambridge, UK, 1998; 361p. [Google Scholar]
- Klingenberg, C.P.; Ekau, W. A combined morphometric and phylogenetic analysis of an ecomorphological trend: Pelagization in Antarctic fishes (Perciformes: Nototheniidae). Biol. J. Linn. Soc. 1996, 59, 143–177. [Google Scholar] [CrossRef]
- Eastman, J.; Barrera-Oro, E.; Moreira, E. Adaptive radiation at a low taxonomic level: Divergence in buoyancy of the ecologically similar Antarctic fish Notothenia coriiceps and N. rossii. Mar. Ecol. Prog. Ser. 2011, 438, 195–206. [Google Scholar] [CrossRef]
- Barrera-Oro, E.R.; Casaux, R.J. Feeding selectivity in Notothenia neglecta, Nybelin, from Potter Cove, South Shetland Islands, Antarctica. Antarct. Sci. 1990, 2, 207–213. [Google Scholar] [CrossRef]
- North, A.W. Locomotory activity and behaviour of the Antarctic teleost Notothenia coriiceps. Mar. Biol. 1996, 126, 125–132. [Google Scholar] [CrossRef]
- Barrera-Oro, E.; Moreira, E.; Seefeldt, M.A.; Francione, M.V.; Quartino, M.L. The importance of macroalgae and associated amphipods in the selective benthic feeding of sister rockcod species Notothenia rossii and N. coriiceps (Nototheniidae) in West Antarctica. Polar Biol. 2018, 42, 317–334. [Google Scholar] [CrossRef]
- Targett, T. Trophic Ecology and Structure of Coastal Antarctic Fish Communities. Mar. Ecol. Prog. Ser. 1981, 4, 243–263. [Google Scholar] [CrossRef]
- Dewitt, H.H.; Heemstra, P.C.; Gon, O. Nototheniidae. In Fishes of the Southern Ocean; Gon, O., Heemstra, P.C., Eds.; J.L.B. Smith Institute of Ichthyology: Grahamstown, South Africa, 1990; pp. 279–331. [Google Scholar]
- Bushula, T.; Pakhomov, E.A.; Kaehler, S.; Davis, S.; Kalin, R.M. Diet and daily ration of two nototheniid fish on the shelf of the sub-Antarctic Prince Edward Islands. Polar Biol. 2005, 28, 585–593. [Google Scholar] [CrossRef]
- Daniels, R.A. Feeding ecology of some fishes of the Antarctic Peninsula. Fish. Bull. 1982, 80, 575–588. [Google Scholar]
- Permitin, Y.E. The consumption of krill by Antarctic fishes. In Antarctic Ecology; Holdgate, M.W., Ed.; Academic Press: London, UK, 1970; Volume 1, pp. 177–182. [Google Scholar]
- Permitin, Y.Y.; Tarverdiyeva, M.I. The food of some Antarctic fish in the South Georgia area. J. Ichthyol. 1972, 12, 104–114. [Google Scholar]
- Barrera-Oro, E.; Tomo, A. Feeding and ecology of Notothenia larseni, Lönnberg. In Antarctic Aquatic Biology; BIOMASS Scientific Series No. 7; El-Sayed, S., Ed.; Cambridge University Press: Cambridge, UK, 1987; pp. 99–106. [Google Scholar]
- Takahashi, M.; Iwami, T. The summer diet of demersal fish at the South Shetland Islands. Antarct. Sci. 1997, 9, 407–413. [Google Scholar] [CrossRef]
- Shust, K.V.; Pinskaya, I.A. Age and rate of growth of six species of nototheniid fish (family Nototheniidae). J. Ichthyol. 1978, 18, 743–749. [Google Scholar]
- Gröhsler, T. Feeding habits as indicators of ecological niches: Investigations of Antarctic fish conducted near Elephant Island in late autumn/winter 1986. Arch. Fish. Mar. Res. 1994, 42, 17–34. [Google Scholar]
- Kock, K.-H. Krill consumption by Antarctic notothenioid fishes. In Antarctic Nutrient Cycles and Food Webs; Siegfried, W.R., Condy, P.R., Laws, R.M., Eds.; Springer: Berlin/Heidelberg, Germany, 1985; pp. 437–444. [Google Scholar]
- Grant, P.R.; Grant, B.R. How and Why Species Multiply: The Radiation of Darwin’s Finches; Princeton University Press: Princeton, NJ, USA, 2008; 218p. [Google Scholar]
- Losos, J.B. Lizards in an Evolutionary Tree: Ecology and Adaptive Radiation of Anoles; University of California Press: Berkeley, CA, USA, 2009; 507p. [Google Scholar]
- Wainwright, P.C.; Richard, B.A. Predicting patterns of prey use from morphology of fishes. Environ. Biol. Fishes 1995, 44, 97–113. [Google Scholar] [CrossRef]
- Knouft, J.H.; Page, L.M. The evolution of body size in extant groups of North American freshwater fishes: Speciation, size distributions, and Cope’s rule. Am. Nat. 2003, 161, 413–421. [Google Scholar] [CrossRef] [PubMed]
- Barneche, D.R.; Robertson, D.R.; White, C.R.; Marshall, D.J. Fish reproductive-energy output increases disproportionately with body size. Science 2018, 360, 642–644. [Google Scholar] [CrossRef] [PubMed]
- Ainley, D.G.; Cziko, P.A.; Nur, N.; Rotella, J.J.; Eastman, J.T.; Larue, M.; Stirling, I.; Abrams, P.A. Further evidence that Antarctic toothfish are important to Weddell seals. Antarct. Sci. 2020, 33, 17–29. [Google Scholar] [CrossRef]
- Balushkin, A.V. Morphological Bases of the Systematics and Phylogeny of the Nototheniid Fishes; Academy of Sciences of the USSR, Zoological Institute: St. Petersburg, Russia, 1984; 153p, [In Russian; English Translation for Division of Polar Programs, National Science Foundation, 1989. 153 pp. Also available as Russian Translation Series No. 73, 1990, from A.A. Balkema, Rotterdam.]. [Google Scholar]
- Kock, K.-H. Antarctic Fish and Fisheries; Cambridge University Press: Cambridge, UK, 1992; 359p. [Google Scholar]
- Moyle, P.B.; Cech, J.J., Jr. Fishes: An Introduction to Ichthyology, 5th ed.; Prentice Hall: Upper Saddle River, NJ, USA, 2004; 726p. [Google Scholar]
- Helfman, G.S.; Collette, B.B.; Facey, D.E.; Bowen, B.W. The Diversity of Fishes: Biology: Evolution, and Ecology, 2nd ed.; Wiley-Blackwell: Chichester, UK, 2009. [Google Scholar]
- Paxton, C. A Cumulative Species Description Curve for Large Open Water Marine Animals. J. Mar. Biol. Assoc. 1998, 78, 1389–1391. [Google Scholar] [CrossRef]
- Roberts, T.R. Systematics, biology, and distribution of the species of the oceanic oarfish genus Regalecus (Teleostei, Lampridiformes, Regalecidae). Mém. Mus. Natl. Hist. Natur. 2012, 202, 1–268. [Google Scholar]
- McClain, C.R.; Balk, M.A.; Benfield, M.C.; Branch, T.A.; Chen, C.; Cosgrove, J.; Dove, A.D.; Gaskins, L.; Helm, R.R.; Hochberg, F.G.; et al. Sizing ocean giants: Patterns of intraspecific size variation in marine megafauna. PeerJ 2015, 3, e715. [Google Scholar] [CrossRef] [PubMed]
- Nelson, J.S. Fishes of the World, 4th ed.; John Wiley & Sons: Hoboken, NJ, USA, 2006; 601p. [Google Scholar]
- Nelson, J.S.; Grande, T.C.; Wilson, M.V.H. Fishes of the World, 5th ed.; John Wiley & Sons: Hoboken, NJ, USA, 2016; 707p. [Google Scholar]
- McCune, A.R.; Carlson, R.L. Twenty ways to lose your bladder: Common natural mutants in zebrafish and widespread convergence of swim bladder loss among teleost fishes. Evol. Dev. 2004, 6, 246–259. [Google Scholar] [CrossRef] [PubMed]
- Meunier, F.J. Skeleton. In Manual of Fish Sclerochronology; Panfili, J., De Pontual, H., Troadec, H., Wright, P.J., Eds.; Ifremer-IRD Coedition: Brest, France, 2002; pp. 65–88. [Google Scholar]
- Zimmermann, C.; Hubold, G. Respiration and activity of Arctic and Antarctic fish with different modes of life: A multivariate analysis of experimental data. In Fishes of Antarctica: A Biological Overview; Di Prisco, G., Pisano, E., Clarke, A., Eds.; Springer: Milan, Italy, 1998; pp. 163–174. [Google Scholar]
- Davenport, J.; Kjørsvik, E. Buoyancy in the Lumpsucker Cyclopterus Lumpus. J. Mar. Biol. Assoc. 1986, 66, 159–174. [Google Scholar] [CrossRef]
- Alexander, R.M. Principles of Animal Locomotion; Princeton University Press: Princeton, NJ, USA, 2003; 371p. [Google Scholar]
- Pelster, B. Buoyancy at depth. In Deep-Sea Fishes: Fish Physiology; Randall, D.J., Farrell, A.P., Eds.; Academic Press: San Diego, CA, USA, 1997; Volume 16, pp. 195–237. [Google Scholar]
- Pelster, B. Buoyancy control in aquatic vertebrates. In Cardio-Respiratory Control in Vertebrates; Glass, M.L., Wood, S.C., Eds.; Springer: Berlin/Heidelberg, Germany, 2009; pp. 65–98. [Google Scholar]
- Griffith, R.W.; Mathews, M.B.; Umminger, B.L.; Grant, B.F.; Pang, P.K.T.; Thomson, K.S.; Pickford, G.E. Composition of fluid from the notochordal canal of the coelacanth, Latimeria chalumnae. J. Exp. Zool. 1975, 192, 165–171. [Google Scholar] [CrossRef] [PubMed]
- Griffith, R.W. Chemistry of the body fluids of the coelacanth, Latimeria chalumnae. Proc. R. Soc. Lond. Ser. B. Biol. Sci. 1980, 208, 329–347. [Google Scholar] [CrossRef]
- Odani, K.; Imamura, H.; Nakaya, K. Osteological description of the Brazilian flathead, Percophis brasiliensis (Actinopterygii: Perciformes: Percophidae), with comments on its phylogenetic position. Species Divers. 2006, 11, 277–294. [Google Scholar] [CrossRef]
- Meunier, F.J.; Lecomte, F.; Duhamel, G. Some histological data on bone and teeth in the grey notothen (Lepidonotothen squamifrons) and in the mackerel icefish (Champsocephalus gunnari) (Notothenioidei; Perciformes; Teleostei). Cybium 2018, 42, 91–97. [Google Scholar]
- Ashique, A.M.; Atake, O.J.; Ovens, K.; Guo, R.; Pratt, I.V.; Detrich, H.W.; Cooper, D.M.L.; Desvignes, T.; Postlethwait, J.H.; Eames, B.F. Bone microstructure and bone mineral density are not systemically different in Antarctic icefishes and related Antarctic notothenioids. J. Anat. 2021, 240, 34–49. [Google Scholar] [CrossRef] [PubMed]
- Schaeffer, B. Differential ossification in the fishes. Trans. N. Y. Acad. Sci. 1961, 23, 501–505. [Google Scholar] [CrossRef]
- Gosline, W.A. Functional Morphology and Classification of Teleostean Fishes; University Press of Hawaii: Honolulu, HI, USA, 1971; 208p. [Google Scholar]
- Witten, P.E.; Hall, B.K. Teleost Skeletal Plasticity: Modulation, Adaptation, and Remodelling. Copeia 2015, 103, 727–739. [Google Scholar] [CrossRef]
- Loeb, V.J.; Kellermann, A.K.; Koubbi, P.; North, A.W.; White, M.G. Antarctic larval fish assemblages: A review. Bull. Mar. Sci. 1993, 53, 416–449. [Google Scholar]
- Andrew, T.G.; Hecht, T.; Heemstra, P.C.; Lutjeharms, J.R.E. Fishes of the Tristan da Cunha Group and Gough Island, South Atlantic Ocean. Ichthyol. Bull. JLB Smith Inst. Ichthyol. 1995, 63, 1–41. [Google Scholar]
- Rodrigues, K.A.; Jaureguizar, A.J.; Guerrero, R.A. Environmental factors that define the spawning and nursery areas for Percophis brasiliensis (Teleostei: Percophididae) in a multispecific reproductive coastal zone, El Rincón (39°–41°S), Argentina. Hydrobiologia 2013, 709, 1–10. [Google Scholar] [CrossRef]
- Voskoboinikova, O.S. Evolutionary significance of heterochronies in the development of the bony skeleton in fishes of the suborder Notothenioidei (Perciformes). J. Ichthyol. 2001, 41, 415–424. [Google Scholar]
- Voskoboinikova, O.S. The growth rate of skeleton in ontogeny of the Antarctic fish from the suborder Notothenioidae (Perciformes, Pisces) and the problem of cold compensation. Doklady Biol. Sci. 2007, 415, 307–309. [Google Scholar] [CrossRef] [PubMed]
- Voskoboinikova, O.S. Ontogenetic Bases of the Notothenioid Evolution. Explorations of the Fauna of the Seas; NAUKA: St. Petersburg, Russia, 2010; Volume 64, 319p. (In Russian) [Google Scholar]
- Voskoboinikova, O.S.; Detrich, H.W., III; Albertson, R.C.; Postlethwait, J.H.; Ghigliotti, L.; Pisano, E. Evolution reshaped life for the water column: The skeleton of the Antarctic silverfish Pleuragrama antarctica Boulenger, 1902. In The Antarctic Silverfish: A Keystone Species in a Changing Ecosystem, Advances in Polar Ecology; Vacchi, M., Pisano, E., Ghigliotti, L., Eds.; Springer International: Cham, Switzerland, 2017; Volume 3, pp. 3–26. [Google Scholar]
- Albertson, R.C.; Yan, Y.-L.; Titus, T.A.; Pisano, E.; Vacchi, M.; Yelick, P.C.; Detrich, H.W.; Postlethwait, J.H. Molecular pedomorphism underlies craniofacial skeletal evolution in Antarctic notothenioid fishes. BMC Evol. Biol. 2010, 10, 4. [Google Scholar] [CrossRef] [PubMed]
- Postlethwait, J.H.; Yan, Y.; Desvignes, T.; Allard, C.; Titus, T.; Le François, N.R.; Detrich, H.W. Embryogenesis and early skeletogenesis in the antarctic bullhead notothen, Notothenia coriiceps. Dev. Dyn. 2016, 245, 1066–1080. [Google Scholar] [CrossRef] [PubMed]
- Eastman, J.T.; DeVries, A.L. Buoyancy adaptations in a swim-bladderless Antarctic fish. J. Morphol. 1981, 167, 91–102. [Google Scholar] [CrossRef] [PubMed]
- Eastman, J.T.; DeVries, A.L. Buoyancy Studies of Notothenioid Fishes in McMurdo Sound, Antarctica. Copeia 1982, 1982, 385. [Google Scholar] [CrossRef]
- Eastman, J.T.; DeVries, A.L. Ultrastructure of the lipid sac wall in the Antarctic notothenioid fish Pleuragramma antarcticum. Polar Biol. 1989, 9, 333–335. [Google Scholar] [CrossRef]
- Chen, L.; Lu, Y.; Li, W.; Ren, Y.; Yu, M.; Jiang, S.; Fu, Y.; Wang, J.; Peng, S.; Bilyk, K.T.; et al. The genomic basis for colonizing the freezing Southern Ocean revealed by Antarctic toothfish and Patagonian robalo genomes. GigaScience 2019, 8, giz016. [Google Scholar] [CrossRef] [PubMed]
- Wainwright, S.A.; Biggs, W.D.; Currey, J.D.; Gosline, J.M. Mechanical Design in Organisms; Princeton University Press: Princeton, NJ, USA, 1976; 423p. [Google Scholar]
- Pelster, B. Buoyancy. In The Physiology of Fishes, 2nd ed.; Evans, D.H., Ed.; CRC Press: Boca Raton, FL, USA, 1998; pp. 25–42. [Google Scholar]
- Voskoboinikova, O.S. Osteological development of the Channichthyidae (Teleostei: Notothenioidei). Cybium 1997, 21, 369–379. [Google Scholar]
- Arratia, G.; Schultze, H.; Casciotta, J. Vertebral column and associated elements in dipnoans and comparison with other fishes: Development and homology. J. Morphol. 2001, 250, 101–172. [Google Scholar] [CrossRef] [PubMed]
- Bensimon-Brito, A.; Cardeira, J.; Cancela, M.L.; Huysseune, A.; Witten, P.E. Distinct patterns of notochord mineralization in zebrafish coincide with the localization of Osteocalcin isoform 1 during early vertebral centra formation. BMC Dev. Biol. 2012, 12, 28. [Google Scholar] [CrossRef] [PubMed]
- Clarke, A.; Doherty, N.; DeVries, A.L.; Eastman, J.T. Lipid content and composition of three species of Antarctic fish in relation to buoyancy. Polar Biol. 1984, 3, 77–83. [Google Scholar] [CrossRef]
- Eastman, J.T. Lipid storage systems and the biology of two neutrally buoyant Antarctic notothenioid fishes. Comp. Biochem. Physiol. Part B Comp. Biochem. 1988, 90, 529–537. [Google Scholar] [CrossRef]
- Eastman, J.T. Phyletic divergence and specialization for pelagic life in the Antarctic nototheniid fish Pleuragramma antarcticum. Comp. Biochem. Physiol. 1997, 118A, 1095–1101. [Google Scholar] [CrossRef]
- Fenaughty, J.M.; Eastman, J.T.; Sidell, B.D. Biological implications of low condition factor “axe handle” specimens of the Antarctic toothfish, Dissostichus mawsoni, from the Ross Sea. Antarct. Sci. 2008, 20, 537–551. [Google Scholar] [CrossRef]
- La Mesa, M.; Eastman, J.T. Antarctic silverfish: Life strategies of a key species in the high-Antarctic ecosystem. Fish Fish. 2011, 13, 241–266. [Google Scholar] [CrossRef]
- Bock, C.; Wermter, F.C.; Mintenbeck, K. MRI and MRS on preserved samples as a tool in fish ecology. Magn. Reson. Imaging 2017, 38, 39–46. [Google Scholar] [CrossRef]
- Hagen, W.; Kattner, G. The role of lipids in the life history of the Antarctic silverfish Pleuragramma antarctica. In The Antarctic Silverfish: A Keystone Species in a Changing Ecosystem, Advances in Polar Ecology; Vacchi, M., Pisano, E., Ghigliotti, L., Eds.; Springer International: Cham, Switzerland, 2017; Volume 3, pp. 131–148. [Google Scholar]
- Fenaughty, J.M.; Stevens, D.W.; Hanchet, S.M. Diet of the Antarctic toothfish (Dissostichus mawsoni) from the Ross Sea, Antarctica (Subarea 88.1). CCAMLR Sci. 2003, 10, 113–123. [Google Scholar]
- Petrov, A.F.; Tatarnikov, V.A. Results of investigation of the diet of Antarctic toothfish Dissostichus mawsoni (Nototheniidae) in the Lazarev Sea. J. Ichthyol. 2011, 51, 131–135. [Google Scholar] [CrossRef]
- Roberts, J.; Xavier, J.C.; Agnew, D.J. The diet of toothfish species Dissostichus eleginoides and Dissostichus mawsoni with overlapping distributions. J. Fish Biol. 2011, 79, 138–154. [Google Scholar] [CrossRef] [PubMed]
- Maes, J.; Van de Putte, A.; Hecq, J.H.; Volckaert, F.A.M. State-dependent energy allocation in the pelagic Antarctic silverfish Pleuragramma antarcticum: Trade-off between winter reserves and buoyancy. Mar. Ecol. Prog. Ser. 2006, 326, 269–282. [Google Scholar] [CrossRef]
- Barton, M. Bond’s Biology of Fishes, 3rd ed.; Thomson Brooks/Cole: Belmont, CA, USA, 2007; 891p. [Google Scholar]
- Friedman, S.T.; Price, S.A.; Corn, K.A.; Larouche, O.; Martinez, C.M.; Wainwright, P.C. Body shape diversification along the benthic-pelagic axis in marine fishes: Effects of habitat on fish body shape. Proc. Biol. Sci. 2020, 287, 20201053. [Google Scholar] [CrossRef]
- Price, S.A.; Friedman, S.T.; Corn, K.A.; Martinez, C.M.; Larouche, O.; Wainwright, P.C. Building a Body Shape Morphospace of Teleostean Fishes. Integr. Comp. Biol. 2019, 59, 716–730. [Google Scholar] [CrossRef] [PubMed]
- Macdonald, J.A.; Wells, R.M.G. Viscosity of body fluids from Antarctic notothenioid fish. In Biology of Antarctic Fish; di Prisco, G., Maresca, B., Tota, B., Eds.; Springer: Berlin/Heidelberg, Germany, 1991; pp. 163–178. [Google Scholar]
- Neat, F.C.; Campbell, N. Proliferation of elongate fishes in the deep sea. J. Fish Biol. 2013, 83, 1576–1591. [Google Scholar] [CrossRef] [PubMed]
- Brey, T.; Gerdes, D. Is Antarctic benthic biomass really higher than elsewhere? Antarct. Sci. 1997, 9, 266–267. [Google Scholar] [CrossRef]
- Clarke, A.; Johnston, N.M. Antarctic marine benthic diversity. Oceanogr. Mar. Biol. Ann. Rev. 2003, 41, 47–114. [Google Scholar]
- Mincks, S.L.; Smith, C.R.; DeMaster, D.J. Persistence of labile organic matter and microbial biomass in Antarctic shelf sediments: Evidence of a sediment ‘food bank’. Mar. Ecol. Prog. Ser. 2005, 300, 3–19. [Google Scholar] [CrossRef]
- Smith, C.R.; Mincks, S.; DeMaster, D.J. A synthesis of bentho-pelagic coupling on the Antarctic shelf: Food banks, ecosystem inertia and global climate change. Deep. Sea Res. Part II Top. Stud. Oceanogr. 2006, 53, 875–894. [Google Scholar] [CrossRef]
- Targett, T.E.; Young, K.E.; Konecki, J.T.; Grecay, P. Research on wintertime feeding in Antarctic fishes. Antarct. J. US 1987, 22, 211–213. [Google Scholar]
- Obermüller, B.E.; Truebano, M.; Peck, L.S.; Eastman, J.T.; Morley, S.A. Reduced seasonality in elemental CHN composition of Antarctic marine benthic predators and scavengers. J. Exp. Mar. Biol. Ecol. 2013, 446, 328–333. [Google Scholar] [CrossRef]
- Casaux, R.J.; Mazzotta, A.S.; Barrera-Oro, E.R. Seasonal aspects of the biology and diet of nearshore nototheniid fish at Potter Cove, South Shetland Islands, Antarctica. Polar Biol. 1990, 11, 63–72. [Google Scholar] [CrossRef]
- Casaux, R.; Barrera-Oro, E.; Baroni, A.; Ramón, A. Ecology of inshore notothenioid fish from the Danco Coast, Antarctic Peninsula. Polar Biol. 2003, 26, 157–165. [Google Scholar] [CrossRef]
- McKenna, J.E., Jr. Trophic relationships within the Antarctic demersal fish community of South Georgia Island. Fish. Bull. 1991, 89, 643–654. [Google Scholar]
- Kiest, K.A. A relationship of diet to prey abundance and the foraging behavior of Trematomus bernacchii. Polar Biol. 1993, 13, 291–296. [Google Scholar] [CrossRef]
- Montgomery, J.C.; Foster, B.A.; Milton, R.C.; Carr, E. Spatial and temporal variations in the diet of nototheniid fish in McMurdo Sound, Antarctica. Polar Biol. 1993, 13, 429–431. [Google Scholar] [CrossRef]
- Vacchi, M.; La Mesa, M.; Castelli, A. Diet of two coastal nototheniid fish from Terra Nova Bay, Ross Sea. Antarct. Sci. 1994, 6, 61–65. [Google Scholar] [CrossRef]
- Vacchi, M.; Cattaneo-Vietti, R.; Chiantore, M.; Dalu, M. Predator-prey relationship between the nototheniid fish Trematomus bernacchii and the Antarctic scallop Adamussium colbecki at Terra Nova Bay (Ross Sea). Antarct. Sci. 2000, 12, 64–68. [Google Scholar] [CrossRef]
- Barrera-Oro, E. Analysis of dietary overlap in Antarctic fish (Notothenioidei) from the South Shetland Islands: No evidence of food competition. Polar Biol. 2003, 26, 631–637. [Google Scholar] [CrossRef]
- La Mesa, M.; Dalú, M.; Vacchi, M. Trophic ecology of the emerald notothen Trematomus bernacchii (Pisces, Nototheniidae) from Terra Nova Bay, Ross Sea, Antarctica. Polar Biol. 2004, 27, 721–728. [Google Scholar] [CrossRef]
- La Mesa, M.; Eastman, J.T.; Vacchi, M. The role of notothenioid fish in the food web of the Ross Sea shelf waters: A review. Polar Biol. 2004, 27, 321–338. [Google Scholar] [CrossRef]
- Brey, T. Population Dynamics in Benthic Invertebrates. A Virtual Handbook; Alfred Wegener Institute for Polar and Marine Research: Bremerhaven, Germany, 2001; Available online: http://www.awi-bremerhaven.de/Benthic/Ecosystem/FoodWeb/Handbook/main.html (accessed on 1 January 2024).
- Barrera-Oro, E. The role of fish in the Antarctic marine food web: Differences between inshore and offshore waters in the southern Scotia Arc and west Antarctic Peninsula. Antarct. Sci. 2002, 14, 293–309. [Google Scholar] [CrossRef]
- Foster, B.A.; Montgomery, J.C. Planktivory in benthic nototheniid fish in McMurdo Sound, Antarctica. Environ. Biol. Fish. 1993, 36, 313–318. [Google Scholar] [CrossRef]
- Pakhomov, E.A. Feeding and exploitation of the food supply by demersal fishes in the Antarctic part of the Indian Ocean. J. Ichthyol. 1997, 37, 360–380. [Google Scholar]
- Stowasser, G.; Pond, D.W.; Collins, M.A. Fatty acid trophic markers elucidate resource partitioning within the demersal fish community of South Georgia and Shag Rocks (Southern Ocean). Mar. Biol. 2012, 159, 2299–2310. [Google Scholar] [CrossRef]
- Casaux, R.; Barrera-Oro, E. Dietary overlap in inshore notothenioid fish from the Danco Coast, western Antarctic Peninsula. Polar Res. 2013, 32, 21319. [Google Scholar] [CrossRef]
- Eastman, J.T. Pleuragramma antarcticum (Pisces, Nototheniidae) as food for other fishes in McMurdo Sound, Antarctica. Polar Biol. 1985, 4, 155–160. [Google Scholar] [CrossRef]
- Friedrich, C.; Hagen, W. Lipid contents of five species of notothenioid fish from high-Antarctic waters and ecological implications. Polar Biol. 1994, 14, 359–369. [Google Scholar] [CrossRef]
- VNIRO. Chemical Composition and Processing Properties of Marine and Ocean Fishes: Handbook-M; VNIRO: Moscow, Russia, 2000; 376p. [Google Scholar]
- Liem, K.F. Adaptive Significance of Intra- and Interspecific Differences in the Feeding Repertoires of Cichlid Fishes. Am. Zool. 1980, 20, 295–314. [Google Scholar] [CrossRef]
- Lauder, G.V. Patterns of Evolution in the Feeding Mechanism of Actinopterygian Fishes. Am. Zool. 1982, 22, 275–285. [Google Scholar] [CrossRef]
- Liem, K.F. Ecomorphology of the teleostean skull. In The Skull, Vol. 3, Functional and Evolutionary Mechanisms; Hanken, J., Hall, B.K., Eds.; University of Chicago Press: Chicago, IL, USA, 1993; pp. 422–452. [Google Scholar]
- Westneat, M.W. Skull biomechanics and suction feeding in fishes. In Fish Physiology: Fish Biomechanics; Shadwick, R.E., Lauder, G.V., Eds.; Academic Press: San Diego, CA, USA, 2006; Volume 23, pp. 29–75. [Google Scholar]
- Norton, S.F.; Brainerd, E.L. Convergence in the Feeding Mechanics of Ecomorphologically Similar Species in the Centrarchidae and Cichlidae. J. Exp. Biol. 1993, 176, 11–29. [Google Scholar] [CrossRef]
- Bansode, M.A.; Eastman, J.T.; Aronson, R.B. Feeding biomechanics of five demersal Antarctic fishes. Polar Biol. 2014, 37, 1835–1848. [Google Scholar] [CrossRef]
- Carlig, E.; Di Blasi, D.; Ghigliotti, L.; Pisano, E.; Faimali, M.; O’driscoll, R.; Parker, S.; Vacchi, M. Diversification of feeding structures in three adult Antarctic nototheniid fish. Polar Biol. 2018, 41, 1701–1715. [Google Scholar] [CrossRef]
- Iwami, T. Osteology and relationships of the family Channichthyidae. Mem. Natl. Inst. Polar Res. Ser. E 1985, 36, 1–69. [Google Scholar]
- Pequeño, G. The geographical distribution and taxonomic arrangement of South American nototheniid fishes (Osteichthyes, Nototheniidae). Bol. Soc. Biol. Concepción 1989, 60, 183–200. [Google Scholar]
- Licandeo, R.R.; Barrientos, C.A.; González, M.T. Age, growth rates, sex change and feeding habits of notothenioid fish Eleginops maclovinus from the central-southern Chilean coast. Env. Biol. Fish. 2006, 77, 51–61. [Google Scholar] [CrossRef]
- Brickle, P.; Arkhipkin, A.I.; Shcherbich, Z.N. age and growth in a temperate euryhaline notothenioid, Eleginops maclovinus from the Falkland islands. J. Mar. Biol. Assoc. 2005, 85, 1217–1221. [Google Scholar] [CrossRef]
- Brickle, P.; Laptikhovsky, V.; Arkhipkin, A. Reproductive strategy of a primitive temperate notothenioid Eleginops maclovinus. J. Fish Biol. 2005, 66, 1044–1059. [Google Scholar] [CrossRef]
- Pavés, H.; Pequeño, G.; Bertrán, C.; Vargas, L. Limnetic feeding in Eleginops maclovinus (Valenciennes, 1930) in the Valdivia River, Chile. Interciencia 2005, 30, 120–125. [Google Scholar]
- Troccoli, G.H.; Aguilar, E.; Martínez, P.A.; Belleggia, M. The diet of the Patagonian toothfish Dissostichus eleginoides, a deep-sea top predator off Southwest Atlantic Ocean. Polar Biol. 2020, 43, 1595–1604. [Google Scholar] [CrossRef]
- Collins, M.A.; Brickle, P.; Brown, J.; Belchier, M. The Patagonian toothfish: Biology, ecology and fishery. In Advances in Marine Biology; Lesser, M., Ed.; Academic Press: San Diego, CA, USA, 2010; Volume 58, pp. 227–300. [Google Scholar]
- Gregory, W.K. Fish Skulls: A Study of the Evolution of Natural Mechanisms. Trans. Am. Philos. Soc. 1933, 23, 75–481. [Google Scholar] [CrossRef]
- Voskoboinikova, O.S. Evolution of the visceral skeleton and phylogeny of Nototheniidae. J. Ichthyol. 1993, 33, 23–47. [Google Scholar]
- Peyer, B. Comparative Odontology; Zangerl, R., Ed.; University of Chicago Press: Chicago, IL, USA, 1968; 347p. [Google Scholar]
- Fink, W.L. Ontogeny and phylogeny of tooth attachment mode in actinopterygian fishes. J. Morphol. 1981, 167, 167–184. [Google Scholar] [CrossRef] [PubMed]
- Cohen, K.E.; Weller, H.I.; Summers, A.P. Not your father’s homodonty—Stress, tooth shape, and the functional homodont. J. Anat. 2020, 237, 837–848. [Google Scholar] [CrossRef] [PubMed]
- Andriashev, A.P.; Balushkin, A.V.; Voskoboinikova, O.S. Morphological validation of the subfamily Gymnodraconinae of the family Bathydraconidae. J. Ichthyol. 1989, 29, 147–156. [Google Scholar]
- Voskoboinikova, O.S. Comparative osteology of dragonfishes of the subfamily Gymnodraconinae (Bathydraconidae). J. Ichthyol. 1991, 31, 24–33. [Google Scholar]
- Balushkin, A.V.; Voskoboinikova, O.S. Antarctic Dragonfishes (Bathydraconidae). Explorations of the Fauna of the Seas; NAUKA: St. Petersburg, Russia, 2011; Volume 65, 222p. (In Russian) [Google Scholar]
- Ojeda, F.P. Morphological characterization of the alimentary tract of Antarctic fishes and its relation to feeding habits. Polar Biol. 1986, 5, 125–128. [Google Scholar] [CrossRef]
- Eastman, J.T.; DeVries, A.L. Morphology of the digestive system of Antarctic nototheniid fishes. Polar Biol. 1997, 17, 1–13. [Google Scholar] [CrossRef]
- Eastman, J.T. Aspects of the biology of the icefish Dacodraco hunteri (Notothenioidei, Channichthyidae) in the Ross Sea, Antarctica. Polar Biol. 1999, 21, 194–196. [Google Scholar] [CrossRef]
- Moreira, E.; Novillo, M.; Eastman, J.T.; Barrera-Oro, E. Degree of herbivory and intestinal morphology in nine notothenioid fishes from the western Antarctic Peninsula. Polar Biol. 2020, 43, 535–544. [Google Scholar] [CrossRef]
- Barrington, E.J.W. The alimentary canal and digestion. In The Physiology of Fishes; Brown, M.E., Ed.; Academic Press: New York, NY, USA, 1957; Volume 1, pp. 109–161. [Google Scholar]
- Eastman, J.T.; Amsler, M.O.; Aronson, R.B.; Thatje, S.; McClintock, J.B.; Vos, S.C.; Kaeli, J.W.; Singh, H.; La Mesa, M. Photographic survey of benthos provides insights into the Antarctic fish fauna from the Marguerite Bay slope and the Amundsen Sea. Antarct. Sci. 2012, 25, 31–43. [Google Scholar] [CrossRef]
- Voronina, E.P.; Neelov, A.V. Structural traits of alimentary tract of fishes of the family Channichthyidae (Notothenioidei). J. Ichthyol. 2001, 41, 778–788. [Google Scholar]
- Iwami, T.; Kock, K.-H. Channichthyidae. In Fishes of the Southern Ocean; Gon, O., Heemstra, P.C., Eds.; J.L.B. Smith Institute of Ichthyology: Grahamstown, South Africa, 1990; pp. 381–399. [Google Scholar]
- Duhamel, G.; Gasco, N.; Davaine, P. Poissons des îles Kerguelen et Crozet. Guide Régional de L’océan Austral. Patrimoines Naturels; Muséum National d’Histoire Naturelle: Paris, France, 2005; Volume 63, 419p. (In French) [Google Scholar]
- Kock, K.-H. Antarctic icefishes (Channichthyidae): A unique family of fishes. A review, Part I. Polar Biol. 2005, 28, 862–895. [Google Scholar] [CrossRef]
- Reid, W.D.K.; Clarke, S.; Collins, M.A.; Belchier, M. Distribution and ecology of Chaenocephalus aceratus (Channichthyidae) around South Georgia and Shag Rocks (Southern Ocean). Polar Biol. 2007, 30, 1523–1533. [Google Scholar] [CrossRef]
- Kock, K.-H.; Gröger, J.; Jones, C.D. Interannual variability in the feeding of ice fish (Notothenioidei, Channichthyidae) in the southern Scotia Arc and the Antarctic Peninsula region (CCAMLR subareas 48.1 and 48.2). Polar Biol. 2013, 36, 1451–1462. [Google Scholar] [CrossRef]
- La Mesa, M.; Vacchi, M.; Iwami, T.; Eastman, J.T. Taxonomic studies of the Antarctic icefish genus Cryodraco Dollo, 1900 (Notothenioidei: Channichthyidae). Polar Biol. 2002, 25, 384–390. [Google Scholar] [CrossRef]
- La Mesa, M.; Vacchi, M. Morphometric analysis of Cryodraco specimens (Notothenioidei: Channichthyidae) from Terra Nova Bay, Ross Sea. Cybium 1997, 21, 363–368. [Google Scholar]
- Ekau, W.; Gutt, J. Notothenioid fishes from the Weddell Sea and their habitat, observed by underwater photography and television. Proc. NIPR Symp. Polar Biol. 1991, 4, 36–49. [Google Scholar]
- Kock, K.-H.; Jones, C.D. The biology of the icefish Cryodraco antarcticus Dollo, 1900 (Pisces, Channichthyidae) in the southern Scotia Arc (Antarctica). Polar Biol. 2002, 25, 416–424. [Google Scholar] [CrossRef]
- Chen, W.-J.; Bonillo, C.; Lecointre, G. Phylogeny of the Channichthyidae (Notothenioidei, Teleostei) based on two mitochondrial genes. In Fishes of Antarctica: A Biological Overview; Di Prisco, G., Pisano, E., Clarke, A., Eds.; Springer: Milan, Italy, 1998; pp. 287–298. [Google Scholar]
- Abe, T.; Suzuki, M. Notes on some fishes associated with the Antarctic krill I. Neopagetopsis ionah. Antarct. Rec. Natl. Inst. Polar Res. 1978, 62, 23–28. [Google Scholar]
- Abe, T.; Suzuki, M. Notes on some fishes associated with the Antarctic krill. II. On Xenocyttus nemotoi Abe, and again on Neopagetopsis ionah Nybelin. Antarct. Rec. Natl. Inst. Polar Res. 1981, 71, 121–129. [Google Scholar]
- O’Driscoll, R.L.; Macaulay, G.J.; Gauthier, S.; Pinkerton, M.; Hanchet, S. Distribution, abundance and acoustic properties of Antarctic silverfish (Pleuragramma antarcticum) in the Ross Sea. Deep Sea Res. Part II Top. Stud. Oceanogr. 2011, 58, 181–195. [Google Scholar] [CrossRef]
- Balushkin, A.V.; Prut’ko, V.G. On occurrences of Chionobathyscus dewitti (Notothenioidei Channichthyidae) in the Ross Sea. J. Ichthyol. 2006, 46, 271–273. [Google Scholar] [CrossRef]
- Sutton, C.P.; Manning, M.J.; Stevens, D.W.; Marriott, P.M. Biological parameters for icefish (Chionobathyscus dewitti) in the Ross Sea, Antarctica. CCAMLR Sci. 2008, 15, 139–165. [Google Scholar]
- Petrov, A.F. New data on the diet of deep-sea icefish Chionobathyscus dewitti (Channichthyidae) in the Ross Sea in 2010. J. Ichthyol. 2011, 51, 692–694. [Google Scholar] [CrossRef]
- Gutt, J.; Ekau, W. Habitat partitioning of dominant high Antarctic demersal fish in the Weddell Sea and Lazarev Sea. J. Exp. Mar. Biol. Ecol. 1996, 206, 25–37. [Google Scholar] [CrossRef]
- Gon, O. Bathydraconidae. In Fishes of the Southern Ocean; Gon, O., Heemstra, P.C., Eds.; J.L.B. Smith Institute of Ichthyology: Grahamstown, South Africa, 1990; pp. 364–380. [Google Scholar]
- Kuhn, K.L.; Near, T.J.; Detrich, H.W., III; Eastman, J.T. Biology of the Antarctic dragonfish Vomeridens infuscipinnis (Notothenioidei: Bathydraconidae). Antarct. Sci. 2011, 23, 18–26. [Google Scholar] [CrossRef]
- Huber, R.; Rylander, M.K. Brain morphology and turbidity preference in Notropis and related genera (Cyprinidae, Teleostei). Environ. Biol. Fishes 1992, 33, 153–165. [Google Scholar] [CrossRef]
- Kotrschal, K.; Palzenberger, M. Neuroecology of cyprinids: Comparative, quantitative histology reveals diverse brain patterns. Environ. Biol. Fishes 1992, 33, 135–152. [Google Scholar] [CrossRef]
- Wagner, H.-J. Brain Areas in Abyssal Demersal Fishes. Brain, Behav. Evol. 2001, 57, 301–316. [Google Scholar] [CrossRef] [PubMed]
- Wagner, H.-J. Volumetric analysis of brain areas indicates a shift in sensory orientation during development in the deep-sea grenadier Coryphaenoides armatus. Mar. Biol. 2003, 142, 791–797. [Google Scholar] [CrossRef]
- Deng, X.; Wagner, H.; Popper, A.N. Interspecific Variations of Inner Ear Structure in the Deep-Sea Fish Family Melamphaidae. Anat. Rec. 2013, 296, 1064–1082. [Google Scholar] [CrossRef] [PubMed]
- Munk, O. The eyes of three benthic deep-sea fishes caught at great depths. Galathea Rep. 1964, 7, 137–149. [Google Scholar]
- Stein, D.L. Description of a new hadal Notoliparis from the Kermadec Trench, New Zealand, and redescription of Notoliparis kermadecensis (Nielsen) (Liparidae, Scorpaeniformes). Copeia 2016, 2016, 907–920. [Google Scholar] [CrossRef]
- Fine, M.L.; Horn, M.H.; Cox, B. Acanthonus armatus, a deep-sea teleost with a minute brain and large ears. Proc. R. Soc. Lond. 1987, 230B, 257–265. [Google Scholar]
- Country, M.W. Retinal metabolism: A comparative look at energetics in the retina. Brain Res. 2017, 1672, 50–57. [Google Scholar] [CrossRef] [PubMed]
- Iglesias, T.L.; Dornburg, A.; Brandley, M.C.; Alfaro, M.E.; Warren, D.L. Life in the unthinking depths: Energetic constraints on encephalization in marine fishes. J. Evol. Biol. 2015, 28, 1080–1090. [Google Scholar] [CrossRef]
- Montgomery, J.C.; MacDonald, J.A. Sensory Tuning of Lateral Line Receptors in Antarctic Fish to the Movements of Planktonic Prey. Science 1987, 235, 195–196. [Google Scholar] [CrossRef]
- Eastman, J.T. Ocular morphology in antarctic notothenioid fishes. J. Morphol. 1988, 196, 283–306. [Google Scholar] [CrossRef] [PubMed]
- Montgomery, J.C.; Macdonald, J.A.; Housley, G.D. Lateral line function in an antarctic fish related to the signals produced by planktonic prey. J. Comp. Physiol. A 1988, 163, 827–833. [Google Scholar] [CrossRef]
- Montgomery, J.; Coombs, S.; Janssen, J. Form and Function Relationships in Lateral Line Systems: Comparative Data from Six Species of Antarctic Notothenioid Fish. Brain, Behav. Evol. 1994, 44, 299–306. [Google Scholar] [CrossRef] [PubMed]
- Pankhurst, N.W.; Montgomery, J.C. Visual Function in Four Antarctic Nototheniid Fishes. J. Exp. Biol. 1989, 142, 311–324. [Google Scholar] [CrossRef]
- Pankhurst, N.W.; Montgomery, J.C. Ontogeny of vision in the Antarctic fish Pagothenia borchgrevinki (Nototheniidae). Polar Biol. 1990, 10, 419–422. [Google Scholar] [CrossRef]
- Janssen, J.; Jones, W.; Slattery, M. Locomotion and feeding responses to mechanical stimuli in Histiodraco velifer (Artedidracondiae). Copeia 1993, 1993, 885–889. [Google Scholar] [CrossRef]
- Eastman, J.T.; Lannoo, M.J. Diversification of brain morphology in antarctic notothenioid fishes: Basic descriptions and ecological considerations. J. Morphol. 1995, 223, 47–83. [Google Scholar] [CrossRef] [PubMed]
- Eastman, J.T.; Lannoo, M.J. Anatomy and histology of the brain and sense organs of the Antarctic plunderfish Dolloidraco longedorsalis (Perciformes: Notothenioidei: Artedidraconidae), with comments on the brain morphology of other artedidraconids and closely related harpagiferids. J. Morphol. 2003, 255, 358–377. [Google Scholar] [CrossRef] [PubMed]
- Eastman, J.T.; Lannoo, M.J. Diversification of brain and sense organ morphology in Antarctic dragonfishes (Perciformes: Notothenioidei: Bathydraconidae). J. Morphol. 2003, 258, 130–150. [Google Scholar] [CrossRef]
- Eastman, J.T.; Lannoo, M.J. Brain and sense organ anatomy and histology in hemoglobinless Antarctic icefishes (Perciformes: Notothenioidei: Channichthyidae). J. Morphol. 2004, 260, 117–140. [Google Scholar] [CrossRef]
- Eastman, J.T.; Lannoo, M.J. Brain and sense organ anatomy and histology of two species of phyletically basal non-Antarctic thornfishes of the Antarctic suborder Notothenioidei (Perciformes: Bovichtidae). J. Morphol. 2007, 268, 485–503. [Google Scholar] [CrossRef] [PubMed]
- Eastman, J.T.; Lannoo, M.J. Brain and sense organ anatomy and histology of the Falkland Islands mullet, Eleginops maclovinus (Eleginopidae), the sister group of the Antarctic notothenioid fishes (Perciformes: Notothenioidei). J. Morphol. 2007, 269, 84–103. [Google Scholar] [CrossRef] [PubMed]
- Eastman, J.T.; Lannoo, M.J. Divergence of brain and retinal anatomy and histology in pelagic Antarctic notothenioid fishes of the sister taxa Dissostichus and Pleuragramma. J. Morphol. 2011, 272, 419–441. [Google Scholar] [CrossRef] [PubMed]
- Janssen, J. Use of the lateral line and tactile senses in feeding in four antarctic nototheniid fishes. Environ. Biol. Fishes 1996, 47, 51–64. [Google Scholar] [CrossRef]
- Montgomery, J.C. An ontogenetic shift in the use of visual and non-visual senses in Antarctic notothenioid fishes. In Antarctic Communities: Species, Structure and Survival; Battaglia, B., Valencia, J., Walton, D.W.H., Eds.; Cambridge University Press: Cambridge, UK, 1997; pp. 217–220. [Google Scholar]
- Lannoo, M.J.; Eastman, J.T. Nervous and sensory system correlates of an epibenthic evolutionary radiation in Antarctic notothenioid fishes, genus Trematomus (Perciformes; Nototheniidae). J. Morphol. 2000, 245, 67–79. [Google Scholar] [CrossRef] [PubMed]
- Wujcik, J.M.; Wang, G.; Eastman, J.T.; Sidell, B.D. Morphometry of retinal vasculature in Antarctic fishes is dependent upon the level of hemoglobin in circulation. J. Exp. Biol. 2007, 210, 815–824. [Google Scholar] [CrossRef] [PubMed]
- Bauchot, R.; Ridet, J.-M.; Bauchot, M.-L. The brain organization of butterflyfishes. Environ. Biol. Fishes 1989, 25, 205–219. [Google Scholar] [CrossRef]
- Huber, R.; Van Staaden, M.J.; Kaufman, L.S.; Liem, K.F. Microhabitat use, trophic patterns, and the evolution of brain structure in African cichlids. Brain Behav. Evol. 1997, 50, 167–182. [Google Scholar] [CrossRef] [PubMed]
- Montgomery, J.; Clements, K. Disaptation and recovery in the evolution of Antarctic fishes. Trends Ecol. Evol. 2000, 15, 267–271. [Google Scholar] [CrossRef]
- Eakin, R.R.; Eastman, J.T. Artedidraconidae. In. Fishes of the Southern Ocean, 2nd ed.; Gon, O., Ed.; South African Institute of Aquatic Biodiversity: Grahamstown, South Africa, 2023; in press. [Google Scholar]
- Coombs, S.; Janssen, J.; Webb, J.F. Diversity of lateral line systems: Evolutionary and functional considerations. In Sensory Biology of Aquatic Animals; Atema, J., Fay, R.R., Popper, A.N., Tavolga, W.N., Eds.; Springer: New York, NY, USA, 1988; pp. 553–593. [Google Scholar]
- Montgomery, J.; Pankhurst, N. Sensory physiology. In Deep-Sea Fishes; Randall, D.J., Farrell, A.P., Eds.; Academic Press: San Diego, CA, USA, 1997; pp. 325–349. [Google Scholar]
- Popper, A.N.; Hawkins, A.D. Hearing. In The Physiology of Fishes, 5th ed.; Currie, S., Evans, D.H., Eds.; CRC Press: Boca Raton, FL, USA, 2020; pp. 143–157. [Google Scholar]
- Walls, G.L. The Vertebrate Eye and Its Adaptive Radiation; Cranbrook Institute of Science Bulletin No. 19.; Cranbrook Press: Bloomfield Hills, MI, USA, 1942; 785p. [Google Scholar]
- Nicol, J.A.C. The Eyes of Fishes; Oxford University Press: Oxford, UK, 1989; 308p. [Google Scholar]
- Wittenberg, J.B.; Haedrich, R.L. The choroid rete mirabile of the fish eye. II. Distribution and relation to the pseudobranch and to the swimbladder rete mirabile. Biol. Bull. 1974, 146, 137–156. [Google Scholar] [CrossRef]
- Berenbrink, M.; Koldkjær, P.; Kepp, O.; Cossins, A.R. Evolution of Oxygen Secretion in Fishes and the Emergence of a Complex Physiological System. Science 2005, 307, 1752–1757. [Google Scholar] [CrossRef] [PubMed]
- Eastman, J.T. Aspects of the morphology of phyletically basal bovichtid fishes of the Antarctic suborder Notothenioidei (Perciformes). Polar Biol. 2006, 29, 754–763. [Google Scholar] [CrossRef]
- Demski, L.S. In a fish’s mind’s eye: The visual pallium of teleosts. In Sensory Processing in Aquatic Environments; Collin, S.P., Marshall, N.J., Eds.; Springer: New York, NY, USA, 2003; pp. 404–419. [Google Scholar]
- Montgomery, J.C.; Wells, R.M.G. Recent advances in the ecophysiology of Antarctic notothenioid fishes: Metabolic capacity and sensory performance. In Fish Ecophysiology; Rankin, J.C., Jensen, F.B., Eds.; Chapman and Hall: London, UK, 1993; pp. 341–374. [Google Scholar]
- Vrieze, L.A.; Bergstedt, R.A.; Sorensen, P.W. Olfactory-mediated stream-finding behavior of migratory adult sea lamprey (Petromyzon marinus). Can. J. Fish. Aquat. Sci. 2011, 68, 523–533. [Google Scholar] [CrossRef]
- Lim, H.; Sorensen, P.W. Common Carp Implanted with Prostaglandin F2α Release a Sex Pheromone Complex that Attracts Conspecific Males in Both the Laboratory and Field. J. Chem. Ecol. 2012, 38, 127–134. [Google Scholar] [CrossRef] [PubMed]
- Pakhomov, E.A. Feeding plasticity of the Antarctic fish Trematomus hansoni Boulenger, 1902 (Pisces: Nototheniidae): The influence of fishery waste on the diet. Polar Biol. 1998, 19, 289–292. [Google Scholar] [CrossRef]
- Montgomery, J.C.; Diebel, C.; Halstead, M.B.D.; Downer, J. Olfactory search tracks in the Antarctic fish Trematomus bernacchii. Polar Biol. 1999, 21, 151–154. [Google Scholar] [CrossRef]
- Smale, D.A.; Barnes, D.K.; Fraser, K.P.; Mann, P.J.; Brown, M.P. Scavenging in Antarctica: Intense variation between sites and seasons in shallow benthic necrophagy. J. Exp. Mar. Biol. Ecol. 2007, 349, 405–417. [Google Scholar] [CrossRef]
- Alfaro, M.E.; Faircloth, B.C.; Harrington, R.C.; Sorenson, L.; Friedman, M.; Thacker, C.E.; Oliveros, C.H.; Černý, D.; Near, T.J. Explosive diversification of marine fishes at the Cretaceous–Palaeogene boundary. Nat. Ecol. Evol. 2018, 2, 688–696. [Google Scholar] [CrossRef] [PubMed]
- Douglas, P.M.J.; Affek, H.P.; Ivany, L.C.; Houben, A.J.P.; Sijp, W.P.; Sluijs, A.; Schouten, S.; Pagani, M. Pronounced zonal heterogeneity in Eocene southern high-latitude sea surface temperatures. Proc. Natl. Acad. Sci. USA 2014, 111, 6582–6587. [Google Scholar] [CrossRef]
- Ivany, L.C.; Lohmann, K.C.; Hasiuk, F.; Blake, D.B.; Glass, A.; Aronson, R.B.; Moody, R.M. Eocene climate record of a high southern latitude continental shelf: Seymour Island, Antarctica. Geol. Soc. Am. Bull. 2008, 120, 659–678. [Google Scholar] [CrossRef]
- Přikryl, T.; Vodrážka, R. A diverse Eocene fish scale assemblage from Seymour Island, Antarctica. Geodiversitas 2012, 34, 895–908. [Google Scholar] [CrossRef]
- Kriwet, J. Additions to the Eocene selachian fauna of Antarctica with comments on Antarctic selachian diversity. J. Vertebr. Paléontol. 2005, 25, 1–7. [Google Scholar] [CrossRef]
- Kriwet, J.; Hecht, T. A review of early gadiform evolution and diversification: First record of a rattail fish skull (Gadiformes, Macrouridae) from the Eocene of Antarctica, with otoliths preserved in situ. Sci. Nat. 2008, 95, 899–907. [Google Scholar] [CrossRef] [PubMed]
- Engelbrecht, A.; Mörs, T.; Reguero, M.A.; Kriwet, J. Revision of Eocene Antarctic carpet sharks (Elasmobranchii, Orectolobiformes) from Seymour Island, Antarctic Peninsula. J. Syst. Palaeontol. 2016, 15, 969–990. [Google Scholar] [CrossRef] [PubMed]
- Engelbrecht, A.; Mörs, T.; Reguero, M.A.; Kriwet, J. A new sawshark, Pristiophorus laevis, from the Eocene of Antarctica with comments on Pristiophorus lanceolatus. Hist. Biol. 2016, 29, 841–853. [Google Scholar] [CrossRef] [PubMed]
- Engelbrecht, A.; Mörs, T.; Reguero, M.; Kriwet, J. New carcharhiniform sharks (Chondrichthyes, Elasmobranchii) from the early to middle Eocene of Seymour Island, Antarctic Peninsula. J. Vert. Paleont. 2017, 37, e1371724. [Google Scholar] [CrossRef] [PubMed]
- Engelbrecht, A.; Mörs, T.; Reguero, M.A.; Kriwet, J. Eocene squalomorph sharks (Chondrichthyes, Elasmobranchii) from Antarctica. J. S. Am. Earth Sci. 2017, 78, 175–189. [Google Scholar] [CrossRef] [PubMed]
- Engelbrecht, A.; Mörs, T.; Reguero, M.A.; Kriwet, J. Skates and rays (Elasmobranchii, Batomorphii) from the Eocene La Meseta and Submeseta formations, Seymour Island, Antarctica. Hist. Biol. 2018, 31, 1028–1044. [Google Scholar] [CrossRef] [PubMed]
- Kriwet, J.; Engelbrecht, A.; Mörs, T.; Reguero, M.; Pfaff, C. Ultimate Eocene (Priabonian) chondrichthyans (Holocephali, Elasmobranchii) of Antarctica. J. Vertebr. Paléontol. 2016, 36, e1160911. [Google Scholar] [CrossRef]
- Schwarzhans, W.; Mörs, T.; Engelbrecht, A.; Reguero, M.; Kriwet, J. Before the freeze: Otoliths from the Eocene of Seymour Island, Antarctica, reveal dominance of gadiform fishes (Teleostei). J. Syst. Palaeontol. 2016, 15, 147–170. [Google Scholar] [CrossRef]
- Reguero, M.; Goin, F.; Acosta Hospitaleche, C.; Dutra, T.; Marenssi, S. South America/West Antarctica: Pacific affinities of the Weddellian Marine/coastal vertebrates. In Late Cretaceous/Paleocene West Antarctica Terrestrial Biota and Its Intercontinental Affinities; Reguero, M., Goin, F., Acosta Hospitaleche, C., Dutra, T., Marenssi, S., Eds.; Springer: Dordrecht, The Netherlands, 2013; pp. 27–54. [Google Scholar]
- Eastman, J.T.; Grande, L. Late Eocene gadiform (Teleostei) skull from Seymour Island, Antarctic Peninsula. Antarct. Sci. 1991, 3, 87–95. [Google Scholar] [CrossRef]
- Balushkin, A.V. Proeleginops grandeastmanorum gen. et sp. nov. (Perciformes, Notothenioidei, Eleginopsidae) from the Late Eocene of Seymour Island (Antarctica) is a fossil notothenioid, not a gadiform. J. Ichthyol. 1994, 34, 10–23. [Google Scholar]
- Claeson, K.M.; Eastman, J.T.; Macphee, R.D.E. Definitive specimens of Merlucciidae (Gadiformes) from the Eocene James Ross Basin of Isla Marambio (Seymour Island), Antarctic Peninsula. Antarct. Sci. 2012, 24, 467–472. [Google Scholar] [CrossRef]
- Grande, L.; Eastman, J.T. A review of Antarctic ichthyofaunas in the light of new fossil discoveries. Palaeontology 1986, 29, 113–137. [Google Scholar]
- Jerzmańska, A.; Świdnicki, J. Gadiform remains from the La Meseta Formation (Eocene) of Seymour Island, West Antarctica. Pol. Polar Res. 1992, 13, 241–253. [Google Scholar]
- Bieńkowska-Wasiluk, M.; Bonde, N.; Møller, P.R.; Gaździcki, A. Eocene relatives of cod icefishes (Perciformes: Notothenioidei) from Seymour Island. Geol. Q. 2013, 57, 567–582. [Google Scholar] [CrossRef]
- Germain, D.; Mondéjar-Fernández, J.; Meunier, F.J. The detection of weakly developed plicidentine in teleost teeth using 3D tomography. Cybium 2016, 40, 75–82. [Google Scholar]
- Schwarzhans, W. Reconstruction of the fossil marine bony fish fauna (Teleostei) from the Eocene to Pleistocene of New Zealand by means of otoliths. Soc. Ital. Sci. Nat. Mus. Stor. Nat. 2019, 46, 1–326. [Google Scholar]
- Mecklenburg, C.W.; Lynghammar, A.; Johannesen, E.; Byrkjedal, I.; Christiansen, J.S.; Dolgov, A.V.; Karamushko, O.V.; Mecklenburg, T.A.; Møller, P.R.; Steinke, D.; et al. Marine Fishes of the Arctic Region; Conservation of Arctic Flora and Fauna: Akureyri, Iceland, 2018; Volume I, Available online: https://caff.is/marine/marine-monitoring-publications/451-marine-fishes-of-the-arctic-region-vol-1 (accessed on 1 January 2024).
- Whittle, R.J.; Quaglio, F.; Griffiths, H.J.; Linse, K.; Crame, J.A. The Early Miocene Cape Melville Formation fossil assemblage and the evolution of modern Antarctic marine communities. Naturwissenschaften 2014, 101, 47–59. [Google Scholar] [CrossRef]
- Quilty, P.G.; Lirio, J.M.; Jillett, D. Stratigraphy of the Pliocene Sørsdal Formation, Marine Plain, Vestfold Hills, East Antarctica. Antarct. Sci. 2000, 12, 205–216. [Google Scholar] [CrossRef]
- Near, T.J.; Ghezelayagh, A.; Ojeda, F.P.; Dornburg, A. Recent diversification in an ancient lineage of Notothenioid fishes (Bovichtus: Notothenioidei). Polar Biol. 2019, 42, 943–952. [Google Scholar] [CrossRef]
- Roa-Varón, A.; Ortí, G. Phylogenetic relationships among families of Gadiformes (Teleostei, Paracanthopterygii) based on nuclear and mitochondrial data. Mol. Phylogenetics Evol. 2009, 52, 688–704. [Google Scholar] [CrossRef] [PubMed]
- Baalsrud, H.T.; Tørresen, O.K.; Solbakken, M.H.; Salzburger, W.; Hanel, R.; Jakobsen, K.S.; Jentoft, S. De Novo Gene Evolution of Antifreeze Glycoproteins in Codfishes Revealed by Whole Genome Sequence Data. Mol. Biol. Evol. 2017, 35, 593–606. [Google Scholar] [CrossRef] [PubMed]
- Zhaung, X.; Yang, C.; Murphy, K.R.; Cheng, C.-H.C. Molecular mechanism and history of non-sense to sense evolution of antifreeze glycoprotein gene in northern gadids. Proc. Natl. Acad. Sci. USA 2019, 116, 4400–4405. [Google Scholar] [CrossRef] [PubMed]
- Coleman, M.; Wernberg, T. The Silver Lining of Extreme Events. Trends Ecol. Evol. 2020, 35, 1065–1067. [Google Scholar] [CrossRef] [PubMed]
- Devries, A.L. Fish antifreeze proteins. In Antifreeze Proteins; Ramløv, H., Friis, D.S., Eds.; Springer Nature International: Cham, Switzerland, 2020; Volume 1, pp. 85–129. [Google Scholar]
- Chen, L.; Devries, A.L.; Cheng, C.-H.C. Evolution of antifreeze glycoprotein gene from a trypsinogen gene in Antarctic notothenioid fish. Proc. Natl. Acad. Sci. USA 1997, 94, 3811–3816. [Google Scholar] [CrossRef] [PubMed]
- DeVries, A.L. Antifreeze Peptides and Glycopeptides in Cold-Water Fishes. Annu. Rev. Physiol. 1983, 45, 245–260. [Google Scholar] [CrossRef] [PubMed]
- Jung, A.; Johnson, P.; Eastman, J.T.; DeVries, A.L. Protein content and freezing avoidance properties of the subdermal extracellular matrix and serum of the Antarctic snailfish, Paraliparis devriesi. Fish Physiol. Biochem. 1995, 14, 71–80. [Google Scholar] [CrossRef] [PubMed]
- Evans, R.P.; Fletcher, G.L. Isolation and characterization of type I antifreeze proteins from Atlantic snailfish (Liparis atlanticus) and dusky snailfish (Liparis gibbus). Biochim. Biophys. Acta (BBA) Protein Struct. Mol. Enzym. 2001, 1547, 235–244. [Google Scholar] [CrossRef]
- Devries, A.L.; Cheng, C.-H.C. Antifreeze proteins and organismal freezing avoidance in polar fishes. In Fish Physiology: The Physiology of Polar Fishes; Farrell, A.P., Steffensen, J.F., Eds.; Academic Press: San Diego, CA, USA, 2005; Volume 22, pp. 155–201. [Google Scholar]
- Anderson, M.E.; Fedorov, V.V. Family Zoarcidae Swainson 1839—Eelpouts. Calif. Acad. Sci. Annot. Checklists Fish. 2004, 34, 1–58. [Google Scholar]
- Stein, D.L. Snailfishes (family Liparidae) of the Ross Sea, Antarctica, and closely adjacent waters. Zootaxa 2012, 3285, 1–120. [Google Scholar] [CrossRef]
- Stein, D.L. A review of the snailfishes (Liparidae, Scorpaeniformes) of New Zealand, including descriptions of a new genus and sixteen new species. Zootaxa 2012, 3588, 1–54. [Google Scholar] [CrossRef]
- Eastman, J.T.; Hikida, R.S.; Devries, A.L. Buoyancy studies and microscopy of skin and subdermal extracellular matrix of the antarctic snailfish, Paraliparis devriesi. J. Morphol. 1994, 220, 85–101. [Google Scholar] [CrossRef] [PubMed]
- Balushkin, A.V.; Moganova, M.V. New species of pelagic eelpouts of the genus Melanostigma (Zoarcidae) from Western Antarctica. J. Ichthyol. 2017, 57, 171–176. [Google Scholar] [CrossRef]
- Gerringer, M.E.; Drazen, J.C.; Linley, T.D.; Summers, A.P.; Jamieson, A.J.; Yancey, P.H. Distribution, composition and functions of gelatinous tissues in deep-sea fishes. R. Soc. Open Sci. 2017, 4, 171063. [Google Scholar] [CrossRef] [PubMed]
- Hobbs, R.S.; Hall, J.R.; Graham, L.A.; Davies, P.L.; Fletcher, G.L. Antifreeze protein dispersion in eelpouts and related fishes reveals migration and climate alteration within the last 20 Ma. PLoS ONE 2020, 15, e0243273. [Google Scholar] [CrossRef]
- Rabosky, D.L.; Santini, F.; Eastman, J.; Smith, S.A.; Sidlauskas, B.; Chang, J.; Alfaro, M.E. Rates of speciation and morphological evolution are correlated across the largest vertebrate radiation. Nat. Commun. 2013, 4, 1958. [Google Scholar] [CrossRef]
Figure 1.
Phylogeny of notothenioids based on RADseq loci (Near et al., 2015, 2018) [2,3], with mapping of characteristic features (not used in the analysis). Eighteen species of the Antarctic clade («cryonotothenioids») have non-Antarctic distributions.

Figure 2.
«Cryonotothenioids» dominate fish diversity, abundance, and biomass in high latitude (73°–77° S) waters exemplified by the southwestern Ross Sea. Data from benthic trawls (Eastman and Hubold, 1999) [17], updated from Eastman (2005, Figure 2) [19].

Figure 3.
Scatterplot showing maximum depths of occurrence for 140 species of notothenioids, with colors indicating the consolidated phylogenetically basal taxa and the five «cryonotothenioid» families. Based on data from Eastman (2017) [43].
Figure 3.
Scatterplot showing maximum depths of occurrence for 140 species of notothenioids, with colors indicating the consolidated phylogenetically basal taxa and the five «cryonotothenioid» families. Based on data from Eastman (2017) [43].

Figure 4.
Species diversity across 100 m depth increments for 31 species of non-Antarctic, 33 species of sub-Antarctic, and 64 species of Antarctic notothenioids. The first dot on the left (without a corresponding tick mark on the x-axis) represents the number of species found at 0–50 m. Subsequent dots and tick marks represent counts that include the total number of species found at depths 51–100 m, 101–200 m, etc. From Eastman (2017) [43].
Figure 4.
Species diversity across 100 m depth increments for 31 species of non-Antarctic, 33 species of sub-Antarctic, and 64 species of Antarctic notothenioids. The first dot on the left (without a corresponding tick mark on the x-axis) represents the number of species found at 0–50 m. Subsequent dots and tick marks represent counts that include the total number of species found at depths 51–100 m, 101–200 m, etc. From Eastman (2017) [43].

Figure 5.
Scatterplot of mean percentage buoyancy values (%B) for 59 species of notothenioids, with colors indicating the consolidated phylogenetically basal families and the five «cryonotothenioid» families. Horizontal lines delimit the biotopes. All species are identified on the horizontal axis. Modified from Eastman (2020) [45].
Figure 5.
Scatterplot of mean percentage buoyancy values (%B) for 59 species of notothenioids, with colors indicating the consolidated phylogenetically basal families and the five «cryonotothenioid» families. Horizontal lines delimit the biotopes. All species are identified on the horizontal axis. Modified from Eastman (2020) [45].

Figure 6.
Ontogenetic change in buoyancy in the demersal sister species Notothenia rossii and N. coriiceps, with trend lines fitted by a quadric model. Regraphed from Eastman et al. (2011 [68], Figure 3a, © Inter-Research, 2011, with permission. Fish drawings reproduced with permission from Fishes of the Southern Ocean (1990, Figures 28 and 30), © South African Institute for Aquatic Biodiversity).
Figure 6.
Ontogenetic change in buoyancy in the demersal sister species Notothenia rossii and N. coriiceps, with trend lines fitted by a quadric model. Regraphed from Eastman et al. (2011 [68], Figure 3a, © Inter-Research, 2011, with permission. Fish drawings reproduced with permission from Fishes of the Southern Ocean (1990, Figures 28 and 30), © South African Institute for Aquatic Biodiversity).

Figure 7.
Histogram showing maximum total lengths of 138 species of notothenioids. Two species of Pogonophryne are omitted because they are known only from their juvenile holotypes. Data are in 10 cm-bins, centered on the midpoint of the bin. From Eastman (2019) [44].
Figure 7.
Histogram showing maximum total lengths of 138 species of notothenioids. Two species of Pogonophryne are omitted because they are known only from their juvenile holotypes. Data are in 10 cm-bins, centered on the midpoint of the bin. From Eastman (2019) [44].

Figure 8.
Scatterplot of maximum total lengths for 138 species of notothenioids, with colors indicating the consolidated phylogenetically basal families and the five «cryonotothenioid» families. Horizontal lines delimit the four size categories. Regraphed with data from Eastman (2019) [44].
Figure 8.
Scatterplot of maximum total lengths for 138 species of notothenioids, with colors indicating the consolidated phylogenetically basal families and the five «cryonotothenioid» families. Horizontal lines delimit the four size categories. Regraphed with data from Eastman (2019) [44].

Figure 9.
Divergence in body size and shape among 11 taxa representing 102 species or 80% of «cryonotothenioid» diversity. Powers of 10 axes accommodate the 39-fold interspecific difference in maximum total lengths. Image sizes are scaled to an assumed zero origin of the x-axis, although this is not shown. Representative species are from largest to smallest: (a) Dissostichus eleginoides, (b) Notothenia rossii, (c) Chaenocephalus aceratus, (d) Parachaenichthys georgianus, (e) Gobionotothen gibberifrons, (f) Trematomus lepidorhinus, (g) Pogonophryne cerebropogon, (h) Vomeridens infuscipinnis, (i) Nototheniops nudifrons, (j) Artedidraco glareobarbatus, and (k) Harpagifer antarcticus.
Figure 9.
Divergence in body size and shape among 11 taxa representing 102 species or 80% of «cryonotothenioid» diversity. Powers of 10 axes accommodate the 39-fold interspecific difference in maximum total lengths. Image sizes are scaled to an assumed zero origin of the x-axis, although this is not shown. Representative species are from largest to smallest: (a) Dissostichus eleginoides, (b) Notothenia rossii, (c) Chaenocephalus aceratus, (d) Parachaenichthys georgianus, (e) Gobionotothen gibberifrons, (f) Trematomus lepidorhinus, (g) Pogonophryne cerebropogon, (h) Vomeridens infuscipinnis, (i) Nototheniops nudifrons, (j) Artedidraco glareobarbatus, and (k) Harpagifer antarcticus.

Figure 10.
Relative dry skeletal masses for 62 individuals representing 20 species of notothenioids, two species of percids (perches), and one species of serranid (sea basses). Note that Eleginops maclovinus clusters among the values for «cryonotothenioids». Regraphed from Eastman et al. (2014) [42].
Figure 10.
Relative dry skeletal masses for 62 individuals representing 20 species of notothenioids, two species of percids (perches), and one species of serranid (sea basses). Note that Eleginops maclovinus clusters among the values for «cryonotothenioids». Regraphed from Eastman et al. (2014) [42].

Figure 11.
Left lateral micro-CT images of heads of adults: (top) Bovichtus diacanthus; (middle) Eleginops maclovinus; (bottom) Chaenodraco wilsoni. Colors are standardized across panels and serve as proxies for the amounts of compact bone (red), cancellous bone (green), and cartilage (blue). The degree of bone mineralization is similar among «cryonotothenioids» where the density-reducing mechanism involves, instead, ontogenetic displacements in the relative amounts of bony, cartilaginous, and notochordal tissues. Bovichtus has a typical well ossified skeleton indicated by red. Eleginops have a cancellous (spongy) bone that registers as green. In Chaenodraco, much of the neurocranial base is cartilage and is not imaged here. Jaw and opercular elements are ossified but extensive cartilage persists in the branchiopharyngeal region and in the pectoral girdle (blue) that also shows foci of weak ossification in the radials (green). The absence of significant bone in the branchiopharyngeal and intercleithral areas facilitates expansion and the wide pharyngeal gape that enables piscivory in some channichthyids. Modified from Eastman et al. (2014) [42].
Figure 11.
Left lateral micro-CT images of heads of adults: (top) Bovichtus diacanthus; (middle) Eleginops maclovinus; (bottom) Chaenodraco wilsoni. Colors are standardized across panels and serve as proxies for the amounts of compact bone (red), cancellous bone (green), and cartilage (blue). The degree of bone mineralization is similar among «cryonotothenioids» where the density-reducing mechanism involves, instead, ontogenetic displacements in the relative amounts of bony, cartilaginous, and notochordal tissues. Bovichtus has a typical well ossified skeleton indicated by red. Eleginops have a cancellous (spongy) bone that registers as green. In Chaenodraco, much of the neurocranial base is cartilage and is not imaged here. Jaw and opercular elements are ossified but extensive cartilage persists in the branchiopharyngeal region and in the pectoral girdle (blue) that also shows foci of weak ossification in the radials (green). The absence of significant bone in the branchiopharyngeal and intercleithral areas facilitates expansion and the wide pharyngeal gape that enables piscivory in some channichthyids. Modified from Eastman et al. (2014) [42].

Figure 15.
Radiographs of an ontogenetic series of Pleuragramma antarcticum exemplifying the neotenic retention and large size of the (a) notochord (N) and the relatively small amount of vertebral bone (white in radiographs) during early ontogeny. Later (b,c) as the mineralization of the notochordal sheath proceeds, the notochord becomes reduced in size and centra becomes more bony. Although the patency of the vertebral canal (VC) decreases as the centra become increasingly amphicelous, it is still evident in larger individuals (c).
Figure 15.
Radiographs of an ontogenetic series of Pleuragramma antarcticum exemplifying the neotenic retention and large size of the (a) notochord (N) and the relatively small amount of vertebral bone (white in radiographs) during early ontogeny. Later (b,c) as the mineralization of the notochordal sheath proceeds, the notochord becomes reduced in size and centra becomes more bony. Although the patency of the vertebral canal (VC) decreases as the centra become increasingly amphicelous, it is still evident in larger individuals (c).

Figure 16.
Transverse histological section of the trunk of an adult Pleuragramma antarcticum (123 mm SL) showing the persistent notochord, relative sizes of the lipid sacs (LS), and a small amount of bone. The vertebral body consists of a thin ring of bone (arrows) surrounding the notochordal vacuoles (V). Arrows also indicate the delicate bone of the hemal arch. The distal pterygiophore is composed of cartilage (C). The lateralis superficialis muscle, consisting of small-diameter red muscle fibers (RM), encircles the trunk. Because it inserts on the caudal fin rays via tendons separate from those of the white myotomal musculature, it provides an energetically efficient means for short bursts of subcarangiform locomotion used for repositioning during “hanging” in the water column, which is typical behavior for this species.
Figure 16.
Transverse histological section of the trunk of an adult Pleuragramma antarcticum (123 mm SL) showing the persistent notochord, relative sizes of the lipid sacs (LS), and a small amount of bone. The vertebral body consists of a thin ring of bone (arrows) surrounding the notochordal vacuoles (V). Arrows also indicate the delicate bone of the hemal arch. The distal pterygiophore is composed of cartilage (C). The lateralis superficialis muscle, consisting of small-diameter red muscle fibers (RM), encircles the trunk. Because it inserts on the caudal fin rays via tendons separate from those of the white myotomal musculature, it provides an energetically efficient means for short bursts of subcarangiform locomotion used for repositioning during “hanging” in the water column, which is typical behavior for this species.

Figure 17.
This ≈65 cm TL Dissostichus mawsoni documents the existence of a benthic phase in the life history of this species. These are screenshots from a video taken by an ROV at 400 m in Gerlache Strait off the western Antarctic Peninsula (≈64° S). When the ROV arrived, the fish was out of sight under the rock ledge behind it. It returned under the ledge after positioning (a) and repositioning (b) itself on the substrate in response to the lights and noise of the ROV. (c) Demersal Trematomus scotti engulfing a krill from a passing swarm. Photos © NHK/National Geographic WILD/ZDF, used with permission.
Figure 17.
This ≈65 cm TL Dissostichus mawsoni documents the existence of a benthic phase in the life history of this species. These are screenshots from a video taken by an ROV at 400 m in Gerlache Strait off the western Antarctic Peninsula (≈64° S). When the ROV arrived, the fish was out of sight under the rock ledge behind it. It returned under the ledge after positioning (a) and repositioning (b) itself on the substrate in response to the lights and noise of the ROV. (c) Demersal Trematomus scotti engulfing a krill from a passing swarm. Photos © NHK/National Geographic WILD/ZDF, used with permission.

Figure 18.
Premaxillae, dentaries, and teeth of: (a) Eleginops maclovinus, 74.5 cm TL; (b) Notothenia coriiceps, 58.7 cm TL; (c) Trematomus bernacchii, 24.6 cm TL; (d) N. rossii, 41.8 cm TL; (e) Dissostichus eleginoides, 65 cm TL. Only the tooth pedicles are present in (a,b); the white arrow in (c) indicates worn tooth surface; black arrows in (d) show the ligament attaching the crown to the pedicle.
Figure 18.
Premaxillae, dentaries, and teeth of: (a) Eleginops maclovinus, 74.5 cm TL; (b) Notothenia coriiceps, 58.7 cm TL; (c) Trematomus bernacchii, 24.6 cm TL; (d) N. rossii, 41.8 cm TL; (e) Dissostichus eleginoides, 65 cm TL. Only the tooth pedicles are present in (a,b); the white arrow in (c) indicates worn tooth surface; black arrows in (d) show the ligament attaching the crown to the pedicle.

Figure 19.
Premaxillae, dentaries, and teeth of: (a) Parachaenichthys charcoti, 37.2 cm SL; (b) Pseudochaenichthys georgianus, 45.8 cm SL; (c) Chaenocephalus aceratus, 27.9 cm SL.
Figure 19.
Premaxillae, dentaries, and teeth of: (a) Parachaenichthys charcoti, 37.2 cm SL; (b) Pseudochaenichthys georgianus, 45.8 cm SL; (c) Chaenocephalus aceratus, 27.9 cm SL.

Figure 20.
Radiographs of the differently shaped heads and jaws of two large species of primarily piscivorous channichthyids. (a) Chaenocephalus aceratus (44.5 cm TL) has a wide U-shaped snout and engulfs large prey head-first. It has more rugged and well-ossified jaws as indicated by the whiteness, especially of the dentary. (b) Neopagetopsis ionah (57.5 cm TL) has a narrow V-shaped snout, slightly bowed more gracile jaws, and consumes Pleuragramma antarcticum and krill. Arteries of both specimens contain a radiopaque compound.
Figure 20.
Radiographs of the differently shaped heads and jaws of two large species of primarily piscivorous channichthyids. (a) Chaenocephalus aceratus (44.5 cm TL) has a wide U-shaped snout and engulfs large prey head-first. It has more rugged and well-ossified jaws as indicated by the whiteness, especially of the dentary. (b) Neopagetopsis ionah (57.5 cm TL) has a narrow V-shaped snout, slightly bowed more gracile jaws, and consumes Pleuragramma antarcticum and krill. Arteries of both specimens contain a radiopaque compound.

Figure 21.
The oral (a) and pharyngeal (b) gapes in the large-headed (≈40% of SL) piscivorous channichthyid Chaenocephalus aceratus. Arrows in (b) indicate the posterior pharyngobranchials (dorsal) and the fifth ceratobranchials (ventral). The pharyngeal lumen/gape is the darker area immediately posterior to these branchial elements.
Figure 21.
The oral (a) and pharyngeal (b) gapes in the large-headed (≈40% of SL) piscivorous channichthyid Chaenocephalus aceratus. Arrows in (b) indicate the posterior pharyngobranchials (dorsal) and the fifth ceratobranchials (ventral). The pharyngeal lumen/gape is the darker area immediately posterior to these branchial elements.

Figure 22.
A gravid female Chaenocephalus aceratus (a) has swallowed a Champsocephalus gunnari that was at least 50% of its own length. Photo courtesy of Professor Lisa Crockett, Ohio University. Semipelagic channichthyid Pagetodes antarcticus (b) in a bipodal stance on the substrate. Screenshot from a video taken by an ROV in the Gerlache Strait off the western Antarctic Peninsula (≈64° S), courtesy of NHK Broadcasting, Japan (Deep Antarctica production). Photo © NHK/National Geographic WILD/ZDF, used with permission.
Figure 22.
A gravid female Chaenocephalus aceratus (a) has swallowed a Champsocephalus gunnari that was at least 50% of its own length. Photo courtesy of Professor Lisa Crockett, Ohio University. Semipelagic channichthyid Pagetodes antarcticus (b) in a bipodal stance on the substrate. Screenshot from a video taken by an ROV in the Gerlache Strait off the western Antarctic Peninsula (≈64° S), courtesy of NHK Broadcasting, Japan (Deep Antarctica production). Photo © NHK/National Geographic WILD/ZDF, used with permission.

Figure 23.
(a) Radiograph of semipelagic channichthyid Dacodraco hunteri (233 mm TL) showing bowed jaws, and large and small, sharp conical teeth; thin laminae of bone (white arrows) on dorsal and ventral surfaces of the cartilaginous neurocranium; and persistent notochordal canals (black arrows) in vertebral centra. (b) Fresh specimen of semipelagic channichthyid Neopagetopsis ionah showing streamlined body and coloration typical of a pelagic species. (c) Semipelagic channichthyid Chionobathyscus dewitti, with characteristic wide non-tapering snout, holding position in the water column above the substrate. Panel c, modified from Eastman et al. (2013) [200].
Figure 23.
(a) Radiograph of semipelagic channichthyid Dacodraco hunteri (233 mm TL) showing bowed jaws, and large and small, sharp conical teeth; thin laminae of bone (white arrows) on dorsal and ventral surfaces of the cartilaginous neurocranium; and persistent notochordal canals (black arrows) in vertebral centra. (b) Fresh specimen of semipelagic channichthyid Neopagetopsis ionah showing streamlined body and coloration typical of a pelagic species. (c) Semipelagic channichthyid Chionobathyscus dewitti, with characteristic wide non-tapering snout, holding position in the water column above the substrate. Panel c, modified from Eastman et al. (2013) [200].

Figure 26.
The bathydraconid Vomeridens infuscipinnis, a semipelagic species that hover above the substrate. Note the elongated body, large pectoral fins (29% of SL in this specimen), and thin skin of the pectoral and pelvic fins. Permission from and courtesy of Professor Thomas J. Near, Yale University.
Figure 26.
The bathydraconid Vomeridens infuscipinnis, a semipelagic species that hover above the substrate. Note the elongated body, large pectoral fins (29% of SL in this specimen), and thin skin of the pectoral and pelvic fins. Permission from and courtesy of Professor Thomas J. Near, Yale University.

Figure 30.
Transverse sections showing retinal histology of Dissostichus mawsoni (a,c) and Pleuragramma antarcticum (b,d). Sections are aligned along the plane of the external limiting membrane. Original magnifications, ×285. Sections (a,b) are stained to highlight cone ellipsoids in the photoreceptor layer that are more numerous in Pleuragramma. Sections (c,d) are stained to visualize nuclei in various layers of the retina. 1, retinal pigment epithelium; 2, outer segments of photoreceptors; 3, inner segments of photoreceptors (myoid and ellipsoids); 4, external limiting membrane; 5, external nuclear layer; 6, internal nuclear layer; 7, ganglion cell layer; 8, optic nerve fibers.
Figure 30.
Transverse sections showing retinal histology of Dissostichus mawsoni (a,c) and Pleuragramma antarcticum (b,d). Sections are aligned along the plane of the external limiting membrane. Original magnifications, ×285. Sections (a,b) are stained to highlight cone ellipsoids in the photoreceptor layer that are more numerous in Pleuragramma. Sections (c,d) are stained to visualize nuclei in various layers of the retina. 1, retinal pigment epithelium; 2, outer segments of photoreceptors; 3, inner segments of photoreceptors (myoid and ellipsoids); 4, external limiting membrane; 5, external nuclear layer; 6, internal nuclear layer; 7, ganglion cell layer; 8, optic nerve fibers.

Figure 37.
Taxonomic composition of (a) 263 species of Antarctic and sub-Antarctic species from the shelf and upper slope depths of the Southern Ocean compared with (b) 205 species from the Arctic Region. Antarctic data from Eastman (unpublished); Arctic data from Mecklenburg et al. (2018) [290].
Figure 37.
Taxonomic composition of (a) 263 species of Antarctic and sub-Antarctic species from the shelf and upper slope depths of the Southern Ocean compared with (b) 205 species from the Arctic Region. Antarctic data from Eastman (unpublished); Arctic data from Mecklenburg et al. (2018) [290].


{kind=link}
{kind=link}
{kind=link}
{kind=link}
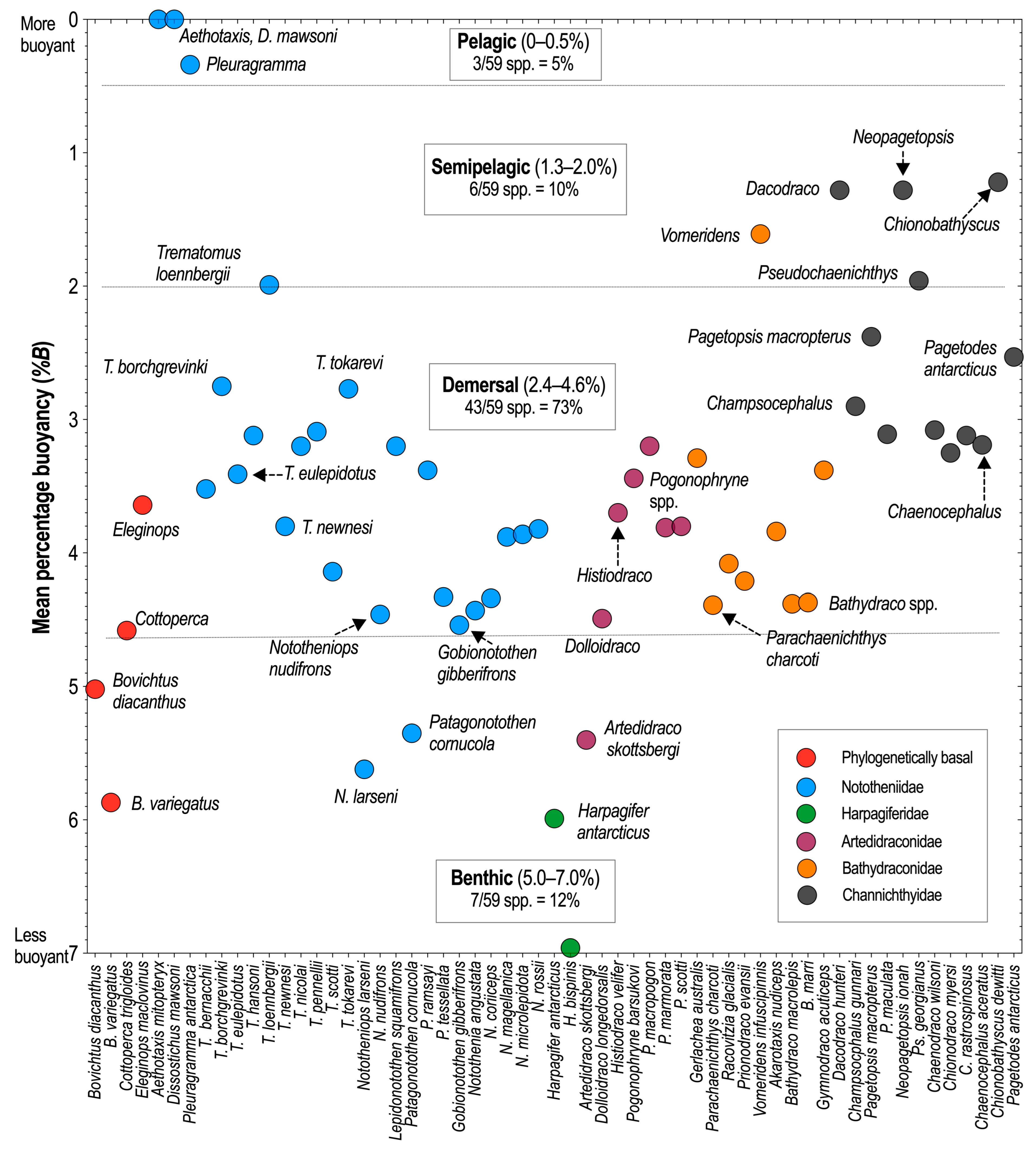{kind=link}
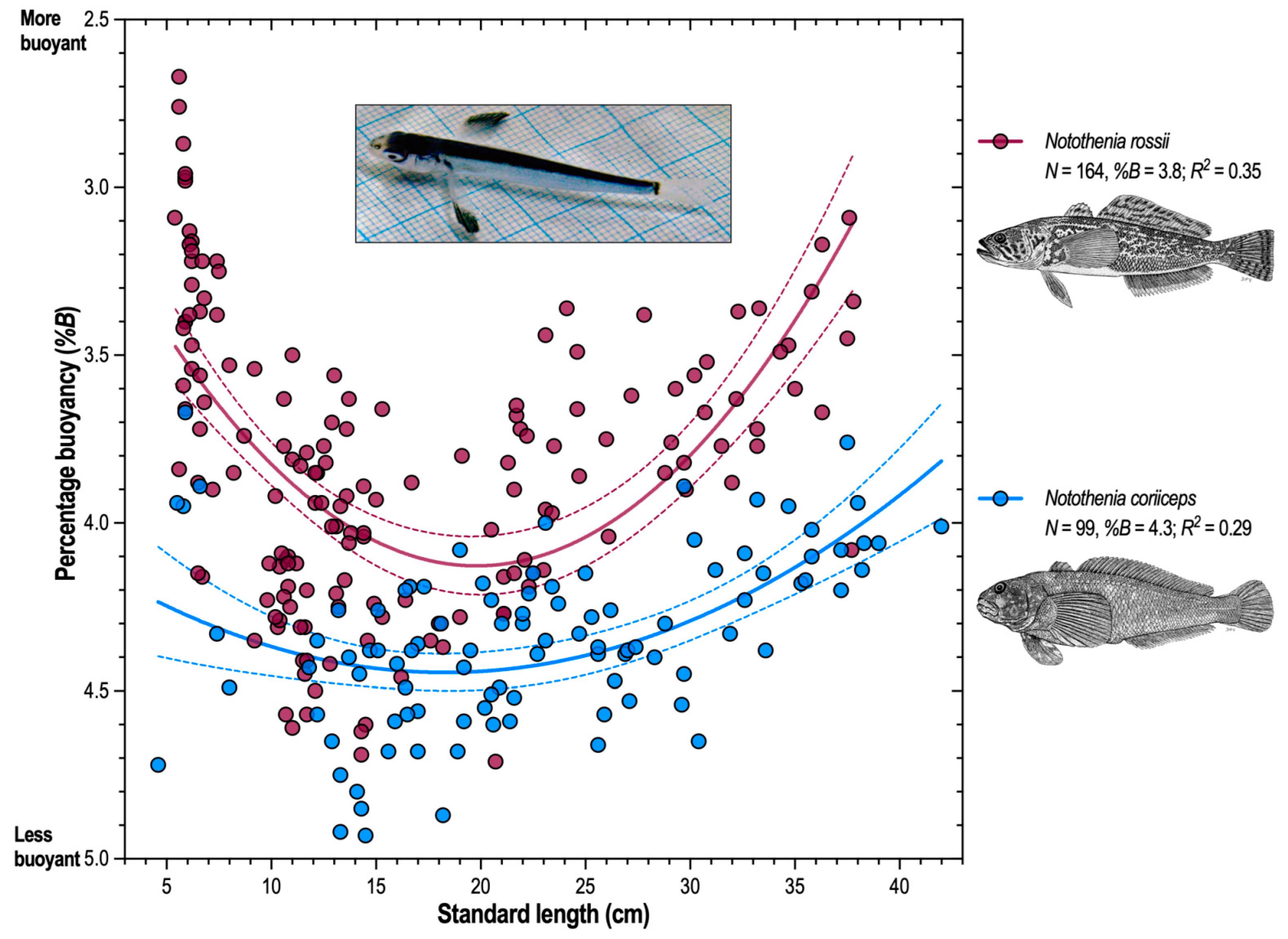{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
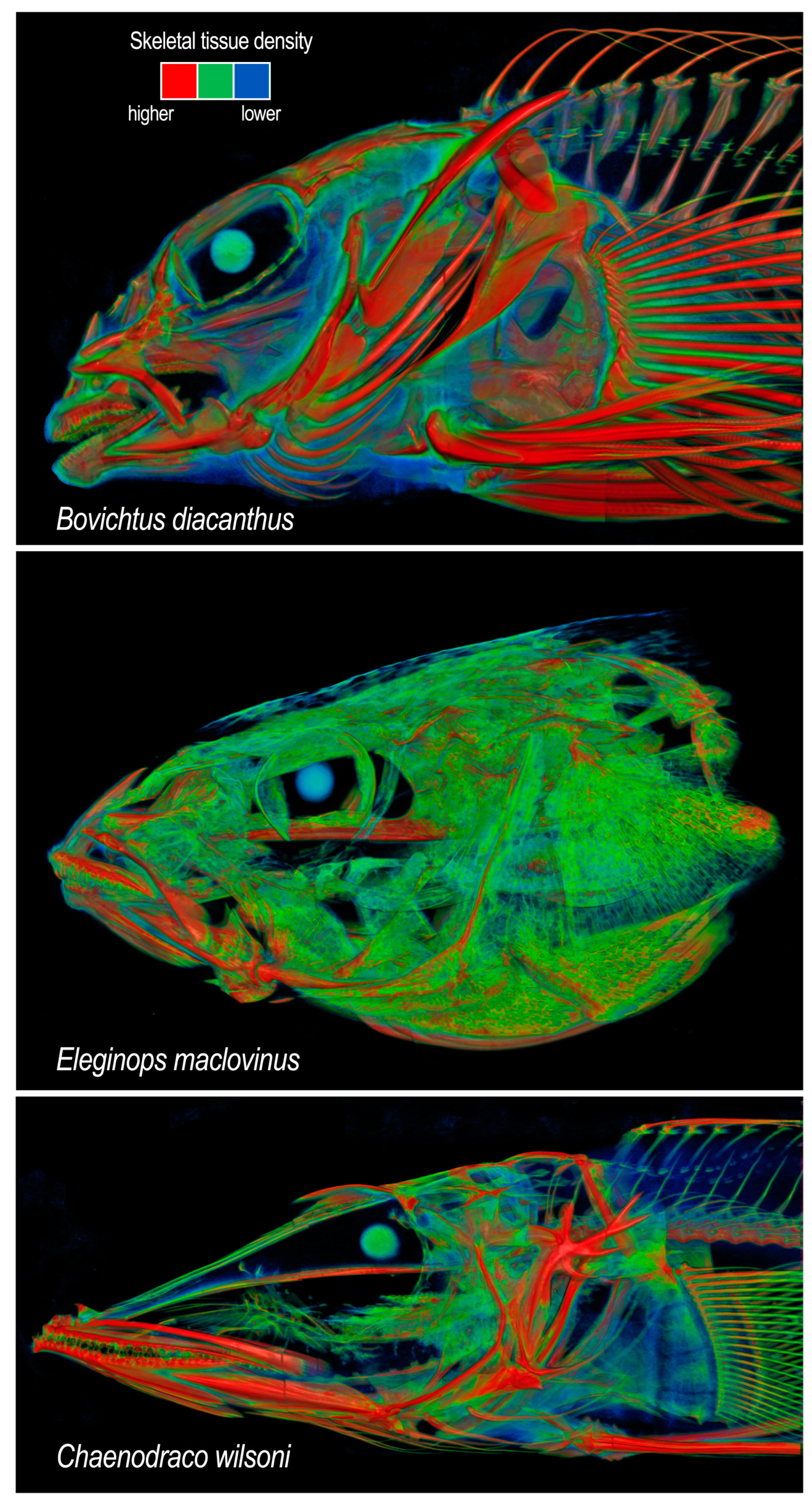{kind=link}
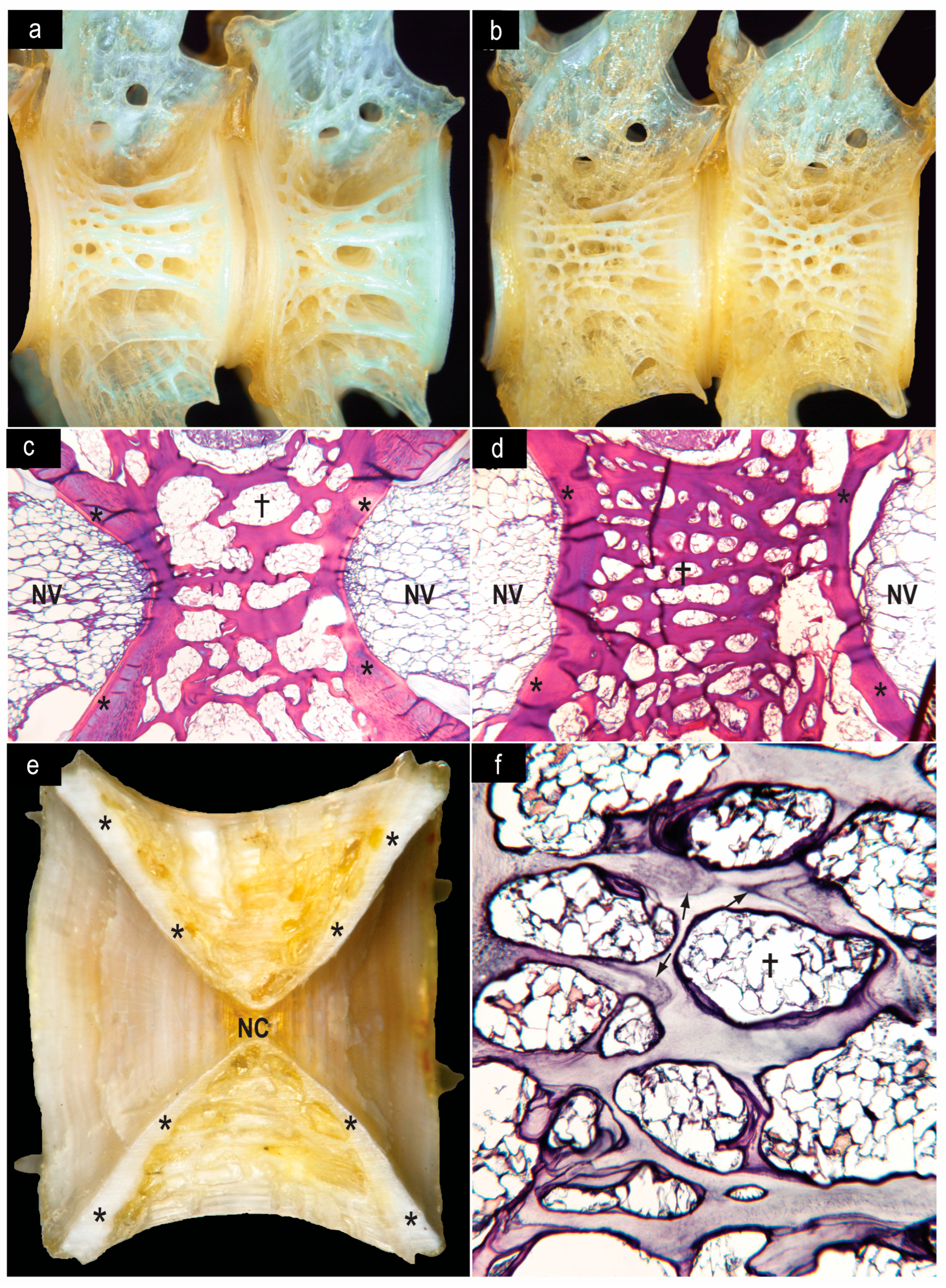{kind=link}
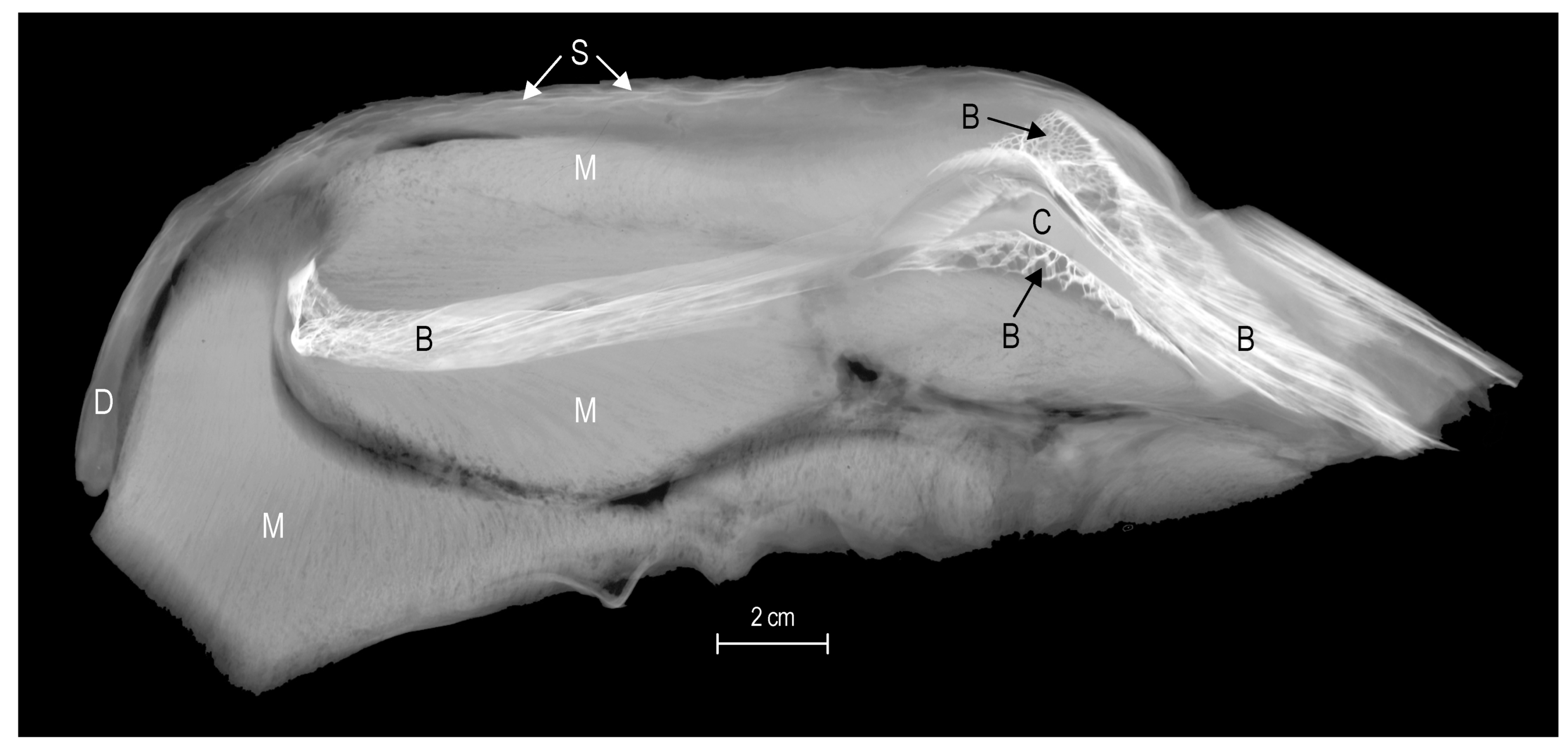{kind=link}
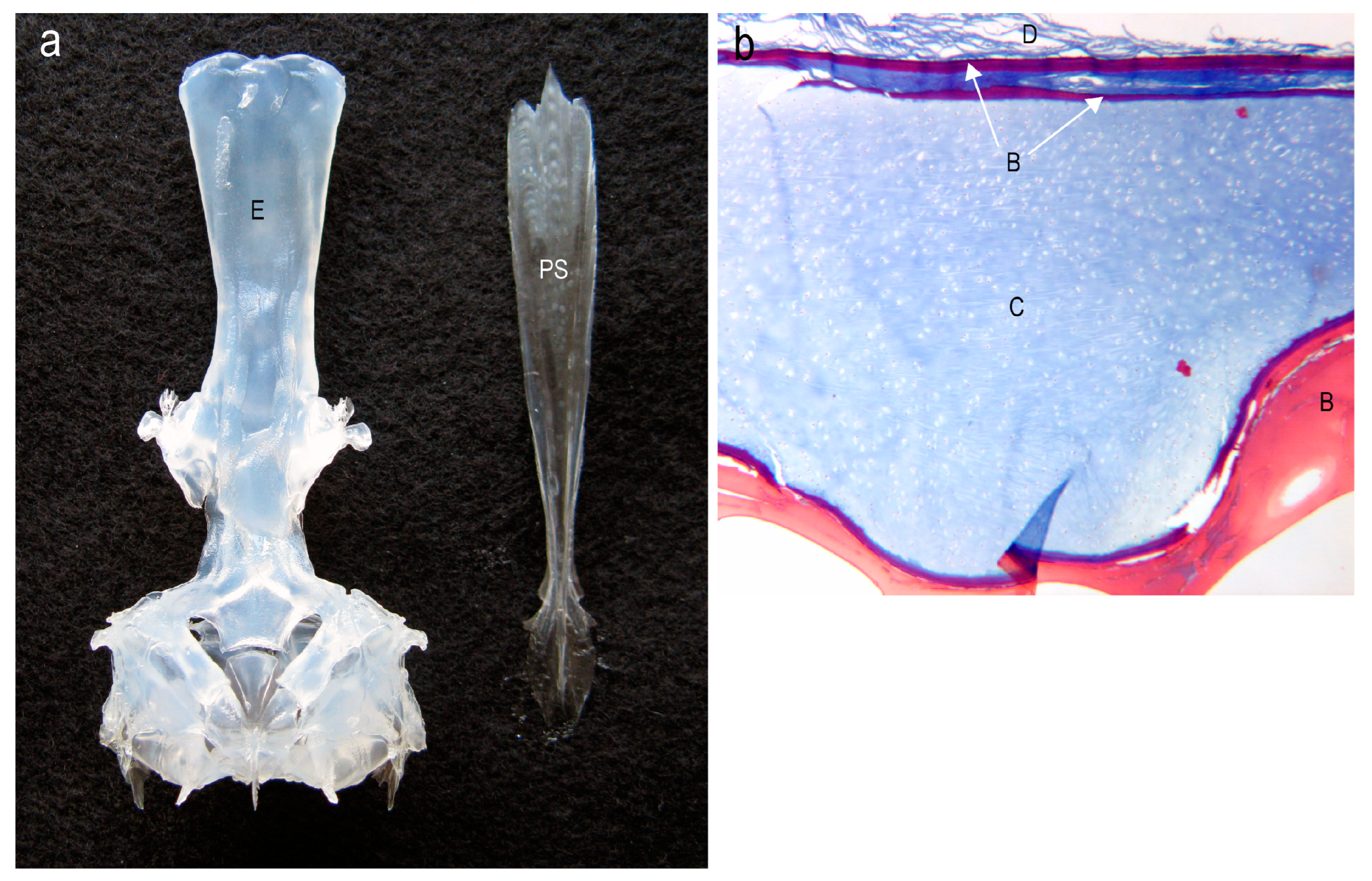{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
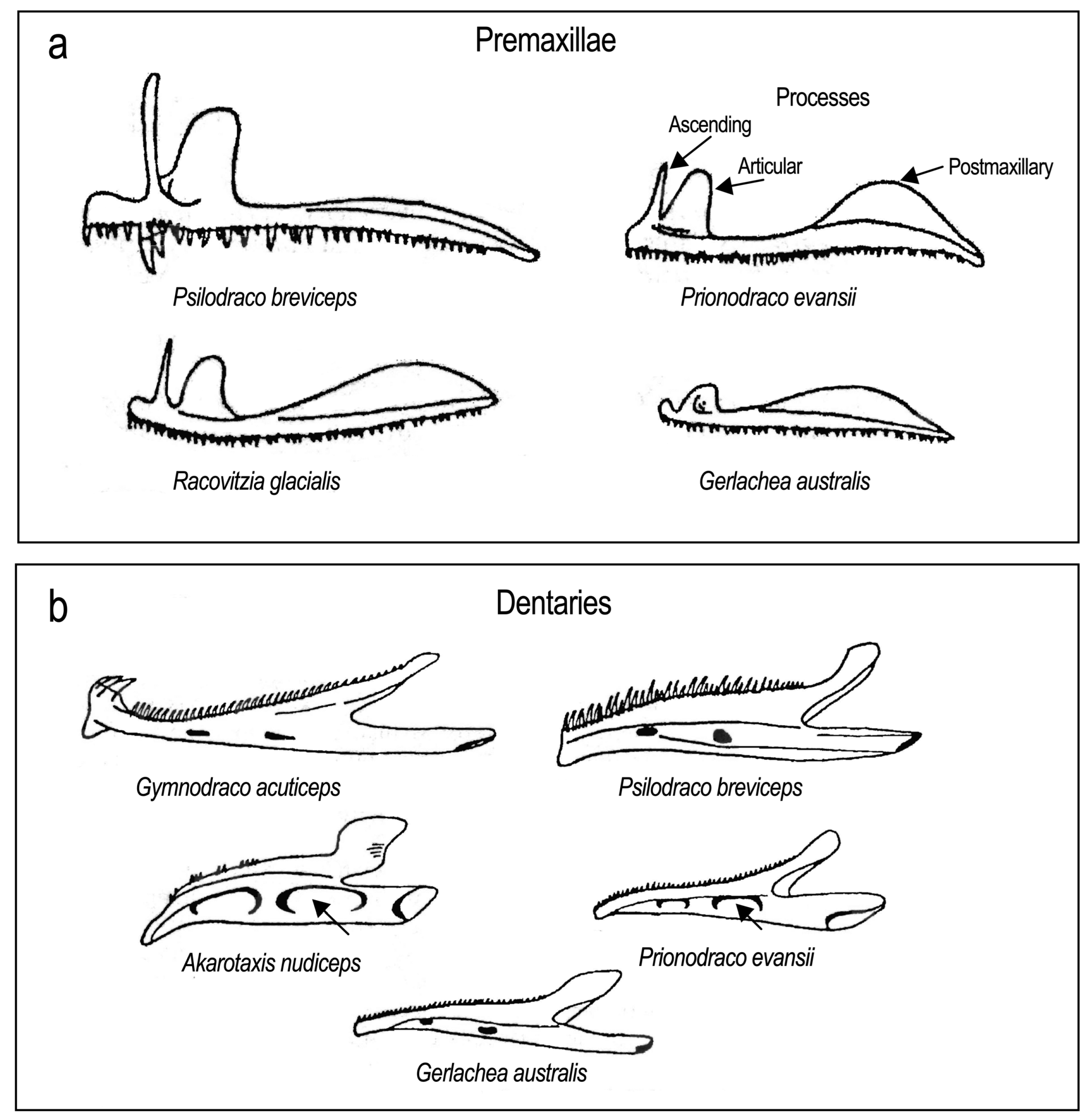{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
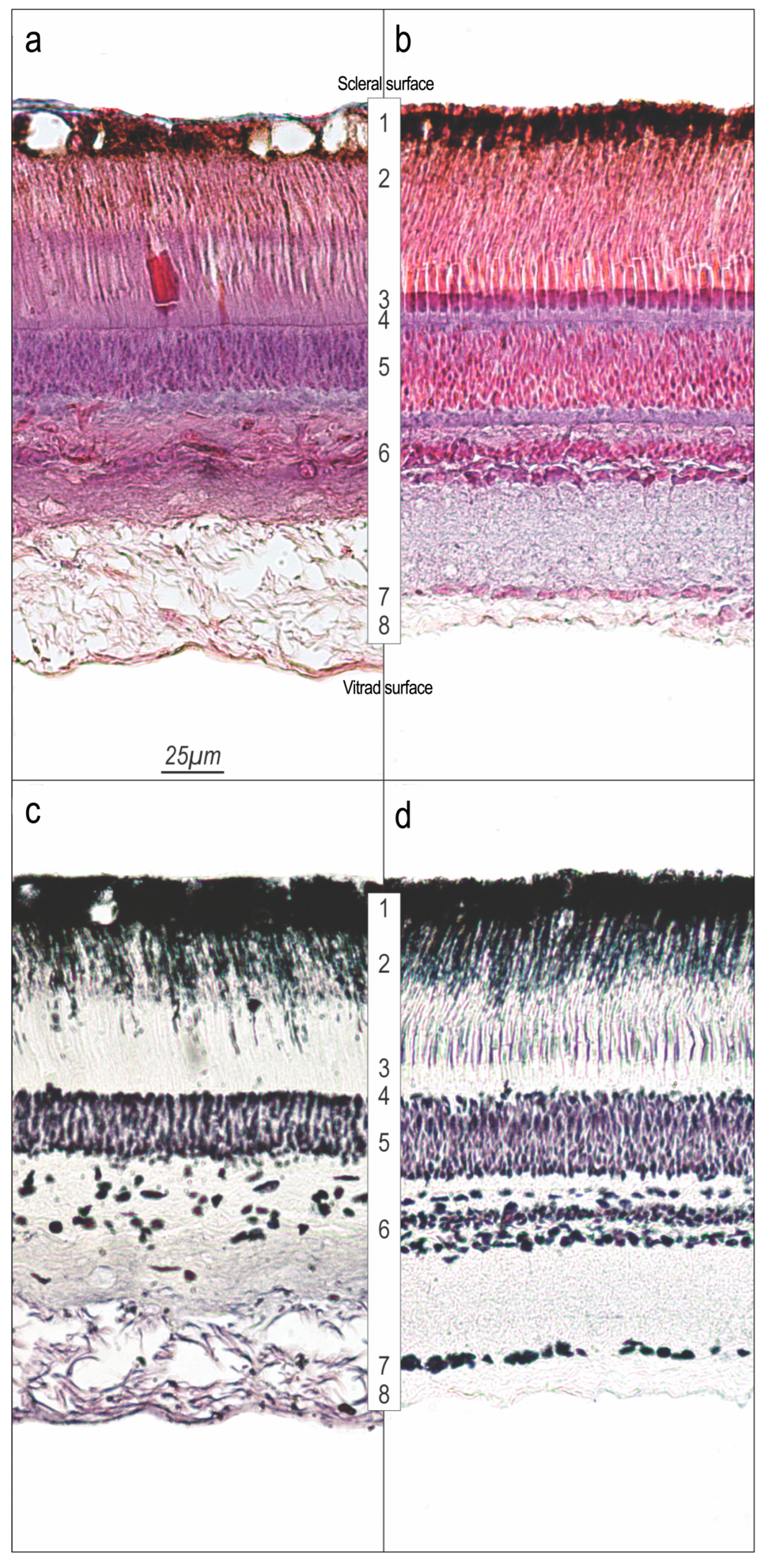{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
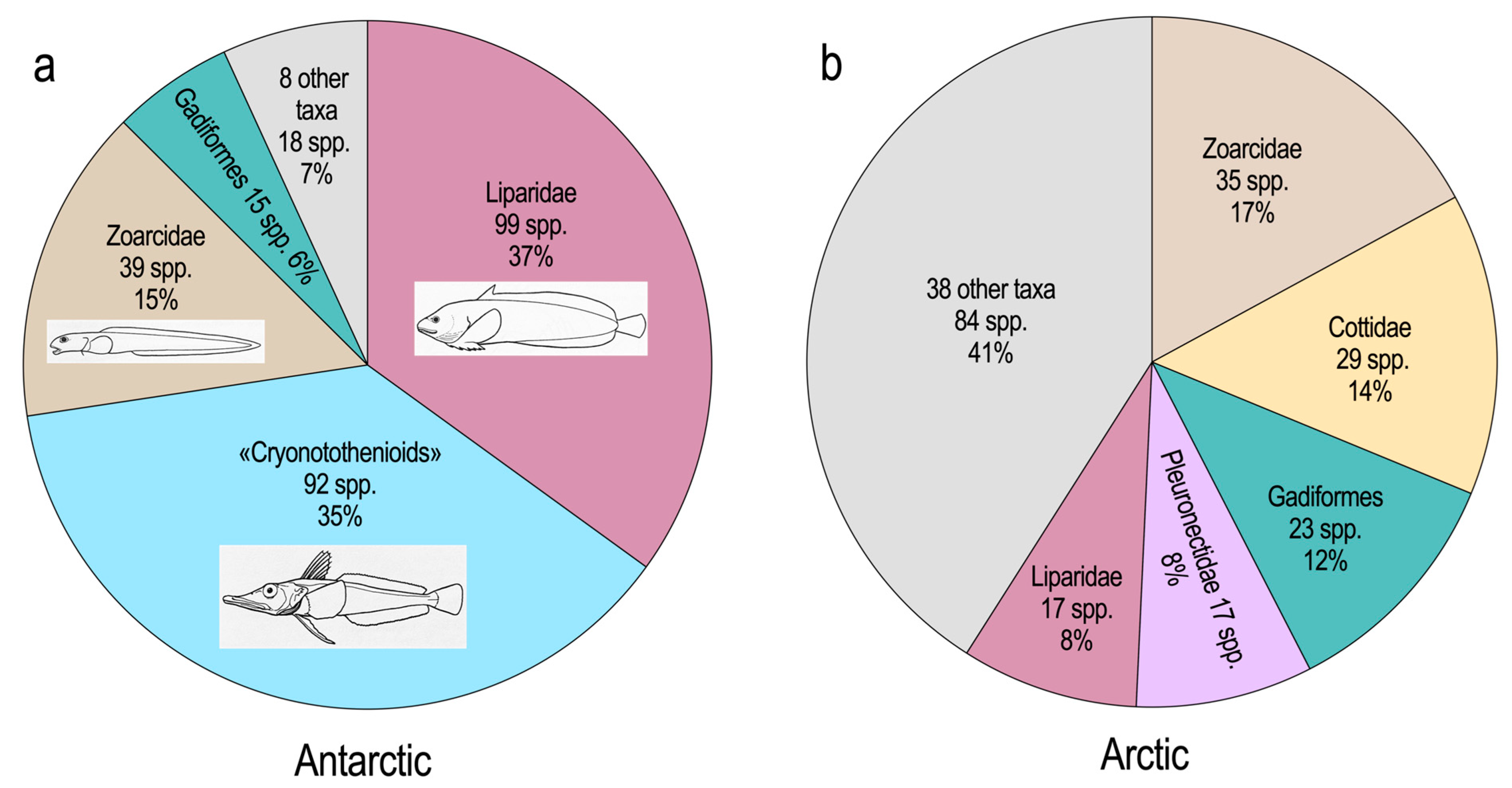{kind=link}
Table 1.
Densities of biomaterials contributing to the hydrostatic balance in the bodies of fishes a,b.
Table 1.
Densities of biomaterials contributing to the hydrostatic balance in the bodies of fishes a,b.
| Substance | Density (kg m–3) |
|---|---|
| Hydroxyapatite, Ca10(PO4)6(OH)2 | 3190 |
| (Mineral component of bone and ) | |
| Calcium carbonate, CaCO3 | 2930 |
| (Mineral component of otoliths) | |
| Bone | 1300–2000 |
| Cartilage | 1060–1180 |
| Muscle | 1040–1080 |
| Viscera | 1040 |
| Gelatinous tissue c | 1025–1050 |
| Notochordal fluid | Unknown d |
| Seawater | 1026 e |
| Freshwater | 1000 |
| Triacylglycerols | 930 |
| Wax esters | 860 |
| Squalene | 860 |
| Gases | ≈1 |
a Davenport and Kjørsvik (1986) [101], Alexander (2003) [102] and Pelster (1997, 2009) [103,104]. b Bodies of most fishes are 60–70% water by weight. c This is a subdermal loose connective tissue consisting primarily of greatly hydrated polysaccharides, especially glycosaminoglycans. d Density has not been determined; the osmolarity of this fluid in coelacanths is 1058 mOsmol (Griffith et al., 1975, Griffith, 1980) [105,106]. e The density of subzero Antarctic seawater is slightly increased from this value at the level of the fourth digit, but the effect on the static buoyancy of notothenioids is minimal.
Table 2.
Ocular vascular structures in notothenioid families a,b.
| Falciform Process d | ||||
|---|---|---|---|---|
| Persistent | ||||
| Choroid | Choroid | Lentiform | ||
| Rete c | Fissure | Dreiecke e | Body f | |
| Percophidae | + | ? | ? | ? |
| Bovichtidae (4 spp.) | + | + | + | + |
| Pseudaphritidae | + | – | – | + |
| Eleginopidae | + | – | – | + |
| Nototheniidae (33 spp.) | + or – | – | – | – or + |
| Harpagiferidae (2 spp.) | – | – | – | – |
| Artedidraconidae (13 spp.) | ± or – | – | – | – |
| Bathydraconidae (13 spp.) | –, ± or + g | – | – | – |
| Channichthyidae (15 spp.) | – | – | – | – |
a Based on Eastman (2006, Table 1) [260], with recent additions representing (for the choroid rete) 242 specimens and 83 of 140 species from 43 of the 45 genera (Eastman, unpublished). b Degree of development: +, present; ±, vestigial; –, absent; ?, unknown. c Vestigial choroid retia are sometimes not macroscopically visible; their presence must be ascertained histologically. d The falciform process is a ridge of pigmented choroidal tissue containing branches of the hyaloid artery. It protrudes into the vitreous cavity through the open embryonic choroid fissure and conveys blood vessels and nerves to the retractor lentis. e This translates from German as “triangles” and the structure is also known as the Triangle of Virchow. The dreiecke is the small anterior-most portion of the falciform process. It consists of smooth muscle, nerves, and blood vessels and is roughly triangular in shape. f The lentiform body is a small vascular bundle situated ventral to the optic nerve at the back of the eye. Although it functions as a rete in some fishes, when present in notothenioids the vessels are relatively large and the existence of capillaries and a countercurrent function are undocumented. When present, the efferent vessels supply the falciform process and retractor lentis. g Cygnodraco mawsoni has a small choroid rete.
Table 3.
Habitat, degree of development of ocular vasculature and retinal thickness, and cell number in notothenioid families based on the sample of 83 specimens in Table 2.
Table 3.
Habitat, degree of development of ocular vasculature and retinal thickness, and cell number in notothenioid families based on the sample of 83 specimens in Table 2.
| Primary Habitat a | Choroid Rete b | Hyaloid Arteries c | Retinal Thickness (µm) d | Number of Retinal Cells e | |
|---|---|---|---|---|---|
| Percophidae | N | ++ | ? | ? | ? |
| Bovichtidae | N | ++ | + | 221–241 | 342–474 |
| Pseudaphritidae | N | ++ | ? | 288 | ? |
| Eleginopidae | N | ++ | – | 272 | 314 |
| Nototheniidae | A, S, N | ± (most) | + or ++ | 150–265 | 106–314 |
| + (some) | (most 110–200) | ||||
| – (few) | |||||
| Harpagiferidae | S | – | ? | ? | ? |
| Artedidraconidae | A | ± or – | ? | 143–225 | 78–79 |
| Bathydraconidae | A | – (most) | ++ or +++ | 132–202 | 98–184 |
| ± (some) | |||||
| + (one) | |||||
| Channichthyidae | A | – | +++ | 166–211 | 99–172 |
a Habitat: N, non-Antarctic; S, sub-Antarctic; A, Antarctic. b Degree of choroid rete development: ++, present and large; +, present and small; ±, vestigial; –, absent; ?, unknown. c Degree of pre-retinal hyaloid arterial development: +++, dense; ++, intermediate; +, sparse. d Obtained from histological sections; thickness excludes optic nerve fiber layer. e Minimum to maximum range for species in each family, with original counts obtained from one Bodian-stained histological section viewed at ×1000; counts are mean number of nuclei (for three replicates) in the entire thickness of the retina (Bruch’s membrane to internal limiting membrane) along a linear extent of 100 µm.
Table 4.
Higher level taxonomic composition of 63 fish species represented by teeth, skeletal elements, scales, or otoliths in the Eocene La Meseta fossil fish fauna from Seymour Island, Antarctica a.
Table 4.
Higher level taxonomic composition of 63 fish species represented by teeth, skeletal elements, scales, or otoliths in the Eocene La Meseta fossil fish fauna from Seymour Island, Antarctica a.
| Taxa | |
|---|---|
| Chondrichthyes (39 species) | Actinopterygii (continued) |
| Callorhinchidae—plownose chimeras | * Notothenioidei – notothenioids |
| Chimaeridae—shortnose chimaeras | †? Proeleginops grandeastmanorum |
| Cetorhinidae—basking sharks | Gadiforms—cods |
| Lamnidae—mackerel sharks | * Moridae—deepsea cods |
| Trikaidae—hound sharks | * Macrouridae—grenadiers |
| Carcharhinidae—requiem sharks | Merlucciidae—hakes |
| Hexanchidae—cow sharks | * Gadidae—cods |
| Stegostomatiidae—zebra sharks | †? Mesetaichthys jerzmanskae |
| Ginglymostomatidae—nurse sharks | Berycidae—alfonsinos |
| Odontaspididae—sand tigers | Trachichthyidae—roughies |
| Squalidae—dogfish sharks | Haemulidae—grunts |
| Squatinidae—angel sharks | Sparidae—porgies |
| Pristiophoridae—saw sharks | Cepolidae—bandfishes |
| Pristidae—sawfishes | Oplegnathidae—knife fishes |
| * Rajidae—skates | Labridae—wrasses |
| Myliobatidae—eagle rays | Trichiuridae—cutlasses fishes |
| † Xiphiorhynchidae—billfishes | |
| Actinopterygii (24 species) | # Pleuronectiformes—flatfishes |
| Clupeidae—herrings | Percoidei, indeterminate |
| Argentinidae—argentines | |
| Paraulopidae—cucumber fishes | |
| Myctophidae—lanternfishes | |
| Siluriforms—catfishes | |
| Ophidiidae—cusk-eels |
a From Schwarzhans et al. (2017, Table 1) [280] and Eastman (2005, Figure 1) [19], with additions from recent the literature cited in the text. *, Represented in modern high latitude Antarctic shelf and upper slope fauna. †, Extinct taxon. ?, Uncertain taxonomic affinity; there is no credible evidence that these are notothenioids (see text). #, Known only from scales.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the author. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Eastman, J.T. The Axes of Divergence for the Evolutionary Radiation of Notothenioid Fishes in Antarctica. Diversity 2024, 16, 214. https://doi.org/10.3390/d16040214
AMA Style
Eastman JT. The Axes of Divergence for the Evolutionary Radiation of Notothenioid Fishes in Antarctica. Diversity. 2024; 16(4):214. https://doi.org/10.3390/d16040214
Chicago/Turabian StyleEastman, Joseph T. 2024. "The Axes of Divergence for the Evolutionary Radiation of Notothenioid Fishes in Antarctica" Diversity 16, no. 4: 214. https://doi.org/10.3390/d16040214
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.