
First Use of Free-Diving Photo-Identification of Porbeagle Shark (Lamna nasus) off the Brittany Coast, France



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
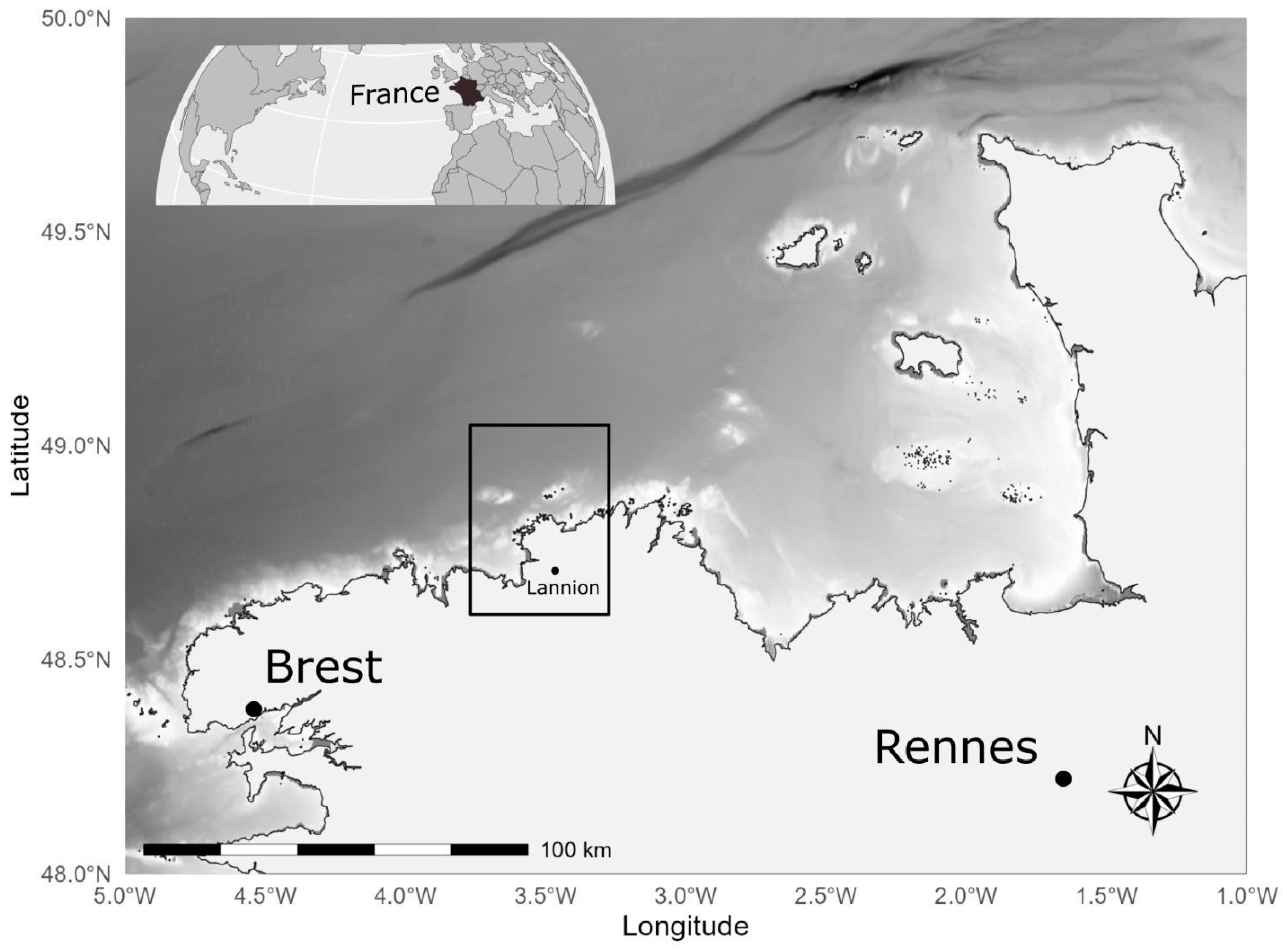
2.1. Field Sampling and Underwater Observations
2.2. Data from Maritime Stakeholders
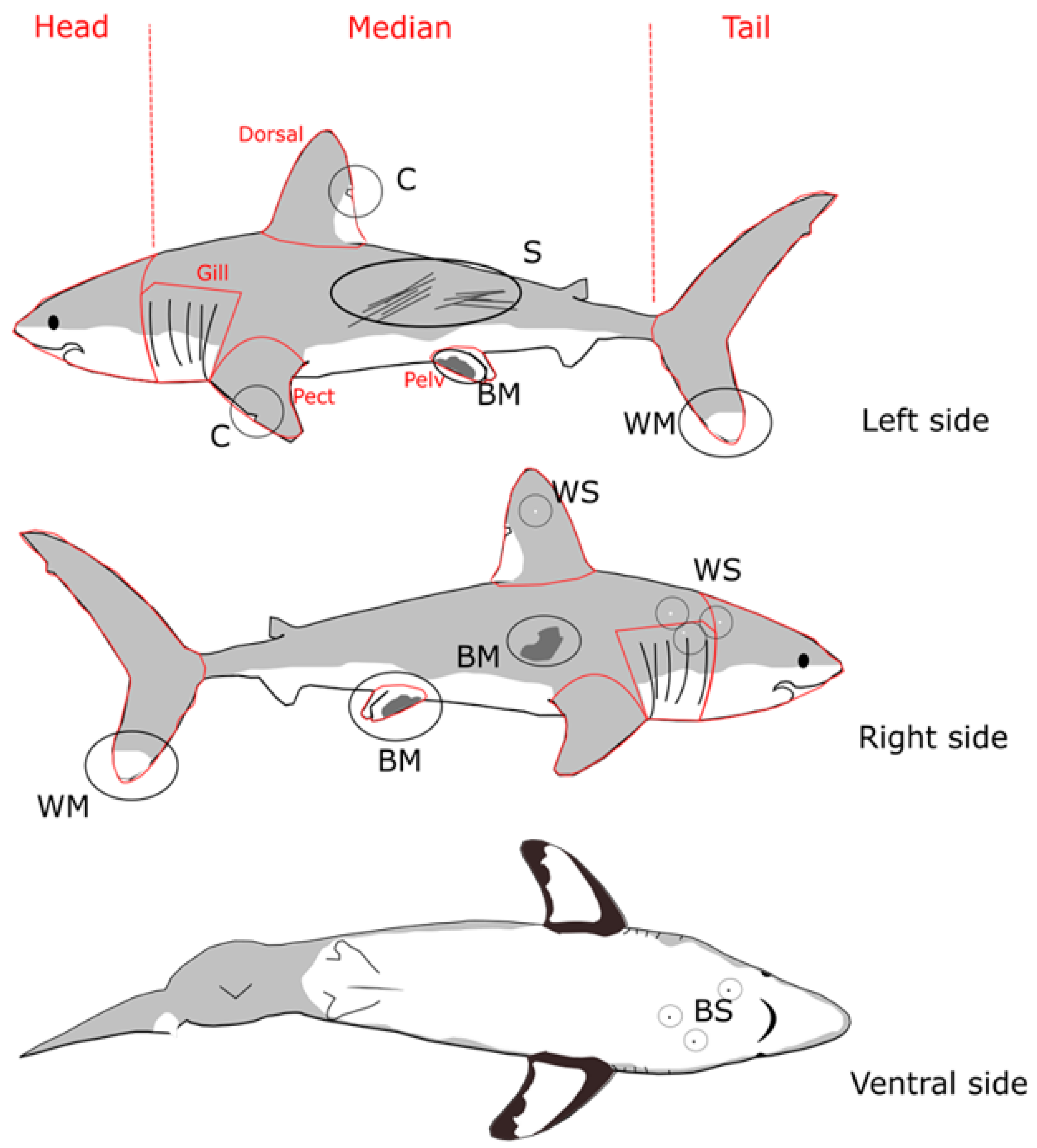
2.3. Phenotypic Markers
2.4. Additional Data
2.5. Data Treatment and Analysis
3. Results
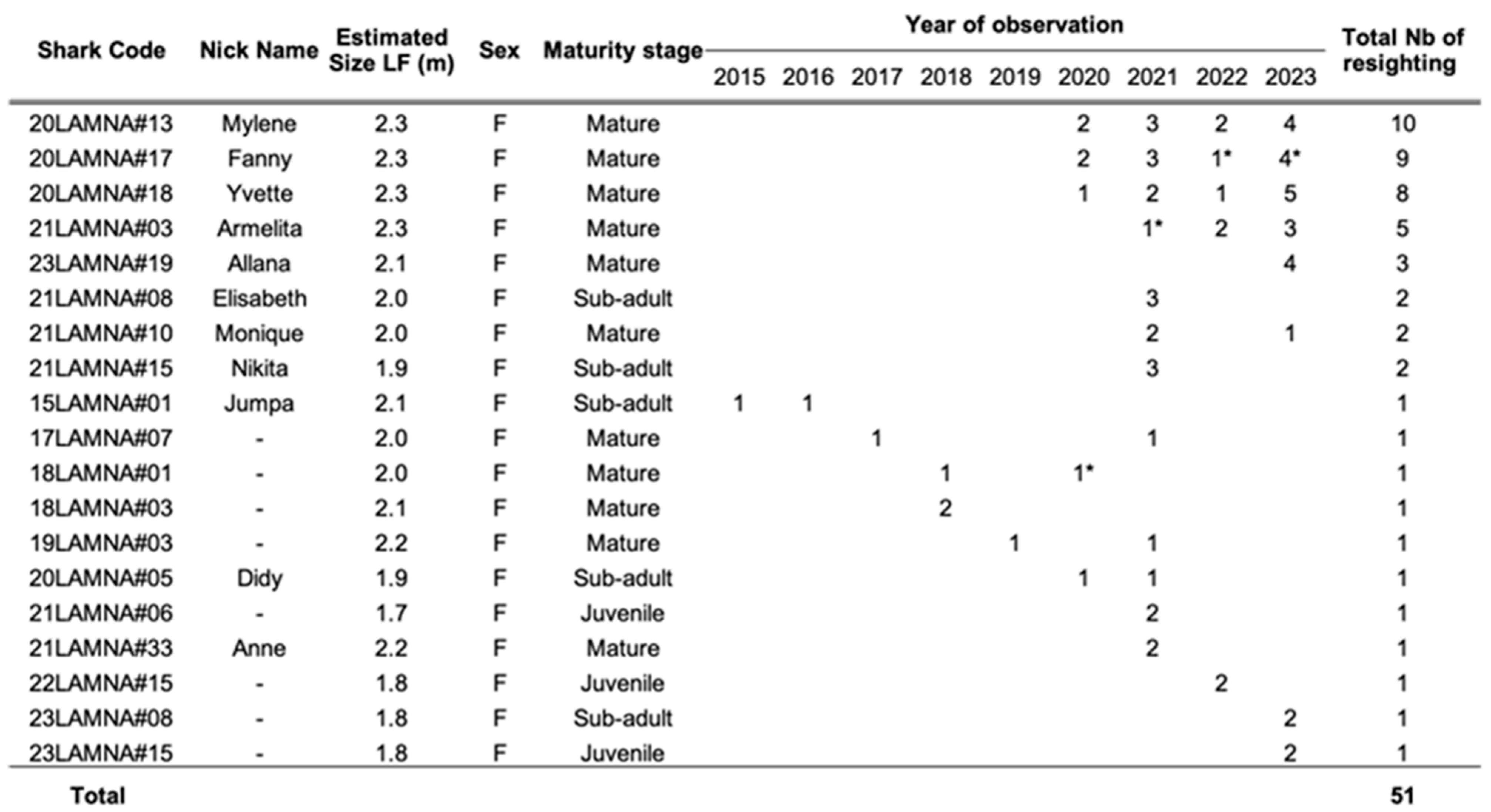
3.1. Porbeagle Photo-ID Catalog
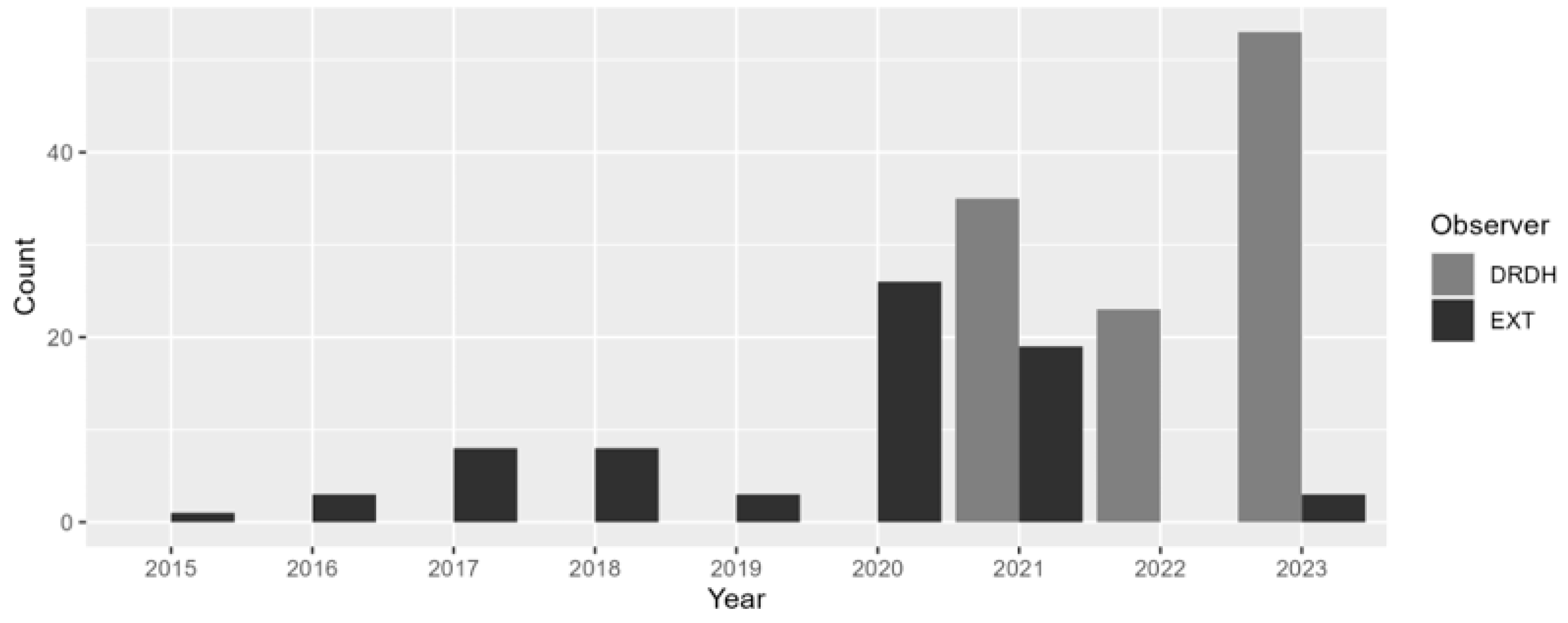
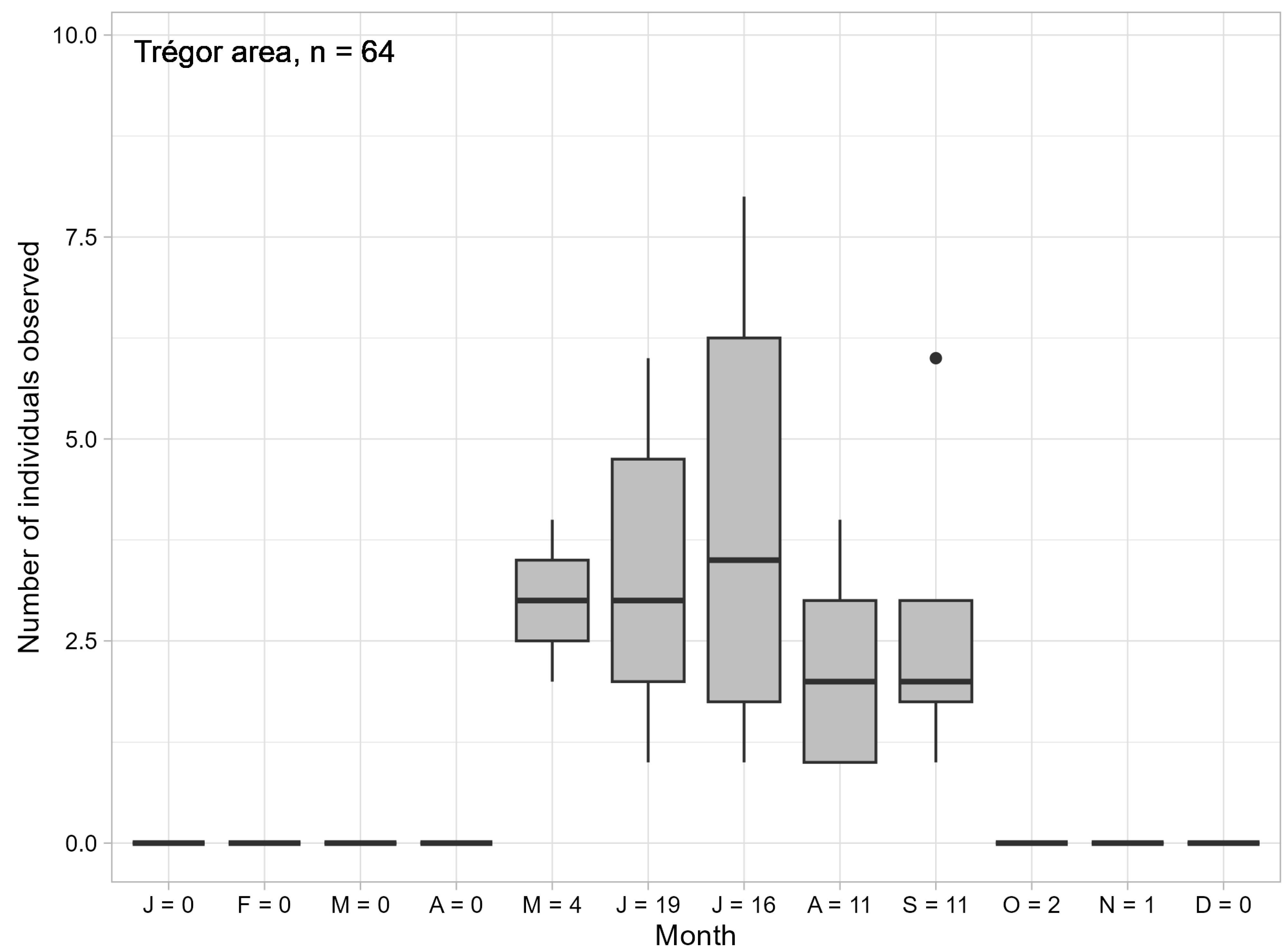
3.2. Seasonal Variations
3.3. Re-Sighting Rate and Site Fidelity
3.4. Biological Information
4. Discussion
4.1. Effectiveness of the Methodology
4.2. Ecology
4.3. Caveats and Limitations
4.4. Perspectives
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Appendix A


References
- Francis, M.P.; Natanson, L.J.; Campana, S.E. The biology and ecology of the porbeagle shark, Lamna nasus. In Sharks of the Open Ocean: Biology, Fisheries and Conservation; Blackwell Publishing: Oxford, UK, 2008; Chapter 9; pp. 105–113. [Google Scholar] [CrossRef]
- Pade, N.G.; Queiroz, N.; Humphries, N.E.; Witt, M.J.; Jones, C.S.; Noble, L.R.; Sims, D.W. First results from satellite-linked archival tagging of porbeagle shark, Lamna nasus: Area fidelity, wider-scale movements and plasticity in diel depth changes. J. Exp. Mar. Biol. Ecol. 2009, 370, 64–74. [Google Scholar] [CrossRef]
- Biais, G.; Coupeau, Y.; Séret, B.; Calmettes, B.; Lopez, R.; Hetherington, S.; Righton, D. Return migration patterns of porbeagle shark (Lamna nasus) in the Northeast Atlantic: Implications for stock range and structure. ICES J. Mar. Sci. 2017, 74, 1268–1276. [Google Scholar] [CrossRef]
- Stevens, J.D. Stomach contents of the blue shark (Prionace glauca L.) off south-west England. J. Mar. Biol. Assoc. U. K. 1973, 53, 357–361. [Google Scholar] [CrossRef]
- Ellis, J.R.; Shackley, S.E. Notes on porbeagle sharks, Lamna nasus, from the Bristol Channel. J. Fish Biol. 1995, 46, 368–370. [Google Scholar] [CrossRef]
- Belleggia, M.; Colonello, J.; Cortés, F.; Figueroa, D.E. Eating catch of the day: The diet of porbeagle shark Lamna nasus (Bonnaterre 1788) based on stomach content analysis, and the interaction with trawl fisheries in the south-western Atlantic (52° S–56° S). J. Fish Biol. 2021, 99, 1591–1601. [Google Scholar] [CrossRef]
- Campana, S.E.; Joyce, W.; Marks, L.; Hurley, P.; Natanson, L.J.; Kohler, N.E.; Jensen, C.F.; Mello, J.J.; Pratt, H.L.J.; Myklevoll, S.; et al. The rise and fall (again) of the porbeagle shark population in the northwest atlantic. In Sharks of the Open Ocean: Biology, Fisheries & Conservation; Camhi, M.D., Pikitch, E.K., Babcock, E.A., Eds.; Blackwell Publishing: Oxford, UK, 2008; pp. 445–461. [Google Scholar]
- Porbeagle (Lamna nasus) in the Northeast Atlantic; Report of the ICES Advisory Committee; ICES: Copenhagen, Denmark, 2010; Volume 9, pp. 85–93.
- Stevens, J.D.; Fowler, S.L.; Soldo, A.; McCord, M.; Baum, J.; Acuña, E.; Domingo, A.; Francis, M. Lamna nasus. The IUCN Red List of Threatened Species. 2006. Available online: https://www.iucnredlist.org/species/11200/3261697 (accessed on 10 October 2023).
- Ellis, J.; Farrell, E.; Jung, A.; McCully, S.; Sims, D.; Soldo, A. Lamna nasus (Europe assessment). The IUCN Red List of Threatened Species 2015: E.T11200A48916453. Available online: https://www.iucnredlist.org/species/11200/48916453 (accessed on 11 December 2023).
- Council Regulation (EU). Council Regulation (EU) No 23/2010 of 14 January 2010 fixing for 2010 the fishing opportunities for certain fish stocks and groups of fish stocks, applicable in EU waters and, for EU vessels, in waters where catch limitations are required and amending Regulations (EC) No 1359/2008, (EC) No 754/2009, (EC) No 1226/2009, and (EC) No 1287/2009. Off. J. Eur. Union 2010, L21, 1–120. [Google Scholar]
- Estes, R.D. A Guide to Watching African Mammals: Including Hoofed Mammals, Carnivores, and Primates; Chelsea Green Publishing: White River Junction, VE, USA, 1993. [Google Scholar]
- Karanth, K.U.; Nichols, J.D. Estimation of tiger densities in India using photographic captures and recaptures. Ecology 1998, 79, 2852–2862. [Google Scholar] [CrossRef]
- Aschettino, J.M.; Baird, R.W.; McSweeney, D.J.; Webster, D.L.; Schorr, G.S.; Huggins, J.L.; Martien, K.K.; Mahaffy, S.D.; West, K.L. Population structure of melon-headed whales (Peponocephala electra) in the Hawaiian Archipelago: Evidence of multiple populations based on photo-identification. Mar. Mammal Sci. 2012, 28, 666–689. [Google Scholar] [CrossRef]
- Wursig, B.; Jefferson, T.A. Methods of photo-identification for small cetaceans. In Individual Recognition of Cetaceans: Use of Photo-Identification and Other Techniques to Estimate Population Parameters; Hammond, P.S., Mizroch, S.A., Donovan, G.P., Eds.; Reports of the International Whaling Commission, Special Issue 12; International Whaling Commission: Cambridge, UK, 1990; pp. 43–52. [Google Scholar]
- Vincent, C.; Meynier, L.; Ridoux, V. Photo-identification in grey seals: Legibility and stability of natural markings. Mammalia 2001, 65, 363–372. [Google Scholar] [CrossRef]
- Mazzoil, M.; McCulloch, S.D.; Defran, R.H.; Murdoch, M.E. Use of digital photography and analysis of dorsal fins for photo-identification of bottlenose dolphins. Aquat. Mamm. 2004, 30, 209–219. [Google Scholar] [CrossRef]
- Taylor, G. Whale Sharks, the Giants of Ningaloo Reef; Angus & Robertson: Sydney, NSW, Australia, 1994; ISBN 0207184984/13. [Google Scholar]
- Corcoran, M.J.; Gruber, S.H. The use of photo-identification to study the social organization of the spotted eagle ray (Aetobatus narinari). Bahamas J. Sci. 1999, 11, 21–27. [Google Scholar]
- Gore, M.A.; Frey, P.H.; Ormond, R.F.; Allan, H.; Gilkes, G. Use of Photo-Identification and Mark-Recapture Methodology to Assess Basking Shark (Cetorhinus maximus) Populations. PLoS ONE 2016, 11, e0150160. [Google Scholar] [CrossRef]
- Graham, R.; Roberts, C.M. Assessing the size, growth, and structure of a seasonal population of whale sharks (Rhincodon typus, Smith 1828) using conventional tagging and photo identification. Fish. Res. 2007, 84, 71–80. [Google Scholar] [CrossRef]
- Couturier, L.I.E.; Jaine, F.R.A.; Townsend, K.A.; Weeks, S.J.; Richardson, A.J.; Bennett, M.B. Distribution, site affinity and regional movements of the manta ray, Manta alfredi (Kreft, 1868), along the east coast of Australia. Mar. Freshw. Res. 2011, 62, 628–637. [Google Scholar] [CrossRef]
- Marshall, A.D.; Pierce, S.J. The use and abuse of photographic identification in sharks and rays. J. Fish Biol. 2012, 80, 1361–1379. [Google Scholar] [CrossRef]
- Dudgeon, C.L.; Noad, M.J.; Lanyon, J.M. Abundance and demography of a seasonal aggregation of zebra sharks Stegostoma fasciatum. Mar. Ecol. Prog. Ser. 2008, 368, 269–281. [Google Scholar] [CrossRef]
- Hewitt, A.M.; Kock, A.A.; Booth, A.J.; Griffiths, C.L. Trends in sightings and population structure of white sharks, Carcharodon carcharias, at Seal Island, False Bay, South Africa, and the emigration of subadult female sharks approaching maturity. Environ. Biol. Fishes 2018, 101, 39–54. [Google Scholar] [CrossRef]
- Bettcher, V.B.; Santos, L.N.; Bertoncini, Á.A.; Silva, M.B.; Castro, A.L. Evidence of the Atlantic nurse shark (Ginglymostoma cirratum) population shrink at Rocas Atoll, Southwestern Atlantic. Aquat. Conserv. Mar. Freshw. Ecosyst. 2023, 33, 845–852. [Google Scholar] [CrossRef]
- Castro, A.L.; Rosa, R.S. Use of natural marks on population estimates of the nurse shark, Ginglymostoma cirratum, at Atol das Rocas Biological Reserve, Brazil. Environ. Biol. Fishes 2005, 72, 213–221. [Google Scholar] [CrossRef]
- Nakachi, K.I. Heeding the History of Kahu Manō: Developing and Validating a Pono Photo-Identification Methodology for Tiger Sharks (Galeocerdo cuvier) in Hawaii. Ph.D. Thesis, University of Hawaii at Hilo, Hilo, HI, USA, 2021. [Google Scholar]
- Madon, B.; Gimenez, O.; McArdle, B.; Baker, C.S.; Garrigue, C. A New Method For Estimating Animal Abundance with Two Sources of Data in Capture-Recapture Studies. Methods Ecol. Evol. 2011, 2, 390–400. [Google Scholar] [CrossRef]
- Holmberg, J.; Norman, B.; Arzoumanian, Z. Estimating population size, structure, and residency time for whale sharks Rhincodon typus through collaborative photo-identification. Endanger. Species Res. 2009, 7, 39–53. [Google Scholar] [CrossRef]
- Lewis, R.; Dawson, S.; Rayment, W. Estimating population parameters of broadnose sevengill sharks (Notorynchus cepedianus) using photo identification capture-recapture. J. Fish Biol. 2020, 97, 987–995. [Google Scholar] [CrossRef]
- Porbeagle (Lamna nasus) in Subareas 1–10, 12, and 14 (the Northeast Atlantic and Adjacent Waters); Report of the ICES Advisory Committee; ICES: Copenhagen, Denmark, 2022. [CrossRef]
- Le Person, A. La Pêche au Gros en Baie de Lannion: Thons Rouges et Requins; Embanner, Y., Ed.; Embanner: Fouesnant, France, 2013; 336p, ISBN 978-2916579573. [Google Scholar]
- Siorat, F. L’évolution de la colonie de Fous de Bassan, Morus bassanus, de l’île Rouzic de 1939 à 1996. Le Cormoran 1998, 47, 173–175. [Google Scholar]
- Chauvaud, S.; Jean, F. Inventaire Patrimonial de la Macrofaune de L’Estran de la Réserve Naturelle des Sept-îles; Rapport C.R.O.E.M.I, CROEMI-UBO, Université de Bretagne Occidentale: Brest, France, 1999; 22p. [Google Scholar]
- Siorat, F.; Bentz, G. Réserve Naturelle des Sept-Îles. Activity Report. 2004. Available online: https://www.maia-network.org (accessed on 20 October 2023).
- Martelet, G.; Gloaguen, E.; Døssing, A.; Lima Simoes da Silva, E.; Linde, J.; Rasmussen, T.M. Airborne/UAV Multisensor Surveys Enhance the Geological Mapping and 3D Model of a Pseudo-Skarn Deposit in Ploumanac’h, French Brittany. Minerals 2021, 11, 1259. [Google Scholar] [CrossRef]
- Kalmijn, A.J. Electric and magnetic sensory world of sharks, skates, and rays. In Sensory Biology of Sharks, Skates, and Rays; Hodgson, E.S., Mathewson, R., Eds.; Office of Naval Research, Department of the Navy: Arlington, VA, USA, 1978; pp. 507–528. [Google Scholar]
- Tricas, T.C. The neuroecology of the elasmobranch electrosensory world: Why peripheral morphology shapes behavior. In The Behavior and Sensory Biology of Elasmobranch Fishes: An Anthology in Memory of Donald Richard Nelson; Tricas, T.C., Gruber, S.H., Eds.; Springer: Dordrecht, The Netherlands, 2001; Volume 20. [Google Scholar] [CrossRef]
- Hutchison, Z.; Sigray, P.; He, H.; Gill, A.B.; King, J.; Gibson, C. Electromagnetic Field (EMF) Impacts on Elasmobranch (Shark, Rays, and Skates) and American Lobster Movement and Migration from Direct Current Cables; US Department of the Interior, Bureau of Ocean Energy Management: Sterling, VA, USA, 2018. [Google Scholar] [CrossRef]
- Serre, S.; Jung, A.; Cherel, Y.; Gamblin, C.; Hennache, C.; Le Loc’h, F.; Lorrain, A.; Priac, A.; Schaal, G.; Stephan, E. Stable isotopes reveal intrapopulation heterogeneity of porbeagle shark (Lamna nasus). Reg. Stud. Mar. Sci. 2024, 69, 103340. [Google Scholar] [CrossRef]
- Domeier, M.L.; Nasby-Lucas, N. Annual re-sightings of photographically identified white sharks (Carcharodon carcharias) at an eastern Pacific aggregation site (Guadalupe Island, Mexico). Mar. Biol. 2007, 150, 977–984. [Google Scholar] [CrossRef]
- Anderson, S.D.; Chapple, T.K.; Jorgensen, S.J.; Klimley, A.P.; Block, B.A. Long-term individual identification and site fidelity of white sharks, Carcharodon carcharias, off California using dorsal fins. Mar. Biol. 2011, 158, 1233–1237. [Google Scholar] [CrossRef]
- Jensen, C.F.; Natanson, L.J.; Pratt, H.L., Jr.; Kohler, N.E.; Campana, S.E. The reproductive biology of the porbeagle shark (Lamna nasus) in the western North Atlantic Ocean. Fish. Bull. 2002, 100, 727–738. [Google Scholar]
- Dureuil, M.; Towner, A.V.; Ciolfi, L.G.; Beck, L.A. A computer-aided framework for subsurface identification of white shark pigment patterns. Afr. J. Mar. Sci. 2015, 37, 363–371. [Google Scholar] [CrossRef]
- Jung, A. A Preliminary Assessment of the French Fishery Targeted Porbeagle Shark (Lamna nasus) in the Northeast Atlantic Ocean: Biology and Catch Statistics; Standing Committee on Research and Statistics, ICCAT: Madrid, Spain, 2008; p. 11. [Google Scholar]
- Saunders, R.A.; Royer, F.; Clarke, M.W. Winter migration and diving behaviour of porbeagle shark, Lamna nasus, in the Northeast Atlantic. ICES J. Mar. Sci. 2011, 68, 166–174. [Google Scholar] [CrossRef]
- Francis, M.P.; Stevens, J.D. Reproduction, embryonic development, and growth of the porbeagle shark, Lamna nasus, in the southwest Pacific Ocean. Fish. Bull. 2000, 98, 41–63. [Google Scholar]
- Penfold, L.M.; Wyffels, J.T. Reproductive science in sharks and rays. In Reproductive Sciences in Animal Conservation, Advances in Experimental Medicine and Biology; Comizzoli, P., Brown, J., Holt, W., Eds.; Springer: Cham, Switzerland, 2019; Volume 1200. [Google Scholar] [CrossRef]
- Anderson, B.; Belcher, C.; Slack, J.A.; Gelsleichter, J. Evaluation of the use of portable ultrasonography to determine pregnancy status and fecundity in bonnethead shark Sphyrna tiburo. J. Fish Biol. 2018, 93, 1163–1170. [Google Scholar] [CrossRef]
- Barnett, A.; Abrantes, K.G.; Stevens, J.D.; Semmens, J.M. Site fidelity and sex-specific migration in a mobile apex predator: Implications for conservation and ecosystem dynamics. Anim. Behav. 2011, 81, 1039–1048. [Google Scholar] [CrossRef]
- Domeier, M.L. Global Perspectives on the Biology and Life History of the White Shark; CRC Press: Boca Raton, FL, USA, 2012. [Google Scholar]
- Pillans, R.D.; Rochester, W.; Babcock, R.C.; Thomson, D.P.; Haywood, M.D.E. Long-Term Acoustic Monitoring Reveals Site Fidelity, Reproductive Migrations, and Sex Specific Differences in Habitat Use and Migratory Timing in a Large Coastal Shark (Negaprion acutidens). Front. Mar. Sci. 2021, 8, 616633. [Google Scholar] [CrossRef]
- Hulbert, L.B.; Aires-da-Silva, A.M.; Gallucci, V.F.; Rice, J.S. Seasonal foraging movements and migratory patterns of female Lamna ditropis tagged in Prince William Sound, Alaska. J. Fish Biol. 2005, 67, 490–509. [Google Scholar] [CrossRef]
- Druon, J.-N.; Campana, S.; Vandeperre, F.; Hazin, F.H.V.; Bowlby, H.; Coelho, R.; Queiroz, N.; Serena, F.; Abascal, F.; Damalas, D.; et al. Global-Scale Environmental Niche and Habitat of Blue Shark (Prionace glauca) by Size and Sex: A Pivotal Step to Improving Stock Management. Front. Mar. Sci. 2022, 9, 828412. [Google Scholar] [CrossRef]
- Hennache, C.; Jung, A. Etude de la Pêcherie Palangrière de Requin Taupe de L’île D’Yeu. Brest, France: Association Pour L’étude et la Conservation des Sélaciens (APECS). 2010. Available online: https://www.researchgate.net/profile/Armelle-Jung/publication/313667107_Etude_de_la_pecherie_palangriere_de_requin_taupe_Lamna_nasus_de_l’Ile_d’Yeu_EPPARTIY/links/58a2315daca272046aafe5f1/Etude-de-la-pecherie-palangriere-de-requin-taupe-Lamna-nasus-de-lIle-dYeu-EPPARTIY.pdf (accessed on 20 April 2021).
- Bradford, R.; Patterson, T.A.; Rogers, P.J.; McAuley, R.; Mountford, S.; Huveneers, C.; Robbins, R.; Fox, A.; Bruce, B.D. Evidence of diverse movement strategies and habitat use by white sharks, Carcharodon carcharias, off southern Australia. Mar. Biol. 2020, 167, 96. [Google Scholar] [CrossRef]
- Barker, S.M.; Williamson, J.E. Collaborative photo-identification and monitoring of grey nurse sharks (Carcharias taurus) at key aggregation sites along the eastern coast of Australia. Mar. Freshw. Res. 2010, 61, 971–979. [Google Scholar] [CrossRef]
- Van Tienhoven, A.M.; Den Hartog, J.E.; Reijns, R.A.; Peddemors, V.M. A computer-aided program for pattern-matching of natural marks on the spotted raggedtooth shark Carcharias taurus. J. Appl. Ecol. 2007, 44, 273–280. [Google Scholar] [CrossRef]
- Sarano, F.; Girardet, J.; Sarano, V.; Vitry, H.; Preud’Homme, A.; Heuzey, R.; Garcia-Cegarra, A.M.; Madon, B.; Delfour, F.; Glotin, H.; et al. Kin relationships in cultural species of the marine realm: Case study of a matrilineal social group of sperm whales off Mauritius island, Indian Ocean. R. Soc. Open Sci. 2021, 8, 201794. [Google Scholar] [CrossRef]
- González, M.T.; Sepúlveda, F.A.; Zárate, P.M.; Baeza, J.A. Regional population genetics and global phylogeography of the endangered highly migratory shark Lamna nasus: Implications for fishery management and conservation. Aquat. Conserv. Mar. Freshw. Ecosyst. 2021, 31, 620–634. [Google Scholar] [CrossRef]
- Madon, B.; Le Guyader, D.; Jung, J.-L.; De Montgolfier, B.; Lopez, P.J.; Foulquier, E.; Bouveret, L.; Le Berre, I. Pairing AIS data and underwater topography to assess maritime traffic pressures on cetaceans: Case study in the Guadeloupean waters of the Agoa sanctuary. Mar. Policy 2022, 143, 105160. [Google Scholar] [CrossRef]
- Séguigne, C.; Mourier, J.; Clua, É.; Buray, N.; Planes, S. Citizen science provides valuable data to evaluate elasmobranch diversity and trends throughout the French Polynesia’s shark sanctuary. PLoS ONE 2023, 18, e0282837. [Google Scholar] [CrossRef] [PubMed]
- Coché, L.; Arnaud, E.; Bouveret, L.; David, R.; Foulquier, E.; Gandilhon, N.; Jeannesson, E.; Le Bras, Y.; Lerigoleur, E.; Lopez, P.J.; et al. Kakila database: Towards a FAIR community approved database of cetacean presence in the waters of the Guadeloupe Archipelago, based on citizen science. Biodivers. Data J. 2021, 9, e69022. [Google Scholar] [CrossRef] [PubMed]







Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Jung, A.; Ory, A.; Abaut, P.; Zaccagnini, L. First Use of Free-Diving Photo-Identification of Porbeagle Shark (Lamna nasus) off the Brittany Coast, France. Diversity 2024, 16, 155. https://doi.org/10.3390/d16030155
Jung A, Ory A, Abaut P, Zaccagnini L. First Use of Free-Diving Photo-Identification of Porbeagle Shark (Lamna nasus) off the Brittany Coast, France. Diversity. 2024; 16(3):155. https://doi.org/10.3390/d16030155
Chicago/Turabian StyleJung, Armelle, Arthur Ory, Paul Abaut, and Lucas Zaccagnini. 2024. "First Use of Free-Diving Photo-Identification of Porbeagle Shark (Lamna nasus) off the Brittany Coast, France" Diversity 16, no. 3: 155. https://doi.org/10.3390/d16030155