
Detection of Glacial Refugia and Post-Glacial Colonization Routes of Morphologically Cryptic Marsh Frog Species (Anura: Ranidae: Pelophylax) Using Environmental Niche Modeling


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
3. Results
3.1. Current Distribution of Water Frog Species
3.2. Distribution Modeling of Species under the Current Environmental Conditions
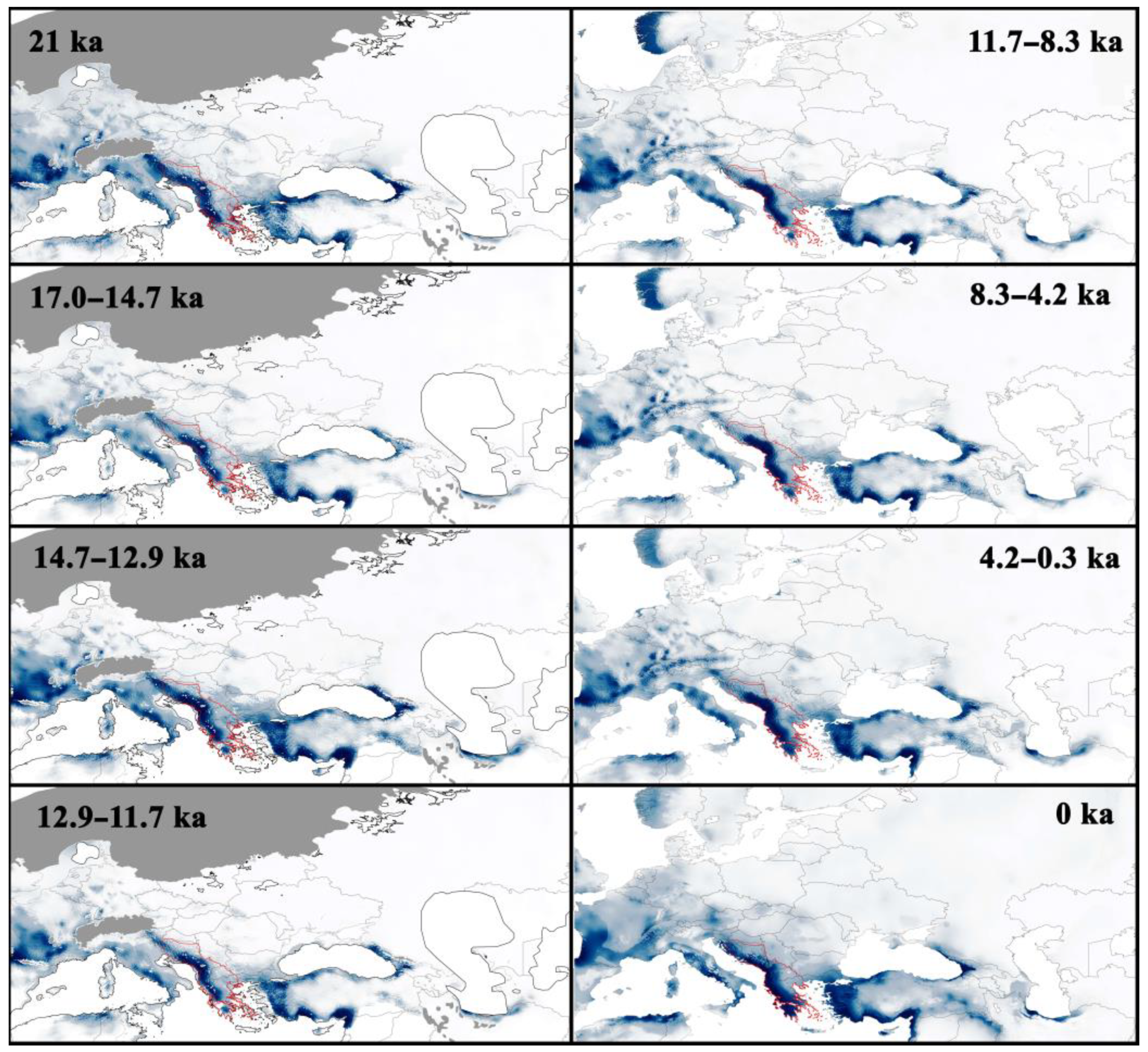
3.3. Distribution of Species at the Late-Quaternary
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Conflicts of Interest
References
- Wiens, J.J. Speciation and ecology revisited: Phylogenetic niche conservatism and the origin of species. Evolution 2004, 58, 193–197. [Google Scholar] [CrossRef] [PubMed]
- Sillero, N.; Barbosa, A.M. Common mistakes in ecological niche models. Int. J. Geogr. Inf. Sci. 2021, 35, 213–226. [Google Scholar] [CrossRef]
- Wiens, J.A.; Stralberg, D.; Jongsomjit, D.; Howell, C.A.; Snyder, M.A. Niches, models, and climate change: Assessing the assumptions and uncertainties. Proc. Natl. Acad. Sci. USA 2009, 106, 19729–19736. [Google Scholar] [CrossRef] [PubMed]
- Elith, J.; Franklin, J. Species Distribution Modeling. Enc. Biodiv. 2013, 6, 692–705. [Google Scholar] [CrossRef]
- Alvarado-Serrano, D.F.; Knowles, L.L. Ecological niche models in phylogeographic studies: Applications, advances and precautions. Mol. Ecol. Res. 2014, 14, 233–248. [Google Scholar] [CrossRef] [PubMed]
- Bartlein, P.J.; Hostetler, S.W. Modeling paleoclimates. Dev. Quart. Sci. 2004, 1, 565–584. [Google Scholar] [CrossRef]
- Sillero, N.; Carretero, M.A. Modelling the past and future distribution of contracting species. The Iberian lizard Podarcis carbonelli (Squamata: Lacertidae) as a case study. Zool. Anz. J. Comp. Zool. 2013, 252, 289–298. [Google Scholar] [CrossRef]
- González-Fernández, A.; Manjarrez, J.; García-Vázquez, U.; D’Addario, M.; Sunny, A. Present and future ecological niche modeling of garter snake species from the Trans-Mexican Volcanic Belt. PeerJ 2018, 6, e4618. [Google Scholar] [CrossRef]
- Gassert, F.; Schulte, U.; Husemann, M.; Ulrich, W.; Rödder, D.; Hochkirch, A.; Engel, E.; Meyer, J.; Habel, J.C. From southern refugia to the northern range margin: Genetic population structure of the common wall lizard, Podarcis muralis. J. Biogeogr. 2013, 40, 1475–1489. [Google Scholar] [CrossRef]
- Dufresnes, C.; Berroneau, M.; Dubey, S.; Litvinchuk, S.N.; Perrin, N. The effect of phylogeographic history on species boundaries: A comparative framework in Hyla tree frogs. Sci. Rep. 2020, 10, 5502. [Google Scholar] [CrossRef]
- Macaluso, L.; Bertini, A.; Carnevale, G.; Eronen, J.T.; Martinetto, E.; Saarinen, J.; Villa, A.; Capasso, F.; Delfino, M. A combined palaeomodelling approach reveals the role as selective refugia of the Mediterranean peninsulas. Palaeogeogr. Palaeoclim. Palaeoecol. 2023, 625, 111699. [Google Scholar] [CrossRef]
- Plötner, J. Die Westpaläarktische Wasserfrösche; Laurenti-Verlag: Bielefeld, Germany, 2005; p. 160. [Google Scholar]
- Sánchez-Montes, G.; Recuero, E.; Gutiérrez-Rodríguez, J.; Gomez-Mestre, I.; Martínez-Solano, I. Species assignment in the Pelophylax ridibundus × P. perezi hybridogenetic complex based on 16 newly characterised microsatellite markers. Herpetol. J. 2016, 26, 99–108. [Google Scholar]
- Lyapkov, S.M.; Ermakov, O.A.; Titov, S.V. Distribution and origin of two forms of the marsh frog Pelophylax ridibundus complex (Anura, Ranidae) from Kamchatka, based on mitochondrial and nuclear DNA data. Zool. Zh. 2017, 96, 1384–1391. (In Russian) [Google Scholar] [CrossRef]
- Jablonski, D.; Wynn, A.; Masroor, R.; Papenfuss, T.; Litvinchuk, S.N.; Mazepa, G. The genus Pelophylax (Amphibia, Ranidae) in Pakistan: Museum collections and possible distribution. Herpetozoa 2021, 34, 141–147. [Google Scholar] [CrossRef]
- Papežík, P.; Sciberras, A.; Benovics, M.; Sciberras, J.; Deidun, A.; Mikulíček, P. Far from Home: Tracing the non-native origin of water frogs (genus Pelophylax) in Malta by molecular markers. Biol. Invasions 2024, preprint. [Google Scholar] [CrossRef]
- Akın, C.; Bilgin, C.C.; Beerli, P.; Westaway, R.; Ohst, T.; Litvinchuk, S.N.; Uzzell, T.; Bilgin, M.; Hotz, H.; Guex, G.-D.; et al. Phylogeographic patterns of genetic diversity in eastern Mediterranean water frogs were determined by geological processes and climate change in the Late Cenozoic. J. Biogeogr. 2010, 37, 2111–2124. [Google Scholar] [CrossRef] [PubMed]
- Plötner, J.; Baier, F.; Akın, C.; Mazepa, G.; Schreiber, R.; Beerli, P.; Litvinchuk, S.N.; Bilgin, C.C.; Borkin, L.; Uzzell, T. Genetic data reveal that water frogs of Cyprus (genus Pelophylax) are an endemic species of Messinian origin. Zoosyst. Evol. 2012, 88, 261–283. [Google Scholar] [CrossRef]
- Bülbül, U.; Matsui, M.; Kutrup, B.; Eto, K. Taxonomic relationships among Turkish water frogs as revealed by phylogenetic analyses using mtDNA gene sequences. Zool. Sci. 2011, 12, 930–936. [Google Scholar] [CrossRef]
- Pesarakloo, A.; Rastegar-Pouyani, E.; Rastegar-Pouyani, N.; Kami, H.; Najibzadeh, M.; Khosravani, A.; Oraie, H. The first taxonomic revaluation of the Iranian water frogs of the genus Pelophylax (Anura: Ranidae) using sequences of the mitochondrial genome. Mitochondrial DNA 2017, 28, 392–398. [Google Scholar] [CrossRef]
- Dufresnes, C.; Litvinchuk, S.N. Diversity, distribution and molecular species delimitation in frogs and toads from the Eastern Palearctic. Zool. J. Linn. Soc. 2022, 195, 695–760. [Google Scholar] [CrossRef]
- Ualiyeva, D.; Ermakov, O.A.; Litvinchuk, S.N.; Guo, X.; Ivanov, A.Y.; Xu, R.; Li, J.; Xu, F.; Arifulova, I.I.; Kaptyonkina, A.G.; et al. Diversity, phylogenetic relationships and distribution of marsh frogs (the Pelophylax ridibundus complex) from Kazakhstan and Northwest China. Diversity 2022, 14, 869. [Google Scholar] [CrossRef]
- Safaei-Mahroo, B.; Ghaffari, H.; Niamir, A. A synoptic review of the Amphibians of Iran: Bibliography, taxonomy, synonymy, distribution, conservation status, and identification key to the eggs, larvae, and adults. Zootaxa 2023, 5279, 1–112. [Google Scholar] [CrossRef]
- Dufresnes, C.; Denoël, M.; Di Santo, L.; Dubey, S. Multiple uprising invasions of Pelophylax water frogs, potentially inducing a new hybridogenetic complex. Sci. Rep. 2017, 7, 6506. [Google Scholar] [CrossRef] [PubMed]
- Dufresnes, C.; Leuenberger, J.; Amrhein, V.; Bühler, C.; Thiébaud, J.; Bohnenstengel, T.; Dubey, S. Invasion genetics of marsh frogs (Pelophylax ridibundus sensu lato) in Switzerland. Biol. J. Linn. Soc. 2018, 123, 402–410. [Google Scholar] [CrossRef]
- Litvinchuk, S.N.; Ivanov, A.Y.; Lukonina, S.A.; Ermakov, O.A. A record of alien Pelophylax species and widespread mitochondrial DNA transfer in Kaliningradskaya Oblast’ (the Baltic coast, Russia). BioInvasions Rec. 2020, 9, 599–617. [Google Scholar] [CrossRef]
- Plötner, J.; Ohst, T. New hypotheses on the systematic of the western Palearctic water frog complex (Anura, Ranidae). Mitt. Mus. Zoosystematics Evol. 2001, 77, 5–21. [Google Scholar] [CrossRef]
- Lymberakis, P.; Poulakakis, N.; Manthalou, G.; Tsigenopoulos, C.S.; Magoulas, A.; Mylonas, M. Mitochondrial phylogeography of Rana (Pelophylax) populations in the Eastern Mediterranean region. Mol. Phylogenet. Evol. 2007, 44, 115–125. [Google Scholar] [CrossRef] [PubMed]
- Hofman, S.; Pabijan, M.; Osikowski, A.; Litvinchuk, S.N.; Szymura, J.M. Phylogenetic relationships among four new complete mitogenome sequences of Pelophylax (Amphibia: Anura) from the Balkans and Cyprus. Mitochondrial DNA Part A 2016, 27, 3434–3437. [Google Scholar] [CrossRef]
- Svinin, A.O.; Dedukh, D.V.; Borkin, L.J.; Ermakov, O.A.; Ivanov, A.Y.; Litvinchuk, J.S.; Zamaletdinov, R.I.; Mikhaylova, R.I.; Trubyanov, A.B.; Skorinov, D.V.; et al. Genetic structure, morphological variation, and gametogenic peculiarities in water frogs (Pelophylax) from northeastern European Russia. J. Zool. Syst. Evol. Res. 2021, 59, 646–662. [Google Scholar] [CrossRef]
- Papežík, P.; Mikulíček, P.; Benovics, M.; Balogová, M.; Choleva, L.; Doležálková-Kaštánková, M.; Lymberakis, P.; Mizsei, E.; Papežíková, S.; Poulakakis, N.; et al. Comparative mitochondrial phylogeography of water frogs (Ranidae: Pelophylax spp.) from the southwestern Balkans. Vert. Zool. 2023, 73, 525–544. [Google Scholar] [CrossRef]
- Dufresnes, C.; Mazepa, G.; Jablonski, D.; Oliveira, R.C.; Wenseleers, T.; Shabanov, D.A.; Auer, M.; Ernst, R.; Koch, C.; Ramírez-Chaves, H.E.; et al. Fifteen shades of green: The evolution of Bufotes toads revisited. Mol. Phylogenet. Evol. 2019, 141, 106615. [Google Scholar] [CrossRef]
- Fick, S.E.; Hijmans, R.J. WorldClim 2: New 1-km spatial resolution climate surfaces for global land areas. Int. J. Climatol. 2017, 37, 4302–4315. [Google Scholar] [CrossRef]
- Brown, J.L.; Hill, D.J.; Dolan, A.M.; Carnaval, A.C.; Haywood, A.M. PaleoClim, high spatial resolution paleoclimate surfaces for global land areas. Sci. Data 2018, 5, 180254. [Google Scholar] [CrossRef] [PubMed]
- Hauswaldt, J.S.; Höer, M.; Ogielska, M.; Christiansen, D.G.; Dziewulska-Szwajkowska, D.; Czernicka, E.; Vences, M. A simplified molecular method for distinguishing among species and ploidy levels in European water frogs (Pelophylax). Mol. Ecol. Res. 2012, 12, 797–805. [Google Scholar] [CrossRef] [PubMed]
- Ermakov, O.; Ivanov, A.; Titov, S.; Svinin, A.; Litvinchuk, S.N. New multiplex PCR method for identification of East European green frog species and their hybrids. Russ. J. Herpetol. 2019, 26, 367–370. [Google Scholar] [CrossRef]
- Wielstra, B.; Arntzen, J.W. Exploring the effect of asymmetric mitochondrial DNA introgression on estimating niche divergence in morphologically cryptic species. PLoS ONE 2014, 9, e95504. [Google Scholar] [CrossRef] [PubMed]
- Toews, D.P.L.; Brelsford, A. The biogeography of mitochondrial and nuclear discordance in animals. Mol. Ecol. 2012, 21, 3907–3930. [Google Scholar] [CrossRef] [PubMed]
- Sequeira, F.; Bessa-Silva, A.; Tarroso, P.; Sousa-Neves, T.; Vallinoto, M.; Gonçalves, H.; Martínez-Solano, I. Discordant patterns of introgression across a narrow hybrid zone between two cryptic lineages of an Iberian endemic newt. J. Evol. Biol. 2019, 33, 202–216. [Google Scholar] [CrossRef] [PubMed]
- Buggs, R.J.A. Empirical study of hybrid zone movement. Heredity 2007, 99, 301–312. [Google Scholar] [CrossRef]
- Wielstra, B.; Burke, T.; Butlin, R.K.; Avcı, A.; Üzüm, N.; Bozkurt, E.; Olgun, K.; Arntzen, J.W. A genomic footprint of hybrid zone movement in crested newts. Evol. Lett. 2017, 1, 93–101. [Google Scholar] [CrossRef]
- Akin Pekşen, Ç. Molecular Evolution and Phylogeography of the Eastern Mediterranean Water Frog (Pelophylax) Complex. Ph.D. Thesis, Middle East Technical University, Ankara, Turkey, 2015; 342p. [Google Scholar]
- Plötner, J.; Akın Pekşen, Ç.; Baier, F.; Uzzell, T.; Bilgin, C.C. Genetic evidence for human-mediated introduction of Anatolian water frogs (Pelophylax cf. bedriagae) to Cyprus (Amphibia: Ranidae). Zool. Middle East 2015, 61, 125–132. [Google Scholar] [CrossRef]
- Ohst, T. Genetische Einflüsse Allochthoner Wasserfrösche auf Endemische Wasserfroschpopulationen (R. kl. esculenta Komplex). Ph.D. Thesis, Humboldt-Universität zu Berlin, Berlin, Germany, 2008; p. 126. [Google Scholar]
- Ermakov, O.A.; Simonov, E.P.; Ivanov, A.J.; Zamaletdinov, R.I.; Fayzulin, A.I. Genetic characteristics of marsh frog (Pelophylax ridibundus complex) from the Western Caucasus based on mitochondrial and nuclear DNA data. Мoлекулярная генетика гидрoбиoнтoв/[oтв. ред. БА Лёвин].–Ярoславль: Филигрань, 2016.–78 с.–(РАН, Институт биoлoгии внутренних вoд им. ИД Папанина. Труды; вып. 2016, 73, 70–76. (In Russian) [Google Scholar]
- Dubey, S.; Dufresnes, C. An extinct vertebrate preserved by its living hybridogenetic descendant. Sci. Rep. 2017, 7, 12768. [Google Scholar] [CrossRef] [PubMed]
- Kukushkin, O.V.; Ivanov, A.Y.; Ermakov, O.A. Genetic heterogeneity of the marsh frog (Pelophylax (ridibundus) complex; Anura, Ranidae) population in Crimea revealed by mitochondrial and nuclear DNA analyses. Univ. Proc. Volga Reg. Nat. Sci. Zool. Penza 2018, 3, 32–54. (In Russian) [Google Scholar] [CrossRef]
- Faizulin, A.I.; Kukushkin, O.V.; Ivanov, A.Y.; Ermakov, O.A. Preliminary data on the molecular genetic structure of Pelophylax ridibundus (Amphibia: Anura: Ranidae) from the southern part of the Crimean Peninsula, based on mitochondrial and nuclear DNA analysis. Curr. Stud. Herpetol. 2017, 17, 56–65. (In Russian) [Google Scholar] [CrossRef]
- Plötner, J.; Köhler, F.; Uzzell, T.; Beerli, P.; Schreiber, R.; Guex, G.-D.; Hotz, H. Evolution of serum albumin intron-1 is shaped by a 50 truncated non-long terminal repeat retrotransposon in western Palearctic water frogs (Neobatrachia). Mol. Phylogenet. Evol. 2009, 53, 784–791. [Google Scholar] [CrossRef] [PubMed]
- Ermakov, O.A.; Fayzulin, A.I.; Askenderov, A.D.; Ivanov, A.J. Molecular-genetic characteristics of marsh frog from the Republic of Dagestan (based on mitochondrial and nuclear DNA data). Izv. Samarskogo Nauchnogo Tsentra Rossiyskoy Akademii Nauk 2016, 18, 94–97. (In Russian) [Google Scholar]
- Ivanov, A.Y.; Korzikov, V.A.; Alekseev, S.K.; Ermakov, O.A. Molecular and genetic characteristic of marsh frogs of Pelophylax ridibundus s.l. from Top Poochye. In Modern Problems of Zoology, Ecology and Conservancy; Ostapenko, V.A., Ed.; Selskokhozyaystvennye Tekhnologii: Moscow, Russia, 2015; pp. 228–232. (In Russian) [Google Scholar]
- Veith, M.; Kosuch, J.; Vences, M. Climatic oscillations triggered post-Messinian speciation of Western Palearctic brown frogs (Amphibia, Ranidae). Mol. Phylogenet. Evol. 2003, 26, 310–327. [Google Scholar] [CrossRef]
- Litvinchuk, S.N.; Matiushova, M.I.; Bezman-Moseyko, O.S.; Lada, G.A.; Skorinov, D.V.; Rosanov, J.M.; Borkin, L.J. Distribution, population systems, and peculiarities of hybrid gametogenesis in water frogs (Pelophylax esculentus complex) in the Dniester River valley (Moldova). Russ. J. Herpetol. 2023, 30, 88–100. [Google Scholar] [CrossRef]
- Barkhatov, A.S.; Snegin, E.A. Identification of cryptic forms of the hybridogenic complex of European water frogs (Pelophylax esculentus complex) in the conditions of transformed biotopes of the south of the Central Russian Upland based on DNA markers. Ecol. Genet. 2022, 20, 247–260. (In Russian) [Google Scholar] [CrossRef]
- Snegin, E.; Barkhatov, A.; Sychev, A.; Snegina, E. Species composition of green frogs (Pelophylax esculentus complex) of the Belgorod agglomeration based on DNA markers. BIO Web Conf. 2021, 30, 04008. (In Russian) [Google Scholar] [CrossRef]
- Ermakov, O.A.; Zaks, M.M.; Titov, S.V. Diagnostics and distribution of “western” and “eastern” forms of marsh frog Pelophylax ridibundus s. l. in the Penza province (on data of analysis of mtDNA cytochrome oxidase gene). Vestn. Tambov St. Univ. 2013, 18, 2999–3002. (In Russian) [Google Scholar]
- Vershinin, V.L.; Sitnikova, I.A.; Vershinina, S.D.; Trofimov, A.G.; Lebedinsky, A.A.; Miura, I.J. Mitochondrial heteroplasmia in marsh frog (Pelophylax ridibundus Pallas, 1771). Genetica Moscow 2019, 55, 972–977. (In Russian) [Google Scholar]
- Miura, I.; Vershinin, V.; Vershinina, S.; Lebedinskii, A.; Trofimov, A.; Sitnikov, I.; Ito, M. Hybridogenesis in the water frogs from western Russian territory: Intrapopulation variation in genome elimination. Genes 2021, 12, 244. [Google Scholar] [CrossRef]
- Kolenda, K.; Pietras-Lebioda, A.; Hofman, S.; Ogielska, M.; Pabijan, M. Preliminary genetic data suggest the occurrence of the Balkan water frog, Pelophylax kurtmuelleri, in southwestern Poland. Amphib.-Reptil. 2017, 38, 187–196. [Google Scholar] [CrossRef]
- Bellati, A.; Chiocchio, A.; Razzetti, E.; Bisconti, R.; Canestrelli, D. A hotchpotch of water frogs in northern Italy. Biol. Invasions 2023, 25, 2737–2745. [Google Scholar] [CrossRef]
- Zimić, A.; Vrhovac, B.; Šunje, E.; Ćurić, A.; Kalamujić Stroil, B. Molecular identification of green frogs (Anura: Ranidae: Pelophylax) of Western Balkans (Bosnia & Herzegovina and Montenegro). In Symposium of Biologists and Ecologists of Republic of Srpska with International Participation—SBERS2020; Jojić, D., Ed.; University of Banja Luka: Banja Luka, Herzegovina, 2020; pp. 152+presentation. [Google Scholar]
- Plötner, J.; Uzzell, T.; Beerli, P.; Spolsky, C.; Ohst, T.; Litvinchuk, S.N.; Guex, G.-D.; Reyer, H.-U.; Hotz, H. Widespread unidirectional transfer of mitochondrial DNA: A case in western Palaearctic water frogs. J. Evol. Biol. 2008, 21, 668–681. [Google Scholar] [CrossRef] [PubMed]
- Hotz, H.; Beerli, P.; Uzzell, T.; Guex, G.-D.; Pruvost, N.B.M.; Schreiber, R.; Plötner, J. Balancing a cline by influx of migrants: A genetic transition in water frogs of eastern Greece. J. Hered. 2013, 104, 57–71. [Google Scholar] [CrossRef]
- Vucić, M.; Jelić, D.; Klobučar, G.I.V.; Prkljačić, B.; Jelić, M. Molecular identification of species and hybrids of water frogs (genus Pelophylax) from Lake Skadar, Southeast Adriatic drainages (Amphibia: Ranidae). Salamandra 2018, 54, 147–157. [Google Scholar]
- Jelić, M.; Franjević, D.; Džikić, D.; Korlević, P.; Vucić, M.; Jelić, D.; Becking, T.; Grandjean, F.; Klobučar, G. Hybrid swarm as a result of hybridization between two alien and two native water frog species (genus Pelophylax) in Central Croatia: Crna Mlaka fishpond acting as a species melting pot? Biol. Invasions 2022, 24, 3291–3304. [Google Scholar] [CrossRef]
- Doležálková-Kaštánková, M.; Pyszko, P.; Choleva, L. Early development survival of Pelophylax water frog progeny is primarily affected by paternal genomic input. Front. Biosci. 2022, 27, 233. [Google Scholar] [CrossRef] [PubMed]
- Mayer, M.; Hawlitschek, O.; Zahn, A.; Glaw, F. Composition of twenty green frog populations (Pelophylax) across Bavaria, Germany. Salamandra 2013, 49, 31–44. [Google Scholar]
- Hawlitschek, O.; Morinière, J.; Dunz, A.; Franzen, M.; Rödder, D.; Glaw, F.; Haszprunar, G. Comprehensive DNA barcoding of the herpetofauna of Germany. Mol. Ecol. Resour. 2015, 16, 242–253. [Google Scholar] [CrossRef] [PubMed]
- Herczeg, D.; Vörös, J.; Christiansen, D.G.; Benovics, M.; Mikulíček, P. Taxonomic composition and ploidy level among European water frogs (Anura: Ranidae: Pelophylax) in eastern Hungary. J. Zool. Syst. Evol. Res. 2017, 55, 129–137. [Google Scholar] [CrossRef]
- Weigand, H.; Lopez de Llergo, J.C.; Frantz, A.C. Genomic basis for an informed conservation management of Pelophylax water frogs in Luxembourg. Ecol. Evol. 2022, 12, e8810. [Google Scholar] [CrossRef] [PubMed]
- Hoffmann, A.; Plötner, J.; Pruvost, N.B.M.; Christiansen, D.G.; Röthlisberger, S.; Choleva, L.; Mikulíček, P.; Cogălniceanu, D.; Sas-Kovács, I.; Shabanov, D.; et al. Genetic diversity and distribution patterns of diploid and polyploid hybrid water frog populations (Pelophylax esculentus complex) across Europe. Mol. Ecol. 2015, 24, 4371–4391. [Google Scholar] [CrossRef] [PubMed]
- Ermakov, O.A.; Fayzulin, A.I.; Zaks, M.M.; Kaybeleva, E.I.; Zaripova, F.F. Distribution «western» and «eastern» forms of marsh frog Pelophylax ridibundus s. l. in the Samara and Saratov region (on data of analysis of mtDNA and nDNA). Izvestiya Samarskogo Nauchnogo Tsentra Rossiyskoy Akademii Nauk 2014, 16, 409–412. (In Russian) [Google Scholar]
- Zamaletdinov, R.I.; Pavlov, A.V.; Zaks, M.M.; Ivanov, A.Y.; Ermakov, O.A. Molecular-genetic characteristic of Pelophylax esculentus complex from the eastern range of distribution (Volga region, Tatarstan Republic). Vest. Tambov St. Univ. Biol. 2015, 3, 54–66. (In Russian) [Google Scholar] [CrossRef]
- Svinin, A.O.; Ivanov, A.Y.; Zaks, M.M.; Litvinchuk, S.N.; Borkin, L.J.; Rosanov, J.M.; Ermakov, O.A. Distribution of the “eastern” and “western” forms of the marsh frog, Pelophylax ridibundus, and their participation in the origin of hemiclonal hybrids, P. esculentus in Mari El Republic. Curr. Stud. Herpetol. 2015, 15, 120–129. (In Russian) [Google Scholar]
- Dubey, S.; Maddalena, T.; Bonny, L.; Jeffries, D.L.; Dufresnes, C. Population genomics of an exceptional hybridogenetic system of Pelophylax water frogs. BMC Evol. Biol. 2019, 19, 164. [Google Scholar] [CrossRef]
- Matson, C.W.; Lambert, M.M.; McDonald, T.J.; Autenrieth, R.L.; Donnelly, K.C.; Islamzadeh, A.; Politov, D.I.; Bickham, J.W. Evolutionary toxicology: Population-level effects of chronic contaminant exposure on the marsh frogs (Rana ridibunda) of Azerbaijan. Environ. Health Perspect. 2006, 114, 547–552. [Google Scholar] [CrossRef] [PubMed]
- Holsbeek, G.; Mergeay, J.; Hotz, H.; Plötner, J.; Volckaert, F.A.M.; De Meester, L. A cryptic invasion within an invasion and widespread introgression in the European water frog complex: Consequences of uncontrolled commercial trade and weak international legislation. Mol. Ecol. 2008, 17, 5023–5035. [Google Scholar] [CrossRef] [PubMed]
- Doležálková-Kaštánková, M.; Pruvost, N.B.M.; Plötner, J.; Reyer, H.-U.; Janko, K.; Choleva, L. All-male hybrids of a tetrapod Pelophylax esculentus share its origin and genetics of maintenance. Biol. Sex Diff. 2018, 9, 13. [Google Scholar] [CrossRef] [PubMed]
- Zangl, L.; Daill, D.; Schweiger, S.; Gassner, G.; Koblmüller, S. A reference DNA barcode library for Austrian amphibians and reptiles. PLoS ONE 2020, 15, e0229353. [Google Scholar] [CrossRef] [PubMed]
- Holsbeek, G.; Maes, G.E.; De Meester, L.; Volckaert, F.A.M. Conservation of the introgressed European water frog complex using molecular tools. Mol. Ecol. 2009, 18, 1071–1087. [Google Scholar] [CrossRef] [PubMed]
- Mikulíček, P.; Kautman, M.; Demovič, B.; Janko, K. When a clonal genome finds its way back to a sexual species: Evidence from ongoing but rare introgression in the hybridogenetic water frog complex. J. Evol. Biol. 2014, 27, 628–642. [Google Scholar] [CrossRef] [PubMed]
- Dubey, S.; Leuenberger, J.; Perrin, N. Multiple origins of invasive and ‘native’ water frogs (Pelophylax spp.) in Switzerland. Biol. J. Linn. Soc. 2014, 112, 442–449. [Google Scholar] [CrossRef]
- Phillips, S.J.; Anderson, R.P.; Schapire, R.E. Maximum entropy modeling of species geographic distributions. Ecol. Model. 2006, 190, 231–259. [Google Scholar] [CrossRef]
- Warren, D.L.; Glor, R.E.; Turelli, M. ENMTools: A toolbox for comparative studies of environmental niche models. Ecography 2010, 33, 607–611. [Google Scholar] [CrossRef]
- Cobos, M.E.; Peterson, A.T.; Barve, N.; Osorio-Olvera, L. kuenm: An R package for detailed development of ecological niche models using Maxent. PeerJ 2019, 7, e6281. [Google Scholar] [CrossRef]
- Allouche, O.; Tsoar, A.; Kadmon, R. Assessing the accuracy of species distribution models: Prevalence, kappa and the true skill statistic (TSS). J. Appl. Ecol. 2006, 43, 1223–1232. [Google Scholar] [CrossRef]
- Phillips, S.J.; Anderson, R.P.; Dudík, M.; Schapire, R.E.; Blair, M.E. Opening the black box: An open-source release of Maxent. Ecography 2017, 40, 887–893. [Google Scholar] [CrossRef]
- Ratnikov, V.Y. Fossil remains of modern amphibian and reptile species as the material for studying of their areas history. Tr. Nauchno-Issledovatel’skogo Instituta Geol. Voronezhskogo Gos. Univ. Voronezh 2009, 59, 1–91. (In Russian) [Google Scholar]
- Böhme, G.; Günther, R. Osteological studies in the European water frogs Rana ridibunda, Rana lessonae and Rana “esculenta” (Anura, Ranidae). Mitt. Zool. Mus. Berlin 1979, 55, 203–215. [Google Scholar]
- Böhme, G. Skelettreste von Amphibien (Urodela, Salenia) aus dem fossilen Tierbautensystem von Pisede bei Malchin. Teil 1: Taxonomie und Biostrationomie. Wissenschaft. Zeitschr. Humbolt-Univer. Berlin Math.-Nat. R. 1983, 32, 657–670. [Google Scholar]
- Böhme, G. Skelettreste von Amphibien (Urodela, Salenia) aus dem fossilen Tierbautensystem von Pisede bei Malchin. Teil 2: Palaookologische und faunengeschichtliche Auswertung. Wissenschaft. Zeitschr. Humbolt-Univer. Berlin Math.-Nat. R. 1983, 32, 671–680. [Google Scholar]
- Höglund, J.; Bolender, L.; Cortazar-Chinarro, M.; Meurling, S.; Laurila, A.; Hermaniuk, A.; Dufresnes, C. Low neutral and immunogenetic diversity in northern fringe populations of the green toad Bufotes viridis: Implications for conservation. Conserv. Genet. 2022, 23, 139–149. [Google Scholar] [CrossRef]
- Sagonas, K.; Karameta, E.; Kotsakiozi, P.; Poulakakis, N. Cross-species testing of nuclear markers in Pelophylax water frogs in Greece and examination of their power to detect genetic admixture. Amphib.-Reptil. 2020, 41, 253–259. [Google Scholar] [CrossRef]
- Bellati, A.; Razzetti, E.; Resteghini, M.; Sacchi, R.; Pellitteri-Rosa, D.; Casiraghi, M.; Bernini, F.; Galeotti, P.; Fasola, M. First molecular characterization of invasive alien populations of Pelophylax kurtmuelleri (Gayda, 1940) and new records from Italy. In Atti IX Congresso Nazionale della Societas Herpetologica Italica; Scillitani, G., Liuzzi, C., Lorusso, L., Mastropasqua, F., Ventrella, P., Eds.; Pineta: Conversano, Italy, 2013; pp. 287–289. [Google Scholar]
- Bellati, A.; Bassu, L.; Nulchis, V.; Corti, C. Detection of alien Pelophylax species in Sardinia (western Mediterranean, Italy). BioInvasions Rec. 2019, 8, 8–25. [Google Scholar] [CrossRef]
- Hewitt, G.M. Some genetic consequences of ice ages, and their role in divergence and speciation. Biol. J. Linn. Soc. 1996, 58, 247–276. [Google Scholar] [CrossRef]
- Hewitt, G. The genetic legacy of the Quaternary ice ages. Nature 2000, 405, 907–913. [Google Scholar] [CrossRef] [PubMed]
- Hewitt, G.M. Speciation, hybrid zones and phylogeography—Or seeing genes in space and time. Mol. Ecol. 2001, 10, 537–549. [Google Scholar] [CrossRef] [PubMed]
- Hewitt, G.M. Quaternary phylogeography: The roots of hybrid zones. Genetica 2011, 139, 617–638. [Google Scholar] [CrossRef]
- Taberlet, P.; Cheddadi, R. Quaternary refugia and persistence of biodiversity. Science 2002, 297, 2009–2010. [Google Scholar] [CrossRef] [PubMed]
- Taberlet, P.; Fumagalli, L.; Wust-Saucy, A.-G.; Cosson, J.-F. Comparative phylogeography and post-glacial colonization route in Europe. Mol. Ecol. 1998, 7, 453–464. [Google Scholar] [CrossRef] [PubMed]
- Petit, R.J.; Aguinagalde, I.; de Beaulieu, J.-L.; Bittkau, C.; Brewer, S.; Cheddadi, R.; Ennos, R.; Fineschi, S.; Grivet, D.; Lascoux, M.; et al. Glacial refugia: Hotspots but not melting pots of genetic diversity. Science 2003, 300, 1563–1565. [Google Scholar] [CrossRef] [PubMed]
- Schmitt, T. Molecular biogeography of Europe: Pleistocene cycles and post-glacial trends. Front. Zool. 2007, 4, 11. [Google Scholar] [CrossRef]
- Bhagwat, S.A.; Willis, K.J. Species persistence in northerly glacial refugia of Europe: A matter of chance or biogeographical traits? J. Biogeogr. 2008, 35, 464–482. [Google Scholar] [CrossRef]
- Habel, J.C.; Drees, C.; Schmitt, T.; Assmann, T. Review refugial areas and post-glacial colonizations in the Western Palearctic. In Relict Species: Phylogeography and Conservation Biology; Habel, J.C., Assmann, T., Eds.; Springer: Berlin/Heidelberg, Germany, 2010; pp. 189–197. [Google Scholar]
- Nève, G.; Verlaque, R. Genetic differentiation between and among refugia. In Relict Species: Phylogeography and Conservation Biology; Habel, J.C., Assmann, T., Eds.; Springer: Berlin/Heidelberg, Germany, 2010; pp. 277–294. [Google Scholar]
- Feliner, G.N. Southern European glacial refugia: A tale of tales. Taxon 2011, 60, 365–372. [Google Scholar] [CrossRef]
- Provan, J.; Bennett, K.D. Phylogeographic insights into cryptic glacial refugia. Trends Ecol. Evol. 2008, 23, 564–571. [Google Scholar] [CrossRef]
- Schmitt, T. Biogeographical and evolutionary importance of the European high mountain systems. Front. Zool. 2009, 6, 9. [Google Scholar] [CrossRef] [PubMed]
- Stewart, J.R.; Lister, A.M.; Barnes, I.; Dalén, L. Refugia revisited: Individualistic responses of species in space and time. Proc. R. Soc. B 2010, 277, 661–671. [Google Scholar] [CrossRef] [PubMed]
- Varga, Z. Extra-Mediterranean refugia, post-glacial vegetation history and area dynamics in Eastern Central Europe. In Relict Species: Phylogeography and Conservation Biology; Habel, J.C., Assmann, T., Eds.; Springer: Berlin/Heidelberg, Germany, 2010; pp. 57–87. [Google Scholar]
- Ratnikov, V. Do northern species of amphibians and reptiles of Eurasia need southern refugia? Biol. J. Linn. Soc. 2023. [Google Scholar] [CrossRef]
- Grichuk, V.P. Vegetation of Europe in the Late Pleistocene. In Paleogeography of Europe for the Last 100 Thousand Years; Gerasimov, I.P., Velichko, A.A., Eds.; Nauka: Moscow, Russia, 1982; pp. 92–109. (In Russian) [Google Scholar]
- Allen, J.R.M.; Hickler, T.; Singarayer, J.S.; Sykes, M.T.; Valdes, P.J.; Huntley, B. Last glacial vegetation of northern Eurasia. Quart. Sci. Rew. 2010, 29, 2604–2618. [Google Scholar] [CrossRef]
- Kireev, V.A. Fauna of Kalmykia. Amphibians and Reptiles; Kalmytskoe Knizhnoe Izdatel’stvo: Elista, Russia, 1983; p. 113. (In Russian) [Google Scholar]
- Belik, V. Recourses of Biodiversity of the Steppe Don River Region. Amphibians and Reptiles; Lap Lambert Academic Publishing: Saarbrücken, Germany, 2011; p. 94. (In Russian) [Google Scholar]
- Schmitt, T.; Varga, Z. Extra-Mediterranean refugia: The rule and not the exception? Front. Zool. 2012, 9, 22. [Google Scholar] [CrossRef] [PubMed]
- Litvinchuk, S.N.; Crottini, A.; Federici, S.; De Pous, P.; Donaire, D.; Andreone, F.; Kalezić, M.L.; Džukić, G.; Lada, G.A.; Borkin, L.J.; et al. Phylogeographic patterns of genetic diversity in the common spadefoot toad, Pelobates fuscus (Anura: Pelobatidae), reveals evolutionary history, post-glacial range expansion and secondary contact. Organ. Div. Evol. 2013, 13, 433–451. [Google Scholar] [CrossRef]
- Dufresnes, C.; Perrin, N. Effect of biogeographic history on population vulnerability in European amphibians. Conserv. Biol. 2014, 29, 1235–1241. [Google Scholar] [CrossRef]
- Skorinov, D.V.; Litvinchuk, S.N. Tracing glacial refugia of the smooth newt (Lissotriton vulgaris) based on species distribution modelling. Vest. St. Petersburg St. Univ. Biol. 2016, 3, 136–143. (In Russian) [Google Scholar] [CrossRef]
- Dufresnes, C.; Suchan, T.; Smirnov, N.A.; Denoël, M.; Rosanov, J.M.; Litvinchuk, S.N. Revisiting a speciation classic: Comparative analyses support sharp but leaky transitions between Bombina toads. J. Biogeogr. 2021, 48, 548–560. [Google Scholar] [CrossRef]
- Babik, W.; Branicki, W.; Sandera, M.; Litvinchuk, S.; Borkin, L.J.; Irwin, J.T.; Rafinski, J. Mitochondrial phylogeography of the moor frog, Rana arvalis. Mol. Ecol. 2004, 13, 1469–1480. [Google Scholar] [CrossRef]
- Roček, Z.; Šandera, M. Distribution of Rana arvalis in Europe: A historical perspective. Zeitschr. Feldherpetol. 2008, Suppl. 13, 135–150. [Google Scholar]
- Knopp, T.; Merilä, J. The post-glacial recolonization of Northern Europe by Rana arvalis as revealed by microsatellite and mitochondrial DNA analyses. Heredity 2009, 102, 174–181. [Google Scholar] [CrossRef]
- Litvinchuk, S.N.; Borkin, L.J. Evolution, Systematics and Distribution of Crested Newts (Triturus cristatus complex) in the Territory of Russia and Adjacent Countries; Evropeyskiy dom: St. Petersburg, Russia, 2009; pp. 592+24. (In Russian) [Google Scholar]
- Teacher, A.G.F.; Garner, T.W.J.; Nichols, R.A. European phylogeography of the common frog (Rana temporaria): Routes of post-glacial colonization into the British Isles, and evidence for an Irish glacial refugium. Heredity 2009, 102, 490–496. [Google Scholar] [CrossRef]
- Wielstra, B.; Babik, W.; Arntzen, J.W. The crested newt Triturus cristatus recolonized temperate Eurasia from an extra-Mediterranean glacial refugium. Biol. J. Linn. Soc. 2015, 114, 574–587. [Google Scholar] [CrossRef]
- Vörös, J.; Mikulíček, P.; Major, A.; Recuero, E.; Arntzen, J.W. Phylogeographic analysis reveals northerly refugia for the riverine amphibian Triturus dobrogicus (Caudata: Salamandridae). Biol. J. Linn. Soc. 2016, 119, 974–991. [Google Scholar] [CrossRef]
- van Rensburg, A.J.; Robin, M.; Phillips, B.; Buskirk, J.V. European common frog (Rana temporaria) recolonized Switzerland from multiple glacial refugia in northern Italy via trans-and circum-Alpine routes. Ecol. Evol. 2021, 11, 15984–15994. [Google Scholar] [CrossRef] [PubMed]






Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Litvinchuk, S.N.; Skorinov, D.V.; Ivanov, A.Y.; Ermakov, O.A. Detection of Glacial Refugia and Post-Glacial Colonization Routes of Morphologically Cryptic Marsh Frog Species (Anura: Ranidae: Pelophylax) Using Environmental Niche Modeling. Diversity 2024, 16, 94. https://doi.org/10.3390/d16020094
Litvinchuk SN, Skorinov DV, Ivanov AY, Ermakov OA. Detection of Glacial Refugia and Post-Glacial Colonization Routes of Morphologically Cryptic Marsh Frog Species (Anura: Ranidae: Pelophylax) Using Environmental Niche Modeling. Diversity. 2024; 16(2):94. https://doi.org/10.3390/d16020094
Chicago/Turabian StyleLitvinchuk, Spartak N., Dmitriy V. Skorinov, Alexander Yu. Ivanov, and Oleg A. Ermakov. 2024. "Detection of Glacial Refugia and Post-Glacial Colonization Routes of Morphologically Cryptic Marsh Frog Species (Anura: Ranidae: Pelophylax) Using Environmental Niche Modeling" Diversity 16, no. 2: 94. https://doi.org/10.3390/d16020094