

Coexistence of Two Copepods, Recorded for the First Time, in NW Iberian Shelf: The Case of Oithona atlantica and the Allochthonous O. davisae

Abstract
:1. Introduction
2. Materials and Methods
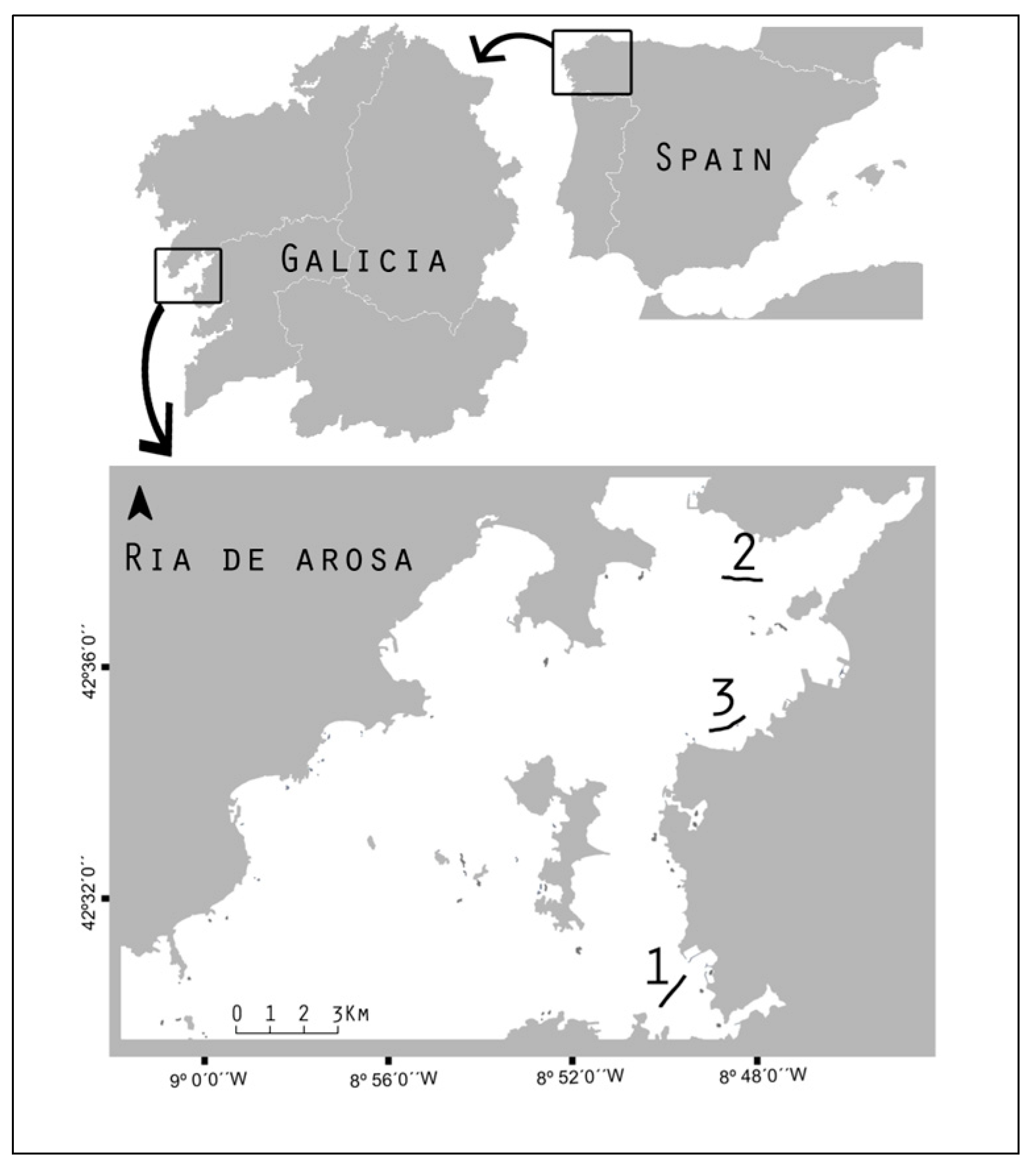
2.1. Sampling
2.2. Morphological Analyses
2.3. Molecular Analyses
3. Results
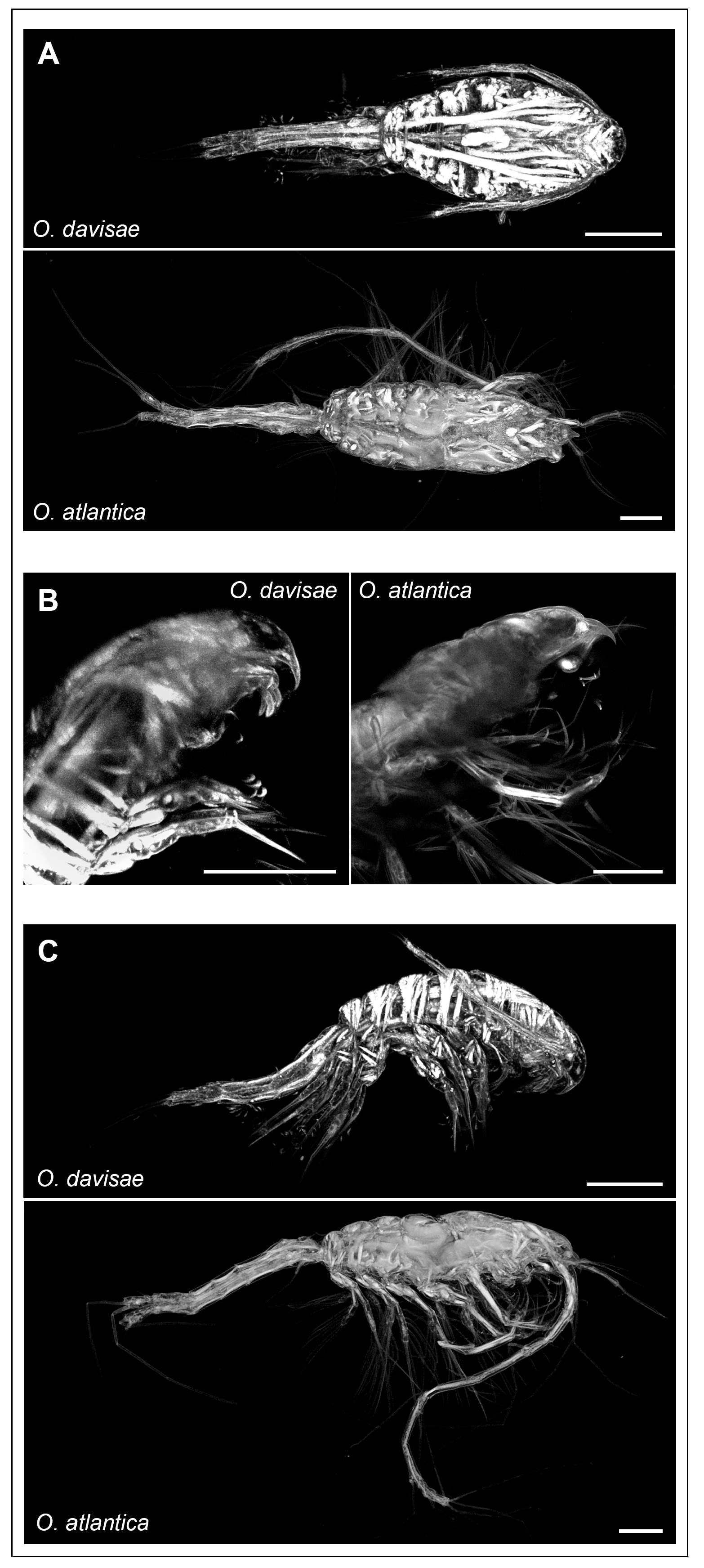
3.1. Morphological Analyses
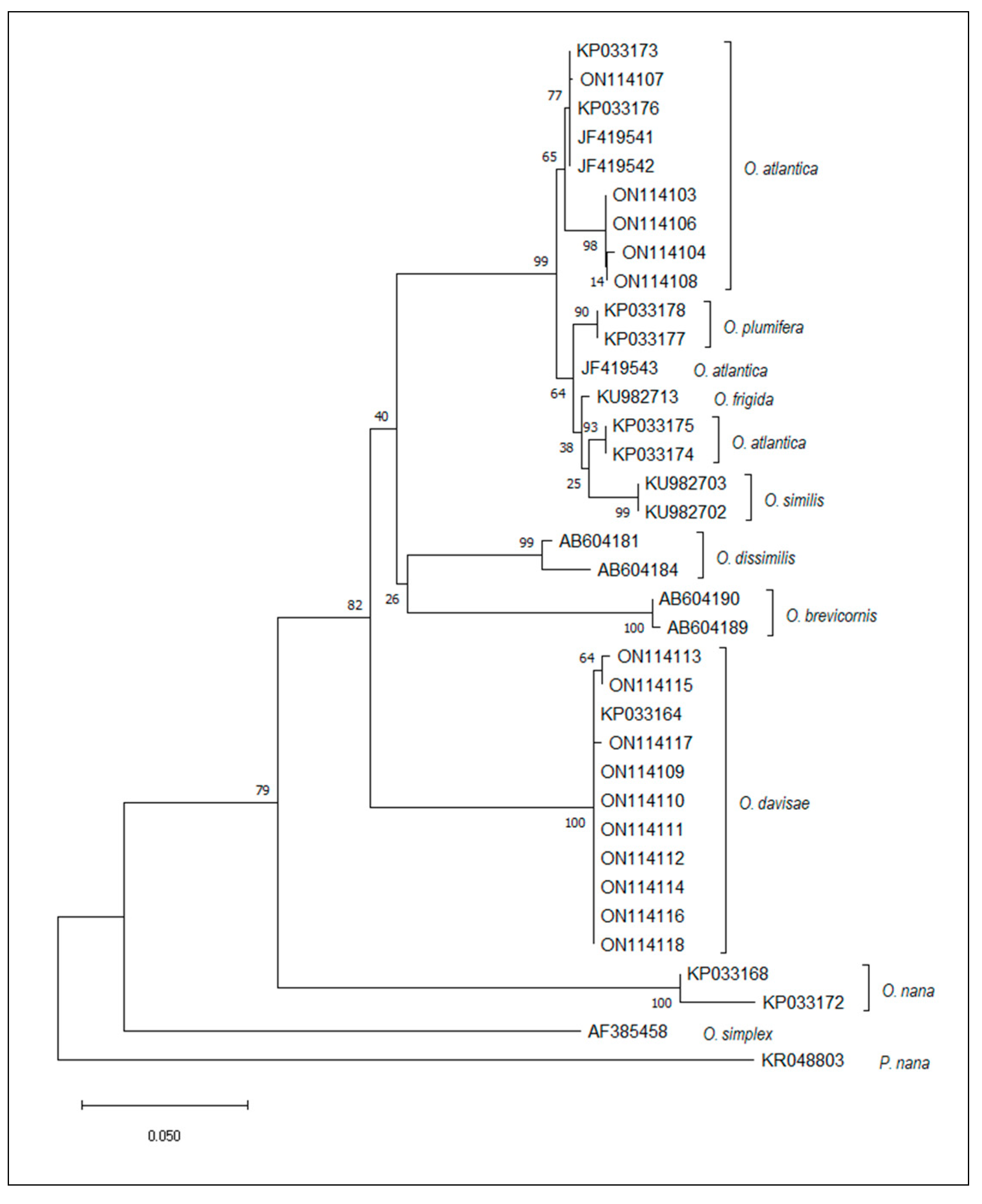
3.2. Molecular Analyses
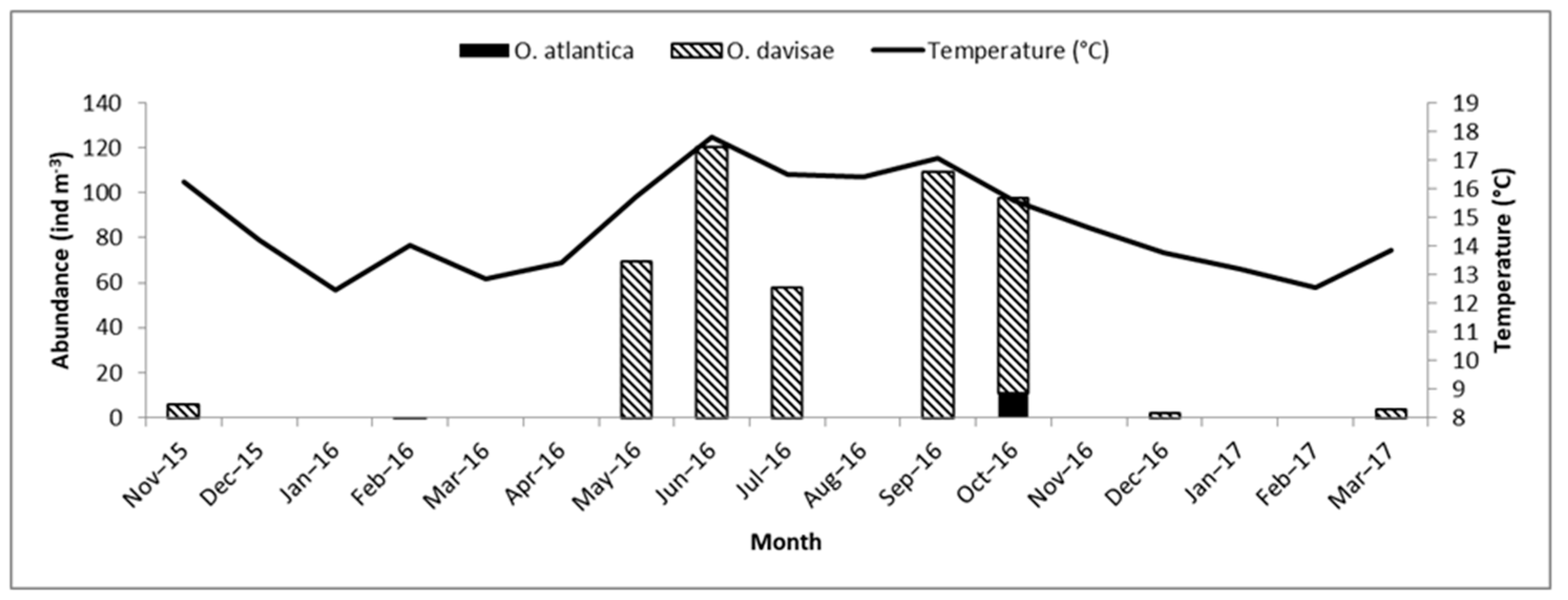
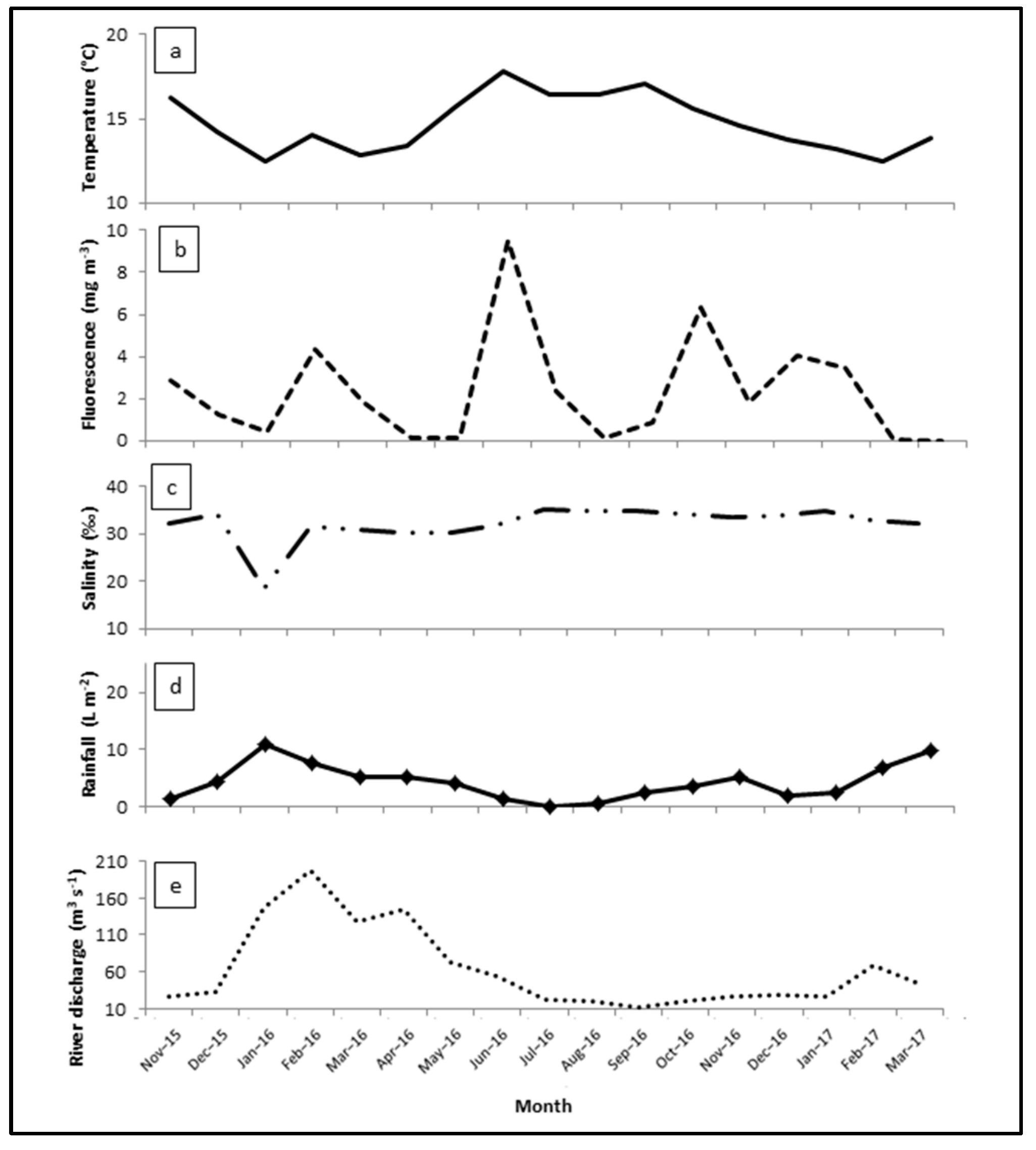
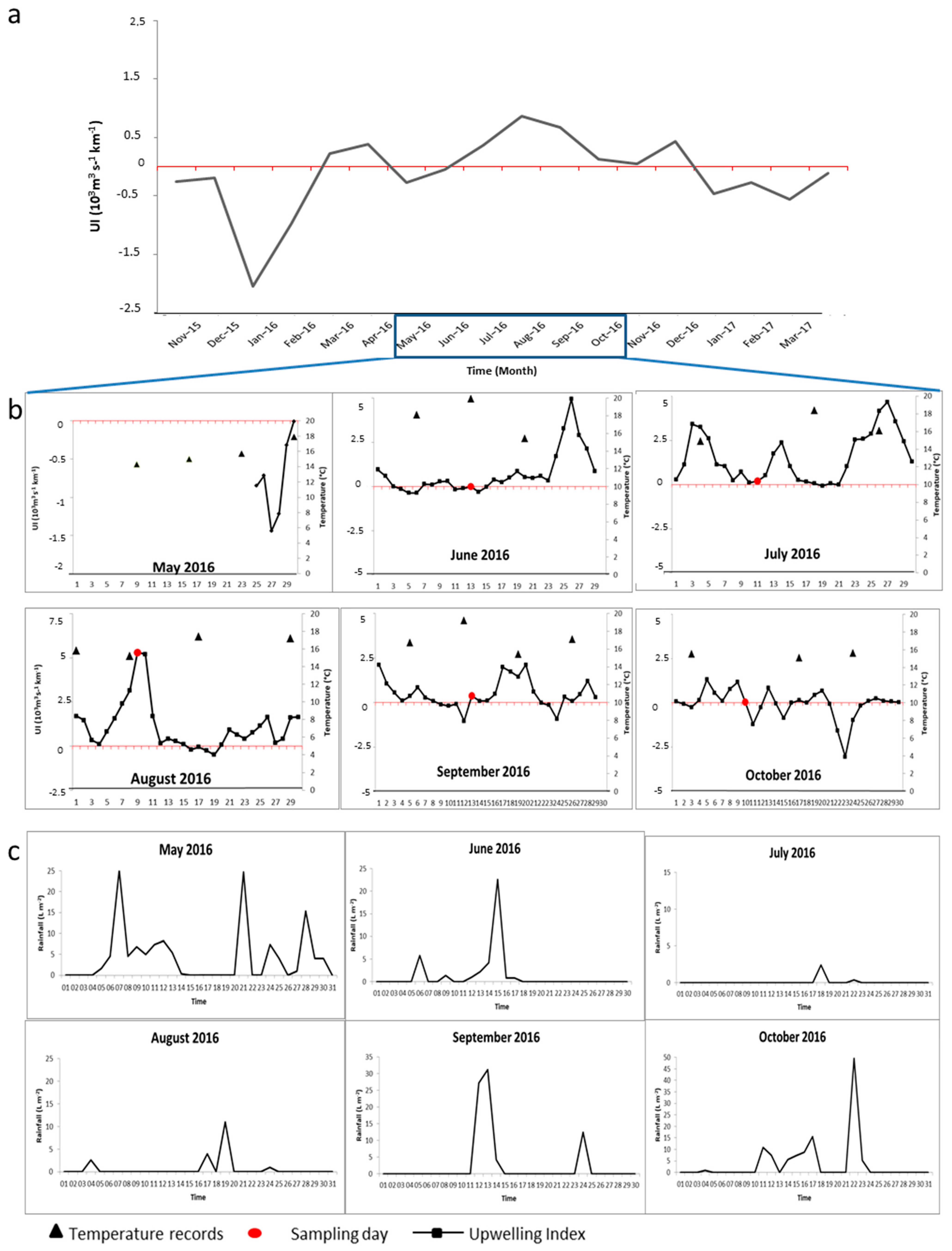
3.3. Seasonal Abundance Variation
3.4. Expansion of the Two Copepod Species in the Ría de Arosa
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Turner, J.F. The importance of small planktonic copepods and their roles in pelagic marine food webs. Zool. Stud. 2004, 43, 255–266. [Google Scholar]
- Valdés, V.; Carlotti, F.; Escribano, R.; Donoso, K.; Pagano, M.; Molina, V.; Fernandez, C. Nitrogen and phosphorus recycling mediated by copepods in Western Tropical South Pacific. Biogeosciences 2018, 15, 6019–6032. [Google Scholar] [CrossRef]
- Taylor, A.H.; Allen, J.I.; Clark, P.A. Extraction of a weak climatic signal by an ecosystem. Nature 2002, 416, 629–632. [Google Scholar] [CrossRef] [PubMed]
- Perry, R.I.; Batchelder, H.P.; Mackas, D.L.; Chiba, S.; Durbin, E.; Greve, W.; Verheye, H.M. Identifying global synchronies in marine zooplankton populations: Issues and opportunities. ICES J. Mar. Sci. 2004, 61, 445–456. [Google Scholar] [CrossRef]
- Hays, G.C.; Richardson, A.J.; Robinson, C. Climate change and marine plankton. Trends Ecol. Evol. 2005, 20, 337–344. [Google Scholar] [CrossRef]
- Alvarez-Salgado, X.A.; Rosón, G.; Pérez, F.F.; Figueiras, F.G.; Pazos, Y. Hydrographic variability off the Rías Baixas (NW Spain) during the upwelling season. J. Geohys. Res. 1993, 98, 14447–14455. [Google Scholar] [CrossRef]
- Blanton, J.O.; Tenore, K.R.; Castillejo, F.F.; Atkinson, L.P.; Schwing, F.B.; Lavin, A. The relationship of upwelling to mussel production in the rias on the western coast of Spain. J. Mar. Res. 1987, 45, 497–511. [Google Scholar] [CrossRef]
- Figueiras, F.G.; Pazos, Y. Microplankton assemblages in three Rías Baixas (Vigo, Arosa and Muros, Spain) with a subsurface chlorophyll maximum: Their relationships to hydrography. Mar. Ecol. Prog. Ser. 1991, 76, 219–233. Available online: http://www.jstor.org/stable/24825583 (accessed on 6 July 2021). [CrossRef]
- Bode, A.; Álvarez-Ossorio, M.T.; González, N.; Stephens, J.; Jordan, M.; Taylor, A.; Proctor, R. Estimations of mesozooplankton biomass in a coastal upwelling area off NW Spain. J. Plankton Res. 1998, 20, 1005–1014. [Google Scholar] [CrossRef]
- Bode, A.; Álvarez-Ossorio, M.T.; González, N.; Lorenzo, J.; Rodríguez, C.; Varela, M.; Varela, M.M. Seasonal variability of plankton blooms in the Ria de Ferrol (NW Spain): II. Plankton abundance, composition and biomass. Estuar. Coast. Shelf Sci. 2005, 63, 285–300. [Google Scholar] [CrossRef]
- Varela, M.; Penas, E. Primary production of benthic microalgae in an intertidal sand flat of the Ria de Arosa, NW Spain. Mar. Ecol. Prog. Ser. 1985, 25, 111–119. [Google Scholar] [CrossRef]
- Hanson, R.B.; Alvarez-Ossorio, M.T.; Cal, R.; Campos, M.J.; Roman, M.; Santiago, G.; Varela, M.; Yoder, J.A. Plankton response following a spring upwelling event in the Ria de Arosa, Spain. Mar. Ecol. Prog. Ser. 1986, 32, 101–113. [Google Scholar] [CrossRef]
- Barciela, R.M.; García, E.; Fernández, E. Modelling primary production in a coastal embayment affected by upwelling using dynamic ecosystem models and artificial neural networks. Ecol. Model. 1999, 120, 199–211. [Google Scholar] [CrossRef]
- Corral, J.; Alvarez-Ossorio, M. El zooplancton de la Ria de Arosa (NW de España): Composicion y distribucion de las comunidades en un ciclo anual. Bol. Inst. Esp. Oceanogr. 1978, 265, 133–163. [Google Scholar]
- Molares, J.; Fuentes, J. Recruitment of the mussel Mytilus galloprovincialis on collectors situated on the intertidal zone in the Riá de Arousa (NW Spain). Aquaculture 1995, 138, 131–137. [Google Scholar] [CrossRef]
- Macho, G.; Molares, J.; Vázquez, E. Timing of larval release by three barnacles from the NW Iberian Peninsula. Mar. Ecol. Prog. Ser. 2005, 298, 251–260. [Google Scholar] [CrossRef]
- Macho, G. Ecología Reproductiva y Larvaria del Percebe y otros Cirrípedos en Galicia. Ph.D. Thesis, Universidade de Vigo, Vigo, Spain, 2006. [Google Scholar]
- Macho, G.; Vázquez, E.; Giráldez, R.; Molares, J. Spatial and temporal distribution of barnacle larvae in the partially mixed estuary of the Ría de Arousa (Spain). J. Exp. Mar. Biol. Ecol. 2010, 392, 129–139. [Google Scholar] [CrossRef]
- Miranda, A.; Alvarez-Ossorio, M.; Valdés, L.; Casas, G. Copepoda. In Inventario de la Biodiversidad Marina de Galicia: Proyecto LEMGAL; Bañón, R., Ed.; Consellería do Mar, Xunta de Galicia: Santiago de Compostela, Spain, 2017; pp. 392–401. [Google Scholar]
- Mazzocchi, M.G. Oithona Baird, 1843; ICES Identification Leaflets for Plankton No. 188; ICES: Copenhagen, Denmark, 2019. [Google Scholar] [CrossRef]
- Temnykh, A.; Nishida, S. New record of the planktonic copepod Oithona davisae Ferrari and Orsi in the Black Sea with notes on the identity of “Oithona brevicornis”. Aquat. Invasions 2012, 7, 425–431. [Google Scholar] [CrossRef]
- Ferrari, F.D.; Orsi, J. Oithona Davisae, New Species, and Limnoithona sinensis (Burckhardt, 1912) (Copepoda: Oithonidae) from the Sacramento-San Joaquin Estuary, California. J. Crustac. Biol. 1984, 4, 106–126. [Google Scholar] [CrossRef]
- Mihneva, V.; Stefanova, K. The non-native copepod Oithona davisae (Ferrari F.D. and Orsi, 1984) in the Western Black Sea: Seasonal and annual abundance variability. BioInvasions Rec. 2013, 2, 119–124. [Google Scholar] [CrossRef]
- Cornils, A.; Wend-Heckmann, B. First report of the planktonic copepod Oithona davisae in the northern Wadden Sea (North Sea): Evidence for recent invasion? Helgol. Mar. Res. 2015, 69, 243–248. [Google Scholar] [CrossRef]
- Cordell, J.R.; Lawrence, D.J.; Ferm, N.C.; Tear, L.M.; Smith, S.S.; Herwig, R.P. Factors influencing densities of non-indigenous species in the ballast water of ships arriving at ports in Puget Sound, Washington, United States. Aquat. Conserv. Mar. Freshw. Ecosyst. 2008, 19, 322–343. [Google Scholar] [CrossRef]
- Isinibilir, M.; Svetlichny, L.S.; Hubareva, E.S. Small oithonid copepods in the northeastern Marmara Sea. In Proceedings of the 41st CIESM Congress, Kiel, Germany, 12–16 September 2016; Volume 41, p. 483. [Google Scholar]
- Vidjak, O.; Bojanić, N.; de Olazabal, A.; Benzi, M.; Brautović, I.; Camatti, E.; Hure, M.; Lipej, L.; Lučić, D.; Pansera, M.; et al. Zooplankton in Adriatic port environments: Indigenous communities and non-indigenous species. Mar. Pollut. Bull. 2018, 147, 133–149. [Google Scholar] [CrossRef] [PubMed]
- Pansera, M.; Camatti, E.; Schroeder, A.; Zagami, G.; Bergamasco, A. The non-indigenous Oithona davisae in a Mediterranean transitional environment: Coexistence patterns with competing species. Sci. Rep. 2021, 11, 8341. [Google Scholar] [CrossRef]
- Altukhov, D.A.; Gubanova, A.D.; Mukhanov, V.S. New invasive copepod Oithona davisae Ferrari and Orsi, 1984: Seasonal dynamics in Sevastopol Bay and expansion along the Black Sea coasts. Mar. Ecol. 2014, 35, 28–34. [Google Scholar] [CrossRef]
- Zagami, G.; Brugnano, C.; Granata, A.; Guglielmo, L.; Minutoli, R.; Aloise, A. Biogeographical distribution and ecology of the planktonic copepod Oithona davisae: Rapid invasion in Lakes Faro and Ganzirri (Central Mediterranean Sea). In Trends in Copepod Studies—Distribution, Biology and Ecology; Uteri, M., Ed.; Nova Science Publishers: New York, NY, USA, 2019; pp. 59–82. [Google Scholar]
- Saiz, E.; Calbet, A.; Broglio, E. Effects of small-scale turbulence on copepods: The case of Oithona davisae. Limnol. Oceanogr. 2003, 48, 1304–1311. [Google Scholar] [CrossRef]
- Uriarte, I.; Villate, F.; Iriarte, A. Zooplankton recolonization of the inner estuary of Bilbao: Influence of pollution abatement, climate and non-indigenous species. J. Plankton Res. 2015, 38, 718–731. [Google Scholar] [CrossRef]
- Cinar, M.E.; Bilecenoglu, M.; Ozturk, B.; Katagan, T.; Aysel, V. Alien species on the coasts of Turkey. Mediterr. Mar. Sci. 2005, 6, 119. [Google Scholar] [CrossRef]
- Nishida, S. Taxonomy and distribution of the family Oithonidae (Copepoda, Cyclopoida) in the Pacific and Indian Oceans. Bull. Ocean. Res. Inst. Univ. Tokyo 1985, 20, 167. [Google Scholar]
- Ortman, B.D. DNA Barcoding the Medusozoa and Ctenophora. Ph.D. Thesis, University of Connecticut, Storrs, CT, USA, 2008. [Google Scholar]
- Cepeda, G.D.; Blanco-Bercial, L.; Bucklin, A.; Berón, C.M.; Viñas, M.D. Molecular Systematic of Three Species of Oithona (Copepoda, Cyclopoida) from the Atlantic Ocean: Comparative Analysis Using 28S rDNA. PLoS ONE 2012, 7, e35861. [Google Scholar] [CrossRef]
- Thompson, J.D.; Higgins, D.G.; Gibson, T.J. CLUSTAL W: Improving the sensitivity of progressive multiple sequence alignment through sequence weighting, position-specific gap penalties and weight matrix choice. Nucleic Acids Res. 1994, 22, 4673–4680. [Google Scholar] [CrossRef] [PubMed]
- Kumar, S.; Stecher, G.; Tamura, K. MEGA7: Molecular Evolutionary Genetics Analysis version 7.0 for bigger datasets. Mol. Biol. Evol. 2016, 33, 1870–1874. [Google Scholar] [CrossRef] [PubMed]
- Darriba, D.; Taboada, G.L.; Doallo, R.; Posada, D. jModelTest 2: More models, new heuristics and parallel computing. Nat. Methods 2012, 9, 772. [Google Scholar] [CrossRef]
- Cornils, A.; Wend-Heckmann, B.; Held, C. Global phylogeography of Oithona similis s.l. (Crustacea, Copepoda, Oithonidae)—A cosmopolitan plankton species or a complex of cryptic lineages? Mol. Phylogenetics Evol. 2017, 107, 473–485. [Google Scholar] [CrossRef] [PubMed]
- Ueda, H.; Yamaguchi, A.; Saitoh, S.-I.; Sakaguchi, S.O.; Tachihara, K. Speciation of two salinity-associated size forms of Oithona dissimilis (Copepoda: Cyclopoida) in estuaries. J. Nat. Hist. 2011, 45, 2069–2079. [Google Scholar] [CrossRef]
- Pansera, M.; Granata, A.; Guglielmo, L.; Minutoli, R.; Zagami, G.; Brugnano, C. How does mesh-size selection reshape the description of zooplankton community structure in coastal lakes? Estuar. Coast. Shelf Sci. 2014, 151, 221–235. [Google Scholar] [CrossRef]
- Kurt, T.T.; Beşiktepe, S. First distribution record of the invasive copepod Oithona davisae Ferrari and Orsi, 1984, in the coastal waters of the Aegean Sea. Mar. Ecol. 2019, 40, e12548. [Google Scholar] [CrossRef]
- Yıldız, I.; Feyzioglu, A.M.; Besiktepe, S. First observation and seasonal dynamics of the new invasive planktonic copepod Oithona davisae Ferrari and Orsi, 1984 along the southern Black Sea (Anatolian Coast). J. Nat. Hist. 2017, 51, 127–139. [Google Scholar] [CrossRef]
- Atkinson, A. Omnivory and feeding selectivity in five copepod species during spring in the Bellingshausen Sea, Antarctica. ICES J. Mar. Sci. 1995, 52, 385–396. [Google Scholar] [CrossRef]
- Nakamura, Y.; Turner, J.T. Predation and respiration by the small cyclopoid copepod Oithona similisr: How important is feeding on ciliates and heterotrophic flagellates? J. Plankton Res. 1997, 19, 1275–1288. [Google Scholar] [CrossRef]
- Broglio, E.; Saiz, E.; Calbet, A.; Trepat, I.; Alcaraz, M. Trophic impact and prey selection by crustacean zooplankton on the microbial communities of an oligotrophic coastal area (NW Mediterranean Sea). Aquat. Microb. Ecol. 2004, 35, 65–78. [Google Scholar] [CrossRef]
- Saiz, E.; Griffell, K.; Calbet, A.; Isari, S. Feeding rates and prey: Predator size ratios of the nauplii and adult females of the marine cyclopoid copepod Oithona davisae. Limnol. Oceanogr. 2014, 59, 2077–2088. [Google Scholar] [CrossRef]
- Uchima, M.; Hirano, R. Food of Oithona davisae (Copepoda: Cyclopoida) and the effect of food concentration at first feeding on the larval growth. Bull. Plankton Soc. Jpn. 1986, 33, 21–28. [Google Scholar]
- Alvarez-Salgado, X.A.; Rosón, G.; Pérez, F.F.; Figueiras, F.G.; Pazos, Y. Nitrogen cycling in an estuarine upwelling system, the Ría de Arousa (NW Spain). I. Short-time-scale patterns of hydrodynamic and biogeochemical circulation. Mar. Ecol. Prog. Ser. 1996, 135, 259–273. [Google Scholar] [CrossRef]
- Villacieros-Robineau, N.; Herrera, J.L.; Castro, C.G.; Piedracoba, S.; Roson, G. Hydrodynamic characterization of the bottom boundary layer in a coastal upwelling system (Ría de Vigo, NW Spain). Cont. Shelf Res. 2013, 68, 67–79. [Google Scholar] [CrossRef]
- Blanco-Bercial, L.; Álvarez-Marqués, F.; Cabal, J.A. Changes in the mesozooplankton community associated with the hidrographic off the northwestern Iberian Peninsula. ICES J. Mar. Sci. 2006, 63, 799–810. [Google Scholar] [CrossRef]
- Bode, A.; Alvarez-Ossorio, M.T.; Cabanas, J.M.; Miranda, A.; Varela, M. Recent trends in plankton and upwelling intensity off Galicia (NW Spain). Prog. Oceanogr. 2009, 83, 342–350. [Google Scholar] [CrossRef]
- Buttay, L.; Miranda, A.; Casas, G.; González-Quirós, R.; Nogueira, E. Long-term and seasonal zooplankton dynamics in the northwest Iberian shelf and its relationship with meteo-climatic and hydrographic variability. J. Plankton Res. 2015, 38, 106–121. [Google Scholar] [CrossRef]
- Olmos, L. Ecology of Cephalopod Paralarvae in a Seasonal Upwelling System. Ph.D. Thesis, Universidade de Vigo, Vigo, Spain, 2018. [Google Scholar]
- Uye, S.; Sano, K. Seasonal reproductive biology of the small cyclopoid copepod Oithona davisae in a temperate eutrophic inlet. Mar. Ecol. Prog. Ser. 1995, 118, 121–128. [Google Scholar] [CrossRef]
- Fraga, S.; Anderson, D.M.; Bravo, I.; Reguera, B.; Steidinger, K.A.; Yentsch, C.M. Influence of upwelling relaxation on dinoflagellates and shellfish toxicity in Ria de Vigo, Spain. Estuar. Coast. Shelf Sci. 1988, 27, 349–361. [Google Scholar] [CrossRef]
- Crespo, B.G.; Figueiras, F.G.; Porras, P.; Teixeira, I. Downwelling and dominance of autochthonous dinoflagellates in the NW Iberian margin: The example of the Ría de Vigo. Harmful Algae 2006, 5, 770–781. [Google Scholar] [CrossRef]
- Uye, S.; Sano, K. Seasonal variations in biomass, growth rate and production rate of the small cyclopoid copepod Oithona davisae in a temperate eutrophic inlet. Estuar. Coast. Shelf Sci. 1998, 163, 37–44. [Google Scholar]
- Martens, P.; Beusekom, J.E.E. Zooplankton response to a warmer northern Wadden Sea. Helgol. Mar. Res. 2008, 62, 67–75. [Google Scholar] [CrossRef]
- Álvarez-Salgado, X.A.; Labarta, U.; Fernández-Reiriz, M.J.; Figueiras, F.; Rosón, G.; Piedracoba, S.; Filgueira, R.; Cabanas, J.M. Renewal time and the impact of harmful algal blooms on the extensive mussel raft culture of the Iberian coastal upwelling system (SW Europe). Harmful Algae 2008, 7, 849–855. [Google Scholar] [CrossRef]
- Sanchez-Velasco, L. Diet composition and feeding habits of fish larvae of two co-occurring species (Pisces: Callionymidae and Bothidae) in the North-western Mediterranean. ICES J. Mar. Sci. 1998, 55, 299–308. [Google Scholar] [CrossRef]
- Uye, S.-I. Replacement of large copepods by small ones with eutrophication of embayments: Cause and consequence. Hydrobiologia 1994, 292, 513–519. [Google Scholar] [CrossRef]
- Belmonte, G.; Cavallo, A. Body size and its variability in the copepod Acartia margalefi (Calanoida) from Lake Acquatina (SE Italy). Ital. J. Zoól. 1997, 64, 377–382. [Google Scholar] [CrossRef]
- Belmonte, G.; Rubino, F. Cysts and Resting Eggs from Marine Zooplankton: Dimension of the Phenomenon, Physiology of Rest, and Ecological and Biogeographic Implications. In Dormancy in Aquatic Organisms. Theory, Human Use and Modeling; Editors Taylor & Francis: Abingdon, UK, 2019; pp. 71–94. [Google Scholar] [CrossRef]
- Arzul, I.; Chollet, B.; Boyer, S.; Bonnet, D.; Gaillard, J.; Baldi, Y.; Robert, M.; Joly, J.P.; Garcia, C.; Bouchoucha, M. Contribution to the understanding of the cycle of the protozoan parasite Marteilia refringens. Parasitology 2014, 141, 227–240. [Google Scholar] [CrossRef]
- Carrasco, N.; López-Flores, I.; Alcaraz, M.; Furones, M.D.; Berthe, F.C.J.; Arzul, I. Dynamics of the parasite Marteilia refringens (Paramyxea) in Mytilus galloprovincialis and zooplankton populations in Alfacs Bay (Catalonia, Spain). Parasitology 2007, 134, 1541–1550. [Google Scholar] [CrossRef]
- Foster, R.; Peeler, E.; Bojko, J.; Clark, P.F.; Morritt, D.; Roy, H.E.; Stebbing, P.; Tidbury, H.J.; Wood, L.E.; Bass, D. Pathogens co-transported with invasive non-native aquatic species: Implications for risk analysis and legislation. NeoBiota 2021, 69, 79–102. [Google Scholar] [CrossRef]
- Gregori, M.; Aznar, F.J.; Abollo, E.; Roura, Á.; González, Á.F.; Pascual, S. Nyctiphanes couchii as intermediate host for the acanthocephalan Bolbosoma balaenae in temperate waters of the NE Atlantic. Dis. Aquat. Org. 2012, 99, 37–47. [Google Scholar] [CrossRef] [PubMed]
- Gregori, M.; Aznar, F.J.; Abollo, E.; Roura, A.; González, A.F.; Pascual, S. Nyctiphanes couchii as intermediate host for Rhadinorhynchus sp. (Acanthocephala, Echinorhynchidae) from NW Iberian Peninsula waters. Dis. Aquat. Org. 2013, 105, 9–20. [Google Scholar] [CrossRef] [PubMed]
- Gregori, M.; Roura, Á.; Abollo, E.; González, Á.F.; Pascual, S. Anisakis simplex complex (Nematoda: Anisakidae) in zooplankton communities from temperate NE Atlantic waters. J. Nat. Hist. 2015, 49, 755–773. [Google Scholar] [CrossRef]
- Gregori, M.; Fernández-Leborans, G.; Roura, A.; Abollo, E.; González, Á.F.; Pascual, S. Description of a new epibiotic relationship (Suctorian-Copepoda) in NE Atlantic waters: From morphological to phylogenetic analyses. Acta Zoól. 2016, 97, 165–176. [Google Scholar] [CrossRef]
- Peeler, E.J.; Oidtmann, B.C.; Midtlyng, P.J.; Miossec, L.; Gozlan, R.E. Non-native aquatic animals introductions have driven disease emergence in Europe. Biol. Invasions 2011, 13, 1291–1303. [Google Scholar] [CrossRef]
- Carnegie, R.B.; Arzul, I.; Bushek, D. Managing marine mollusc diseases in the context of regional and international commerce: Policy issues and emerging concerns. Philos. Trans. R. Soc. B Biol. Sci. 2016, 371, 20150215. [Google Scholar] [CrossRef]
- Roy, H.E.; Hesketh, H.; Purse, B.V.; Eilenberg, J.; Santini, A.; Scalera, R.; Stentiford, G.D.; Adriaens, T.; Bacela-Spychalska, K.; Bass, D.; et al. Alien Pathogens on the Horizon: Opportunities for Predicting their Threat to Wildlife. Conserv. Lett. 2016, 10, 477–484. [Google Scholar] [CrossRef]
- Villalba, A.; Iglesias, D.; Ramilo, A.; Darriba, S.; Parada, J.B.; No, E.; Abollo, E.; Molares, J.; Carballal, M.J. Cockle Cerastoderma edule fishery collapse in the Ría de Arousa (Galicia, NW Spain) associated with the protistan parasite Marteilia cochillia. Dis. Aquat. Org. 2014, 109, 55–80. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Species | N | Accession Number |
|---|---|---|
| O. atlantica | 12 | KP033173–KP033176; JF419541–JF419543; ON114103; ON114104; ON114106–ON114108 |
| O. plumifera | 2 | KP033177–KP0331778 |
| O. frigida | 1 | KU982713 |
| O. similis | 55 | JF419529–JF419540; FM991723; EF460779; KF153700; KX495626; KP033160–KP033162; KU982646–KU982648; KU982650–KU982651; KU982655; KU982657; KU982659-KU982660; KU982663; KU982665; KU982671–KU982674; KU982676; KU982678–KU982686; KU982702–KU982703; KU982694; KU982699; KU982705–KU982711 |
| O. dissimilis | 12 | AB604177–AB604188 |
| O. brevicornis | 2 | AB604189–AB604190 |
| O. davisae | 15 | KP033163–KP033167; ON114109–ON114118 |
| O. nana | 11 | FM991725–FM991727; JF419544; JF419546–JF419547; KP033168–KP033172 |
| O. simplex | 1 | AF385458 |
| Intraspecific Distances | Interspecific Distances | ||
|---|---|---|---|
| O. atlantica Gal | O. davisae Gal | ||
| O. davisae Gal | 0–0.005 | - | - |
| O. davisae | 0–0.002 | 0.120–0.130 | 0–0.005 |
| O. atlantica Gal | 0–0.012 | - | 0.120–0.133 |
| O. atlantica | 0–0.019 | 0–0.022 | 0.117–0.126 |
| O. plumifera | 0.000 | 0.014–0.027 | 0.126–0.129 |
| O. frigida | - | 0.012–0.024 | 0.120–0.146 |
| O. similis | 0–0.010 | 0.025–0.043 | 0.138–0.146 |
| O. dissimilis | 0–0.020 | 0.103–0.125 | 0.134–0.145 |
| O. brevicornis | 0.002 | 0.130–0.147 | 0.156–0.168 |
| O. nana | 0–0.027 | 0.180–0.220 | 0.195–0.230 |
| O. simplex | - | 0.130–0.147 | 0.156–0.163 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
García-Alves, L.; Ramilo, A.; Pascual, S.; González, Á.F.; Abollo, E. Coexistence of Two Copepods, Recorded for the First Time, in NW Iberian Shelf: The Case of Oithona atlantica and the Allochthonous O. davisae. Diversity 2024, 16, 61. https://doi.org/10.3390/d16010061
García-Alves L, Ramilo A, Pascual S, González ÁF, Abollo E. Coexistence of Two Copepods, Recorded for the First Time, in NW Iberian Shelf: The Case of Oithona atlantica and the Allochthonous O. davisae. Diversity. 2024; 16(1):61. https://doi.org/10.3390/d16010061
Chicago/Turabian StyleGarcía-Alves, Lara, Andrea Ramilo, Santiago Pascual, Ángel F. González, and Elvira Abollo. 2024. "Coexistence of Two Copepods, Recorded for the First Time, in NW Iberian Shelf: The Case of Oithona atlantica and the Allochthonous O. davisae" Diversity 16, no. 1: 61. https://doi.org/10.3390/d16010061