
Characterization of Lacinutrix neustonica sp. nov., Isolated from the Sea Surface Microlayer of Brackish Lake Shihwa, South Korea
 , , and
, , and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Habitat, Isolation and Cultivation Conditions
2.2. 16S rRNA Gene Sequencing and Phylogenetic Analysis
2.3. Genome Sequencing, Assembly, Annotation, and Phylogenomic Analysis
2.4. Physiological, Morphological and Biochemical Characteristics
3. Results and Discussion
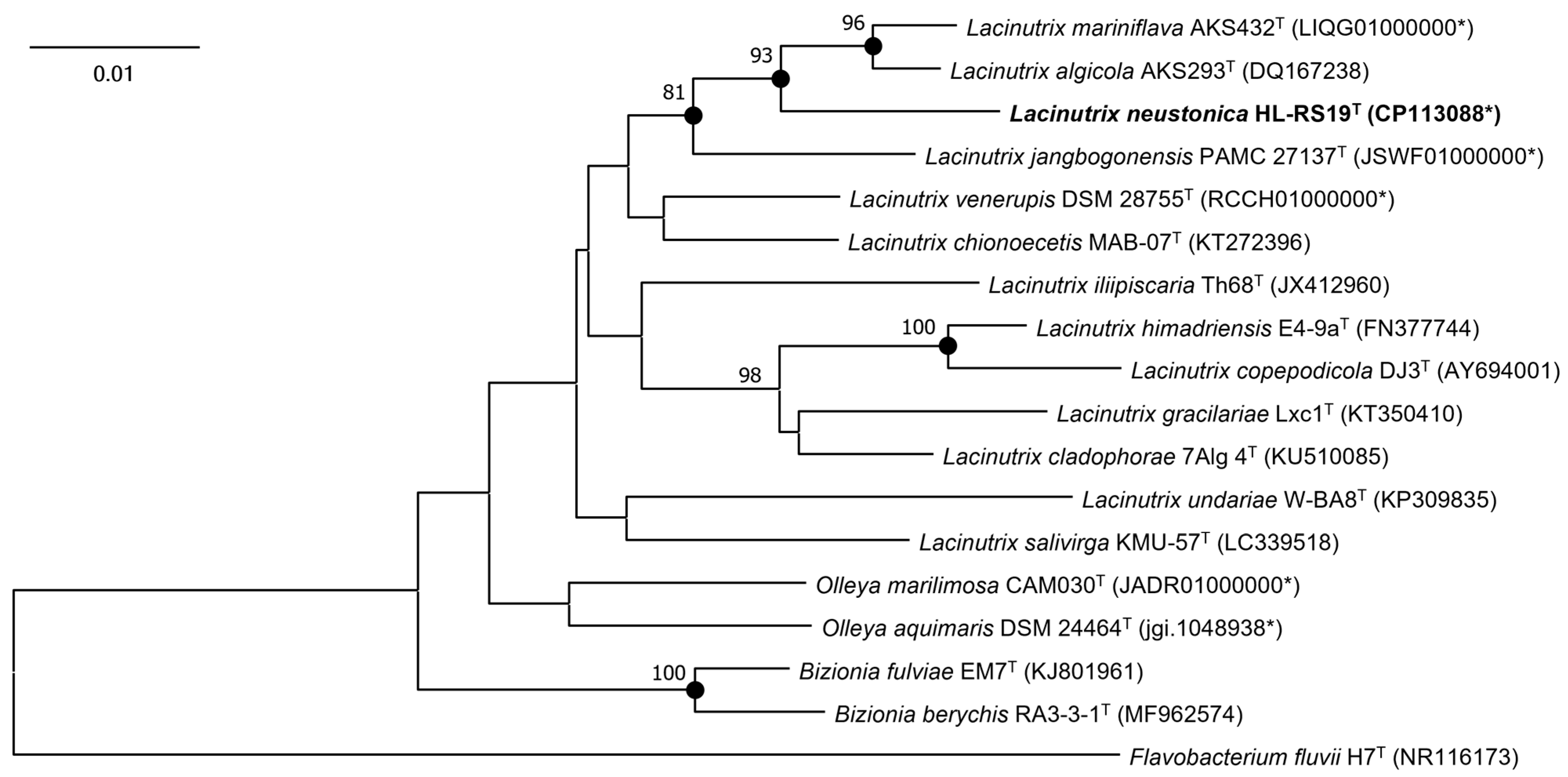
3.1. 16S rRNA Gene Sequencing and Phylogenetic Analysis
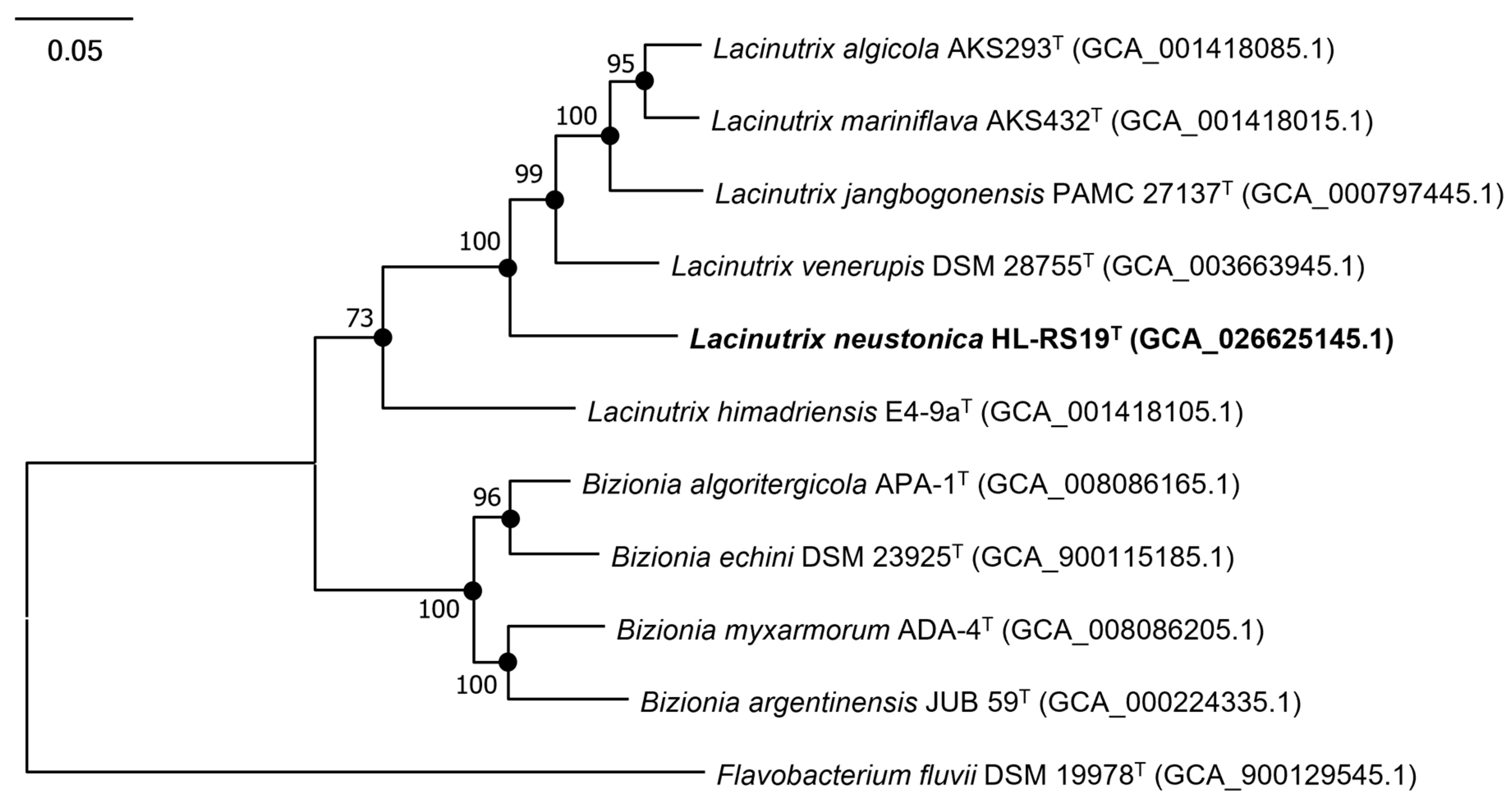
3.2. Genome Analysis and Genomic Features
3.3. Physiological, Morphological and Biochemical Characteristics
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Conflicts of Interest
References
- Hunter, K.A. Processes affecting particulate trace metals in the sea surface microlayer. Mar. Chem. 1980, 9, 49–70. [Google Scholar] [CrossRef]
- Zancker, B.; Cunliffe, M.; Engel, A. Bacterial community composition in the sea surface microlayer off the peruvian coast. Front. Microbiol. 2018, 9, 2699. [Google Scholar] [CrossRef] [PubMed]
- Carlucci, A.F.; Wolgast, D.M.; Craven, D.B. Microbial populations in surface films: Amino acid dynamics in nearshore and offshore waters off Southern California. J. Geophys. Res. Oceans 1992, 92, 5271–5280. [Google Scholar] [CrossRef]
- Cunliffe, M.; Engel, A.; Frka, S.; Gasparovic, B.; Guitart, C.; Murrell, J.C.; Salter, M.; Stolle, C.; Upstill-Goddard, R.; Wurl, O. Sea surface microlayers: A unified physicochemical and biological perspective of the air-ocean interface. Prog. Oceanogr. 2013, 109, 104–116. [Google Scholar] [CrossRef]
- Wurl, O.; Obbard, J.P. A review of pollutants in the sea-surface microlayer (SML): A unique habitat for marine organisms. Mar. Pollut. Bull. 2004, 48, 1016–1030. [Google Scholar] [CrossRef] [PubMed]
- Cincinelli, A.; Stortini, A.M.; Checchini, L.; Martellini, T.; Del Bubba, M.; Lepri, L. Enrichment of organic pollutants in the sea surface microlayer (SML) at Terra Nova Bay, Antarctica: Influence of SML on superfacial snow composition. J. Environ. Monitor. 2005, 7, 1305–1312. [Google Scholar] [CrossRef] [PubMed]
- Hardy, J.T.; Crecelius, E.A.; Antrim, L.D.; Kiesser, S.L.; Broadhurst, V.L.; Boehm, P.D.; Steinhauer, W.G.; Coogan, T.H. Aquatic surface microlayer contamination in Chesapeake Bay. Mar. Chem. 1990, 28, 333–351. [Google Scholar] [CrossRef]
- Williams, P.M.; Carlucci, A.F.; Henrichs, S.M.; Vanvleet, E.S.; Horrigan, S.G.; Reid, F.M.H.; Robertson, K.J. Chemical and microbiological studies of sea-surface films in the Southern Gulf of California and off the West-Coast of Baja-California. Mar. Chem. 1986, 19, 17–98. [Google Scholar] [CrossRef]
- Hardy, J.T.; Apts, C.W.; Crecelius, E.A.; Bloom, N.S. Sea-surface microlayer metals enrichments in an urban and rural bay. Estuar. Coast. Shelf Sci. 1985, 20, 299–312. [Google Scholar] [CrossRef]
- Lee, I.; Jang, G.I.; Cho, Y.; Yoon, S.J.; Pham, H.M.; Nguyen, A.V.; Lee, Y.M.; Park, H.; Rhee, T.S.; Kim, S.H.; et al. Sandaracinobacter neustonicus sp. nov., isolated from the sea surface microlayer in the Southwestern Pacific Ocean, and emended description of the genus Sandaracinobacter. Int. J. Syst. Evol. Microbiol. 2020, 70, 4698–4703. [Google Scholar] [CrossRef] [PubMed]
- Jang, G.I.; Lee, I.; Ha, T.T.; Yoon, S.J.; Hwang, Y.J.; Yi, H.; Yun, S.; Lee, W.S.; Hwang, C.Y. Pseudomonas neustonica sp. nov., isolated from the sea surface microlayer of the Ross Sea (Antarctica). Int. J. Syst. Evol. Microbiol. 2020, 70, 3832–3838. [Google Scholar] [CrossRef]
- Hwang, C.Y.; Lee, I.; Hwang, Y.J.; Yoon, S.J.; Lee, W.S.; Cho, B.C. Pseudoalteromonas neustonica sp. nov., isolated from the sea surface microlayer of the Ross Sea (Antarctica), and emended description of the genus Pseudoalteromonas. Int. J. Syst. Evol. Microbiol. 2016, 66, 3377–3382. [Google Scholar] [CrossRef] [PubMed]
- Wong, S.K.; Yoshizawa, S.; Nakajima, Y.; Cuadra, M.J.; Nogi, Y.; Nakamura, K.; Takami, H.; Ogura, Y.; Hayashi, T.; Chiura, H.X.; et al. Amylibacter kogurei sp. nov., a novel marine alphaproteobacterium isolated from the coastal sea surface microlayer of a marine inlet. Int. J. Syst. Evol. Microbiol. 2018, 68, 2872–2877. [Google Scholar] [CrossRef]
- Urios, L.; Agogue, H.; Intertaglia, L.; Lesongeur, F.; Lebaron, P. Melitea salexigens gen. nov., sp. nov., a gammaproteobacterium from the Mediterranean Sea. Int. J. Syst. Evol. Microbiol. 2008, 58, 2479–2483. [Google Scholar] [CrossRef]
- Urios, L.; Intertaglia, L.; Lesongeur, F.; Lebaron, P. Haliea salexigens gen. nov., sp. nov., a member of the Gammaproteobacteria from the Mediterranean Sea. Int. J. Syst. Evol. Microbiol. 2008, 58, 1233–1237. [Google Scholar] [CrossRef]
- Bowman, J.P.; Nichols, D.S. Novel members of the family Flavobacteriaceae from Antarctic maritime habitats including Subsaximicrobium wynnwilliamsii gen. nov., sp. nov., Subsaximicrobium saxinquilinus sp. nov., Subsaxibacter broadyi gen. nov., sp. nov., Lacinutrix copepodicola gen. nov., sp. nov., and novel species of the genera Bizionia, Gelidibacter and Gillisia. Int. J. Syst. Evol. Microbiol. 2005, 55, 1471–1486. [Google Scholar] [CrossRef]
- Parte, A.C.; Carbasse, J.S.; Meier-Kolthoff, J.P.; Reimer, L.C.; Goker, M. List of Prokaryotic names with Standing in Nomenclature (LPSN) moves to the DSMZ. Int. J. Syst. Evol. Microbiol. 2020, 70, 5607–5612. [Google Scholar] [CrossRef]
- Bowman, J.P. Lacinutrix. In Bergey’s Manual of Systematics of Archaea and Bacteria; Wiley: Hoboken, NJ, USA, 2015; pp. 1–3. [Google Scholar]
- Nedashkovskaya, O.I.; Kim, S.G.; Zhukova, N.V.; Lee, J.S.; Mikhailov, V.V. Lacinutrix cladophorae sp. nov., a flavobacterium isolated from the green alga Cladophora stimpsonii, transfer of Flavirhabdus iliipiscaria Shakeela et al. 2015 to the genus Lacinutrix as Lacinutrix iliipiscaria comb. nov. and emended description of the genus Lacinutrix. Int. J. Syst. Evol. Microbiol. 2016, 66, 4339–4346. [Google Scholar] [CrossRef] [PubMed]
- Srinivas, T.N.; Prasad, S.; Manasa, P.; Sailaja, B.; Begum, Z.; Shivaji, S. Lacinutrix himadriensis sp. nov., a psychrophilic bacterium isolated from a marine sediment, and emended description of the genus Lacinutrix. Int. J. Syst. Evol. Microbiol. 2013, 63, 729–734. [Google Scholar] [CrossRef]
- Huang, Z.; Li, G.; Lai, Q.; Gu, L.; Shao, Z. Lacinutrix gracilariae sp. nov., isolated from the surface of a marine red alga Gracilaria sp. Int. J. Syst. Evol. Microbiol. 2016, 66, 587–591. [Google Scholar] [CrossRef]
- Kim, H.; Yoon, S.C.; Choi, K.H.; Kim, S.T.; Lee, J.B.; Kim, D.S.; Han, H.L.; Bae, K.S.; Park, D.S. Lacinutrix chionocetis sp. nov., isolated from gut of a red snow crab. Arch. Microbiol. 2017, 199, 597–603. [Google Scholar] [CrossRef]
- Lasa, A.; Dieguez, A.L.; Romalde, J.L. Description of Lacinutrix venerupis sp. nov.: A novel bacterium associated with reared clams. Syst. Appl. Microbiol. 2015, 38, 115–119. [Google Scholar] [CrossRef]
- Lee, Y.M.; Hwang, C.Y.; Lee, I.; Jung, Y.J.; Cho, Y.; Baek, K.; Hong, S.G.; Kim, J.; Chun, J.; Lee, H.K. Lacinutrix jangbogonensis sp. nov., a psychrophilic bacterium isolated from Antarctic marine sediment and emended description of the genus Lacinutrix. Anton. Leeuw. 2014, 106, 527–533. [Google Scholar] [CrossRef]
- Nedashkovskaya, O.I.; Kwon, K.K.; Yang, S.H.; Lee, H.S.; Chung, K.H.; Kim, S.J. Lacinutrix algicola sp. nov. and Lacinutrix mariniflava sp. nov., two novel marine alga-associated bacteria and emended description of the genus Lacinutrix. Int. J. Syst. Evol. Microbiol. 2008, 58, 2694–2698. [Google Scholar] [CrossRef]
- Park, S.; Park, J.M.; Jung, Y.T.; Kang, C.H.; Yoon, J.H. Lacinutrix undariae sp. nov., isolated from a brown algae reservoir. Int. J. Syst. Evol. Microbiol. 2015, 65, 2696–2701. [Google Scholar] [CrossRef] [PubMed]
- Yoon, J.; Lee, J.S.; Lee, K.C. Description of Lacinutrix salivirga sp. nov., a marine member of the family Flavobacteriaceae isolated from seawater. Arch. Microbiol. 2018, 200, 1159–1165. [Google Scholar] [CrossRef] [PubMed]
- Shakeela, Q.; Shehzad, A.; Zhang, Y.; Tang, K.; Zhang, X.H. Flavirhabdus iliipiscaria gen. nov., sp. nov., isolated from intestine of flounder (Paralichthys olivaceus) and emended descriptions of the genera Flavivirga, Algibacter, Bizionia and Formosa. Int. J. Syst. Evol. Microbiol. 2015, 65, 1347–1353. [Google Scholar] [CrossRef]
- Englen, M.D.; Kelley, L.C. A rapid DNA isolation procedure for the identification of Campylobacter jejuni by the polymerase chain reaction. Lett. Appl. Microbiol. 2000, 31, 421–426. [Google Scholar] [CrossRef] [PubMed]
- Lane, D.J. 16S/23S rRNA Sequencing. In Nucleic Acid Techniques in Bacterial Systematics, 2nd ed.; Stackebrandt, E., Goodfellow, M., Eds.; John Wiley and Sons: Chichester, UK, 1991; pp. 115–175. [Google Scholar]
- Anzai, Y.; Kudo, Y.; Oyaizu, H. The phylogeny of the genera Chryseomonas, Flavimonas, and Pseudomonas supports synonymy of these three genera. Int. J. Syst. Bacteriol. 1997, 47, 249–251. [Google Scholar] [CrossRef] [PubMed]
- Yoon, S.H.; Ha, S.M.; Kwon, S.; Lim, J.; Kim, Y.; Seo, H.; Chun, J. Introducing EzBioCloud: A taxonomically united database of 16S rRNA gene sequences and whole-genome assemblies. Int. J. Syst. Evol. Microbiol. 2017, 67, 1613–1617. [Google Scholar] [CrossRef] [PubMed]
- Jeon, Y.S.; Lee, K.; Park, S.C.; Kim, B.S.; Cho, Y.J.; Ha, S.M.; Chun, J. EzEditor: A versatile sequence alignment editor for both rRNA- and protein-coding genes. Int. J. Syst. Evol. Microbiol. 2014, 64, 689–691. [Google Scholar] [CrossRef]
- Tamura, K.; Stecher, G.; Kumar, S. MEGA11: Molecular evolutionary genetics analysis version 11. Mol. Biol. Evol. 2021, 38, 3022–3027. [Google Scholar] [CrossRef] [PubMed]
- Saitou, N.; Nei, M. The neighbor-joining method: A new method for reconstructing phylogenetic trees. Mol. Biol. Evol. 1987, 4, 406–425. [Google Scholar] [CrossRef] [PubMed]
- Jukes, T.H.; Cantor, C.R. Evolution of Protein Molecules. In Mammalian Protein Metabolism; Academic Press: New York, NY, USA, 1969; Volume 3, pp. 21–132. [Google Scholar]
- Felsenstein, J. Evolutionary trees from DNA-Sequences—A maximum-likelihood approach. J. Mol. Evol. 1981, 17, 368–376. [Google Scholar] [CrossRef] [PubMed]
- Kimura, M.A. Simple method for estimating evolutionary rates of base substitutions through comparative studies of nucleotide sequences. J. Mol. Evol. 1980, 16, 111–120. [Google Scholar] [CrossRef] [PubMed]
- Fitch, W.M. Toward defining the course of evolution: Minimum change for a specific tree topology. Syst. Biol. 1971, 20, 406–416. [Google Scholar] [CrossRef]
- Nei, M.; Kumar, S. Molecular Evolution and Phylogenetics; Oxford University Press: New York, NY, USA, 2000. [Google Scholar]
- Felsenstein, J. Confidence limits on phylogenies: An approach using the bootstrap. Evolution 1985, 39, 783–791. [Google Scholar] [CrossRef]
- Kolmogorov, M.; Yuan, J.; Lin, Y.; Pevzner, P.A. Assembly of long, error-prone reads using repeat graphs. Nat. Biotechnol. 2019, 37, 540–546. [Google Scholar] [CrossRef] [PubMed]
- Mikheenko, A.; Prjibelski, A.; Saveliev, V.; Antipov, D.; Gurevich, A. Versatile genome assembly evaluation with QUAST-LG. Bioinformatics 2018, 34, 142–150. [Google Scholar] [CrossRef]
- Danecek, P.; Bonfield, J.K.; Liddle, J.; Marshall, J.; Ohan, V.; Pollard, M.O.; Whitwham, A.; Keane, T.; McCarthy, S.A.; Davies, R.M.; et al. Twelve years of SAMtools and BCFtools. Gigascience 2021, 10, giab008. [Google Scholar] [CrossRef]
- Yoon, S.H.; Ha, S.M.; Lim, J.; Kwon, S.; Chun, J. A large-scale evaluation of algorithms to calculate average nucleotide identity. Anton. Leeuw. 2017, 110, 1281–1286. [Google Scholar] [CrossRef] [PubMed]
- Meier-Kolthoff, J.P.; Carbasse, J.S.; Peinado-Olarte, R.L.; Göker, M. TYGS and LPSN: A database tandem for fast and reliable genome-based classification and nomenclature of prokaryotes. Nucleic Acids Res. 2021, 50, D801–D807. [Google Scholar] [CrossRef] [PubMed]
- Lee, I.; Chalita, M.; Ha, S.M.; Na, S.I.; Yoon, S.H.; Chun, J. ContEst16S: An algorithm that identifies contaminated prokaryotic genomes using 16S RNA gene sequences. Int. J. Syst. Evol. Microbiol. 2017, 67, 2053–2057. [Google Scholar] [CrossRef]
- Chaumeil, P.A.; Mussig, A.J.; Hugenholtz, P.; Parks, D.H. GTDB-Tk: A toolkit to classify genomes with the Genome Taxonomy Database. Bioinformatics 2019, 36, 1925–1927. [Google Scholar] [CrossRef] [PubMed]
- Tatusova, T.; DiCuccio, M.; Badretdin, A.; Chetvernin, V.; Nawrocki, E.P.; Zaslavsky, L.; Lomsadze, A.; Pruitt, K.; Borodovsky, M.; Ostell, J. NCBI prokaryotic genome annotation pipeline. Nucleic Acids Res. 2016, 44, 6614–6624. [Google Scholar] [CrossRef] [PubMed]
- Caspi, R.; Billington, R.; Keseler, I.M.; Kothari, A.; Krummenacker, M.; Midford, P.E.; Ong, W.K.; Paley, S.; Subhraveti, P.; Karp, P.D. The MetaCyc database of metabolic pathways and enzymes—A 2019 update. Nucleic Acids Res. 2020, 48, D445–D453. [Google Scholar] [CrossRef] [PubMed]
- Karp, P.D.; Billington, R.; Caspi, R.; Fulcher, C.A.; Latendresse, M.; Kothari, A.; Keseler, I.M.; Krummenacker, M.; Midford, P.E.; Ong, Q.; et al. The BioCyc collection of microbial genomes and metabolic pathways. Brief Bioinform. 2019, 20, 1085–1093. [Google Scholar] [CrossRef]
- Blin, K.; Shaw, S.; Kloosterman, A.M.; Charlop-Powers, Z.; Van Wezel, G.P.; Medema, M.H.; Weber, T. antiSMASH 6.0: Improving cluster detection and comparison capabilities. Nucleic Acids Res. 2021, 49, W29–W35. [Google Scholar] [CrossRef]
- Liu, D.; Zhang, Y.; Fan, G.; Sun, D.; Zhang, X.; Yu, Z.; Wang, J.; Wu, L.; Shi, W.; Ma, J. IPGA: A handy integrated prokaryotes genome and pan-genome analysis web service. iMeta 2022, 1, e55. [Google Scholar] [CrossRef]
- Bernardet, J.F.; Nakagawa, Y.; Holmes, B. Proposed minimal standards for describing new taxa of the family Flavobacteriaceae and emended description of the family. Int. J. Syst. Evol. Microbiol. 2002, 52, 1049–1070. [Google Scholar] [CrossRef]
- Lee, Y.S.; Lee, D.H.; Kahng, H.Y.; Sohn, S.H.; Jung, J.S. Polaribacter gangjinensis sp. nov., isolated from seawater. Int. J. Syst. Evol. Microbiol. 2011, 61, 1425–1429. [Google Scholar] [CrossRef] [PubMed]
- Yoon, J.H.; Kang, S.J.; Jung, Y.T.; Lee, M.H.; Oh, T.K. Alkalibacillus flavidus sp. nov., isolated from a marine solar saltern. Int. J. Syst. Evol. Microbiol. 2010, 60, 434–438. [Google Scholar] [CrossRef]
- Jordan, E.O.; Caldwell, M.E.; Reiter, D. Bacterial motility. J. Bacteriol. 1934, 27, 165–174. [Google Scholar] [CrossRef]
- Skerman, V.B.D. A Guide to the Identification of the Genera of Bacteria, with Methods and Digests of Generic Characteristics, 2nd ed.; Williams & Wilkins Co.: Baltimore, MD, USA, 1967. [Google Scholar]
- Gosink, J.J.; Woese, C.R.; Staley, J.T. Polaribacter gen. nov., with three new species, P. irgensii sp. nov., P. franzmannii sp. nov., and P. filamentus sp. nov., gas vacuolate polar marine bacteria of the Cytophaga-Flavobacterium-Bacteroides group and reclassification of ‘Flectobacillus glomeratus’ as Polaribacter glomeratus comb. nov. Int. J. Syst. Evol. Microbiol. 1998, 48, 223–235. [Google Scholar] [CrossRef]
- Fautz, E.; Reichenbach, H. A simple test for flexirubin-type pigments. Fems Microbiol. Lett. 1980, 8, 87–91. [Google Scholar] [CrossRef]
- Tindall, B.J.; Sikorski, J.; Smibert, R.A.; Krieg, N.R. Phenotypic Characterization and the Principles of Comparative Systematics. In Methods for General and Molecular Microbiology; Reddy, C.A., Beveridge, T.J., Breznak, J.A., Marzluf, G., Schmidt, T.M., Snyder, L.R., Eds.; American Society for Microbiology: Washington, DC, USA, 2007; pp. 330–393. [Google Scholar] [CrossRef]
- Smibert, R.M.; Krieg, N.R. Methods for General and Molecular Bacteriology; American Society for Microbiology: Washington, DC, USA, 1994; pp. 607–654. [Google Scholar]
- Cowan, S.T.; Steel, K.J. Manual for the Identification of Medical Bacteria; Cambridge University Press: London, UK, 1965. [Google Scholar]
- Lányi, B. Classical and Rapid Identification Methods for Medically Important Bacteria. In Current Methods for Classification and Idenfication of Mircroorganisms; Colwell, R.R., Grigorova, R., Eds.; Academic Press: London, UK, 1988; Volume 19, pp. 1–67. [Google Scholar]
- Komagata, K.; Suzuki, K. Lipid and cell-wall analysis in bacterial systematics. Methods Microbiol. 1987, 19, 161–207. [Google Scholar] [CrossRef]
- Minnikin, D.E.; Odonnell, A.G.; Goodfellow, M.; Alderson, G.; Athalye, M.; Schaal, A.; Parlett, J.H. An integrated procedure for the extraction of bacterial isoprenoid quinones and polar lipids. J. Microbiol. Meth. 1984, 2, 233–241. [Google Scholar] [CrossRef]
- Collins, M.D. Analysis of isoprenoid quinones. Methods Microbiol. 1985, 18, 329–366. [Google Scholar] [CrossRef]
- Goris, J.; Konstantinidis, K.T.; Klappenbach, J.A.; Coenye, T.; Vandamme, P.; Tiedje, J.M. DNA–DNA hybridization values and their relationship to whole genome sequence similarities. Int. J. Syst. Evol. Microbiol. 2007, 57, 81–91. [Google Scholar] [CrossRef] [PubMed]
- Dieser, M.; Greenwood, M.; Foreman, C.M. Carotenoid pigmentation in Antarctic heterotrophic bacteria as a strategy to withstand environmental stresses. Arct. Antarct. Alp. Res. 2010, 42, 396–405. [Google Scholar] [CrossRef]
- Stafsnes, M.H.; Josefsen, K.D.; Kildahl-Andersen, G.; Valla, S.; Ellingsen, T.E.; Bruheim, P. Isolation and characterization of marine pigmented bacteria from Norwegian coastal waters and screening for carotenoids with UVA-blue light absorbing properties. J. Microbiol. 2010, 48, 16–23. [Google Scholar] [CrossRef] [PubMed]
- Armstrong, G.A. Genetics of eubacterial carotenoid biosynthesis: A colorful tale. Annu. Rev. Microbiol. 1997, 51, 629–659. [Google Scholar] [CrossRef] [PubMed]
- Misawa, N.; Satomi, Y.; Kondo, K.; Yokoyama, A.; Kajiwara, S.; Saito, T.; Ohtani, T.; Miki, W. Structure and functional-analysis of a marine bacterial carotenoid biosynthesis gene-cluster and astaxanthin biosynthetic pathway proposed at the gene level. J. Bacteriol. 1995, 177, 6575–6584. [Google Scholar] [CrossRef]
- Crowley, D.J.; Boubriak, I.; Berquist, B.R.; Clark, M.; Richard, E.; Sullivan, L.; DasSarma, S.; McCready, S. The uvrA, uvrB and uvrC genes are required for repair of ultraviolet light induced DNA photoproducts in Halobacterium sp. NRC-1. Saline Syst. 2006, 2, 11. [Google Scholar] [CrossRef]
- Donaldson, J.R.; Courcelle, C.T.; Courcelle, J. RuvABC is required to resolve Holliday junctions that accumulate following replication on damaged templates in Escherichia coli. J. Biol. Chem. 2006, 281, 28811–28821. [Google Scholar] [CrossRef]
- Rosen, B.P. Families of arsenic transporters. Trends Microbiol. 1999, 7, 207–212. [Google Scholar] [CrossRef] [PubMed]
- Mateos, L.M.; Villadangos, A.F.; de la Rubia, A.G.; Mourenza, A.; Marcos-Pascual, L.; Letek, M.; Pedre, B.; Messens, J.; Gil, J.A. The arsenic detoxification system in Corynebacteria: Basis and application for bioremediation and redox control. Adv. Appl. Microbiol. 2017, 99, 103–137. [Google Scholar] [CrossRef]
- Villadangos, A.F.; Ordóñez, E.; Pedre, B.; Messens, J.; Gil, J.A.; Mateos, L.M. Engineered coryneform bacteria as a bio-tool for arsenic remediation. Appl. Microbiol. Biotechnol. 2014, 98, 10143–10152. [Google Scholar] [CrossRef]
- Feo, J.C.; Ordonez, E.; Letek, M.; Castro, M.A.; Munoz, M.I.; Gil, J.A.; Mateos, L.M.; Aller, A.J. Retention of inorganic arsenic by coryneform mutant strains. Water Resour. Res. 2007, 41, 531–542. [Google Scholar] [CrossRef]
- Sousa, T.; Branco, R.; Piedade, A.P.; Morais, P.V. Hyper accumulation of arsenic in mutants of Ochrobactrum tritici silenced for arsenite efflux pumps. PLoS ONE 2015, 10, e0131317. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| ANI/dDDH Value (%) | ||||||
|---|---|---|---|---|---|---|
| dDDH\ANI | 1 | 2 | 3 | 4 | 5 | 6 |
| 1 | − | 75.2 | 75.3 | 75.5 | 75.3 | 73.5 |
| 2 | 19.7 | − | 86.5 | 82.0 | 80.6 | 75.8 |
| 3 | 20.2 | 31.8 | − | 81.8 | 80.7 | 76.0 |
| 4 | 20.1 | 25.9 | 25.6 | − | 80.2 | 75.4 |
| 5 | 19.6 | 23.5 | 23.7 | 23.4 | − | 75.5 |
| 6 | 19.8 | 21.2 | 21.5 | 20.6 | 19.9 | − |
| Characteristics | 1 | 2 | 3 |
|---|---|---|---|
| Growth temperature range (optimum; °C) | 10–30 (20–25) | 5–20 (15–20) | 0–25 (17.5) |
| Salt tolerance range (optimum;%, w/v) | 1.0–7.0 (3.0–3.5) | 1.5–6.0 (2.5–3.0) | 0–2.5 (0.5) |
| pH range (optimum) | 6.0–8.5 (6.5–7.0) | 6.0–8.5 (6.5–7.0) | 5.5–8.5 (6.5) |
| Hydrolysis of: | |||
| Casein | − | + | + |
| Gelatin | − * | + * | + |
| Tween 40 | − | + | − |
| Tween 80 | + | + | − |
| Urea | − | − | + |
| Utilization as sole carbon source: | |||
| N-Acetyl-D-glucosamine | − | − | + |
| D-Glucose | − | + | − |
| D-Maltose | + | + | − |
| Mannitol | − | − | + |
| D-Mannose | + | − | − |
| Enzymatic activity (API ZYM) of: | |||
| β-Galactosidase | − | − | + |
| DNA G+C content (mol%) | 35.2 | 34.7 ** | 37.0 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Choi, J.Y.; Kim, S.Y.; Hong, Y.W.; Kim, B.J.; Shin, D.Y.; Kang, J.K.; Cho, B.C.; Hwang, C.Y. Characterization of Lacinutrix neustonica sp. nov., Isolated from the Sea Surface Microlayer of Brackish Lake Shihwa, South Korea. Diversity 2023, 15, 1004. https://doi.org/10.3390/d15091004
Choi JY, Kim SY, Hong YW, Kim BJ, Shin DY, Kang JK, Cho BC, Hwang CY. Characterization of Lacinutrix neustonica sp. nov., Isolated from the Sea Surface Microlayer of Brackish Lake Shihwa, South Korea. Diversity. 2023; 15(9):1004. https://doi.org/10.3390/d15091004
Chicago/Turabian StyleChoi, Jy Young, Soo Yoon Kim, Yeon Woo Hong, Bok Jin Kim, Dong Young Shin, Jin Kyeong Kang, Byung Cheol Cho, and Chung Yeon Hwang. 2023. "Characterization of Lacinutrix neustonica sp. nov., Isolated from the Sea Surface Microlayer of Brackish Lake Shihwa, South Korea" Diversity 15, no. 9: 1004. https://doi.org/10.3390/d15091004