


Ancient Plant Lineages Endemic to Africa and Its Islands: An Analysis on the Distribution and Diversity


Abstract
:1. Introduction
2. Materials and Methods
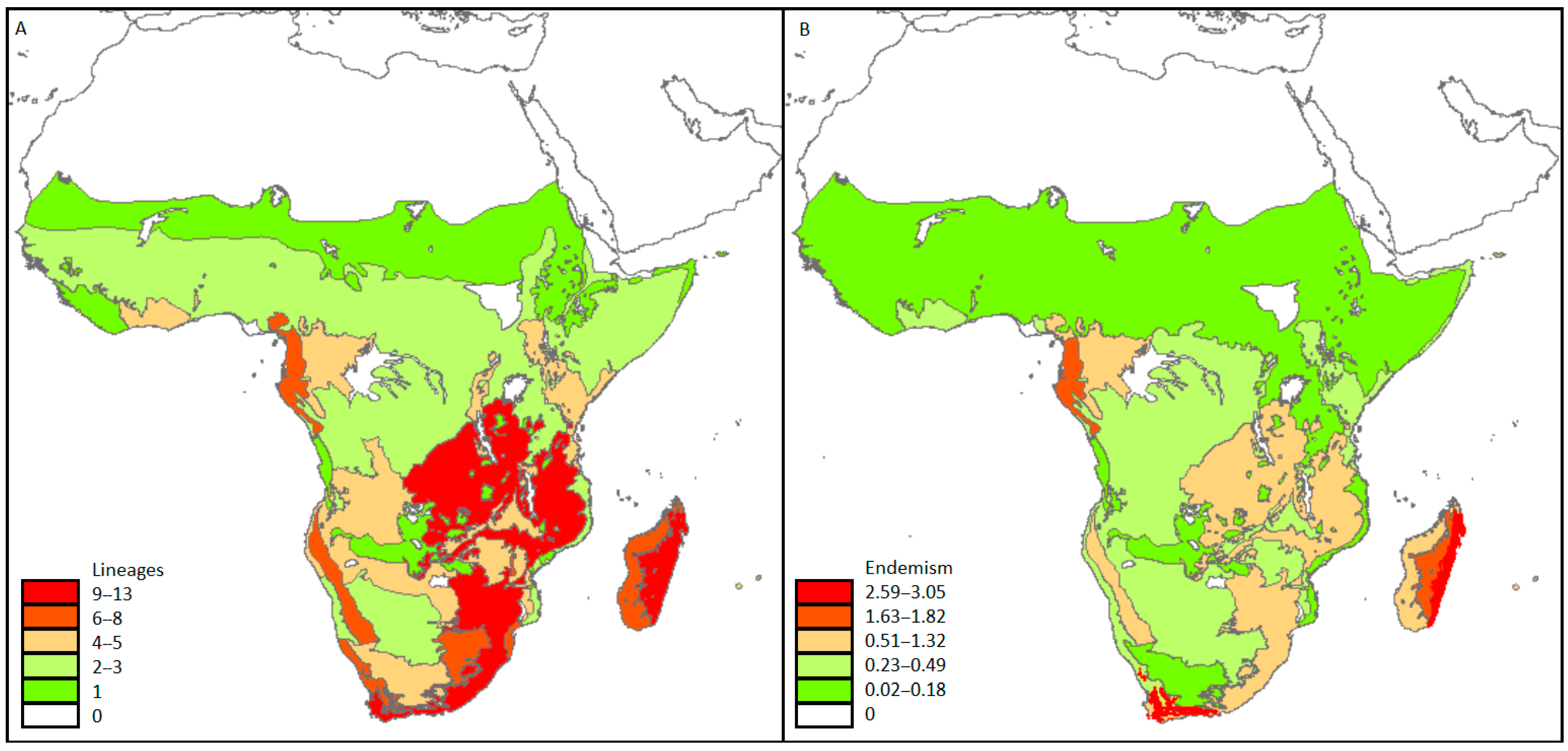
3. Results
4. Discussion
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Forest, F.; Grenyer, R.; Rouget, M.; Davies, T.J.; Cowling, R.M.; Faith, D.P.; Balmford, A.; Manning, J.C.; Procheş, Ş.; van der Bank, M.; et al. Preserving the evolutionary potential of floras in biodiversity hotspots. Nature 2007, 445, 757–760. [Google Scholar] [CrossRef] [PubMed]
- Mishler, B.D. Spatial phylogenetics. J. Biogeogr. 2023, 50, 1454–1463. [Google Scholar] [CrossRef]
- Lowenthal, D. Conserving nature and antiquity. In Man, Nature and Technology; Baark, E., Svedin, U., Eds.; Palgrave Macmillan: London, UK, 1988; pp. 122–134. [Google Scholar] [CrossRef]
- Donoghue, M.J.; Sanderson, M.J. Confluence, synnovation, and depauperons in plant diversification. New Phytol. 2015, 207, 260–274. [Google Scholar] [CrossRef] [PubMed]
- Procheş, Ş.; Ramdhani, S.; Perera, S.J.; Ali, J.R.; Gairola, S. Global hotspots in the present-day distribution of ancient animal and plant lineages. Sci. Rep. 2015, 5, 15457. [Google Scholar] [CrossRef] [PubMed]
- McLoughlin, S. The breakup history of Gondwana and its impact on pre-Cenozoic floristic provincialism. Aust. J. Bot. 2001, 49, 271–300. [Google Scholar] [CrossRef]
- Janssens, S.B.; Couvreur, T.L.; Mertens, A.; Dauby, G.; Dagallier, L.P.; Abeele, S.V.; Vandelook, F.; Mascarello, M.; Beeckman, H.; Sosef, M.; et al. A large-scale species level dated angiosperm phylogeny for evolutionary and ecological analyses. Biodivers. Data J. 2020, 8, e39677. [Google Scholar] [CrossRef] [PubMed]
- Dagallier, L.P.; Janssens, S.B.; Dauby, G.; Blach-Overgaard, A.; Mackinder, B.A.; Droissart, V.; Svenning, J.C.; Sosef, M.S.; Stévart, T.; Harris, D.J.; et al. Cradles and museums of generic plant diversity across tropical Africa. New Phytol. 2019, 225, 2196–2213. [Google Scholar] [CrossRef] [PubMed]
- Stevens, P.F. Angiosperm Phylogeny Website. Available online: http://www.mobot.org/MOBOT/research/APweb/ (accessed on 1 March 2023).
- Liu, Y.; Wang, S.; Li, L.; Yang, T.; Dong, S.; Wei, T.; Wu, S.; Liu, Y.; Gong, Y.; Feng, X.; et al. The Cycas genome and the early evolution of seed plants. Nat. Plants 2022, 8, 389–401. [Google Scholar] [CrossRef] [PubMed]
- iNaturalist. Available online: https://www.inaturalist.org/observations (accessed on 4 March 2023).
- Encyclopedia of Life. Available online: https://eol.org/ (accessed on 3 March 2023).
- Olson, D.M.; Dinerstein, E.; Wikramanayake, E.D.; Burgess, N.D.; Powell, G.V.; Underwood, E.C.; D’amico, J.A.; Itoua, I.; Strand, H.E.; Morrison, J.C.; et al. Terrestrial ecoregions of the world: A new map of life on Earth. BioScience 2001, 51, 933–938. [Google Scholar] [CrossRef]
- Environmental Systems Research Institute (ERSI). ArcGIS; Version 10.8; Environmental Systems Research Institute: Redlands, CA, USA, 2020. [Google Scholar]
- Palazzesi, L.; Gottschling, M.; Barreda, V.; Weigend, M. First Miocene fossils of Vivianiaceae shed new light on phylogeny, divergence times, and historical biogeography of Geraniales. Biol. J. Linn. Soc. 2012, 107, 67–85. [Google Scholar] [CrossRef]
- Barthlott, W.; Lauer, W.; Placke, A. Global distribution of species diversity in vascular plants: Towards a world map of phytodiversity. Erdkunde 1996, 50, 317–327. [Google Scholar] [CrossRef]
- Jansson, R. Global patterns in endemism explained by past climatic change. Proc. R. Soc. B 2003, 270, 583–590. [Google Scholar] [PubMed]
- Pie, M.R.; Caron, F.S. Substantial variation in species ages among vertebrate clades. bioRxiv 2023. [Google Scholar] [CrossRef]
- Wickens, G.E. Ecophysiology of Economic Plants in Arid and Semi-Arid Lands; Springer: Berlin/Heidelberg, Germany, 1998; ISBN 364-208-089-8. [Google Scholar]
- Procheş, Ş.; Cowling, R.M.; Goldblatt, P.; Manning, J.C.; Snijman, D.A. An overview of the Cape geophytes. Biol. J. Linn. Soc. 2006, 87, 27–43. [Google Scholar] [CrossRef]
- Mokany, K.; Jordan, G.J.; Harwood, T.D.; Harrison, P.A.; Keppel, G.; Gilfedder, L.; Carter, O.; Ferrier, S. Past, present and future refugia for Tasmania’s palaeoendemic flora. J. Biogeogr. 2017, 44, 1537–1546. [Google Scholar] [CrossRef]
- Linder, H.P. The radiation of the Cape flora, southern Africa. Biol. Rev. 2003, 78, 597–638. [Google Scholar] [CrossRef] [PubMed]
- Procheş, Ş.; Johnson, S.D. Beetle pollination of the fruit-scented cones of the South African cycad Stangeria eriopus. Am. J. Bot. 2009, 96, 1722–1730. [Google Scholar] [CrossRef] [PubMed]
- Olson, S.L. The fossil record of birds. In Avian Biology; Farner, D.S., King, J.R., Parkes, K.C., Eds.; Academic Press: New York, NY, USA, 1985; pp. 79–238. ISBN 012-249-408-3. [Google Scholar]


{kind=link}
| Lineage Number | Order | Family | Genus/Genera | Species | Age (Mya) | Distribution |
|---|---|---|---|---|---|---|
| 1 | Cycadales | Zamiaceae | Stangeria | 1 | 149 | SE Africa |
| 2 | Cycadales | Zamiaceae | Encephalartos | 68 | 101 | much of Africa, mostly SE |
| 3 | Welwitschiales | Welwitschiaceae | Welwitschia | 1 | 168 | SW Africa |
| 4 | Laurales | Lauraceae | Hypodaphnis | 1 | 112 | W Africa |
| 5 | Asparagales | Lanariaceae | Lanaria | 1 | 131 | Cape |
| 6 | Asparagales | Amaryllidaceae | Agapanthus | 8 | 124 | southern Africa |
| 7 | Asparagales | Iridaceae | Nivenia, Klattia, Witsenia | 14 | 103 | Cape |
| 8 | Asparagales | Iridaceae | Aristea | 55 | 113 | much of Africa, Madagascar |
| 9 | Asparagales | Tecophileaceae | Cyanastrum, Cyanella, Kabuyea, Walleria | 16 | 119 | much of Africa |
| 10 | Asparagales | Aparagaceae | Eriospermum | 114 | 120 | much of Africa, mostly Cape |
| 11 | Dioscoreales | Afrothismiaceae | Afrothismia | 16 | 123 | central Africa |
| 12 | Buxales | Buxaceae | Didymeles | 2 | 170 | Madagascar |
| 13 | Gunnerales | Myrothamnaceae | Myrothamnus | 2 | 168 | southern and East Africa, Madagascar |
| 14 | Saxifragales | Peridiscaceae | Medusandra | 2 | 137 | central Africa |
| 15 | Rosales | Dirachmaceae | Dirachma | 2 | 104 | Horn of Africa, Socotra |
| 16 | Oxalidales | Huaceae | Afrostyrax, Hua | 4 | 151 | central and W Africa |
| 17 | Myrtales | Combretaceae | Strephonema | 2 | 109 | central and W Africa |
| 18 | Crossosomatales | Aphloiaceae | Aphloia | 1 | 146 | southern and East Africa, Madagascar |
| 19 | Malvales | Sphaerosepalaceae | Dialyceras, Rhopalocarpus | 18 | 130 | Madagascar |
| 20 | Brassicales | Pentadiplandraceae | Pentadiplandra | 1 | 109 | central Africa |
| 21 | Huerteales | Gerrardinaceae | Gerrardina | 2 | 148 | southern and East Africa |
| 22 | Sapindales | Kirkiaceae | Kirkia | 8 | 106 | southern and East Africa, Madagascar |
| 23 | Santalales | Octoknemaceae | Octoknema | 14 | 117 | central and W Africa |
| 24 | Caryophyllales | Asteropeiaceae, Physenaceae | Asteropeia, Physena | 10 | 121 | Madagascar |
| 25 | Caryophyllales | Kewaceae | Kewa | 6 | 137 | southern Africa, mostly Cape |
| 26 | Caryophyllales | Barbeuiaceae | Barbeuia | 1 | 124 | Madagascar |
| 27 | Caryophyllales | Didiereaceae | Portulacaria, Calyptrotheca, Alluaudia, Alluaudiopsis, Decarya, Didierea | 20 | 113 | southern and East Africa, Madagascar |
| 28 | Cornales | Hydrostachydaceae | Hydrostachys | 21 | 103 | southern and East Africa, Madagascar |
| 29 | Cornales | Curtisiaceae, Grubbiaceae | Curtisia, Grubbia | 4 | 110 | southern and East Africa |
| 30 | Ericales | Sladeniaceae | Ficalhoa | 1 | 143 | southern and East Africa |
| 31 | Ericales | Roridulaceae | Roridula | 2 | 132 | Cape |
| 32 | Asterales | Rousseaceae | Roussea | 1 | 135 | Mauritius |
| 33 | Bruniales | Bruniaceae | Audouinia, Berzelia, Brunia, Linconia, Staavia, Thamnea | 81 | 152 | southern Africa, mostly Cape |
| 34 | Apiales | Apiaceae | Lichtensteinia | 7 | 103 | southern Africa, mostly Cape |
| 35 | Solanales | Convolvulaceae | Humbertia | 1 | 132 | Madagascar |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Procheş, Ş.; Ramdhani, S. Ancient Plant Lineages Endemic to Africa and Its Islands: An Analysis on the Distribution and Diversity. Diversity 2023, 15, 1000. https://doi.org/10.3390/d15091000
Procheş Ş, Ramdhani S. Ancient Plant Lineages Endemic to Africa and Its Islands: An Analysis on the Distribution and Diversity. Diversity. 2023; 15(9):1000. https://doi.org/10.3390/d15091000
Chicago/Turabian StyleProcheş, Şerban, and Syd Ramdhani. 2023. "Ancient Plant Lineages Endemic to Africa and Its Islands: An Analysis on the Distribution and Diversity" Diversity 15, no. 9: 1000. https://doi.org/10.3390/d15091000