
Bioacoustic Monitoring Reveals the Calling Activity of an Endangered Mountaintop Frog (Philoria kundagungan) in Response to Environmental Conditions

Abstract
:1. Introduction
2. Materials and Methods
2.1. Study Species
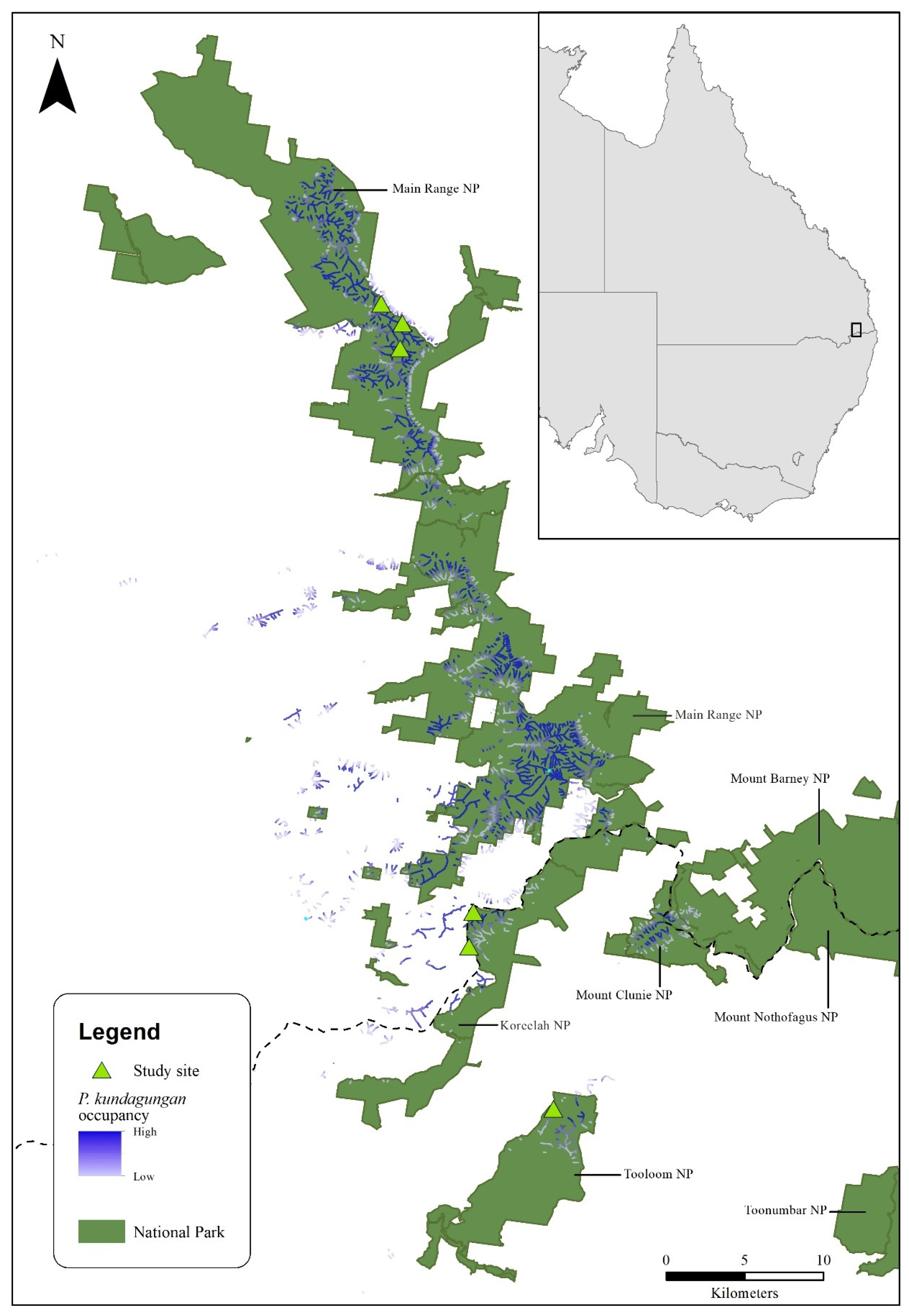
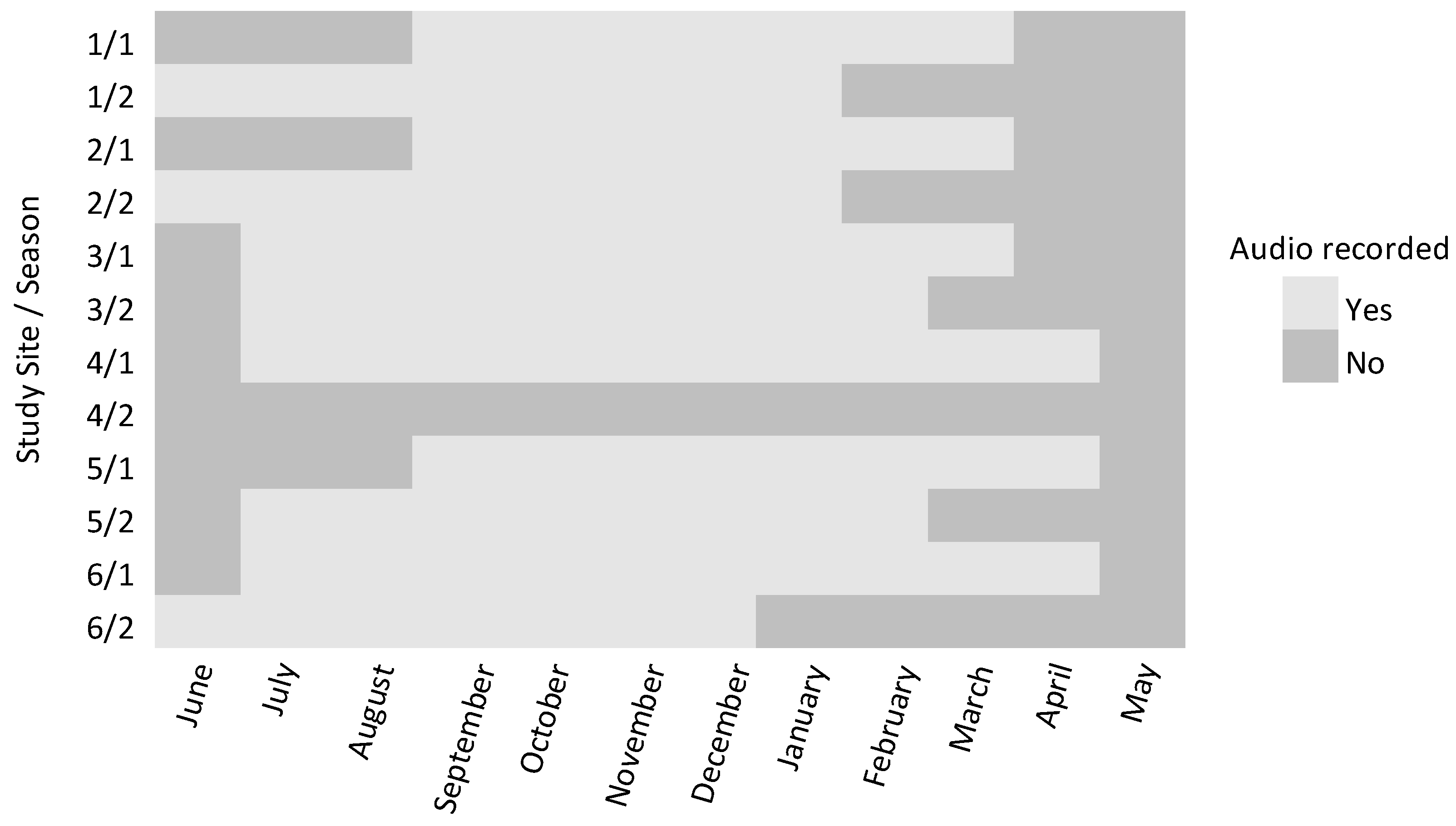
2.2. Data Collection
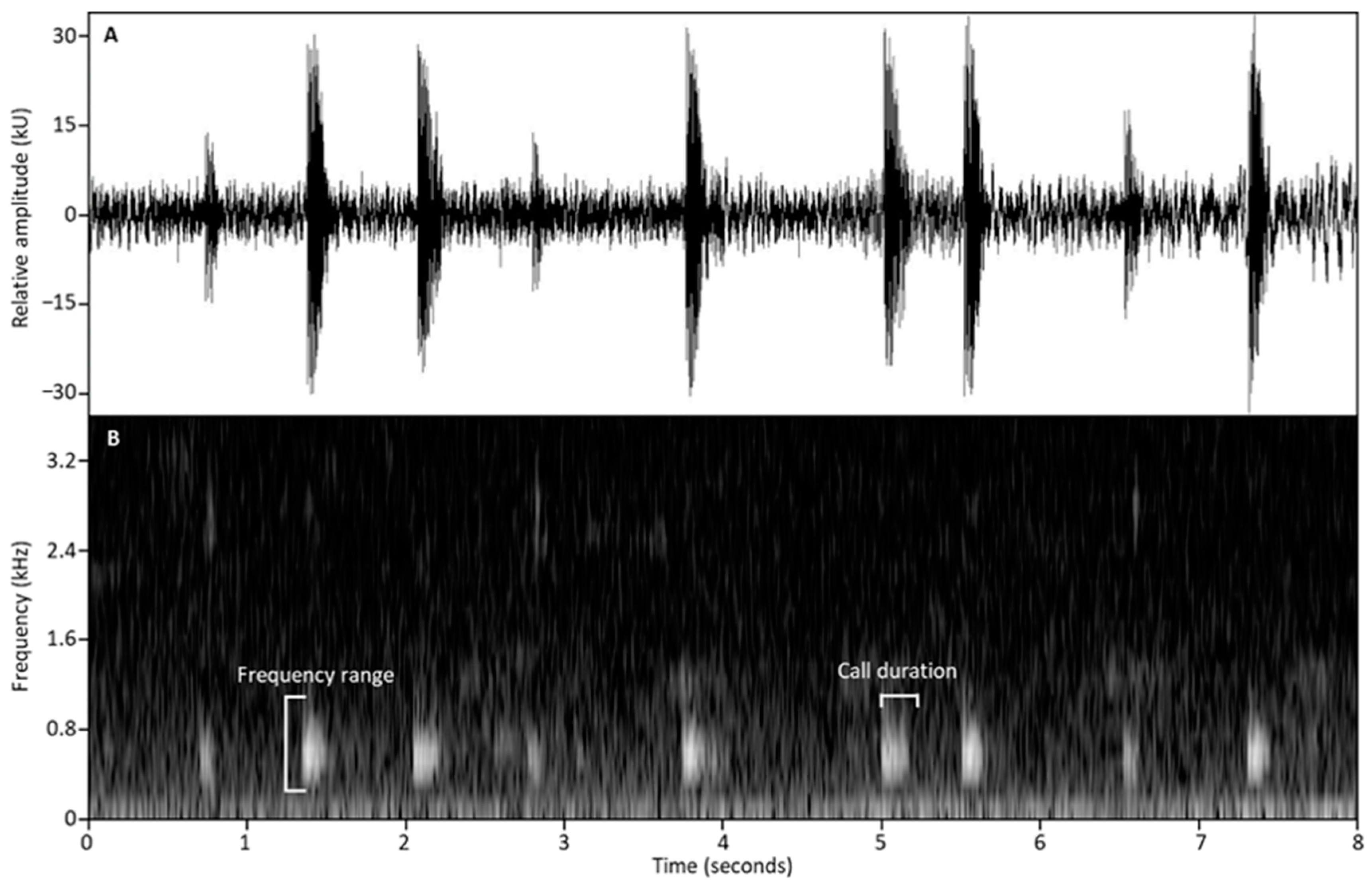
2.3. Development of Automated Species-Specific Recogniser
2.4. Recogniser Performance Assessment
2.5. Statistical Analysis
3. Results
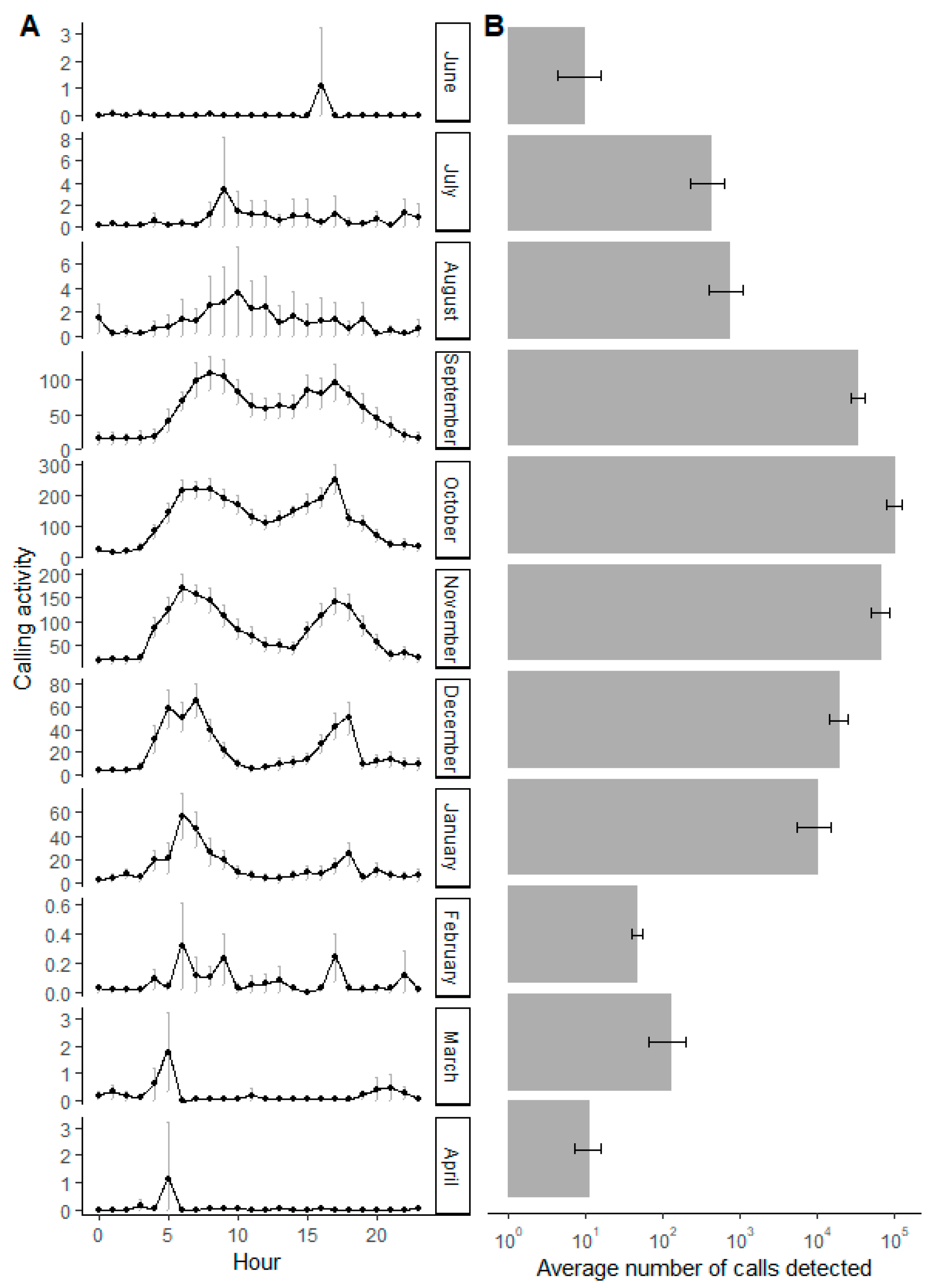
3.1. Annual and Daily Calling Activity
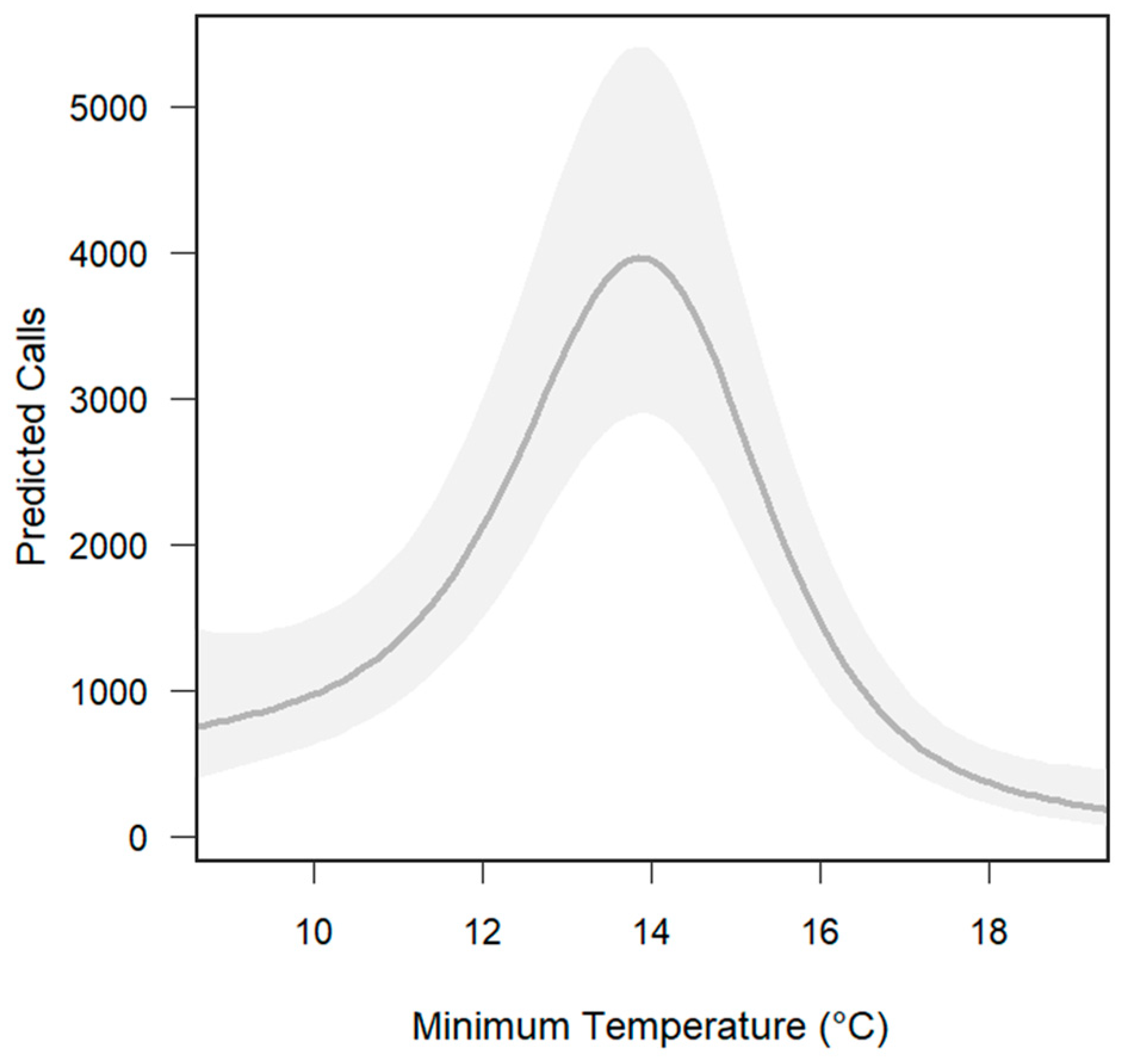
3.2. The Influence of Environmental Variables on Calling Activity
4. Discussion
4.1. Diel and Seasonal Calling Activity
4.2. Environmental Factors
4.3. Implications of Ongoing Climate Change
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- The International Union for Conservation of Nature Red List of Threatened Species. Available online: https://www.iucnredlist.org/ (accessed on 1 March 2023).
- Cushman, S.A. Effects of habitat loss and fragmentation on amphibians: A review and prospectus. Biol. Conserv. 2006, 128, 231–240. [Google Scholar] [CrossRef]
- Fisher, M.C.; Garner, T.W.J. Chytrid fungi and global amphibian declines. Nat. Rev. Microbiol. 2020, 18, 332–343. [Google Scholar] [CrossRef] [PubMed]
- Scheele, B.C.; Pasmans, F.; Skerratt, L.F.; Berger, L.; Martel, A.; Beukema, W.; Acevedo, A.A.; Burrowes, P.A.; Carvalho, T.; Catenazzi, A.; et al. Amphibian fungal panzootic causes catastrophic and ongoing loss of biodiversity. Science 2019, 363, 1459–1463. [Google Scholar] [CrossRef]
- Blaustein, A.R.; Walls, S.C.; Bancroft, B.A.; Lawler, J.J.; Searle, C.L.; Gervasi, S.S. Direct and indirect effects of climate change on amphibian populations. Diversity 2010, 2, 281–313. [Google Scholar] [CrossRef]
- Hof, C.; Araújo, M.B.; Jetz, W.; Rahbek, C. Additive threats from pathogens, climate and land-use change for global amphibian diversity. Nature 2011, 480, 516–519. [Google Scholar] [CrossRef]
- Foden, W.B.; Butchart, S.H.M.; Stuart, S.N.; Vié, J.C.; Akçakaya, H.R.; Angulo, A.; DeVantier, L.M.; Gutsche, A.; Turak, E.; Cao, L.; et al. Identifying the World’s Most Climate Change Vulnerable Species: A Systematic Trait-Based Assessment of all Birds, Amphibians and Corals. PLoS ONE 2013, 8, e65427. [Google Scholar] [CrossRef] [Green Version]
- Li, Y.; Cohen, J.M.; Rohr, J.R. Review and synthesis of the effects of climate change on amphibians. Integr. Zool. 2013, 8, 145–161. [Google Scholar] [CrossRef]
- Hoffmann, E.P.; Williams, K.; Hipsey, M.R.; Mitchell, N.J. Drying microclimates threaten persistence of natural and translocated populations of threatened frogs. Biodivers. Conserv. 2021, 30, 15–34. [Google Scholar] [CrossRef]
- Taylor, E.N.; Diele-Viegas, L.M.; Gangloff, E.J.; Hall, J.M.; Halpern, B.; Massey, M.D.; Rödder, D.; Rollinson, N.; Spears, S.; Sun, B.-J.; et al. The thermal ecology and physiology of reptiles and amphibians: A user’s guide. J. Exp. Zool. Part A Ecol. Integr. Physiol. 2021, 335, 13–44. [Google Scholar] [CrossRef]
- Botts, E.A.; Erasmus, B.F.N.; Alexander, G.J. Observed range dynamics of South African amphibians under conditions of global change. Austral Ecol. 2015, 40, 309–317. [Google Scholar] [CrossRef]
- Abram, N.J.; Henley, B.J.; Sen Gupta, A.; Lippmann, T.J.R.; Clarke, H.; Dowdy, A.J.; Sharples, J.J.; Nolan, R.H.; Zhang, T.; Wooster, M.J.; et al. Connections of climate change and variability to large and extreme forest fires in southeast Australia. Commun. Earth Environ. 2021, 2, 8. [Google Scholar] [CrossRef]
- Pyne, S.J. From Pleistocene to Pyrocene: Fire Replaces Ice. Earth’s Future 2020, 8, e2020EF001722. [Google Scholar] [CrossRef]
- Heard, G.W.; Bolitho, L.J.; Newell, D.; Hines, H.B.; Norman, P.; Willacy, R.J.; Scheele, B.C. Drought, fire, and rainforest endemics: A case study of two threatened frogs impacted by Australia’s “Black Summer”. Ecol. Evol. 2023, 13, e10069. [Google Scholar] [CrossRef]
- Legge, S.; Woinarski, J.C.Z.; Scheele, B.C.; Garnett, S.T.; Lintermans, M.; Nimmo, D.G.; Whiterod, N.S.; Southwell, D.M.; Ehmke, G.; Buchan, A.; et al. Rapid assessment of the biodiversity impacts of the 2019–2020 Australian megafires to guide urgent management intervention and recovery and lessons for other regions. Divers. Distrib. 2022, 28, 571–591. [Google Scholar] [CrossRef]
- Mahony, M.J.; Hines, H.B.; Lemckert, F.; Newell, D.; Roberts, J.D.; Rowley, J.J.; Scheele, B.C.; West, M. The impacts of the 2019–2020 wildfires on Australian frogs. In Australia’s Megafires: Biodiversity Impacts and Lessons From 2019–2020; Rumpff, L., Legge, S., Leeuwen, S.V., Wintle, B., Woinarski, J.C.Z., Eds.; CSIRO Publishing: Melbourne, Australia, 2023; pp. 167–181. [Google Scholar]
- Bolitho, L.; Newell, D. Extensive range contraction predicted under climate warming for two endangered mountaintop frogs from the rainforests of subtropical Australia. Sci. Rep. 2022, 12, 20215. [Google Scholar] [CrossRef] [PubMed]
- Knowles, R.; Mahony, M.; Armstrong, J.; Donnellan, S. Systematics of Sphagnum Frogs of the Genus Philoria (Anura: Myobatrachidae) in Eastern Australia, With the Description of Two New Species. Rec. Aust. Mus. 2004, 56, 57–74. [Google Scholar] [CrossRef] [Green Version]
- Mahony, M.J.; Hines, H.B.; Bertozzi, T.; Mahony, S.V.; Newell, D.A.; Clarke, J.M.; Donnellan, S.C. A new species of Philoria (Anura: Limnodynastidae) from the uplands of the Gondwana Rainforests World Heritage Area of eastern Australia. Zootaxa 2022, 5104, 209–241. [Google Scholar] [CrossRef]
- Newell, D. An update on frog declines from the forests of subtropical eastern Australia. In Status of Conservation and Decline of Amphibians: Australia, New Zealand, and Pacific Islands; Heatwole, H., Rowley, J.L., Eds.; CSIRO Publishing: Melbourne, Australia, 2018; pp. 29–37. [Google Scholar]
- Anstis, M. Tadpoles and frogs of Australia, 2nd ed.; New Holland Publishers Pty Limited: Sydney, Australia, 2017; pp. 500–518. [Google Scholar]
- Hill, A.P.; Prince, P.; Piña Covarrubias, E.; Doncaster, C.P.; Snaddon, J.L.; Rogers, A. AudioMoth: Evaluation of a smart open acoustic device for monitoring biodiversity and the environment. Methods Ecol. Evol. 2018, 9, 1199–1211. [Google Scholar] [CrossRef] [Green Version]
- Shonfield, J.; Heemskerk, S.; Bayne, E.M. Utility of automated species recognition for acoustic monitoring of owls. J. Raptor Res. 2018, 52, 42–55. [Google Scholar] [CrossRef] [Green Version]
- Hoffmann, E.P.; Mitchell, N.J. Breeding phenology of a terrestrial-breeding frog is associated with soil water potential: Implications for conservation in a changing climate. Austral Ecol. 2022, 47, 353–364. [Google Scholar] [CrossRef]
- Wells, K.D. The Ecology and Behavior of Amphibians; University of Chicago Press: Chicago, CA, USA, 2010; pp. 213–225. [Google Scholar]
- Ingram, G.; Corben, C. A new species of Kyarranus (Anura: Leptodactylidae) from Queensland, Australia. Mem. Qld. Mus. 1975, 17, 335–339. [Google Scholar]
- Bolitho, L.J.; Rowley, J.J.L.; Hines, H.B.; Newell, D. Occupancy modelling reveals a highly restricted and fragmented distribution in a threatened montane frog (Philoria kundagungan) in subtropical Australian rainforests. Aust. J. Zool. 2021, 67, 231–240. [Google Scholar] [CrossRef]
- Jeffrey, S.J.; Carter, J.O.; Moodie, K.B.; Beswick, A.R. Using spatial interpolation to construct a comprehensive archive of Australian climate data. Environ. Model. Softw. 2001, 16, 309–330. [Google Scholar] [CrossRef]
- Sokolova, M.; Lapalme, G. A systematic analysis of performance measures for classification tasks. Inf. Process. Manag. 2009, 45, 427–437. [Google Scholar] [CrossRef]
- Wood, S.N. Fast stable restricted maximum likelihood and marginal likelihood estimation of semiparametric sgeneralised linear models. J. R. Stat. Soc. Ser. B (Stat. Methodol.) 2011, 73, 3–36. [Google Scholar] [CrossRef] [Green Version]
- R Development Core Team. R: A Language and Environment for Statistical Computing; Version 3.5.3; The R Foundation for Statistical Computing: Vienna, Austria, 2019; Available online: https://www.R-project.org/ (accessed on 1 March 2023).
- Marchal, J.; Fabianek, F.; Aubry, Y. Software performance for the automated identification of bird vocalisations: The case of two closely related species. Bioacoustics 2022, 31, 397–413. [Google Scholar] [CrossRef]
- Pérez-Granados, C.; Schuchmann, K.-L. Diel and seasonal variation of Striped Cuckoo (Tapera naevia) svocalisations revealed using automated signal recognition. IBIS 2023, 165, 179–189. [Google Scholar] [CrossRef]
- Willacy, R.J.; Mahony, M.; Newell, D.A. If a frog calls in the forest: Bioacoustic monitoring reveals the breeding phenology of the endangered Richmond Range mountain frog (Philoria richmondensis). Austral Ecol. 2015, 40, 625–633. [Google Scholar] [CrossRef]
- Callaghan, C.; Rowley, J. A continental assessment of diurnality in frog calling behaviour. Austral Ecol. 2020, 46, 65–71. [Google Scholar] [CrossRef]
- Chen, Z.; Wiens, J.J. The origins of acoustic communication in vertebrates. Nat. Commun. 2020, 11, 369. [Google Scholar] [CrossRef] [Green Version]
- Seymour, R.S.; Mahony, M.J.; Knowles, R. Respiration of embryos and larvae of the terrestrially breeding frog Kyarranus loveridgei. Herpetologica 1995, 51, 369–376. [Google Scholar]
- Ophir, A.; Schrader, S.; Gillooly, J. Energetic cost of calling: General constraints and species-specific differences. J. Evol. Biol. 2010, 23, 1564–1569. [Google Scholar] [CrossRef] [PubMed]
- Oseen, K.L.; Wassersug, R.J. Environmental factors influencing calling in sympatric anurans. Oecologia 2002, 133, 616–625. [Google Scholar] [CrossRef]
- Hu, J.; Riveros-Iregui, D.A. Life in the clouds: Are tropical montane cloud forests responding to changes in climate? Oecologia 2016, 180, 1061–1073. [Google Scholar] [CrossRef] [PubMed]
- Nishant, N.; Evans, J.P.; Di Virgilio, G.; Downes, S.M.; Ji, F.; Cheung, K.K.; Tam, E.; Miller, J.; Beyer, K.; Riley, M.L. Introducing NARCliM1. 5: Evaluating the performance of regional climate projections for southeast Australia for 1950–2100. Earth’s Future 2021, 9, e2020EF001833. [Google Scholar] [CrossRef]
- Tilman, D.; Clark, M.; Williams, D.R.; Kimmel, K.; Polasky, S.; Packer, C. Future threats to biodiversity and pathways to their prevention. Nature 2017, 546, 73–81. [Google Scholar] [CrossRef]
- Intergovernmental Panel on Climate Change. Climate Change 2021—The Physical Science Basis: Working Group I Contribution to the Sixth Assessment Report of the Intergovernmental Panel on Climate Change; Cambridge University Press: Cambridge, UK, 2023. [Google Scholar]
- Raxworthy, C.J.; Pearson, R.G.; Rabibisoa, N.; Rakotondrazafy, A.M.; Ramanamanjato, J.B.; Raselimanana, A.P.; Wu, S.; Nussbaum, R.A.; Stone, D.A. Extinction vulnerability of tropical montane endemism from warming and upslope displacement: A preliminary appraisal for the highest massif in Madagascar. Glob. Chang. Biol. 2008, 14, 1703–1720. [Google Scholar] [CrossRef] [Green Version]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Model | edf | AIC | ΔAIC |
|---|---|---|---|
| Minimum temperature | 11.65 | 23,653.52 | 0.00 |
| Minimum temperature + Precipitation | 13.81 | 23,702.40 | 48.88 |
| Minimum temperature + Precipitation7 | 13.21 | 23,766.99 | 113.47 |
| Temperature | 11.57 | 23,875.15 | 221.63 |
| Precipitation7 | 8.20 | 23,912.72 | 259.20 |
| Temperature + Precipitation | 13.73 | 23,920.04 | 266.52 |
| Precipitation | 10.54 | 23,941.22 | 287.70 |
| Temperature + Precipitation7 | 12.57 | 24,149.59 | 496.07 |
| Day Length + Precipitation | 13.10 | 24,165.91 | 512.39 |
| Day Length | 10.86 | 24,193.22 | 539.70 |
| Constant | 4.00 | 25,404.02 | 1750.50 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bolitho, L.; Newell, D.; Hines, H. Bioacoustic Monitoring Reveals the Calling Activity of an Endangered Mountaintop Frog (Philoria kundagungan) in Response to Environmental Conditions. Diversity 2023, 15, 931. https://doi.org/10.3390/d15080931
Bolitho L, Newell D, Hines H. Bioacoustic Monitoring Reveals the Calling Activity of an Endangered Mountaintop Frog (Philoria kundagungan) in Response to Environmental Conditions. Diversity. 2023; 15(8):931. https://doi.org/10.3390/d15080931
Chicago/Turabian StyleBolitho, Liam, David Newell, and Harry Hines. 2023. "Bioacoustic Monitoring Reveals the Calling Activity of an Endangered Mountaintop Frog (Philoria kundagungan) in Response to Environmental Conditions" Diversity 15, no. 8: 931. https://doi.org/10.3390/d15080931