
Multiple Chemical Signals in Male Rock Lizards: Femoral Gland Secretions and Feces May Provide Information on Body Size but Using Different Compounds

Abstract
:1. Introduction
2. Materials and Methods
2.1. Study Animals and Sampling Procedures
2.2. Chemical Analyses
2.3. Statistical Analyses of Data
3. Results
3.1. Chemicals in Feces of Male Lizards
3.2. Chemicals in Femoral Gland Secretions of Male Lizards
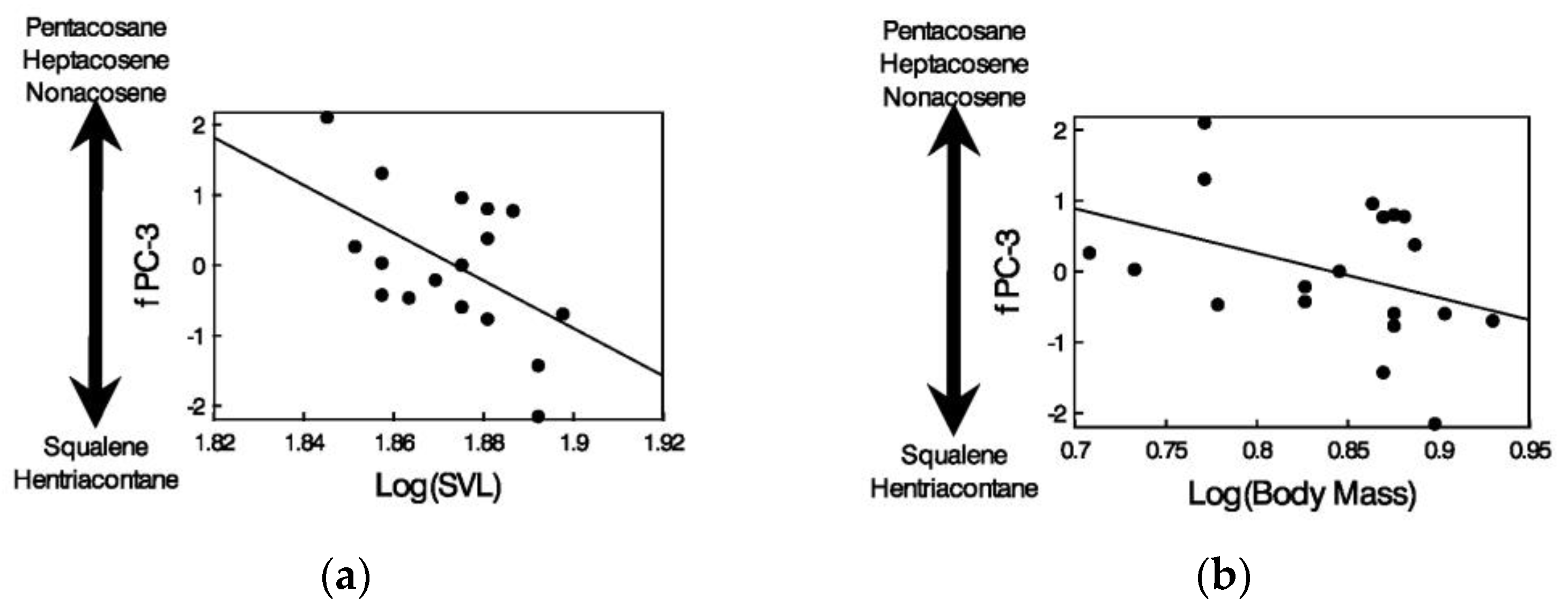
3.3. Relationships between Chemicals in Feces and Body Size of Male Lizards
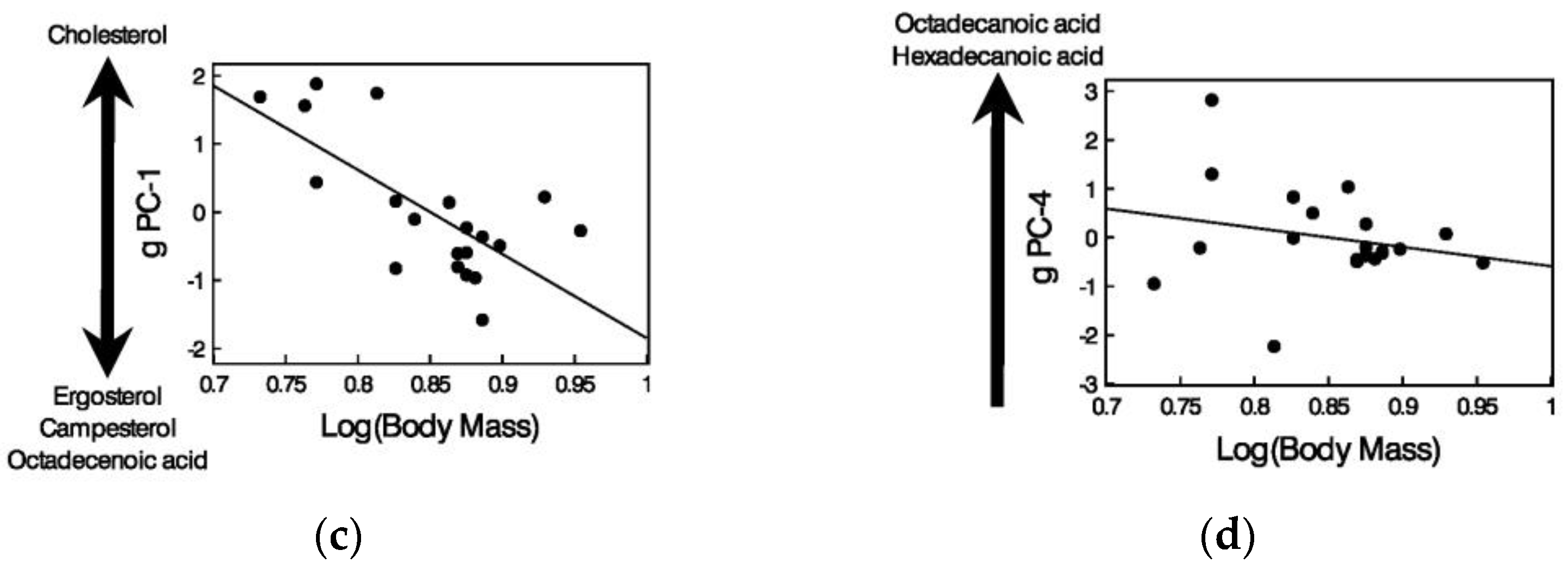
3.4. Relationships between Chemicals in Femoral Gland Secretions and Body Size of Male Lizards
3.5. Intersexual Differences in the Chemical Profiles of Feces
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Mason, R.T. Reptilian pheromones. In Biology of the Reptilia; Gans, C., Crews, D., Eds.; University of Chicago Press: Chicago, IL, USA, 1992; Volume 18, pp. 114–228. [Google Scholar]
- Mason, R.T.; Parker, M.R. Social behavior and pheromonal communication in reptiles. J. Comp. Physiol. A 2010, 196, 729–749. [Google Scholar] [CrossRef] [PubMed]
- Martín, J.; López, P. Pheromones and reproduction in Reptiles. In Hormones and Reproduction of Vertebrates, Volume 3. Reptiles; Norris, D.O., Lopez, K.H., Eds.; Academic Press: Cambridge, MA, USA, 2011; pp. 141–167. [Google Scholar]
- Wyatt, T.D. Pheromones and Animal Behaviour: Chemical Signals and Signatures; Cambridge University Press: Cambridge, UK, 2014. [Google Scholar]
- Baeckens, S. Evolution of animal chemical communication: Insights from non-model species and phylogenetic comparative methods. Belg. J. Zool. 2019, 149, 63–93. [Google Scholar] [CrossRef]
- Labra, A.; Escobar, C.A.; Aguilar, P.M.; Niemeyer, H.M. Sources of pheromones in the lizard Liolaemus tenuis. Rev. Chil. Hist. Nat. 2002, 75, 141–147. [Google Scholar] [CrossRef]
- Weldon, P.J.; Flachsbarth, B.; Schulz, S. Natural products from the integument of nonavian reptiles. Nat. Prod. Rep. 2008, 25, 738–756. [Google Scholar] [CrossRef]
- Mayerl, C.; Baeckens, S.; Van Damme, R. Evolution and role of the follicular epidermal gland system in non-ophidian squamates. Amphib.-Reptil. 2015, 36, 185–206. [Google Scholar] [CrossRef] [Green Version]
- Duvall, D.; Graves, B.D.; Carpenter, G.C. Visual and chemical composite signalling effects of Sceloporus lizards fecal boli. Copeia 1987, 1987, 1028–1031. [Google Scholar] [CrossRef]
- Carpenter, G.C.; Duvall, D. Fecal scent marking in the western banded gecko (Coleonyx variegatus). Herpetologica 1995, 51, 33–38. [Google Scholar]
- López, P.; Aragón, P.; Martín, J. Iberian rock lizards (Lacerta monticola cyreni) assess conspecific information using composite signals from faecal pellets. Ethology 1998, 104, 809–820. [Google Scholar] [CrossRef]
- Bull, C.M.; Griffin, C.L.; Johnston, G.R. Olfactory discrimination in scat-piling lizards. Behav. Ecol. 1999, 10, 136–140. [Google Scholar] [CrossRef] [Green Version]
- Bull, C.M.; Griffin, C.L.; Perkins, M.V. Some properties of a pheromone allowing individual recognition from the scats of an Australian lizard, Egernia striolata. Acta Ethol. 1999, 2, 35–42. [Google Scholar] [CrossRef]
- Bull, C.M.; Griffin, C.L.; Bonnett, M.; Gardner, M.G.; Cooper, S.J. Discrimination between related and unrelated individuals in the Australian lizard Egernia striolata. Behav. Ecol. Sociobiol. 2001, 50, 173–179. [Google Scholar] [CrossRef]
- Aragón, P.; López, P.; Martín, J. Size-dependent chemosensory responses to familiar and unfamiliar conspecific faecal pellets by the Iberian rock-lizard, Lacerta monticola. Ethology 2000, 106, 1115–1128. [Google Scholar] [CrossRef] [Green Version]
- Wilgers, D.J.; Horne, E.A. Discrimination of chemical stimuli in conspecific fecal pellets by visually adept iguanid lizard, Crotaphytus collaris. J. Ethol. 2009, 27, 157–163. [Google Scholar] [CrossRef]
- Valdecantos, S.; Ruiz-Monachesi, M.R.; Labra, A. Testing the functionality of lipids from feces in the conspecific recognition of the weeping lizard, Liolaemus chiliensis. J. Herp. 2020, 54, 476–479. [Google Scholar] [CrossRef] [PubMed]
- Nisa-Ramiro, C.; Rodríguez-Ruiz, G.; López, P.; da Silva Junior, P.I.; Trefaut-Rodrigues, M.; Martín, J. Chemosensory discrimination of male age by female Psammodromus algirus lizards based on femoral secretions and feces. Ethology 2019, 125, 802–809. [Google Scholar] [CrossRef]
- Szabo, B.; Ringler, E. Geckos differentiate self from other using both skin and faecal chemicals: Evidence towards self-recognition? Anim. Cogn. 2023, 26, 1011–1019. [Google Scholar] [CrossRef]
- Martín, J.; López, P. Pheromones and chemical communication in lizards. In Reproductive Biology and Phylogeny of Lizards and Tuatara; Rheubert, J.L., Siegel, D.S., Trauth, S.E., Eds.; CRC Press: Boca Raton, FL, USA, 2014; pp. 43–77. [Google Scholar]
- López, P.; Amo, L.; Martín, J. Reliable signaling by chemical cues of male traits and health state in male lizards, Lacerta monticola. J. Chem. Ecol. 2006, 32, 473–488. [Google Scholar] [CrossRef]
- Martín, J.; López, P. Links between male quality, male chemical signals, and female mate choice in Iberian rock lizards. Funct. Ecol. 2006, 20, 1087–1096. [Google Scholar] [CrossRef]
- Cooper, W.E., Jr.; Pérez-Mellado, V.; Vitt, L.J. Lingual and biting responses to selected lipids by the lizard Podarcis lilfordi. Physiol. Behav. 2002, 75, 237–241. [Google Scholar] [CrossRef]
- Martín, J.; López, P. Chemosensory responses by female Iberian wall lizards, Podarcis hispanica, to selected lipids found in femoral gland secretions of males. J. Herpetol. 2006, 40, 556–561. [Google Scholar] [CrossRef]
- Martín, J.; López, P. Intersexual differences in chemosensory responses to selected lipids reveal different messages conveyed by femoral secretions of male Iberian rock lizards. Amphib.-Reptil. 2008, 29, 572–578. [Google Scholar]
- Martín, J.; López, P. Supplementation of male pheromone on rock substrates attracts female rock lizards to the territories of males: A field experiment. PLoS ONE 2012, 7, e30108. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Møller, A.P.; Pomiankowski, A. Why have birds got multiple sexual ornaments? Behav. Ecol. Sociobiol. 1993, 32, 167–176. [Google Scholar] [CrossRef]
- Johnstone, R.A. Honest advertisement of multiple qualities using multiple signals. J. Theor. Biol. 1995, 177, 87–94. [Google Scholar] [CrossRef]
- Johnstone, R.A. Multiple displays in animal communication: ‘backup signals’ and ‘multiple messages’. Philos. Trans. R. Soc. B Biol. Sci. 1996, 351, 329–338. [Google Scholar]
- Rowe, C. Receiver psychology and the evolution of multicomponent signals. Anim. Behav. 1999, 58, 921–931. [Google Scholar] [CrossRef] [Green Version]
- Partan, S.R.; Marler, P. Issues in the classification of multimodal communication signals. Am. Nat. 2005, 166, 231–245. [Google Scholar] [CrossRef]
- Doucet, S.; Montgomerie, R. Multiple sexual ornaments in satin bowerbirds: Ultraviolet plumage and bowers signal different aspects of male quality. Behav. Ecol. 2003, 14, 503–509. [Google Scholar] [CrossRef] [Green Version]
- Meyers, J.J.; Irschick, D.J.; Vanhooydonck, B.; Herrel, A. Divergent roles for multiple sexual signals in a polygynous lizard. Funct. Ecol. 2006, 20, 709–716. [Google Scholar] [CrossRef]
- Martín, J.; López, P. Multiple color signals may reveal multiple messages in male Schreiber’s Green lizards, Lacerta schreiberi. Behav. Ecol. Sociobiol. 2009, 63, 1743–1755. [Google Scholar] [CrossRef]
- López, P.; Martín, J. Chemosensory exploration of male scent by female rock lizards result from multiple chemical signals of males. Chem. Sens. 2012, 37, 47–54. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Heyman, Y.; Shental, N.; Brandis, A.; Hefetz, A.; Feinerman, O. Ants regulate colony spatial organization using multiple chemical road-signs. Nat. Commun. 2017, 8, 15414. [Google Scholar] [CrossRef] [PubMed]
- Romero-Diaz, C.; Campos, S.M.; Herrmann, M.A.; Soini, H.A.; Novotny, M.V.; Hews, D.K.; Martins, E.P. Composition and compound proportions affect the response to complex chemical signals in a spiny lizard. Behav. Ecol. Sociobiol. 2021, 75, 42. [Google Scholar] [CrossRef]
- Joshi, M.; Ellsworth, B.; Thaker, M. Single components of complex chemical signals convey sex identity and individual variation. Anim. Behav. 2022, 187, 1–13. [Google Scholar] [CrossRef]
- Olsson, M.; Madsen, T.; Nordby, J.; Wapstra, E.; Ujvari, B.; Wittsell, H. Major histocompatibility complex and mate choice in sand lizards. Proc. R. Soc. Lond. B 2003, 270 (Suppl. 2), 254–256. [Google Scholar] [CrossRef] [PubMed]
- Martín, J.; López, P. Vitamin D supplementation increases the attractiveness of males’ scent for female Iberian rock lizards. Proc. R. Soc. Lond. B 2006, 273, 2619–2624. [Google Scholar] [CrossRef] [PubMed]
- Martín, J.; López, P. Condition-dependent chemosignals in reproductive behavior of lizards. Horm. Behav. 2015, 68, 14–24. [Google Scholar] [CrossRef] [Green Version]
- Aragón, P.; López, P.; Martín, J. Discrimination of femoral gland secretions from familiar and unfamiliar conspecifics by male Iberian rock-lizards, Lacerta monticola. J. Herp. 2001, 35, 346–350. [Google Scholar] [CrossRef]
- Aragón, P.; López, P.; Martín, J. Chemosensory discrimination of familiar and unfamiliar conspecifics by lizards: Implications of field spatial relationships between males. Behav. Ecol. Sociobiol. 2001, 50, 128–133. [Google Scholar] [CrossRef]
- Martín, J.; López, P. Scent may signal fighting ability in male Iberian rock lizards. Biol. Lett. 2007, 3, 125–127. [Google Scholar] [CrossRef] [Green Version]
- López, P.; Martín, J. Chemical compounds from femoral gland secretions of male Iberian rock lizards, Lacerta monticola cyreni. Z. Naturforschung C 2005, 60, 632–636. [Google Scholar] [CrossRef] [PubMed]
- Mangiacotti, M.; Martín, J.; López, P.; Reyes-Olivares, C.V.; Rodríguez-Ruiz, G.; Coladonato, A.J.; Scali, S.; Zuffi, M.A.L.; Sacchi, R. Proteins from femoral gland secretions of male rock lizards Iberolacerta cyreni allow self—But not individual—Recognition of unfamiliar males. Behav. Ecol. Sociobiol. 2020, 74, 68. [Google Scholar] [CrossRef]
- Martín, J.; Rodríguez-Ruiz, G.; Cuervo, J.J.; López, P. Intersexual and body size-related variation in chemical constituents from feces and cloacal products involved in intraspecific communication of a fossorial amphisbaenian. PeerJ 2023, 11, e15002. [Google Scholar] [CrossRef] [PubMed]
- García-Roa, R.; Saiz, J.; Gómara, B.; López, P.; Martín, J. How to tackle chemical communication? Relative proportions versus semiquantitative determination of compounds in lizard chemical secretions. Ecol. Evol. 2018, 8, 2032–2040. [Google Scholar] [CrossRef] [Green Version]
- Aebischer, N.J.; Robertson, P.A.; Kenward, R.E. Compositional analysis of habitat use from animal radio-tracking data. Ecology 1993, 74, 1313–1325. [Google Scholar] [CrossRef]
- Clarke, K.R.; Gorley, R.N. PRIMER v6: User Manual/Tutorial; PRIMER-E Ltd.: Plymouth, UK, 2006. [Google Scholar]
- Anderson, M.J.; Gorley, R.N.; Clarke, K.R. PERMANOVA+ for PRIMER: Guide to Software and Statistical Methods; PRIMER-E Ltd.: Plymouth, UK, 2008. [Google Scholar]
- Anderson, M.J. A new method for non-parametric multivariate analysis of variance. Austral Ecol. 2001, 26, 32–46. [Google Scholar]
- Anderson, M.J.; Willis, T.J. Canonical analysis of principal coordinates: A useful method of constrained ordination for ecology. Ecology 2003, 84, 511–525. [Google Scholar] [CrossRef]
- Martín, J.; López, P. Condition-dependent pheromone signalling by male rock lizards: More oily scents are more attractive. Chem. Sens. 2010, 35, 253–262. [Google Scholar] [CrossRef] [Green Version]
- Pérez-Mellado, V.; Bauwens, D.; Gil, M.; Guerrero, F.; Lizana, M.; Ciudad, M.J. Diet composition and prey selection in the lizard Lacerta monticola. Can. J. Zool. 1991, 69, 1728–1735. [Google Scholar] [CrossRef]
- Martín, J.; Raya-García, E.; Ortega, J.; López, P. How to maintain underground social relationships? Chemosensory sex, partner and self recognition in a fossorial amphisbaenian. PLoS ONE 2020, 15, e0237188. [Google Scholar] [CrossRef]
- Mason, R.T.; Jones, T.H.; Fales, H.M.; Pannell, L.K.; Crews, D. Characterization, synthesis, and behavioral responses to sex attractiveness pheromones of red-sided garter snakes (Thamnophis sirtalis parietalis). J. Chem. Ecol. 1990, 16, 2353–2369. [Google Scholar] [CrossRef]
- Eliyahu, D.; Nojima, S.; Mori, K.; Schal, C. New contact sex pheromone components of the German cockroach, Blattella germanica, predicted from the proposed biosynthetic pathway. J. Chem. Ecol. 2008, 34, 229–237. [Google Scholar] [CrossRef] [PubMed]
- Aragón, P.; López, P.; Martín, J. The ontogeny of spatio-temporal tactics and social relationships of adult male Iberian rock lizards, Lacerta monticola. Ethology 2004, 110, 1001–1019. [Google Scholar] [CrossRef]
- López, P.; Martín, J. Potential chemosignals associated with male identity in the amphisbaenian Blanus cinereus. Chem. Sens. 2009, 34, 479–486. [Google Scholar] [CrossRef] [PubMed]
- Shine, R.; Langkilde, T.; Wall, M.; Mason, R.T. Do female garter snakes evade males to avoid harassment or to enhance mate quality? Am. Nat. 2005, 165, 660–668. [Google Scholar] [CrossRef] [PubMed]
- Scheline, R.R. Metabolism of foreign compounds by gastrointestinal microorganisms. Pharmacol. Rev. 1973, 25, 451–523. [Google Scholar] [PubMed]
- Whittaker, D.J.; Slowinski, S.P.; Greenberg, M.; Alian, O.; Winters, A.D.; Ahmad, M.M.; Burrell, M.J.E.; Soini, H.A.; Novotny, M.V.; Ketterson, E.D.; et al. Experimental evidence that symbiotic bacteria produce chemical cues in a songbird. J. Exp. Biol. 2019, 222, jeb202978. [Google Scholar] [CrossRef] [Green Version]
- Baeckens, S.; De Meester, W.; Zoran Tadić, Z.; Van Damme, R. Where to do number two: Lizards prefer to defecate on the largest rock in the territory. Behav. Process. 2019, 167, 103937. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| Chemical Class | Males | Females |
|---|---|---|
| Steroids | 38.61 ± 20.84% | 26.93 ± 12.04% |
| Linear alkanes | 26.92 ± 10.51% | 26.50 ± 9.96% |
| Branched alkanes | 25.15 ± 15.95% | 36.64 ± 14.14% |
| Alcohols | 2.95 ± 4.70% | 3.38 ± 3.28% |
| Carboxyilic acids | 2.60 ± 6.28% | 3.20 ± 5.14% |
| Alkenes | 1.64 ± 2.10% | 1.31 ± 2.85% |
| Terpenoids | 0.76 ± 0.72% | 0.69 ± 0.45% |
| Esters of carboxylic acids | 0.56 ± 1.16% | 0.59 ± 0.67% |
| Waxy esters | 0.30 ± 0.66% | 0.21 ± 0.22% |
| Aldehydes | 0.30 ± 0.17% | 0.35 ± 0.10% |
| Tocopherols | 0.16 ± 0.26% | 0.11 ± 0.20% |
| Ketones | 0.03 ± 0.08% | 0.08 ± 0.17% |
| Aromatic heterocyclic | 0.02 ± 0.04% | 0.01 ± 0.01% |
| Cyclic octa-atomic sulfur | 0.01 ± 0.02% | 0 |
| Furanones | 0 | 0.01 ± 0.04% |
| Compound | Mean ± SD | fPC-1 | fPC-2 | fPC-3 | fPC-4 |
|---|---|---|---|---|---|
| Cholesterol | 13.85 ± 7.06 | 0.71 | −0.30 | −0.23 | 0.35 |
| Sitosterol | 11.34 ± 9.28 | 0.92 | −0.06 | −0.17 | −0.09 |
| Heptacosane | 9.78 ± 5.76 | −0.68 | −0.03 | 0.40 | −0.33 |
| Nonacosane | 5.63 ± 3.68 | 0.00 | 0.74 | −0.04 | −0.20 |
| Pentacosane | 4.15 ± 3.18 | −0.27 | 0.03 | 0.81 | −0.07 |
| 13-Methylheptacosane | 3.47 ± 5.89 | −0.61 | 0.31 | −0.17 | −0.42 |
| Hentriacontane | 2.80 ± 3.30 | 0.25 | 0.55 | −0.51 | 0.38 |
| 13-Methylnonacosane | 2.76 ± 3.12 | −0.68 | −0.15 | −0.08 | 0.42 |
| Campesterol | 2.74 ± 2.03 | 0.85 | 0.01 | −0.23 | 0.02 |
| Unid. Branched Alkane at RT = 55.55 | 2.55 ± 2.37 | −0.68 | 0.50 | −0.02 | −0.19 |
| Docosanol | 2.37 ± 3.79 | 0.01 | 0.17 | 0.13 | 0.51 |
| Stigmasta-5,24(28)-dien-3-ol | 1.93 ± 2.17 | 0.58 | 0.01 | 0.40 | −0.11 |
| Unid. Branched Alkane at RT = 58.18 | 1.65 ± 2.01 | −0.67 | 0.01 | −0.10 | 0.39 |
| 9-Octadecenoic acid | 1.53 ± 4.08 | −0.23 | 0.45 | 0.20 | 0.32 |
| Unid. Branched Alkane at RT = 56.80 | 1.37 ± 1.61 | −0.41 | 0.17 | −0.02 | −0.62 |
| Octacosane | 1.29 ± 1.12 | −0.80 | 0.08 | −0.10 | 0.19 |
| Unid. Branched Alkane at RT = 53.97 | 1.24 ± 2.31 | −0.59 | −0.44 | 0.36 | −0.16 |
| Unid. Branched Alkane at RT = 55.41 | 1.14 ± 1.37 | −0.83 | 0.01 | 0.04 | 0.17 |
| 15,19-Dimethyl-pentatriacontane? | 1.09 ± 1.03 | 0.00 | 0.32 | −0.01 | 0.16 |
| Tricosane | 1.07 ± 0.95 | 0.08 | 0.48 | 0.29 | 0.09 |
| Unid. Steroid at RT = 68.05 | 1.03 ± 1.59 | 0.61 | 0.30 | 0.30 | −0.19 |
| Unid. Branched Alkane at RT = 55.03 | 0.97 ± 1.85 | −0.21 | 0.58 | 0.02 | 0.21 |
| Unid. Branched Alkane at RT = 58.42 | 0.89 ± 0.74 | 0.08 | 0.73 | 0.30 | −0.36 |
| Stigmasterol | 0.89 ± 0.85 | 0.80 | 0.12 | 0.00 | 0.24 |
| Unid. Branched Alkane at RT = 55.11 | 0.80 ± 1.10 | −0.59 | 0.46 | −0.08 | −0.28 |
| Ergosta-5,24-dien-3-ol | 0.76 ± 1.17 | 0.75 | −0.06 | 0.41 | 0.11 |
| Squalene | 0.76 ± 0.72 | −0.62 | −0.18 | −0.57 | 0.19 |
| Heptacosene | 0.75 ± 1.68 | −0.31 | −0.34 | 0.64 | 0.07 |
| Hexacosane | 0.75 ± 0.74 | −0.70 | −0.27 | 0.33 | −0.01 |
| Hexadecanoic acid | 0.71 ± 2.21 | −0.25 | 0.41 | 0.34 | 0.36 |
| Unid. Branched Alkane at RT = 52.04 | 0.65 ± 1.08 | −0.43 | 0.31 | 0.05 | 0.63 |
| Unid. Branched Alkane at RT = 52.63 | 0.62 ± 0.65 | −0.63 | 0.18 | 0.33 | 0.29 |
| Unid. Steroid at RT = 73.77 | 0.62 ± 1.78 | 0.31 | 0.26 | 0.40 | 0.13 |
| Stigmast-7-en-3-ol | 0.61 ± 0.68 | 0.46 | 0.10 | 0.56 | −0.23 |
| Tritriacontane | 0.57 ± 0.81 | 0.32 | 0.81 | 0.06 | 0.09 |
| Stigmastanol | 0.57 ± 0.91 | 0.42 | −0.06 | 0.17 | −0.16 |
| Cholesta-5,7-dien-3-ol | 0.56 ± 0.52 | 0.68 | −0.08 | 0.32 | 0.28 |
| Nonacosene | 0.54 ± 0.66 | 0.22 | −0.13 | 0.72 | 0.39 |
| Unid. Branched Alkane at RT = 67.18 | 0.50 ± 0.65 | 0.52 | 0.47 | −0.35 | −0.09 |
| Eigenvalue | 11.63 | 4.80 | 4.37 | 3.18 | |
| Explained Variance (%) | 29.83 | 12.30 | 11.19 | 8.15 |
| Compound | Mean ± SD | gPC-1 | gPC-2 | gPC-3 | gPC-4 |
|---|---|---|---|---|---|
| Cholesterol | 77.92 ± 2.16 | 0.88 | −0.18 | −0.08 | −0.09 |
| Campesterol | 5.43 ± 1.89 | −0.86 | −0.29 | 0.02 | 0.11 |
| Hexadecanoic acid | 4.04 ± 0.03 | −0.25 | −0.30 | 0.40 | 0.60 |
| 9-Octadecenoic acid | 3.03 ± 0.02 | −0.71 | 0.44 | 0.27 | −0.25 |
| Octadecanoic acid | 2.04 ± 0.03 | 0.10 | 0.11 | 0.55 | 0.75 |
| Cholest-5-en-3-ol-, acetate | 1.58 ± 0.46 | 0.40 | 0.85 | −0.10 | −0.03 |
| Sitosterol | 1.13 ± 0.58 | −0.69 | 0.21 | −0.18 | 0.45 |
| Cholest-4-en-3-one | 0.97 ± 0.70 | 0.67 | 0.16 | 0.43 | −0.15 |
| Cholest-5-en-3-ol, derivative? | 0.95 ± 0.43 | 0.24 | 0.80 | 0.12 | −0.02 |
| 4,4-Dimethyl-cholesta-5,7-dien-3-ol | 0.75 ± 0.27 | −0.79 | −0.21 | 0.13 | −0.20 |
| Ergosta-5,8-dien-3-ol | 0.68 ± 0.13 | −0.75 | 0.05 | −0.32 | −0.12 |
| Cholesta-3,5-diene | 0.51 ± 0.16 | 0.15 | 0.94 | −0.05 | 0.08 |
| Cholesta-4,6-dien-3-ol | 0.31 ± 0.11 | −0.19 | 0.52 | 0.00 | −0.54 |
| Ergosterol | 0.15 ± 0.11 | −0.91 | 0.14 | 0.07 | −0.10 |
| Unid. Steroid (253,269,367,382) | 0.11 ± 0.07 | −0.71 | 0.17 | 0.09 | −0.09 |
| Squalene | 0.11 ± 0.18 | −0.43 | 0.62 | 0.29 | 0.23 |
| Cholesta-2,4-diene | 0.08 ± 0.06 | −0.56 | 0.66 | −0.03 | 0.27 |
| Ergosta-5,22-dien-3-ol | 0.08 ± 0.09 | −0.70 | −0.21 | −0.03 | −0.11 |
| Unid. Steroid (197,251,311,376) | 0.07 ± 0.06 | −0.20 | 0.20 | −0.76 | 0.21 |
| Eicosanol | 0.02 ± 0.03 | −0.38 | 0.10 | 0.51 | −0.51 |
| 9-Hexadecenoic acid | 0.02 ± 0.02 | −0.07 | −0.33 | 0.73 | −0.12 |
| Unid. Steroid (251,363,378) | 0.01 ± 0.02 | −0.44 | −0.60 | −0.21 | −0.02 |
| Eigenvalue | 7.17 | 4.46 | 2.42 | 2.05 | |
| Explained Variance (%) | 32.60 | 20.28 | 10.99 | 9.30 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Martín, J.; Rodríguez-Ruiz, G.; López, P. Multiple Chemical Signals in Male Rock Lizards: Femoral Gland Secretions and Feces May Provide Information on Body Size but Using Different Compounds. Diversity 2023, 15, 858. https://doi.org/10.3390/d15070858
Martín J, Rodríguez-Ruiz G, López P. Multiple Chemical Signals in Male Rock Lizards: Femoral Gland Secretions and Feces May Provide Information on Body Size but Using Different Compounds. Diversity. 2023; 15(7):858. https://doi.org/10.3390/d15070858
Chicago/Turabian StyleMartín, José, Gonzalo Rodríguez-Ruiz, and Pilar López. 2023. "Multiple Chemical Signals in Male Rock Lizards: Femoral Gland Secretions and Feces May Provide Information on Body Size but Using Different Compounds" Diversity 15, no. 7: 858. https://doi.org/10.3390/d15070858