1. Introduction
Desert poplar
Populus pruinosa Schrenk (
Figure S1) is listed in the Red Data Book of Kazakhstan [
1]. In 2007, it was added to the International Red List as a near-threatened species [
2].
P. pruinosa is distributed in Central Asian countries, Iran, Afghanistan, and Xinjiang, but does not form plant communities everywhere.
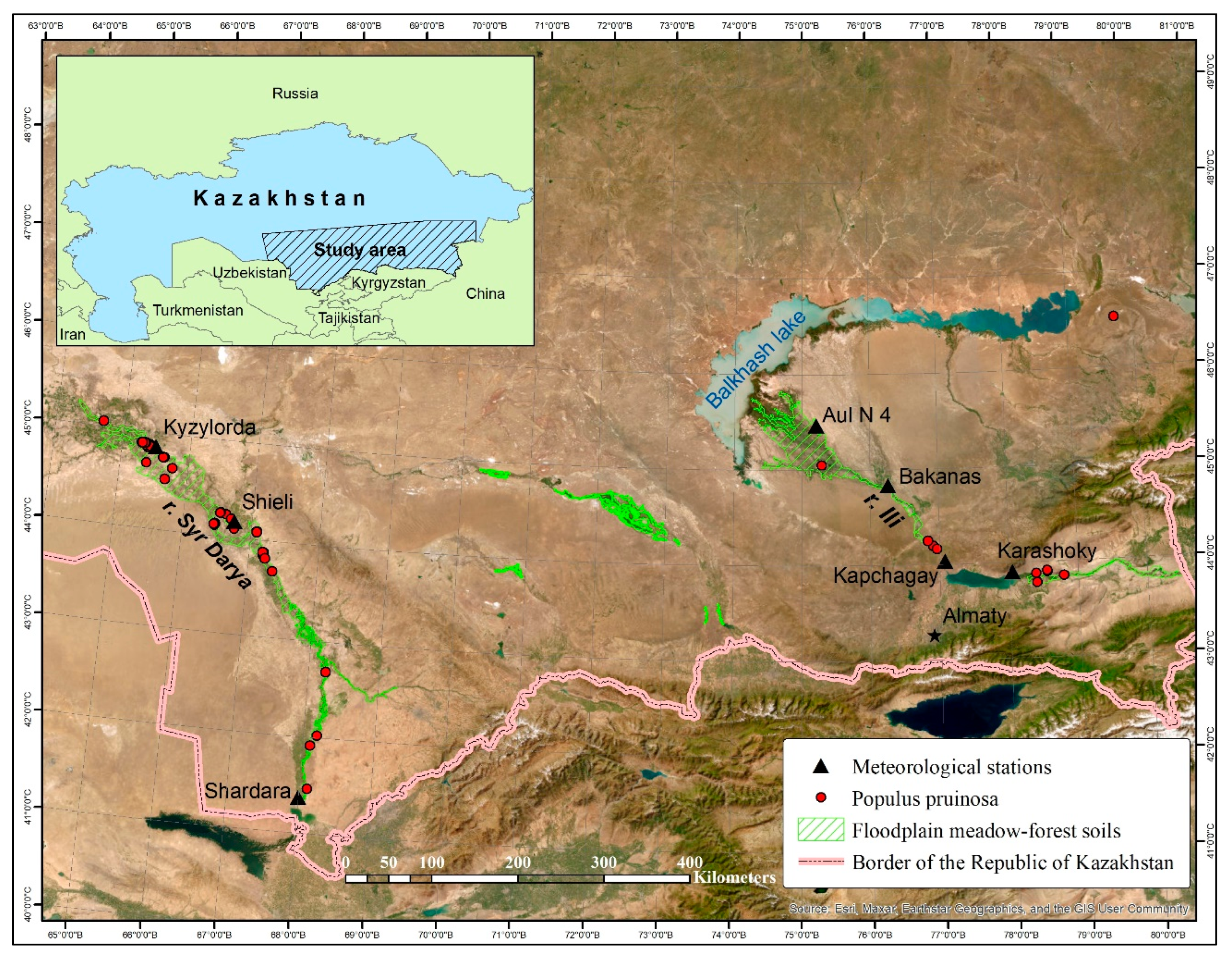
In Kazakhstan, the habitats of the species are associated with the floodplains of desert rivers, including the Syr Darya, the Ili, and their ancient riverbeds (
Figure 1). Floodplain forests (tugai, also tugay) are gallery riparian forests of desert rivers. They found mainly in the Syr Darya River Valley. The distribution in the Ili River Valley is limited where there are only small groves and sparse woodlands. The locations characterized by the participation of two desert poplars from the
Turanga subgenus, including
Populus euphratica Olivier, whose range is wider, are sometimes noted. These two desert poplars have morphological differences in their leaves.
P. euphratica have two types of leaves: juvenile leaves that are narrowly oblong and leaves on mature shoots that are usually broader than long, rhombic, or ovate. In contrast,
P. pruinosa exhibits oval, round, and broad ovate-shaped leaves. [
3,
4]
Studies of both species of turanga have consistently drawn the attention of scientists from Central Asian countries. In our article, we focus on a species that is rarer for Kazakhstan with a narrower range of P. pruinosa.
In Turkmenistan, [
5]
P. pruinosa is found only as an admixture in tugai forests of
P. euphratica. It has been noted that its reproduction is mainly vegetative, not seeded. In Uzbekistan, thickets of
P. pruinosa were found in the lower reaches and the delta of the Amu Darya River, as well as the valleys of the Syr Darya and Zeravshan [
6] rivers. In the middle reaches of the Syr Darya within Uzbekistan and Kazakhstan, turanga forests with a tall-grass meadow cover of
Erianthus ravennae (L.) P. Beauv and
Arundo donax L. are common [
7]. In the Amu Darya Delta, tugai vegetation with the participation of
P. pruinosa has all but disappeared due to the Aral Sea crisis [
8].
According to the iNaturalist website [
9], the locations of the species in Central Asia are marked at 15 points in Uzbekistan (in the valleys of the Zeravshan, the Amu Darya, the Syr Darya, the Surkhan Darya Rivers, the Amu-Bukhara Canal), Tajikistan (the valley of the Vakhsh River), and Kazakhstan (the valleys of the Syr Darya and the Ili Rivers). In the Global Biodiversity Information Facility (GBIF) Secretariat checklist dataset, there are 256 results for findings in China, Afghanistan, Uzbekistan, Kazakhstan, Turkmenistan, Tajikistan, Kyrgyzstan, and Russia [
10]. It should be noted that there are 21 specimens of this species in the herbarium of the Institute of Botany and Phytointroduction (AA). The earliest specimen was collected by A. G. Schrenk in 1843.
Turanga forests used to occupy much larger territories in Kazakhstan than they do at present [
11]. The studies of the initial period covered the forestry characteristics with assessments of wood stocks. Further research considered the conditions of growth, the features of reproduction, and the introduction potential of the species for use in protective forest plantations and the landscaping of settlements in arid regions [
12]. It was noted that the reduction in tugai forests was associated with changes in the hydrological regimes of rivers, the reduction in groundwater, fires, and grazing. After this species was listed in the Red Data Book of Kazakhstan [
1,
13], efforts were aimed primarily at preserving the species in natural reserves and obtaining planting material using in vitro propagation and conservation.
Recent publications reflected the results of a study of the floristic composition of turanga woodlands and forests in key areas [
14,
15] and an evaluation of the soil conditions [
16,
17]. An assessment of the current state of turanga stands in the southeast of Kazakhstan (the Almaty region) was also conducted [
18], which showed the dominance of low-priority, low-field stands that are mainly mature and over-mature in terms of age. The natural renewal in the form of root-bearing individuals was noted only in some small areas, which characterized the plantings as unstable.
Chinese scientists have devoted many publications to these two desert poplars, which are in danger of extinction in the northwest of China. The ecological and phylogeographic features and the genetic differentiation of the turanga species have been studied. There is a fragmentation and a decrease in poplar populations due to the orogeny of the Tien Shan Mountains and Quaternary climatic aridification, which further accelerated the population divergence [
19,
20]. The genetic differentiation was driven by geographical and environmental factors. This knowledge should be helpful for designing strategies to protect these genetic resources in the future.
Among the current problems, water scarcity and low groundwater levels are highlighted. Together, these issues have led to a low rate of seed/seedling regeneration in many populations of
P. euphratica and
P. pruinosa. At the same time, vegetative (clonal) reproduction also depends on floods and the timing and volume of incoming water [
21].
Ecosystems of the turanga forests are threatened by overuse and a continuous decrease in the groundwater level due to the excessive use of water for agriculture, which has resulted in a drastic reduction in the poplar forest area during recent decades [
22]. It should be noted that tugai forests provide essential provisioning, regulating, and cultural ecosystem services. They have significant environment-forming, soil-fixing, and reclamation value, i.e., they generally stabilize the ecological situation in the floodplains of rivers in arid regions [
18,
22].
To better protect and restore the genetic resources of desert poplars, more attention must be paid to the intensity of flood diversion and irrigation, encouraging seed germination and seedling establishment, and protecting populations distributed in the northern Xinjiang region, which hold the highest genetic diversity [
20].
The aim of this research is to advance our knowledge of the woodlands and forests of Populus pruinosa in Kazakhstan. The tasks undertaken to achieve this goal are as follows: (1) to summarize the data on cenoflora; (2) to present the forestry characteristics; (3) to reveal the climatic and soil conditions; (4) to conduct ecological niche modeling of P. pruinosa and select suitable habitats for ecological restoration; and (5) to identify strategies for rehabilitation.
3. Results
3.1. Floristic Composition
A survey of the turanga woodlands in the Ili River Valley on the basis of 10 geobotanical descriptions revealed 103 species of vascular plants. To compile a complete list of the plant species, the floristic list of Stikhareva et al. [
14] at three key sites was analyzed, from which another 61 species were added. Thus, the cenoflora of the turanga woodlands of the Ili River Valley consisted of 164 species of vascular plants belonging to 112 genera and 40 families. The species composition included three species from the Red Data Book of Kazakhstan (
Ferula iliensis,
Tulipa biflora, and
T. kolpakowskiana). The participation of weed species in plant communities was high (24%), indicating the anthropogenic transformation of habitats, which included changes in the hydrological regime, grazing, fires, and locally noted recreation.
Our research in the Syr Darya River Valley supplemented the floristic list of Vesselova et al. [
15] with 16 species. The list currently includes 119 species of vascular plants from 93 genera and 32 families. There were two species from the Red Data Book of Kazakhstan (
Tulipa biflora and
T. borszczowii) in the floodplain forests. Weed species comprised 21% of the cenoflora.
Our analysis of the available materials and publications resulted in a general list of the species found in the turanga woodlands and forests in Kazakhstan, which included 223 species of vascular plants from 136 genera and 42 families (
Table S2). The leading five families were Asteraceae (37 species), Amaranthaceae (27), Poaceae (20), Brassicaceae (18), and Fabaceae (12), which comprised 52% of the cenoflora. The leading genera were
Artemisia, with nine species, and
Salsola, with eight species.
A comparison of the lists from the valleys of the two rivers showed that the Sörensen coefficient was 0.42. This indicated differences, but, at the same time, 42% of the species were common. The differences were due to climatic and botanical geographic features. Both rivers were located in the Sahara–Gobi Desert region of the Iran–Turan subregion, but the turanga woodlands and forests were confined to different botanical and geographic provinces: the Ili River to the north of Turan and the Syr Darya River to the south of Turan [
45].
3.2. Forestry Characteristics
During the field research, the forestry characteristics of 30 turanga communities were assessed. We offer examples from four sites in the Ili River Valley and four sites in the Syr Darya River Valley (
Table S3).
The edificator of the forest communities was P. pruinosa. The basis of the tree layer was created by turanga, and sometimes Elaeagnus angustifolia was mixed with it. However, this occurred only rarely. The turanga communities in the valleys of the Ili and the Syr Darya rivers differed in terms of their closeness to the tree layer, bonitet, and vital conditions.
In the Ili River Valley, the closeness of the woodlands was lower (0.3–0.6) than in the Syr Darya River Valley forests (0.7–1.0), and the bonitet varied from high to low class. The height of the trees varied from 9 to 14 m. The age of the trees varied from 40 to 60 years. Undergrowth was found almost in all the plantings and ranged from five to 10 pieces in the key sites. In the Syr Darya, the plantings were low priority, the height of the trees varied from 4 to 20 m, and the stands were of different ages—from 10 to 55 years. The undergrowth was marked only in one site, with an amount of 21.
The turanga communities had a well-developed shrub layer, in which Caragana halodendron was mandatory and dominant. Tamarisk species were found with a high constancy. For the Ili River, a species from the Red Data Book Berberis iliensis was marked. The salinization of the soil was indicated by perennial saltworts: Suaeda microphylla and Kalidium foliatum in the Ili River Valley, and Halostachys belangeriana in the Syr Darya River Valley. The grass layer was well developed in the Ili River Valley, and the projective coverage varied from 60 to 100%. Grasses were predominant and included Calamagrostis epigeios, Leymus angustus, L. multicaulis, Neotrinia splendens, and Phragmites australis. Many weed species (Dodartia orientalis, Hyoscyamus niger, Sophora alopecuroides, Chenopodium album, Lactuca serriola, Atriplex tatarica, Rhaponticum repens, etc.) were noted in the herbal layer, which indicated an anthropogenic influence from grazing and periodic fires.
In the Syr Darya River Valley, the herbal layer accounted for 30–80%, and it was formed by Leymus multicaulis. Annual saltworts (Atriplex sagittata, Petrosimonia sibirica, Suaeda altissima, and Climacoptera lanata) and halophytic herbs (Saussurea salsa, Aeluropus littoralis, and Limonium otolepis) exhibited a high participation in the coverage.
The species richness of the turanga communities in the valleys of the two rivers had some differences, which were determined by the range of variation. In the Ili River Valley, it varied from 4.7 to 9.7, and in the Syr Darya River Valley it varied from 4.1 to 7.5. This showed that the species diversity was slightly higher in the Ili River Valley.
The assessment of the vitality of the stands showed that the turanga communities in the Ili River Valley were in better condition. The species developed normally, reached its normal size, went through the entire cycle of its development, and formed fruits. In almost all the key plots, undergrowth from the seed origin developed.
In the Syr Darya River Valley, a weakening was evident, with insufficient vegetative development where plants did not pass through their entire life cycle. The trees did not reach their normal size and undergrowth was noted only in one site. There was a high closeness of the stands which contributed to competition for water resources and the disruption of self-renewal.
3.3. Ecological Niche Modeling
Ecological niche modeling of
P. pruinosa was undertaken to identify the relationships between the distribution of the species and the habitat [
40,
46] in order to create maps of the potential area and the most suitable places for the restoration of floodplain forests in Kazakhstan. The basis for the mapping were the identified optimal ranges of the key environmental variables [
41].
3.3.1. Climate
The climatic conditions were determined based on the average data of Kazhydromet for the last 30 years (1 January 1991 to 31 December 2020) for seven meteorological stations located within the range of
P. pruinosa [
47]. A thirty-year period was the standard for determining the climatic norms. The Shardara, Shieli, and Kyzylorda weather stations were located near the Syr Darya River. Karashoky, Kapchagay, Bakanas, and Aul N 4 were located along the Ili River.
According to the available data, the average monthly temperatures during the year ranged from −11.8 to +29.2 °C (
Figure 2). In winter, they ranged from −11.8 to +1 °C, in spring from +1.2 to +21.9 °C, in summer from +23.3 to +29.2 °C, and in autumn from −0.9 up to +21.4 °C. The average monthly maximum air temperature varied in winter from −6.2 to +6.8 °C, in spring from +7.9 to +28.7 °C, in summer from +30.6 to +36.6 °C, and in autumn from +5.5 to +29.3 °C.
The average monthly minimum air temperature also varied in winter from −1.7 to −16.7 °C, in spring from −4.4 to +16.2 °C, in summer from +14.8 to +22.8 °C, and in autumn from −5.8 to +14.7 °C. The average annual air temperature ranged from +8.3 to +14.6 °C. All the maximum warm values were marked at stations located along the Syr Darya River, and the cold values were marked along the Ili River. The average monthly precipitation ranged from 1 mm in August at Shardara to 39 mm in May at Kapchagay (
Figure 3).
The seasonal fluctuations at the stations were as follows. In winter, it fluctuated from 8–10 mm (Aul N 4) to 28–37 mm (Shardara); in spring from 10–12 mm (Aul N 4) to 21–36 mm (Shardara) and 24–39 mm (Kapchagay); in summer from 1–6 mm (Shardara) to 15–32 mm (Kapchagay); and in autumn from 2–28 mm (Shardara) to 13–29 mm (Kapchagay). The average annual number of days with precipitation ≥ 1 mm ranged from 32 at Shieli and Aul N 4 up to 60 days for Kapchagay. The average monthly number of days with precipitation ≥ 1 mm ranged from 0.5 in July and September at Shardara and Shieli, to 6.7 in April at Kapchagay.
The average annual precipitation ranged from 117 to 306 mm, with the highest precipitation rates occurring in November and spring, and the minimum occurring from June to October at all the stations. The largest number of places with a distribution of
P. pruinosa was noted at the weather stations located in Kyzylorda and Shieli, with average annual temperatures of +11.3–11.9 °C, average monthly maximum air temperatures for the year of +17.8–19.1 °C, and minimum temperatures of +5.3–5.4 °C, where the average annual precipitation ranged from 145 to 147 mm (
Figure 4).
To create a model of the ecological niche of
P. pruinosa, monthly climate data from WorldClim were used as the environmental variables [
42,
48]. Comparing the WorldClim data for the period 1970–2000 to the current norms described above, we concluded that there was a slight increase in the temperature regime (
Table S1, Figure S3). At the same time, the WorldClim averaged values for the period 2010–2018 exceeded the norm in almost every month. Due to the fact that the period under consideration was nine years and the standard for determining climate norms was a thirty-year period, these data cannot be considered as complete evidence, but only as a trend averaged over the period 2010–2018.
The WorldClim precipitation data for the periods 1970–2000 and 2010–2018 differed significantly from each other and from the norm, although the trends during the year coincided (
Table S1, Figure S4). The difference between the raster datasets and terrestrial datasets was due to the fact that the WorldClim databases were calculated for a different period and relied on reconstructions based on the extrapolation of data from a limited number of weather stations.
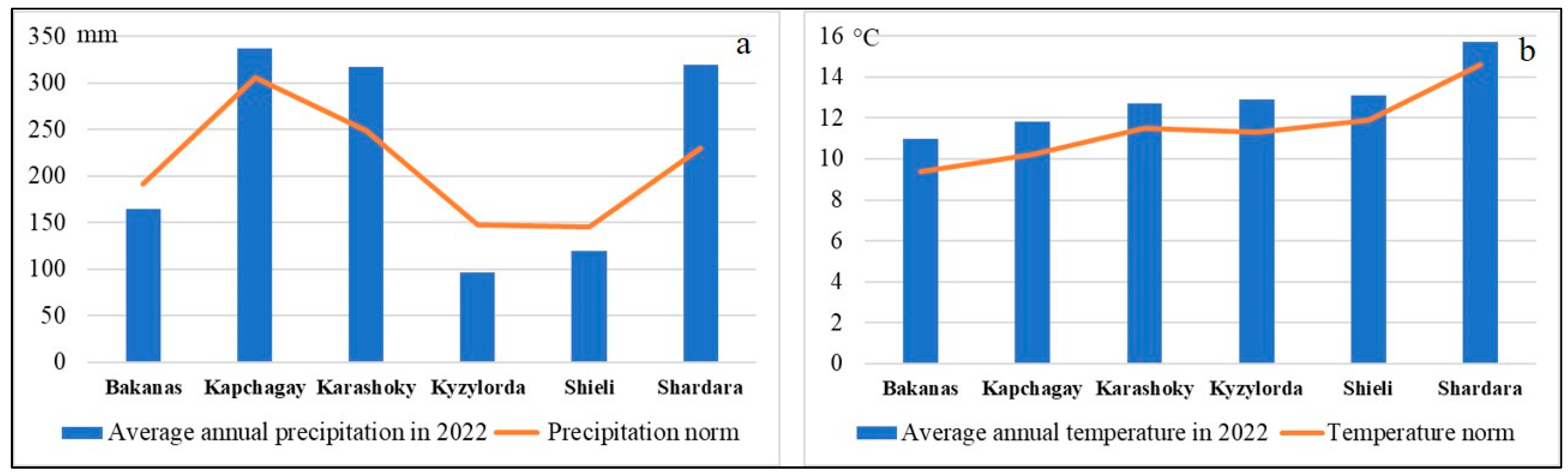
In conjunction with the use of satellite images from 2022 when calculating the distance to watercourses and the normalized difference vegetation index (NDVI), we considered it necessary to analyze the climatic conditions of this year. The analysis of the climatic conditions in 2022 showed that, in terms of temperature, the norm was exceeded by 7–17%. In terms of precipitation, the norm was exceeded by 10–39% at the Shardara, Kapchagay, and Karashoky stations, while at the rest of the stations, the values were 14–39% lower than the norm (
Figure 5).
3.3.2. Soil Conditions
An analysis of the morphogenetic properties of the soils of the Ili and the Syr Darya River Valleys was carried out to obtain comparative data characterizing the edaphic conditions of the communities with the participation of
P. pruinosa. On the floodplain terraces of the Ili River, the floodplain meadow–forest soils of the hydromorphic regime were distributed, forming on the alluvial deposits under the tugai woodlands with meadow cover [
17]. The fresh or slightly saline groundwater from the above-floodplain terraces lied at a depth of 1–3 m and had a chloride-sulfate bicarbonate composition. The soils were layered with alternating interlayers of light texture. The differentiation of the profile into horizons was weakly expressed. In the upper part, there was a low-power (up to 5 cm) turf horizon of dark gray color. Signs of redox processes in the form of rusty and bluish-gray gley spots were noted on the horizon transitional to the soil-forming rock. The buried horizons from the previous stage of soil formation were found at different depths, differing in terms of a dark color and a compacted composition. The humus content in the profile was low and did not exceed 0.1–0.34%, while the number of carbonates was also low (1.5–3.4%) with a maximum in the upper part. The reaction of the soil solution was slightly alkaline (pH = 7.8–8.6) with an increase in the alkalinity with the depth. There was no salinity in a half-meter layer (0.126–0.198%). The absorbed bases were low in the range of 5–6.7 mg-eq per 100 g of soil. The soils were sandy with a predominance of fine (41.3%) and medium (52.2%) sand fractions [
17].
Turanga woodlands were also formed in depressions of the foothill plain by erosive processes in the form of dried riverbeds and temporary watercourses, developing on forest–meadow soils with shallow occurrence (2–5 m) or the wedging of slightly saline groundwater, springs, and additional surface moistening. Such a site was located at a distance of 6.5 km from the Ili River. The soil-forming rocks were layered with sand and pebble deposits. The soil profile was characterized by a weak morphological differentiation, a small thickness of the humus horizon (43 cm), and a carbonate-illuvial horizon with rusty spots under it. The humus content was low and did not exceed 0.79–1.41%, with a sharp drop in the depth of up to 0.27%. The soils were carbonate, the largest number of carbonates was found in the surface horizon (8.55%). In the carbonate horizon, the value changed to 5.06%, and it decreased to 2.43–2.53% in the central part of the profile. The reaction of the soil solution was slightly alkaline, with a pH = 8. The absorption capacity was low in the range of 16–17 mg-eq per 100 g of soil. The upper part of the profile was not saline (0.1%). The salinity increased with the depth (up to 0.431–0.797%), which was due to the influence of mineralized groundwater. The type of salinity was chloride-sulfate, and the degree of salinity was weak. The texture of the soils was heavy loam with a predominance of fine sand fractions (29.76%) and fine dust (21.70%).
Floodplain meadow–forest soils developed on elevated areas of the riverine floodplain and the riverine ramparts of the Syr Darya River [
16,
49]. Fresh and slightly mineralized groundwater was found at depths from 1–3 to 2–5 m. The soil-forming rocks were alluvial deposits of a light granulometric composition. The soil profile was layered with alternating interlayers of different thicknesses and textures (medium-loamy, light-loamy, and sandy varieties), with a predominance of dusty fractions. A turf horizon of 2–3 cm was allocated at the surface. The thickness of the humus horizon was 30–50 cm. The buried humus horizons were found at various depths. The upper horizon was painted dark gray and had a fragile dusty–lumpy structure. The lower horizons were distinguished by the presence of signs of redox processes in the form of rusty spots. The humus content varied from 1.0 to 3.0%. The reaction of the soil solution was alkaline and highly alkaline, pH = 8.1–9.0, with an increase in the alkalinity with the depth. The number of absorbed bases was low, within 10 mg-eq per 100 g of soil. Depending on the depth and granulometric composition of the sediments, there were unsalted and saline soils. The salts were concentrated mainly in the upper horizon with more desalinated deep layers. The amount of salt ranged from 0.749% in sandy soils to 0.953–2.811% in light-loamy soils and up to 3.125% in medium-loamy varieties. The type of salinity was chloride-sulfate and magnesium–calcium sulfate, and the degree of salinity varied from weak to very strong. The high salinity (2.165%) of the lower part of the profile was due to the influence of mineralized groundwater. The soils had a large reserve of moisture available for plants.
According to the presented morphological features, the edaphic conditions of the turanga communities were forest–meadow and floodplain meadow–forest soils. The floodplain meadow–forest soils were distributed in the Ili and the Syr Darya River Valleys. The soil profile had a weak formation and an indistinctly pronounced differentiation into genetic horizons (i.e., layered). There were signs of a past hydromorphic stage of soil formation in the form of iron oxides and buried humus horizons. The floodplain meadow–forest soils had a low level of fertility with a low humus content, a low absorption capacity, and alkaline or slightly alkaline reactions to the soil solutions. They were low in carbonate but had a favorable air regime and were well provided with moisture available for plants. The forest–meadow soils of the depressions of the foothill plain at a distance of 6.5 km from the Ili River developed with additional moistening on the layered sand and pebble deposits. These soils differed in a relatively formed profile and in the isolation of the humus horizon. They had a slightly alkaline reaction to the soil solution and belonged to the saline type, with a weak degree of the chloride-sulfate type of salinization and heavy-loam varieties of the soil texture.
3.3.3. Ecological Niche Modeling and Mapping
The ecological niche modeling of a species includes calculating all the environmental variables that are significant for the species, taken in their optimal ranges, and finding the intersection of all the favorable conditions. To identify the variables, we considered the geographically linked meeting points of the studied species, climatic and soil conditions [
50,
51], relief characteristics (SRTM) and raster sets of the environmental variables (WorldClim [
42,
48]), and distances to watercourses and settlements. A total of 102 environmental variables were used. The sources and optimal ranges for
P. pruinosa are presented in
Table S2.
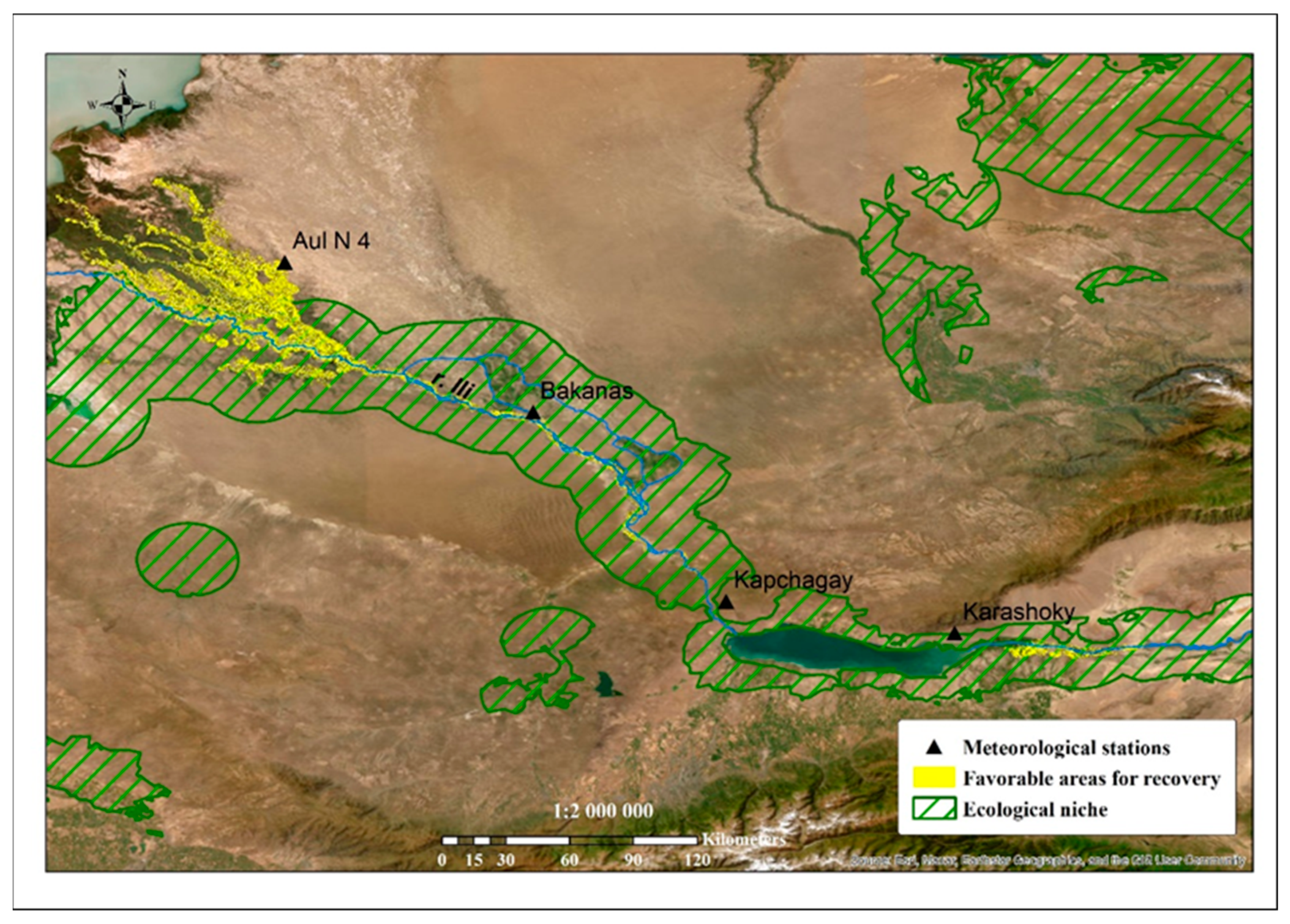
As a result of sampling the correspondence of the favorable environmental variables from 90% and above, an extensive territory was determined (
Figure 6, indicated by a black border). The main limiting factors that determined the possibility of the spreading (reproduction) of turanga were the soil moisture and the frequency of flooding, which promoted seed germination and clonal growth [
15,
21,
52]. In this regard, we calculated the distances to the nearest watercourse using high spatial resolution satellite images (Sentinel-2). The resulting distance ranged from 0 to 17 km, thereby determining the size of the buffer zone around the reservoirs. To isolate this zone and determine the optimal range of turanga growing areas, water bodies were identified according to the remote sensing data at a low spatial resolution (Terra/MODIS). As a result of the calculations, the ecological niche of
P. pruinosa (
Figure 6) was determined.
The final stage of these studies was to determine the most favorable areas for the restoration of turanga groves. For this task, we considered it necessary to expand a larger scale. When performing the calculations based on the ecological niche of the species, we used two limiting factors: the most favorable distance to watercourses and the type of soil. Thus, the area for ecological restoration was determined (
Figure 7 and
Figure 8).
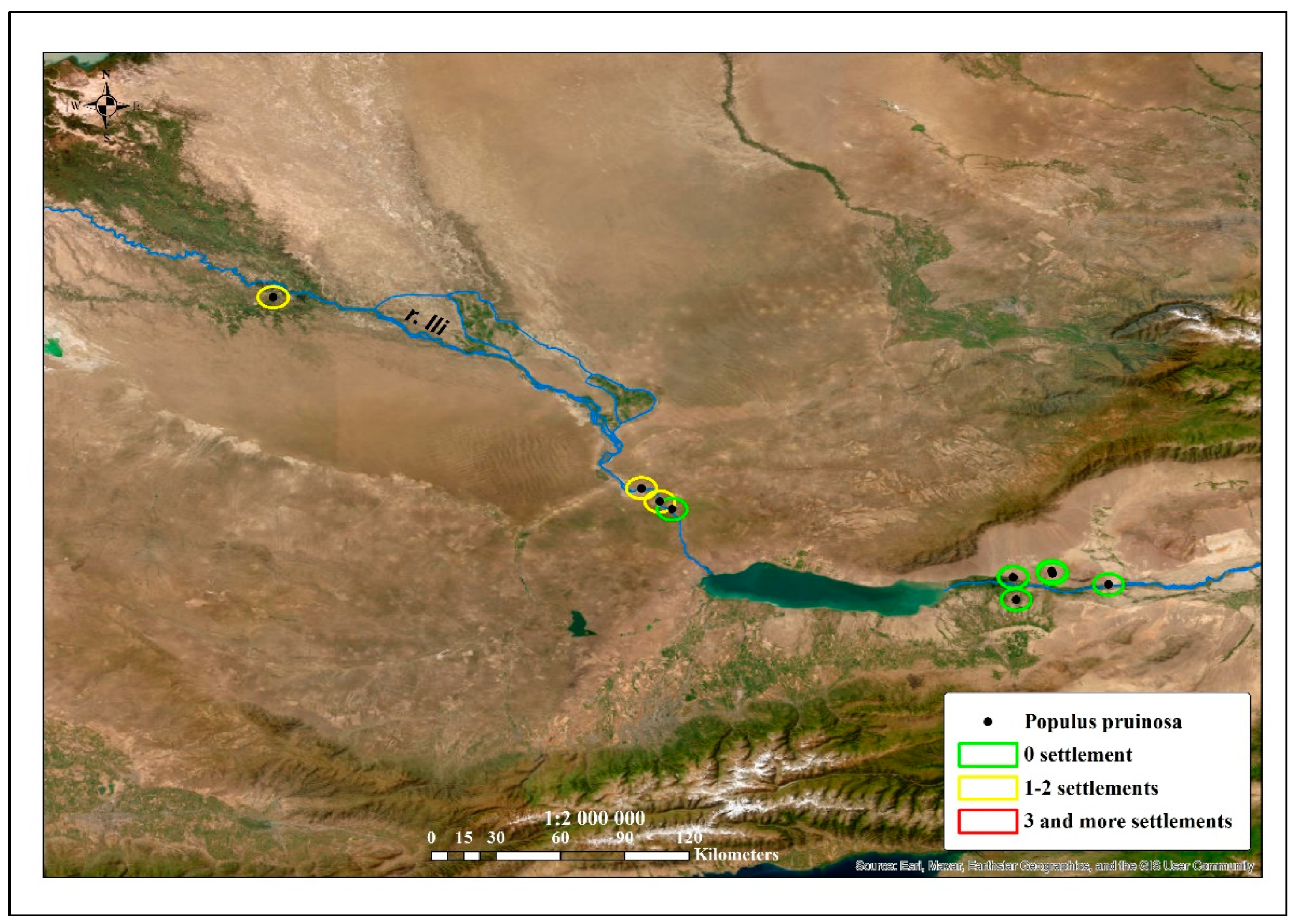
Overgrazing, deforestation, and fires have led to a reduction in the areas of tugai forests [
53] and these influencing factors were primarily the result of their proximity to settlements (
Figure 9 and
Figure 10).
A total of six turanga groves spread along the Ili River are under state protection: five points in the territory of the Altyn–Emel National Park and one grove in the territory of the Balkhash State nature sanctuary. The calculation of the distance of turanga forests to settlements showed that some were located within a five-kilometer zone of several villages, which indicated a high risk of extinction for these groves. Thus, the territories where this species is threatened with extinction have been identified. These sites, concentrated along the Syr Darya River, do not have a conservation status, but they are in need of protection.
4. Discussion
Turanga forests are a special relict type of tugai vegetation that are common in the floodplains of rivers in Central Asia, which have preserved the features of the tertiary flora [
54]. From a global perspective, tugai forests are a unique and threatened floodplain ecosystem found in the desert regions of Central Asia. These forests have the highest biodiversity and provide environmental benefits and natural resources for the human populations in these regions [
55].
The area of modern tugai forests in Central Asia is currently declining. In all places, there is evidence of their degradation and desertification, the halophytization and simplification of their structure, a reduction in species diversity, and a decrease in productivity [
56,
57]. The reasons for the reduction stem from natural climatic changes, the aridization of the territory, and the regulation of the river flows, together with the building of hydrotechnical constructions on the rivers and the absence of floods for rivers with a regulated regime, which leads to an absent possibility for natural renewal. Only the most severe floods, which cannot be regulated anthropogenically, work to ensure the preservation of the rapidly dying tugai forests. Only then is sufficient natural seed renewal facilitated. In the years with an average water supply, turanga does not renew. In addition, turanga is not conducive to artificial afforestation, either by seeds or cuttings [
5,
12,
58].
This is due to the peculiarities of biology. The most important of these peculiarities is the ability of the seeds of tugai species to germinate only in moist unsalted alluvium. In modern conditions of flow regulation and anthropogenic impact, the natural renewal strategy of tugai communities is almost universally impossible. The artificial restoration of tugai requires the essential flooding of territories and the strict selection of areas suitable for reforestation. In the Amu Darya River Valley in Uzbekistan, these areas were selected based on an assessment of the groundwater level and soil salinity [
59]. For restoration of turanga forests, a groundwater level within the range of 2–2.5 m is required. For seed renewal, surface flooding of the territory should occur for 10–15 days during the fruiting period, followed by a month where the ground water level is maintained at 0.3–0.5 m. The soils should be represented by unsalted types. For artificial reforestation, different water qualities can be used (i.e., river or collector drainage), as well as different regimes of watering, ranging from minimal rare initial watering with a lack of water to annual artificial flooding.
In this context, our experience in modeling the ecological niche helped to identify favorable habitats for the restoration of the turanga tugai forests in Kazakhstan. For this purpose, we analyzed the climatic and soil conditions of poplar growth. The climatic conditions corresponded to the following ranges. The average monthly temperatures during the year ranged from −11.8 to +29.2 °C, and the average annual air temperature ranged from +8.3 to +14.6 °C. All the warmest values were noted at stations near the Syr Darya River, and the coldest ones were noted along the Ili River. The average annual precipitation ranged from 117 to 306 mm, with the greatest precipitation in November and spring. The minimum was recorded from June to October, and the average annual number of days with precipitation varied from 32 to 60 days.
Due to some discrepancy between the optimal ranges of ground-based and remote data (WorldClim, and SRTM) during the calculations, we deliberately expanded the territory of the potential distribution of P. pruinosa for some parameters. We used these data to develop maps of the most favorable areas for the ecological restoration of the turanga poplar. The most vulnerable forests located within 5 km of settlements that were in need of protection were also identified.
Our field research confirmed a dangerous trend in changing the range and living conditions for turanga forests in Kazakhstan. In the past, the reason for the reduction in these areas was logging. Currently, the changing of the flood regime can cause a decrease in and even the absence of seed renewal, leading to the further degradation of existing woodlands. In addition, anthropogenic factors such as overgrazing, fires, and recreation have led to the spread of weed species and created competition for water resources with turanga seedlings. In some areas, weeds accounted for 32% of the cenoflora [
60].
Studies on soil cover have shown that floodplain meadow–forest soils have a favorable air regime and are well supplied with moisture that is available for plants. Therefore, for the natural maintenance of the existing plants and the expansion of their area, our efforts should focus on creating conditions for the germination of seeds and the formation of root offspring.
As recommendations for the natural restoration of tugai forests, we can draw on the experience of the Tarim River (Xinjiang, China) where, during the germination period from the middle of July to the middle of August, the artificial regulation of water resources is carried out through the creation of ecological irrigation channels [
61].
The volume and duration of floods affects the clonal reproduction of natural woodland plants [
21]. Therefore, research should focus on determining the timing of flooding, creating artificial drainage, and diverting water for irrigation after flooding forestland, which are of great importance for effectively utilizing seasonal flooding in arid areas.
Floods carry seeds to germination sites and also moisten the soil, facilitating seed germination abilities. In the following year, watering is necessary to replenish the moisture reserves that are required for the vital activity of seedlings and for ensuring that they survive during winter and through the summer drought [
62].
Undoubtedly, the reasonable regulation of water resources is necessary for the conservation of populations. Over the past 20 years, a water release program has been conducted in the lower reaches of the Tarim River. We recommend focusing the diversion of additional (“ecological”) water to the stands with shallow groundwater in order to preserve the functionality of forests and their ability to regenerate from seeds [
22].
Currently,
P. pruinosa is listed as a protected species in Syr Darya in the Turkestan Regional Natural Park, Altyn–Emel, and the Sharyn National Parks. Given that the reduction in woodlands is occurring at a rapid pace, it is necessary to expand the network of protected areas of different statuses (regional, national) to enable conservation and further study. The need to create genetic reserves was discussed as early as in 1973 [
12], and its necessity is confirmed by modern studies of tugai forests in river valleys and the need to preserve their gene pool [
18]. It has been shown that mass seed renewal occurs in areas located close to the mother plants (100–600 m) with sufficient soil moisture and good aeration and illumination. The change in the hydrological regime of rivers leads to the drying and salinization of soils, which contributes to the deterioration of seed renewal conditions. Measures for the conservation of the turanga forest in the Ili-Balkhash region include forestry, biotechnological methods, introduction methods, and recommendations for the creation of nurseries using seed sowing along the ridges alternating between irrigation furrows [
63].
Uzbekistan has experience in organizing nature reserves among farmlands in the developed deltas of the arid zone, based on the concept of a “polarized landscape” proposed by B.B. Rodoman [
64,
65]. The optimal degree of the combination of cultivated lands and the natural landscape was proposed, including the water supply of fragmented areas in the zone of economic development due to the rational placement of fields and irrigation networks. At the same time, an eco-network of specially protected areas was formed. It was built along water protection zones and allowed for the preservation of biodiversity [
66,
67].
To restore the tugai forest into potentially suitable habitats, it is necessary to use high-quality seedlings. The artificial cultivation of turanga from seeds and cuttings in Kazakhstan began in the 1950s–70s, but the results were not encouraging. We returned to this issue in the new millennium. The results were obtained and methods were developed in the Republican Forest Breeding and Seed Center [
68]. The planting material has already been planted in the southeast of Kazakhstan (the Balkhash region). The Mangyshlak Experimental Botanical Garden (Aktau, Western Kazakhstan) has experience in cultivating turanga seedlings from root offspring [
69], which were used in landscaping settlements. Microclonal reproduction should be considered a promising method for growing planting material, and it has been tested on poplar species, including turanga [
70,
71,
72].
5. Conclusions
Our research showed the real state of the tugai forests formed by P. pruinosa in Kazakhstan. The habitats of the rare species were associated with floodplains of desert rivers—the Syr Darya and the Ili. Turanga forests were confined to floodplain meadow–forest soils formed on alluvial deposits under a tugai forest with a developed grass layer. Groundwater was located at a depth of 1–3 m. The soils were layered with alternating river alluvial deposits of a light texture. The differentiation into horizons was weakly expressed, and the buried horizons of previous stages of soil formation were present. The floodplain meadow–forest soils had a low level of fertility with a low humus content (0.1–3.0%), a low absorption capacity, and an alkaline or slightly alkaline reaction to the soil solution (pH = 7.8–9.0). They had a favorable air regime and were well stocked with moisture that was available for plants. The forest–meadow soils in the depressions of the foothill plain differed in a relatively formed profile and an isolation of the humus horizon, had a weak degree of salinization, and their soil texture was of heavy-loam varieties.
The floristic composition of the turanga forests included 223 species of vascular plants from 136 genera and 42 families. A total of 165 species grow in the Ili River Valley and 119 species in the Syr Darya River Valley. The similarity coefficient was 0.42, which was due to botanical and geographical features. The species richness was slightly higher in the Ili River Valley. Weeds accounted for 21 to 24% of the species composition, which indicated an anthropogenic disturbance.
The turanga forests were characterized by a well-defined three-layer vertical structure of vegetation cover. The forestry assessment showed that the vital condition of the turanga in the Ili River Valley was better than that in the Syr Darya River Valley, where there was a high closeness of the stand and the seed renewal was disrupted.
The current locations of P. pruinosa in Kazakhstan were also analyzed. The map of the ecological niche was compiled, on the basis of which potential habitats for the growth of the species and places for ecological restoration were identified. The most vulnerable forests at risk of extinction were located within 5 km of settlements along the Syr Darya River. It is urgently required to create a network of protected areas and genetic reserves in the valleys of the Syr Darya and the Ili Rivers, in order to preserve the gene pool of turanga.
We recommend planning measures for the conservation and restoration of turanga forests in the valleys of the Ili and Syr Darya desert rivers as part of a comprehensive study on the current state of the ecosystems and the selection of territories with favorable conditions for possible seed renewal. The studies of the characteristics of the conditions should include an assessment of morphological features and the physico-chemical properties of the soils of a predominantly light texture in the absence of surface and profile salinization (the amount of salts within 0.3%), and the depth of fresh or slightly mineralized groundwater up to 2–3 m.
After selecting the areas for restoration, methods should be determined to implement an irrigation (or flood) moisture regime for the period of seed germination (up to 15 days). In the case of planting seedlings, a watering system should be developed to maintain the soil moisture across two growing seasons.



 ,
, {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}









