
Mammalian Roadkill in a Semi-Arid Region of Brazil: Species, Landscape Patterns, Seasonality, and Hotspots
 , ,
, ,  and
and
Abstract
:1. Introduction
2. Materials and Methods
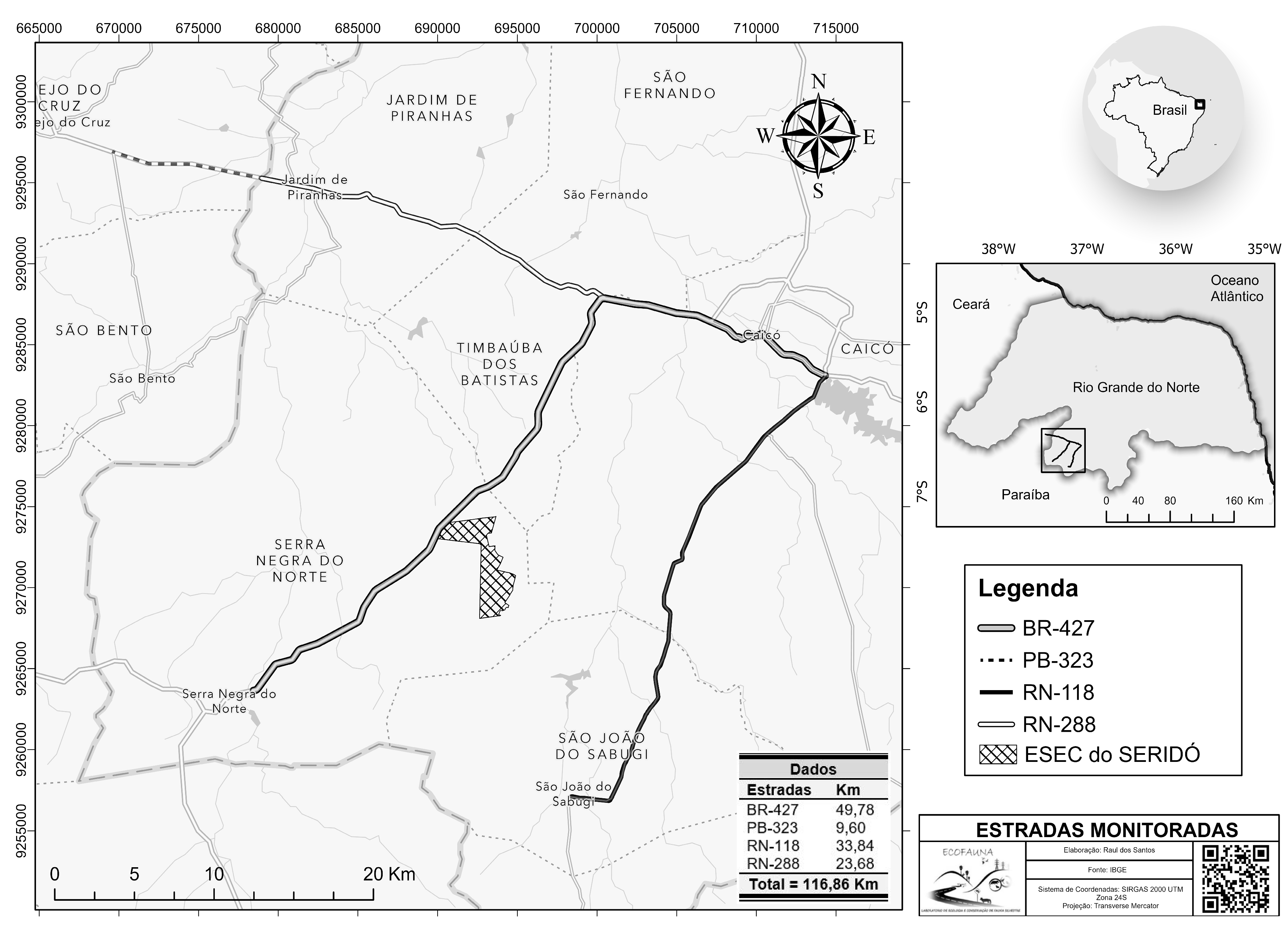
2.1. Study Area
2.2. Roadkill Data
2.3. Landscape and Seasonal Variables
2.4. Statistical Analysis
2.4.1. Hotspots
2.4.2. Landscape versus Roadkill
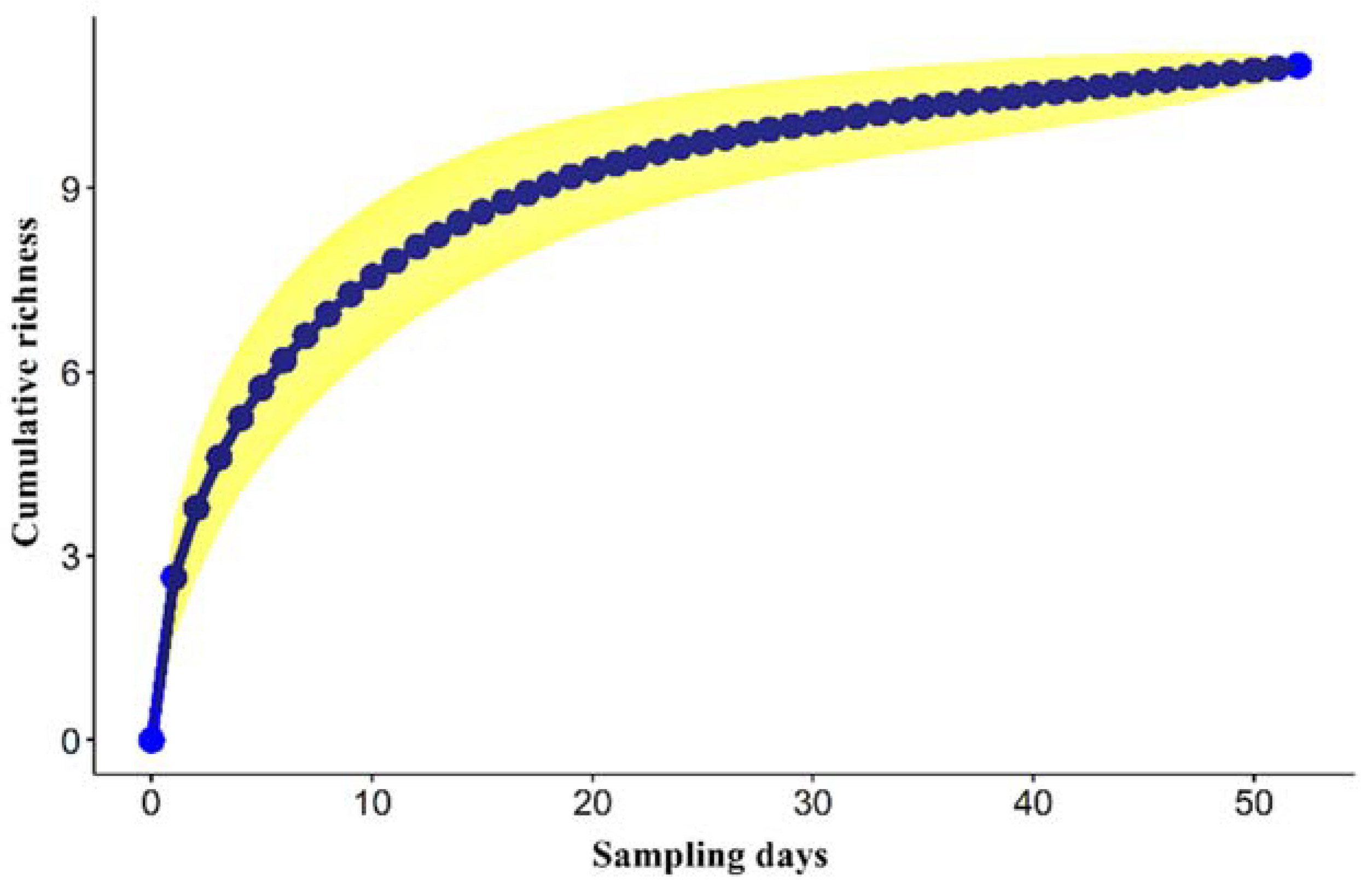
3. Results
3.1. LULC Classification
3.2. Roadkill
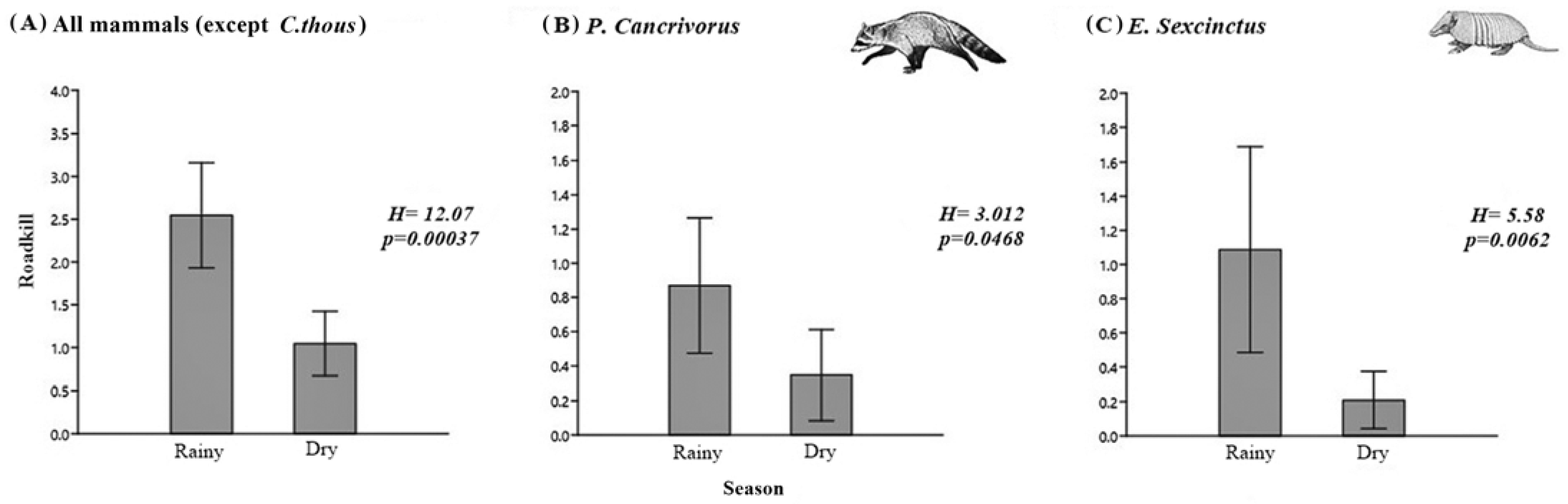
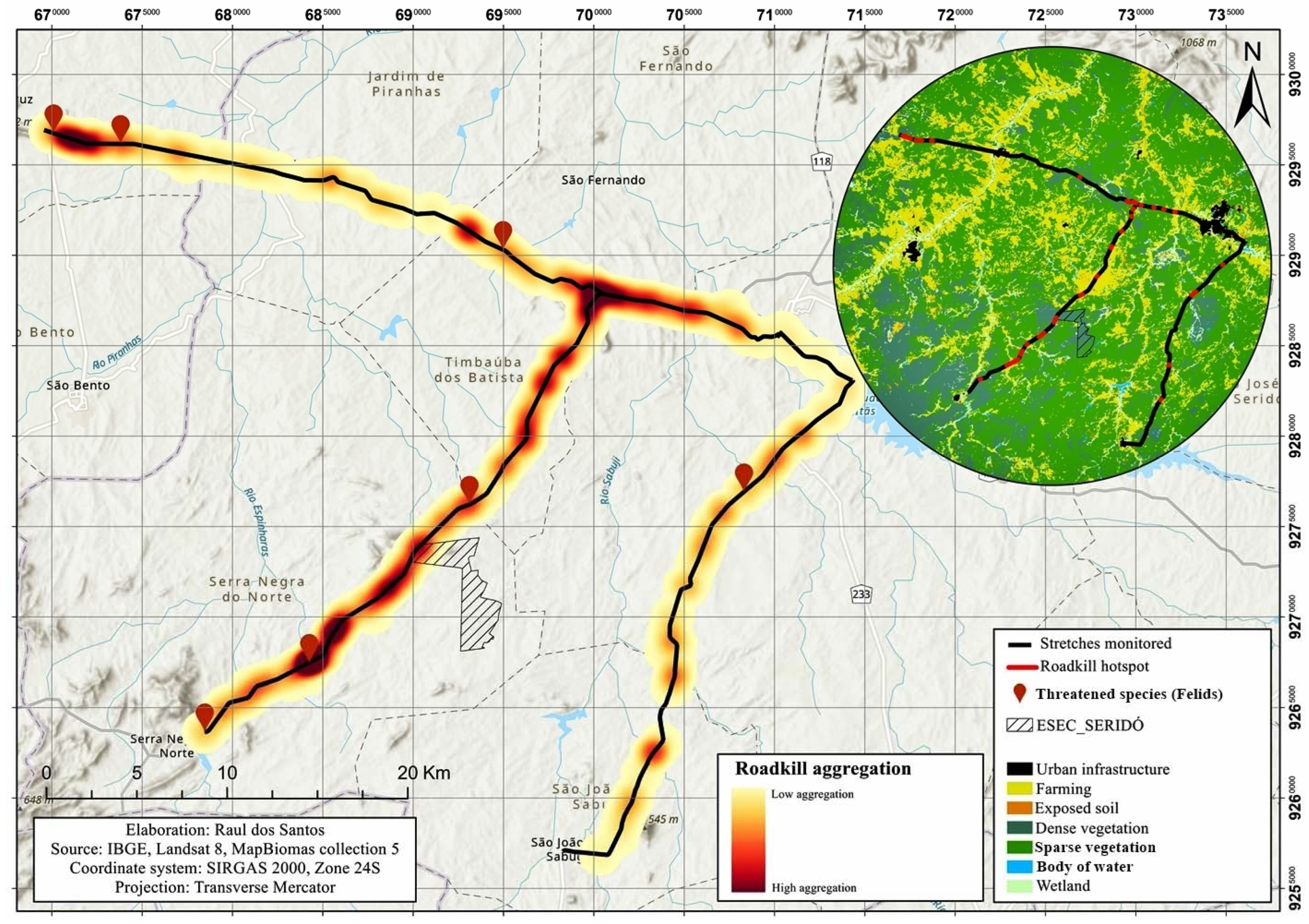
3.3. Roadkill Aggregations and Identification of Hotspots
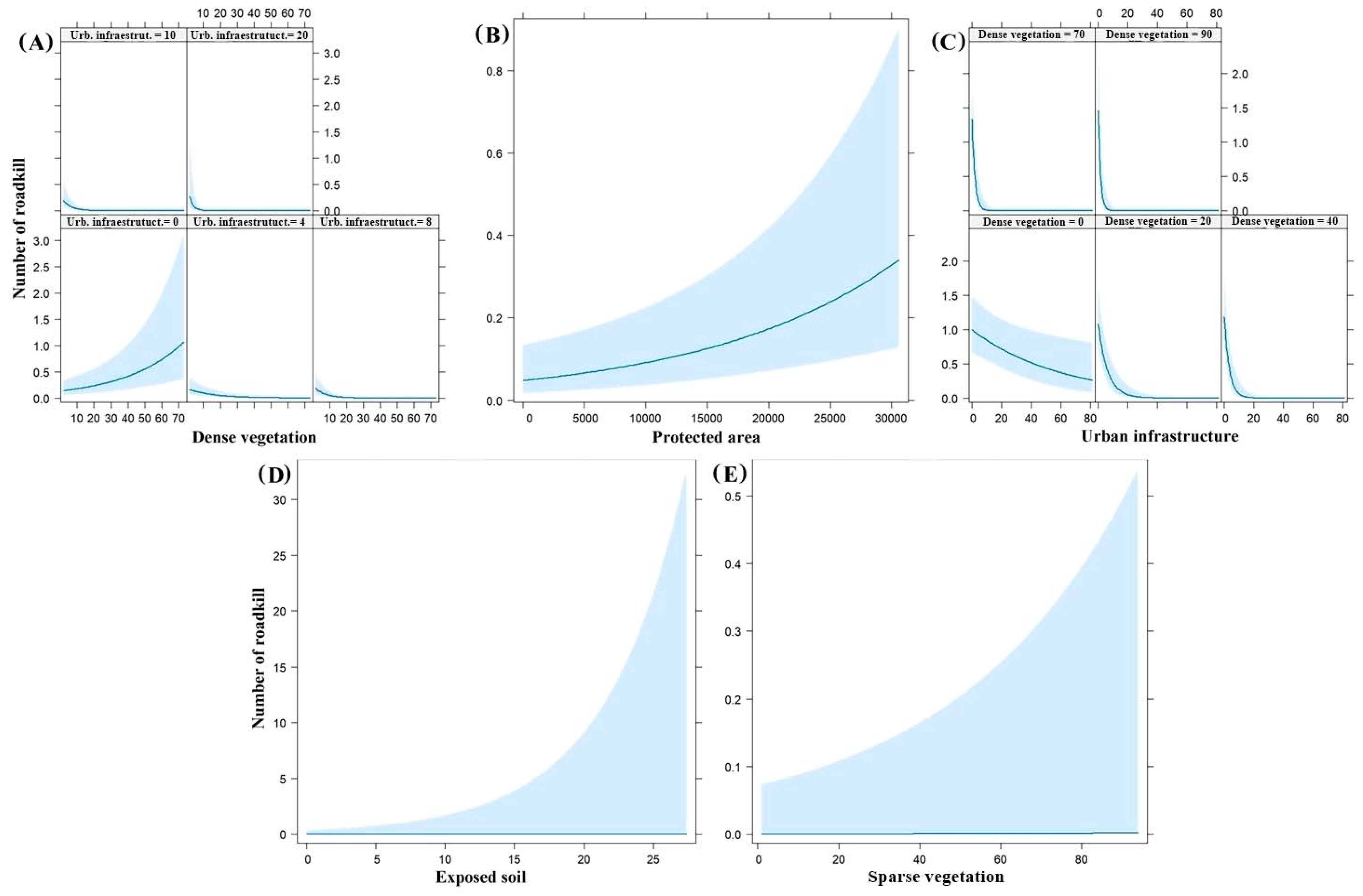
3.4. Landscape Variables Associated with Roadkill
4. Discussion
- I
- The erection of signposts, showing color images of the species most vulnerable to roadkill, together with instructions for the driver on the need to reduce speed to increase their reaction time when confronted with an animal on the road. Signposts should be installed at at least six of the principal hotspots identified in the present study, including four of which are located on the federal highway (684543W, 9267232S; 693296W, 9276207S; 697918W, 9283630S, and 700332W, 9287730S) and two on state highways (672244W, 9296095S and 693573W, 9290888S). These hotspots were selected because, in addition to their relevance for the three principal species, they are also important for threatened species, such as L. emiliae and H. yagouaroundi. It is important to note that each hotspot must have two signposts at an interval of approximately 1 km, to warn drivers approaching from both directions.
- II
- Installation of speed-reducing devices (in accordance with the national traffic legislation), such as electronic monitoring systems or physical structures, such as speed bumps or studs. These devices should be installed after the signposts and within the area of the hotspot, to provide a backup mechanism that contributes to the effectiveness of the signposts.
- III
- The excavation of tunnels under the road as a complementary, but relatively costly measure, which may be more or less viable, depending on the size of the target species and the topography of the region (the need to avoiding flooding in the rainy season).
- IV
- Installation of containment (guide) fences in the vicinity of crossing points, to divert animals and oblige them to cross the road near the speed-reducing devices or in more appropriate stretches of the road.
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Trombulak, S.C.; Frissell, C.A. Review of Ecological Effects of Roads on Terrestrial and Aquatic Communities. Conserv. Biol. 2000, 14, 18–30. [Google Scholar] [CrossRef] [Green Version]
- Bager, A.; Da, P.; Lucas, S.; Bourscheit, A.; Kuczach, A.; Maia, B. Os Caminhos Da Conservação Da Biodiversidade Brasileira Frente Aos Impactos Da Infraestrutura Viária. Biodivers. Bras. BioBrasil 2016, 6, 75–86. [Google Scholar]
- Morelli, F.; Benedetti, Y.; Delgado, J.D. A Forecasting Map of Avian Roadkill-Risk in Europe: A Tool to Identify Potential Hotspots. Biol. Conserv. 2020, 249, 108729. [Google Scholar] [CrossRef]
- Grilo, C.; Borda-de-Água, L.; Beja, P.; Goolsby, E.; Soanes, K.; le Roux, A.; Koroleva, E.; Ferreira, F.Z.; Gagné, S.A.; Wang, Y.; et al. Conservation Threats from Roadkill in the Global Road Network. Glob. Ecol. Biogeogr. 2021, 30, 2200–2210. [Google Scholar] [CrossRef]
- Pinto, F.A.S.; Clevenger, A.P.; Grilo, C. Effects of Roads on Terrestrial Vertebrate Species in Latin America. Environ. Impact Assess. Rev. 2020, 81, 106337. [Google Scholar] [CrossRef]
- CBEE. Centro Brasileiro De Estudos Em Ecologia De Estradas Brasil: Atropelamentos de Fauna Selvagem. Available online: https://sistemaurubu.com.br/dados/ (accessed on 18 March 2023).
- Dornas, R.A.P.; Kindel, A.; Bager, A.; Freitas, S.R. Avaliação Da Mortalidade de Vertebrados Em Rodovias No Brasil. In Ecologia de Estradas: Tendências e Pesquisas; Bager, A., Ed.; UFLA: Lavras, Brazil, 2012; Volume 1, pp. 139–152. ISBN 978-85-903770-3-0. [Google Scholar]
- González-Suárez, M.; Zanchetta Ferreira, F.; Grilo, C. Spatial and Species-Level Predictions of Road Mortality Risk Using Trait Data. Glob. Ecol. Biogeogr. 2018, 27, 1093–1105. [Google Scholar] [CrossRef] [Green Version]
- Pinto, F.A.S.; Cirino, D.W.; Cerqueira, R.C.; Rosa, C.; Freitas, S.R. How Many Mammals Are Killed on Brazilian Roads? Assessing Impacts and Conservation Implications. Diversity 2022, 14, 835. [Google Scholar] [CrossRef]
- Dean, W.R.J.; Seymour, C.L.; Joseph, G.S.; Foord, S.H. A Review of the Impacts of Roads on Wildlife in Semi-Arid Regions. Diversity 2019, 11, 81. [Google Scholar] [CrossRef] [Green Version]
- Garda, A.A.G.; Lion, M.B.; Lima, S.M.D.Q.; Mesquita, D.O.; Araujo, H.F.P.D.; Napoli, M.F. Os Animais Vertebrados Do Bioma Caatinga. Cienc. Cult. 2018, 70, 29–34. [Google Scholar] [CrossRef] [Green Version]
- Lee, E.; Croft, D.B.; Achiron-Frumkin, T. Roads in the Arid Lands: Issues, Challenges and Potential Solutions. In Handbook of Road Ecology; van der Ree, R., Smith, D.J., Grilo, C., Eds.; John Wiley & Sons, Ltd.: Hoboken, NJ, USA, 2015; pp. 382–390. ISBN 9781118568170. [Google Scholar]
- Dixon, J.D.; Oli, M.K.; Wooten, M.C.; Eason, T.H.; Cown, J.W.M.C.; Paetkau, D. Two Florida Black Bear Populations. Conserv. Biol. 2006, 20, 155–162. [Google Scholar] [CrossRef]
- Forman, R.T.T.; Deblinger, R.D. The Ecological Road-Effect Zone of a Massachusetts (U.S.A.) Suburban Highway. Conserv. Biol. 2000, 14, 36–46. [Google Scholar] [CrossRef]
- Laurance, W.F.; Goosem, M.; Laurance, S.G.W. Impacts of Roads and Linear Clearings on Tropical Forests. Trends Ecol. Evol. 2009, 24, 659–669. [Google Scholar] [CrossRef] [PubMed]
- Teixeira, F.Z.; Coelho, I.P.; Esperandio, I.B.; Rosa Oliveira, N.; Porto Peter, F.; Dornelles, S.S.; Rosa Delazeri, N.; Tavares, M.; Borges Martins, M.; Kindel, A. Os Hotsposts de Atropelamentos Nas Estradas São Coincidentes Entre Diferentes Grupos de Vertebrados? Oecologia Aust. 2013, 17, 36–47. [Google Scholar] [CrossRef] [Green Version]
- Santos, E.; Cordova, M.; Rosa, C.; Rodrigues, D. Hotspots and Season Related to Wildlife Roadkill in the Amazonia–Cerrado Transition. Diversity 2022, 14, 657. [Google Scholar] [CrossRef]
- Shimabukuro, A.R.; Santos, R.; Taili, I.; Lima, A.; Freitas, K.; Guimarães, T.; Morlanes, V.; Calabuig, C. Novos Registros e Considerações Sobre a Distribuição Geográfica de Galictis Cuja (Carnivora: Mustelidae) No Rio Grande Do Norte, Brasil. Bol. Do Mus. Para. Emílio Goeldi Ciências Nat. 2022, 17, 545–555. [Google Scholar] [CrossRef]
- Perrino, E.V.; Musarella, C.M.; Magazzini, P. Management of Grazing Italian River Buffalo to Preserve Habitats Defined by Directive 92/43/EEC in a Protected Wetland Area on the Mediterranean Coast: Palude Frattarolo, Apulia, Italy. Euro-Mediterr. J. Environ. Integr. 2021, 6, 32. [Google Scholar] [CrossRef]
- Grilo, C. A Rede Viária e a Fauna: Impactos, Mitigação e Implicações Para a Conservação Das Espécies Em Portugal. In Ecologia de Estradas: Tendências e Pesquisas; Bager, A., Ed.; UFLA: Lavras, Brazil, 2012; Volume 1, pp. 35–58. ISBN 978-85-903770-3-0. [Google Scholar]
- Abra, F.D.; Granziera, B.M.; Huijser, M.P.; de Barros Ferraz, K.M.P.M.; Haddad, C.M.; Paolino, R.M. Pay or Prevent? Human Safety, Costs to Society and Legal Perspectives on Animal-Vehicle Collisions in São Paulo State, Brazil. PLoS ONE 2019, 14, e0215152. [Google Scholar] [CrossRef] [Green Version]
- Melo, E.S.; Santos-Filho, M. Efeitos Da BR-070 Na Província Serrana de Cáceres, Mato Grosso, Sobre a Comunidade de Vertebrados Silvestres. Rev. Bras. Zoociências 2007, 9, 185–192. [Google Scholar]
- Cáceres, N.C.; Casella, J.; Goulart, C.D.S. Variação Espacial e Sazonal Atropelamentos de Mamíferos No Bioma Cerrado, Rodovia BR 262, Sudoeste Do Brasil. Mastozool. Neotrop. 2012, 19, 21–33. [Google Scholar]
- Caires, H.S.; Souza, C.R.; Lobato, D.N.C.; Fernandes, M.N.S.; Damasceno, J.S. Roadkilled Mammals in the Northern Amazon Region and Comparisons with Roadways in Other Regions of Brazil. Iheringia. Ser Zool 2019, 109, 1–9. [Google Scholar] [CrossRef]
- Caro, T.M.; Shargel, J.A.; Stoner, C.J. Frequency of Medium-Sized Mammal Road Kills in an Agricultural Landscape in California. Am. Midl. Nat. 2000, 144, 362–369. [Google Scholar] [CrossRef]
- Caceres, N.C. Biological Characteristics Influence Mammal Road Kill in an Atlantic Forest-Cerrado Interface in South-Western Brazil. Ital. J. Zool. 2011, 78, 379–389. [Google Scholar] [CrossRef]
- Orlandin, E.; Piovesan, M.; Favretto, M.A.; D’Agostini, F.M. Mamíferos de Médio e Grande Porte Atropelados No Oeste de Santa Catarina, Brasil. Biota Amaz. 2015, 5, 125–130. [Google Scholar] [CrossRef]
- Cirino, D.W.; Lupinetti-Cunha, A.; Freitas, C.H.; de Freitas, S.R. Do the Roadkills of Different Mammal Species Respond the Same Way to Habitat and Matrix? Nat. Conserv. 2022, 47, 65–85. [Google Scholar] [CrossRef]
- Moore, L.J.; Petrovan, S.O.; Bates, A.J.; Hicks, H.L.; Baker, P.J.; Perkins, S.E.; Yarnell, R.W. Demographic Effects of Road Mortality on Mammalian Populations: A Systematic Review. In Biological Reviews; Wiley Online Library: Hoboken, NJ, USA, 2023. [Google Scholar] [CrossRef]
- LaRue, M.A.; Nielsen, C.K. Modelling Potential Dispersal Corridors for Cougars in Midwestern North America Using Least-Cost Path Methods. Ecol. Modell. 2008, 212, 372–381. [Google Scholar] [CrossRef]
- Carvalho, C.F.; Custódio, A.E.I.; Júnior, O.M. Agregações de Atropelamentos de Vertebrados Silvestres Na Rodovia Br-050, Minas Gerais, Brasil. Biosci. J. 2015, 31, 951–959. [Google Scholar] [CrossRef] [Green Version]
- Aresco, M.J. Mitigation Measures To Reduce Highway Mortality of Turtles and Other Herpetofauna At a North Florida Lake. J. Wildl. Manag. 2005, 69, 549–560. [Google Scholar] [CrossRef]
- Bager, A.; Piedras, S.R.N.; Tainana, M.S.; Hóbus, Q. Fauna Selvagem e Atropelamento. -Diagnóstico Do Conhecimento Brasileiro. In Áreas Protegidas—Repensando as Escalas de Atuação; Bager, A., Ed.; Bager, Alex: Porto Alegre, Brazil, 2007; Volume 1, pp. 49–62. [Google Scholar]
- Fabrizio, M.; di Febbraro, M.; D’Amico, M.; Frate, L.; Roscioni, F.; Loy, A. Habitat Suitability vs Landscape Connectivity Determining Roadkill Risk at a Regional Scale: A Case Study on European Badger (Meles meles). Eur. J. Wildl. Res. 2019, 65, 7. [Google Scholar] [CrossRef]
- Filius, J.; van der Hoek, Y.; Jarrín-V, P.; van Hooft, P. Wildlife Roadkill Patterns in a Fragmented Landscape of the Western Amazon. Ecol. Evol. 2020, 10, 6623–6635. [Google Scholar] [CrossRef]
- Grilo, C.; Ascensão, F.; Santos-Reis, M.; Bissonette, J.A. Do Well-Connected Landscapes Promote Road-Related Mortality? Eur. J. Wildl. Res. 2011, 57, 707–716. [Google Scholar] [CrossRef]
- Seo, C.; Thorne, J.H.; Choi, T.; Kwon, H.; Park, C.H. Disentangling Roadkill: The Influence of Landscape and Season on Cumulative Vertebrate Mortality in South Korea. Landsc. Ecol. Eng. 2015, 11, 87–99. [Google Scholar] [CrossRef]
- Cândido, J.F., Jr.; Brocardo, C.R. Persistência de Mamíferos de Médio e Grande Porte Em Fragmento de Floresta Ombrófila Mista No Estado Do Paraná, Brasil. Rev. Árvore 2012, 36, 301–310. [Google Scholar]
- Clevenger, A.P.; Chruszcz, B.; Gunson, K.E. Spatial Patterns and Factors Influencing Small Vertebrate Fauna Road-Kill Aggregations. Biol. Conserv. 2003, 109, 15–26. [Google Scholar] [CrossRef]
- Davies, J.M.; Roper, T.J.; Shepherdson, D.D.J. Seasonal Distribution of Road Kills in the European Badger (Meles meles). J. Zool 1987, 211, 525–529. [Google Scholar] [CrossRef]
- Medinas, D.; Marques, J.T.; Mira, A. Assessing Road Effects on Bats: The Role of Landscape, Road Features, and Bat Activity on Road-Kills. Ecol. Res. 2013, 28, 227–237. [Google Scholar] [CrossRef] [Green Version]
- Bastos, D.F.D.O.; Souza, R.A.T.; Zina, J.; da Rosa, C.A. Seasonal and Spatial Variation of Road-Killed Vertebrates on Br-330, Southwest Bahia, Brazil. Oecologia Aust. 2019, 23, 388–402. [Google Scholar] [CrossRef]
- Bager, A.; Alves Da Rosa, C. Priority Ranking of Road Sites for Mitigating Wildlife Roadkill. Biota. Neotrop. 2010, 10, 149–153. [Google Scholar] [CrossRef] [Green Version]
- MMA. Ministério do Meio Ambiente Plano de Manejo ESEC Seridó. Available online: https://www.gov.br/icmbio/ptbr/assuntos/biodiversidade/unidade-de-conservacao/unidades-de-biomas/caatinga/lista-de-ucs/esec-do-serido (accessed on 18 March 2023).
- Quintela, F.M.; DA ROSA, C.A.; Feijó, A. Updated and Annotated Checklist of Recent Mammals from Brazil. An. Acad. Bras. Ciências 2020, 92, 1–57. [Google Scholar] [CrossRef]
- Carmignotto, A.P.; Astúa, D. Mammals of the Caatinga: Diversity, Ecology, Biogeography, and Conservation. In Caatinga: The Largest Tropical Dry Forest Region in South America; Silva, J.M.C.D., Leal, I.R., Tabarelli, M., Eds.; Springer International Publishing: Berlin/Heidelberg, Germany, 2018; pp. 211–254. ISBN 9783319683393. [Google Scholar]
- Bonvicino, C.R.; Bezerra, A.M.R.; Percequillo, A.R.; D’Andrea, P.S. Instituto Chico Mendes de Conservação da Biodiversidade. In Livro Vermelho da Fauna Brasileira Ameaçada de Extinção: Volume II—Mamíferos; ICMBio, Ed.; Instituto Chico Mendes de Conservação da Biodiversidade: Brasília, Brazil, 2018; Volume II. [Google Scholar]
- IUCN. The IUCN Red List of Threatened Species. Available online: http://www.iucnredlist.org (accessed on 18 March 2023).
- MMA—Ministério do Meio Ambiente Salve: Risco de Extinção Da Fauna Brasileira. Available online: https://salve.icmbio.gov.br/#/ (accessed on 18 March 2023).
- MapBiomas—Collection 5 of the Annual Coverage and Land Use Maps Series of the Brazil. Available online: https://mapbiomas.org (accessed on 13 April 2021).
- Queiroz, L.P.; Cardoso, D.; Fernandes, M.F.; Moro, M.F. Diversity and Evolution of Flowering Plants of the Caatinga Domain. In Caatinga: The Largest Tropical Dry Forest Region in South America; Silva, J.M.C.D., Leal, I.R., Tabarelli, M., Eds.; Springer International Publishing: Cham, Switzerland, 2017; pp. 23–63. ISBN 978-3-319-68339-3. [Google Scholar]
- Coelho, A.V.P.; Coelho, I.P.; Kindel, A.; Teixeira, F.Z. Siriema: Road Mortality Software; NERF, UFRGS: Porto Allegre, Brazil, 2014; Volume 2.0. [Google Scholar]
- Bolker, B.M. Ecological Models and Data in R; Princeton University Press: Princeton, NJ, USA, 2008; ISBN 9781400840908. [Google Scholar]
- Burnham, K.P.; Anderson, D.R.; Burnham, K.P. Model Selection and Multimodel Inference: A Practical Information-Theoretic Approach; Springer: New York, NY, USA, 2002; Volume 1, ISBN 0387953647. [Google Scholar]
- Muriel, R.; Ferrer, M.; Balbontín, J.; Cabrera, L.; Calabuig, C.P. Disentangling the Effect of Parental Care, Food Supply, and Offspring Decisions on the Duration of the Postfledging Period. Behav. Ecol. 2015, 26, 1587–1596. [Google Scholar] [CrossRef] [Green Version]
- Grueber, C.E.; Nakagawa, S.; Laws, R.J.; Jamieson, I.G. Multimodel Inference in Ecology and Evolution: Challenges and Solutions. J. Evol. Biol. 2011, 24, 699–711. [Google Scholar] [CrossRef] [PubMed]
- Bolker, B.; Giné-Vázquez, I.; R Core Team. Bbmle: Tools for General Maximum Likelihood Estimation, R package version 0.9; R Core Team: Vienna, Austria, 2010.
- Bates, D.; Mächler, M.; Bolker, B.M.; Walker, S.C. Fitting Linear Mixed-Effects Models Using Lme4. J. Stat. Softw. 2015, 67, 1–48. [Google Scholar] [CrossRef]
- Venables, W.N.; Ripley, B.D. Modern Applied Statistics with S, 4th ed.; Springer: New York, NY, USA, 2002; ISBN 0-387-95457-0. [Google Scholar]
- Fox, J.; Weisberg, S. An {R} Companion to Applied Regression, 3rd ed.; Published online: Thousand Oaks, CA, USA, 2019. [Google Scholar]
- Wickham, H. Ggplot2: Elegant Graphics for Data Analysis; Springer: Cham, Switzerland; New York, NY, USA, 2016; ISBN 978-3-319-24277-4. [Google Scholar]
- Fox, J.; Weisberg, S. Visualizing Fit and Lack of Fit in Complex Regression Models with Predictor Effect Plots and Partial Residuals. J. Stat. Softw. 2018, 87, 1–27. [Google Scholar] [CrossRef]
- Landis, J.R.; Koch, G.G. The Measurement of Observer Agreement for Categorical Data. Biometrics 1977, 33, 159–174. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cezar, H.R.D.A.; Abrantes, S.H.F.; Lima, J.P.R.D.; Medeiros, J.B.D.; Abrantes, M.M.R.; Carreiro, A.D.N.; Barbosa, J.P.D.L. Mamíferos Silvestres Atropelados Em Estradas Da Paraíba, Nordeste Do Brasil. Braz. J. Dev. 2021, 7, 48037–48049. [Google Scholar] [CrossRef]
- Araujo, L.A.D.F.; Hannibal, W.; Costa, R.R.G.F.; Rossi, R.F.; Claro, H.W.P. Effects of Landscape on Roadkill of Medium and Large-Sized Mammals in Southern Goiás, Brazil. Oecologia Aust. 2020, 24, 164–172. [Google Scholar] [CrossRef]
- Oliveira, D.D.S.; Silva, V.M.D. Vertebrados Silvestres Atropelados Na BR 158, RS, Brasil. Biotemas 2012, 25, 229–235. [Google Scholar] [CrossRef]
- Teixeira, F.Z.; Rytwinski, T.; Fahrig, L. Inference in Road Ecology Research: What We Know versus What We Think We Know. Biol. Lett. 2020, 16, 20200140. [Google Scholar] [CrossRef]
- Teixeira, F.Z.; Coelho, A.V.P.; Esperandio, I.B.; Kindel, A. Vertebrate Road Mortality Estimates: Effects of Sampling Methods and Carcass Removal. Biol. Conserv. 2013, 157, 317–323. [Google Scholar] [CrossRef]
- Marinho, P.H.; Bezerra, D.; Antongiovanni, M.; Fonseca, C.R.; Venticinque, E.M. Mamíferos de Médio e Grande Porte Da Caatinga Do Rio Grande Do Norte, Nordeste Do Brasil. Mastozool. Neotrop. 2018, 25, 345–362. [Google Scholar] [CrossRef]
- Alves, R.R.N.; Feijó, A.; Barboza, R.R.D.; Souto, W.M.S.; Fernandes-Ferreira, H.; Cordeiro-Estrela, P.; Langguth, A. Game Mammals of the Caatinga Biome. Ethnobiol. Conserv. 2016, 5, 1–51. [Google Scholar] [CrossRef] [Green Version]
- Feijó, A.; Langguth, A. Mamíferos de Médio e Grande Porte Do Nordeste Do Brasil: Distribuição e Taxonomia, Com Descrição de Novas Espécies. Rev. Nord. Debiologia 2013, 22, 3–225. [Google Scholar]
- Dias, D.M.; Guedes, P.G.; Silva, S.S.P.; Sena, L.M.M. Diversidade de Mamíferos Não Voadores Em Uma Área de Caatinga Do Nordeste Do Brasil. Neotrop. Biol. Conserv. 2017, 12, 200–208. [Google Scholar] [CrossRef] [Green Version]
- Dias, D.D.M.; Bocchiglieri, A. Riqueza e Uso Do Habitat Por Mamíferos de Médio e Grande Porte Na Caatinga, Nordeste Do Brasil. Neotrop. Biol. Conserv. 2016, 11, 38–46. [Google Scholar] [CrossRef]
- Delciellos, A.C. Mammals of Four Caatinga Areas in Northeastern Brazil: Inventory, Species Biology, and Community Structure. Check List 2016, 12, 1916. [Google Scholar] [CrossRef] [Green Version]
- Ramos-Abrantes, M.M.; Carreiro, A.D.N.; Araújo, D.V.F.D.; Souza, J.G.D.; Lima, J.P.R.D.; Cezar, H.R.D.A.; Leite, L.S.; Abrantes, S.H.F. Vertebrados Silvestres Atropelados Na Rodovia BR-230, Paraíba, Brasil. Pubvet 2018, 12, 139. [Google Scholar] [CrossRef]
- Zocche, J.; Costa, S.; Zocche-de-Souza, P.; Viana, I.; Mattia, D.; Scussel, C.; Zocche, C.; Pereira, J.; Carvalho, F. Vertebrados Silvestres Atropelados Em Rodovias Do Sul de Santa Catarina, Brasil. In Geoprocessamento na Análise Ambiental; UNESCO: Paris, France, 2020; pp. 252–289. [Google Scholar]
- Brum, T.R.; Santos-Filho, M.; Canale, G.R.; Ignácio, A.R.A. Efeitos Das Rodovias Mt 235 e 358 Sob a Diversidade de Vertebrados No Sudoeste de Mato Grosso. Braz. J. Biol. 2018, 78, 125–132. [Google Scholar] [CrossRef] [Green Version]
- Hegel, C.G.Z.; Consalter, G.C.; Zanella, N. Mamíferos Silvestres Atropelados Na Rodovia RS-135, Norte Do Estado Do Rio Grande Do Sul. Biotemas 2012, 25, 165–170. [Google Scholar] [CrossRef] [Green Version]
- Beisiegel, B.D.M.; Lemos, F.G.; Azevedo, F.C.D.; Queirolo, D.; Jorge, R.S.P. Avaliação Do Risco de Extinção Do Cachorro-Do-Mato Cerdocyon Thous (Linnaeus, 1766) No Brasil. Biodivers. Bras. 2013, 1, 138–145. [Google Scholar]
- Lima, L.A.Q.; de Oliveira, T.M.A.; Maia, T.G.; Shimabukuro, A.; Taili, I.; dos Santos, R.; Calabuig, C. Predation of Boa Constrictor (Boidae) by Cerdocyon Thous (Canidae) in Caatinga in Brazil. Pak. J. Zool 2022, 54, 1935–1937. [Google Scholar] [CrossRef]
- Cheida, C.C.; Guimarães, F.H.; Beisiegel, B.D.M. Avaliação Do Risco de Extinção Do Guaxinim Procyon Cancrivorus (Cuvier, 1798) No Brasil. Biodivers. Bras. 2013, 1, 283–290. [Google Scholar]
- Macfadem Juarez, K.; Marinho-filho, J. Diet, Habitat Use, and Home Ranges of Sympatric Canids in Central Brazil. J. Mammal. 2002, 83, 925–933. [Google Scholar] [CrossRef]
- Medri, Í.M.; Mourão, G.D.M.; Rodrigues, F.H.G. Ordem Cingulata. In Mamíferos do Brasil; Reis, N.R.D., Peracchi, A.L., Pedro, W.A., Lima, I.P.D., Eds.; Technical Books Editora: Londrina, Brazil, 2011; Volume 2, pp. 75–90. ISBN 978-85-906395-4-1. [Google Scholar]
- Souza, J.L.; Anacleto, T.C.S. Levantamento de Mamíferos Atropelados Na Rodovia BR-158, Estado de Mato Grosso, Brasil. In Ecologia de Estradas: Tendências e Pesquisas; Bager, A., Ed.; UFLA: Lavras, Brazil, 2012; Volume 1, pp. 207–2022. ISBN 978-85-903770-3-0. [Google Scholar]
- Bernegossi, A.M.; Borges, C.H.D.S.; Sandoval, E.D.P.; Cartes, J.L.; Cernohorska, H.; Kubickova, S.; Vozdova, M.; Caparroz, R.; González, S.; Duarte, J.M.B. Resurrection of the Genus Subulo Smith, 1827 for the Gray Brocket Deer, with Designation of a Neotype. J. Mammal. 2022, 104, 619–633. [Google Scholar] [CrossRef]
- Cherem, J.J.; Kammers, M.; Ghizoni, I.R., Jr.; Martins, A. Mamíferos de Médio e Grande Porte Atropelados Em Rodovias Do Estado de Santa Catarina, Sul Do Brasil. Biotemas 2007, 3, 81–96. [Google Scholar]
- Bueno, C.; José, P.; de Almeida, A.L. Sazonalidade de Atropelamentos e Os Padrões de Movimentos Em Mamíferos Na BR-040 (Rio de Janeiro-Juiz de Fora). Rev. Bras. De Zoociências 2010, 12, 219–226. [Google Scholar]
- Bueno, C.; Sousa, C.O.M.; Freitas, S.R. Habitat Ou Matriz: Qual é Mais Relevante Para Prever Atropelamentos de Vertebrados? Braz. J. Biol. 2015, 75, S228–S238. [Google Scholar] [CrossRef] [Green Version]
- Saranholi, B.H.; Bergel, M.M.; Ruffino, P.H.; Rodríguez-C, K.G.; Ramazzotto, L.A.; de Freitas, P.D.; Galetti, P.M., Jr. Roadkill Hotspots in a Protected Area of Cerrado in Brazil: Planning Actions to Conservation Zonas de Alto Impacto de Atropellamientos En Un Área Protegida de Cerrado (Brasil): Planeando Acciones Para La Conservación. Rev. MVZ Córdoba 2016, 21, 5441–5448. [Google Scholar] [CrossRef] [Green Version]
- Rocha, V.J.; Agular, L.M.; Silva-Pereira, J.E.; Moro-Rios, R.F.; Passos, F.C. Feeding Habits of the Crab-Eating Fox, Cerdocyon Thous (Carnivora: Canidae), in a Mosaic Area with Native and Exotic Vegetation in Southern Brazil. Rev. Bras. Zool. 2008, 25, 594–600. [Google Scholar] [CrossRef]
- Cheida, C.C.; Nakano-Oliveira, E.; Fusco-Costa, R.; Rocha-Mendes, F.; Quadros, J. Ordem Carnivora. In Mamíferos do Brasil; Reis, N.R.D., Peracchi, A.L., Pedro, W.A., Lima, I.P.D., Eds.; Technical Books Editora: Londrina, Brazil, 2011; Volume 1, pp. 235–288. [Google Scholar]
- Barros Ferraz, K.M.P.M.; de Siqueira, M.F.; Martin, P.S.; Esteves, C.F.; do Couto, H.T.Z. Assessment of Cerdocyon Thous Distribution in an Agricultural Mosaic, Southeastern Brazil. Mammalia 2010, 74, 275–280. [Google Scholar] [CrossRef] [Green Version]
- Freitas, S.R.D.; Oliveira, A.N.D.; Ciocheti, G.; Vieira, M.V.; Matos, D.M.D.S. How Landscape Features Influence Road-Kill of Three Species of Mammals in the Brazilian Savanna? Oecologia Aust. 2015, 18, 35–45. [Google Scholar] [CrossRef]
- Courtenay, O.; Maffei, L. Crab-Eating Fox (Cerdocyon Thous). In Canids: Foxes, Wolves, Jackals and Dogs. Status Survey and Conservation Action Plan; Sillero-Zubiri, C., Hoffmann, M., Macdonald, D.W., Eds.; IUCN Publications: Gland, Switzerland; Cambridge, UK, 2004; pp. 32–38. ISBN 2-8317-0786-2. [Google Scholar]
- Ascensão, F.; Desbiez, A.L.J.; Medici, E.P.; Bager, A. Spatial Patterns of Road Mortality of Medium-Large Mammals in Mato Grosso Do Sul, Brazil. Wildl. Res. 2017, 44, 135–146. [Google Scholar] [CrossRef]
- Rocha, V.J.; Reis, N.R.D.; Sekiama, M.L. Dieta e Dispersão de Sementes Por Cerdocyon Thous (Linnaeus) (Carnívora, Canidae), Em Um Fragmento Florestal No Paraná, Brasil. Rev. Bras. Zool. 2004, 21, 871–876. [Google Scholar] [CrossRef] [Green Version]
- Fritzell, E.K. Reproduction of Raccoons (Procyon lotor) in North Dakota. Am. Midl. Nat. 1978, 100, 253–256. [Google Scholar] [CrossRef]
- Sanderson, G.C.; Nalbandov, A.V. The Reproductive Cycle of the Raccoon in Illinois. Ill. Nat. Hist. Surv. Bull. 1973, 31, 29–85. [Google Scholar] [CrossRef]
- Stope, M.B. The Raccoon (Procyon lotor) as a Neozoon in Europe. Animals 2023, 13, 273. [Google Scholar] [CrossRef] [PubMed]
- Marques, R.V.; Fábian, M.E. Daily Activity Patterns of Medium and Large Neotropical Mammals during Different Seasons in an Area of High Altitude Atlantic Rain Forest in the South of Brazil. Rev. Bras. De Zoociências 2018, 19, 38–64. [Google Scholar] [CrossRef] [Green Version]
- Lagos, L.; Picos, J.; Valero, E. Temporal Pattern of Wild Ungulate-Related Traffic Accidents in Northwest Spain. Eur. J. Wildl. Res. 2012, 58, 661–668. [Google Scholar] [CrossRef]
- Morelle, K.; Lehaire, F.; Lejeune, P. Spatio-Temporal Patterns of Wildlife-Vehicle Collisions in a Region with a High-Density Road Network. Nat. Conserv. 2013, 5, 53–73. [Google Scholar] [CrossRef] [Green Version]
- Almeida, L.B.D.; Queirolo, D.; Beisiegel, B.D.M.; Oliveira, T.G. de Avaliação Do Estado de Conservação Do Gato-Mourisco Puma Yagouaroundi (É. Geoffroy Saint-Hilaire, 1803) No Brasil. Biodivers. Bras. 2013, 1, 99–106. [Google Scholar]
- Marinho, P.H.; Bezerra, D.; Antongiovanni, M.; Fonseca, C.R.; Venticinque, E.M. Activity Patterns of the Threatened Northern Tiger Cat Leopardus Tigrinus and Its Potential Prey in a Brazilian Dry Tropical Forest. Mamm. Biol. 2018, 89, 30–36. [Google Scholar] [CrossRef]
- Paolino, R.M.; Versiani, N.F.; Pasqualotto, N.; Rodrigues, T.F.; Krepschi, V.G.; Chiarello, A.G. Uso Da Zona de Amortecimento de Uma Unidade de Conservação de Cerrado Por Mamíferos. Biota. Neotrop. 2016, 16, 1–13. [Google Scholar] [CrossRef] [Green Version]
- Astete, S.; Marinho-Filho, J.; Kajin, M.; Penido, G.; Zimbres, B.; Sollmann, R.; Jácomo, A.T.A.; Tôrres, N.M.; Silveira, L. Forced Neighbours: Coexistence between Jaguars and Pumas in a Harsh Environment. J. Arid. Environ. 2017, 146, 27–34. [Google Scholar] [CrossRef]
- Montalvo, V.H.; Saénz-Bolanõs, C.; Alfaro, L.D.; Cruz, J.C.; Guimarães-Rodrigues, F.H.; Carrillo, E.; Sutherland, C.; Fuller, T.K. Seasonal Use of Waterholes and Pathways by Macrofauna in the Dry Forest of Costa Rica. J. Trop. Ecol. 2019, 35, 68–73. [Google Scholar] [CrossRef]
- Najafi, J.; Farashi, A.; Pasha Zanoosi, A.A.; Yadreh, R. Water Resource Selection of Large Mammals for Water Resources Planning. Eur. J. Wildl. Res. 2019, 65, 82. [Google Scholar] [CrossRef]
- Massara, L.R.; Paschoal, A.M.D.O.; Hirsch, A.; Chiarello, A.G. Diet and Habitat Use by Maned Wolf Outside Protected Areas in Eastern Brazil. Trop. Conserv. Sci. 2012, 5, 284–300. [Google Scholar] [CrossRef]
- Costa, R.R.G.F.; Dias, L.A. Mortalidade de Vertebrados Por Atropelamento Em Trecho Da GO-164, No Sudoeste Goiano. Rev. De Biotecnol. Ciência 2013, 2, 58–74. [Google Scholar]
- Evangelho Silva, D.; Liberato Costa Corrêa, L.; Vilges de Oliveira, S.; Helena Cappellari, L. Monitoramento de Vertebrados Atropelados Em Dois Trechos de Rodovias Na Região Central Do Rio Grande Do Sul, Brasil. Rev. De Ciências Ambient. 2013, 7, 26–36. [Google Scholar]
- Glista, D.J.; DeVault, T.L.; DeWoody, J.A. A Review of Mitigation Measures for Reducing Wildlife Mortality on Roadways. Landsc Urban Plan 2009, 91, 1–7. [Google Scholar] [CrossRef] [Green Version]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| 2014 | 2015 | 2016 | 2017 | |||||
|---|---|---|---|---|---|---|---|---|
| Landscape Variable | Rainy (km2) | Dry (km2) | Rainy (km2) | Dry (km2) | Rainy (km2) | Dry (km2) | Rainy (km2) | Dry (km2) |
| Body of water | 50.57 | 23.22 | 39.20 | 14.13 | 22.37 | 12.36 | 21.63 | 9.38 |
| Wetland | 43.25 | 63.35 | 74.70 | 91.22 | 45.53 | 100.28 | 39.86 | 47.43 |
| Urban infrastructure | 16.58 | 16.59 | 16.89 | 16.90 | 17.44 | 17.06 | 19.45 | 21.19 |
| Farming | 417.89 | 422.79 | 453.24 | 457.32 | 449.68 | 452.26 | 454.02 | 456.74 |
| Exposed soil | 25.78 | 34.66 | 42.62 | 57.08 | 29.75 | 44.11 | 18.96 | 163.78 |
| Dense vegetation | 617.71 | 449.13 | 598.86 | 513.68 | 531.10 | 336.29 | 563.01 | 577.86 |
| Sparse vegetation | 1216.10 | 1378.16 | 1162.39 | 1237.56 | 1292.01 | 1425.71 | 1270.92 | 1111.47 |
| Landscape variable | Rainy (%) | Dry (%) | Rainy (%) | Dry (%) | Rainy (%) | Dry (%) | Rainy (%) | Dry (%) |
| Body of water | 2.12 | 0.97 | 1.64 | 0.59 | 0.94 | 0.52 | 0.91 | 0.39 |
| Wetland | 1.81 | 2.65 | 3.13 | 3.82 | 1.91 | 4.20 | 1.67 | 1.99 |
| Urban infrastructure | 0.69 | 0.69 | 0.71 | 0.71 | 0.73 | 0.71 | 0.81 | 0.89 |
| Farming | 17.50 | 17.71 | 18.98 | 19.15 | 18.83 | 18.94 | 19.01 | 19.13 |
| Exposed soil | 1.08 | 1.45 | 1.78 | 2.39 | 1.25 | 1.85 | 0.79 | 6.86 |
| Dense vegetation | 25.87 | 18.81 | 25.08 | 21.51 | 22.24 | 14.08 | 23.58 | 24.20 |
| Sparse vegetation | 50.93 | 57.71 | 48.68 | 51.83 | 54.11 | 59.70 | 53.22 | 46.55 |
| Kappa | 0.80 | 0.81 | 0.81 | 0.79 | 0.80 | 0.79 | 0.78 | 0.81 |
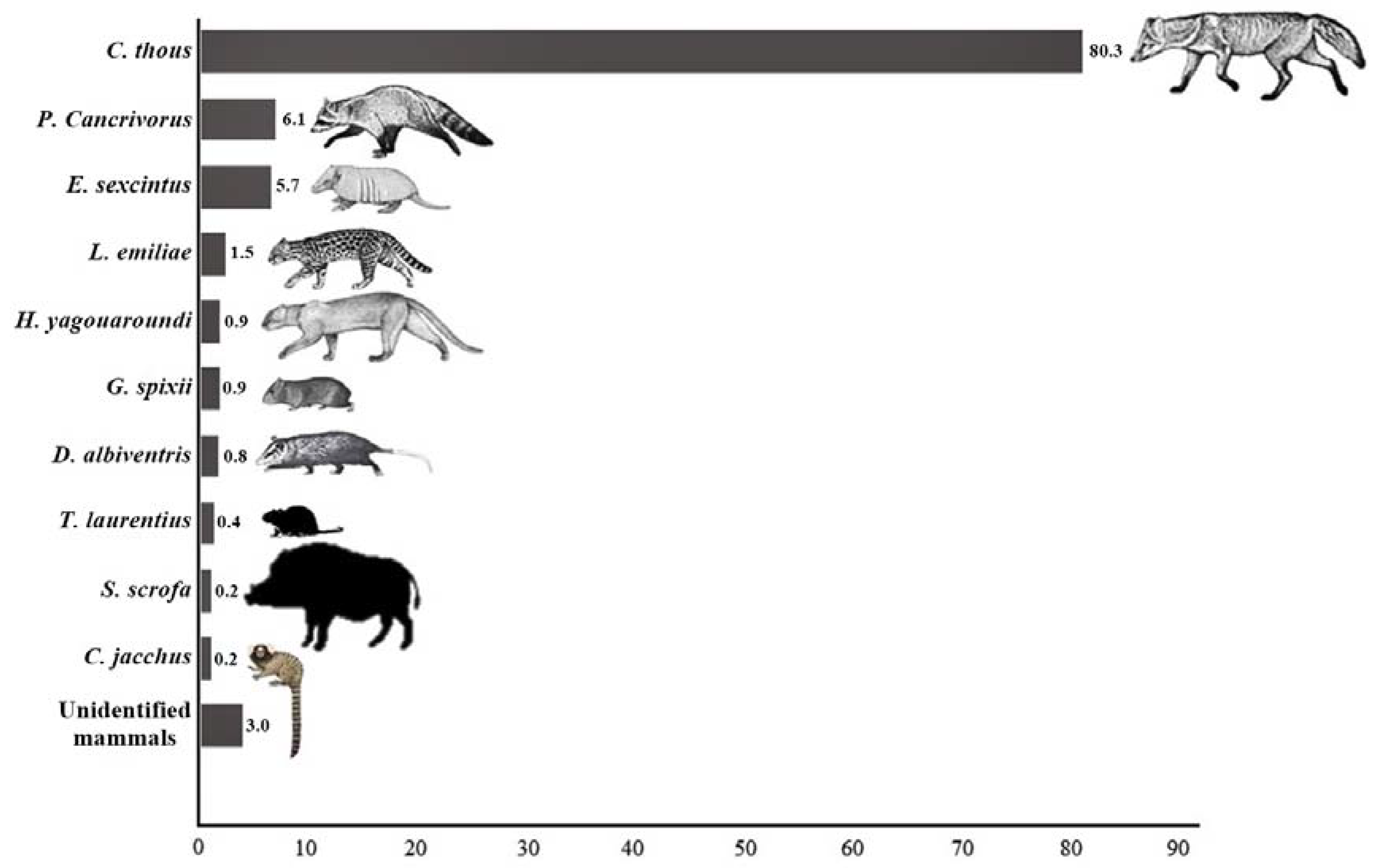
| Taxon | Common Name | Number of Roadkills | Roadkill Rates (ind/km/Day) | Status (Brazil/IUCN) | |||||
|---|---|---|---|---|---|---|---|---|---|
| 2013 | 2014 | 2015 | 2016 | 2017 | Mean | ||||
| CARNIVORA | |||||||||
| Canidae | |||||||||
| Cerdocyon thous (Linnaeus, 1766) | Crab-eating fox | 423 | 0.068 | 0.061 | 0.085 | 0.053 | 0.073 | 0.068 | LC/LC |
| Felidae | |||||||||
| Leopardus emiliae (Thomas, 1914) | Northern tiger cat | 8 | 0 | 0.002 | 0.001 | 0.001 | 0.002 | 0.001 | EN/VU |
| Herpailurus yagouaroundi (É, Geoffroy Saint-Hilare, 1803) | Jaguarundi | 5 | 0.003 | 0.001 | 0.001 | 0.001 | 0 | 0.001 | VU/LC |
| Procyonidae | |||||||||
| Procyon cancrivorus (G, [Baron] Cuvier, 1798) | Crab-eating raccoon | 32 | 0.003 | 0.005 | 0.007 | 0.003 | 0.006 | 0.005 | LC/LC |
| CINGULATA | |||||||||
| Dasypodidae | |||||||||
| Euphractus sexcinctus (Linnaeus, 1758) | Six-banded armadillos | 30 | 0 | 0.003 | 0.003 | 0.013 | 0.003 | 0.005 | LC/LC |
| DIDELPHIMORPHIA | |||||||||
| Didelphidae | |||||||||
| Didelphis albiventris (Lund, 1840) | White-eared opossum | 4 | 0 | 0.002 | 0.001 | 0 | 0 | 0.001 | LC/LC |
| ARTIODACTYLA | |||||||||
| Sus scrofa scrofa (Linnaeus, 1758) | Wild boar | 1 | 0 | 0.001 | 0 | 0 | 0 | 0.000 | LC/LC |
| RODENTIA | |||||||||
| Caviidae | |||||||||
| Galea spixii (Wagler, 1831) | Yellow-toothed cavy | 5 | 0 | 0.002 | 0.001 | 0 | 0.001 | 0.001 | LC/LC |
| Echimyidae | |||||||||
| Thrichomys laurentius (Thomas, 1904) | São Lourenço’s punaré | 2 | 0.003 | 0 | 0 | 0.001 | 0 | 0.000 | LC/LC |
| PRIMATES | |||||||||
| Callitrichidae | |||||||||
| Callithrix jacchus (Linnaeus, 1758) | Common marmoset | 1 | 0 | 0.001 | 0 | 0 | 0 | 0.000 | LC/LC |
| Unidentified Mammal | 16 | 0 | 0.001 | 0.001 | 0.009 | 0.002 | 0.003 | ||
| TOTAL | 527 | 0.077 | 0.076 | 0.099 | 0.080 | 0.088 | 0.085 | ||
| Roadkill | Buffer | Variables Selected in the Model | Β (Standard Error) | p | df | ΔAICc | wAICc |
|---|---|---|---|---|---|---|---|
| All mammals (Except C. thous) | 5 km | Dense vegetation | 2.77 × 10−2 (6.30 × 10−3) | 1.08 × 10−5 * | 9 | 0.0 | 0.9984 |
| Urban infrastructure | 7.87 × 10−2 (4.14 × 10−2) | 5.7 × 10−2 | |||||
| Interaction | −2.36 × 10−2 (4.79 × 10−3) | 8.35 × 10−7 * | |||||
| Period (dry) | −9.51 × 10−1 (1.30 × 10−1) | 3.14 × 10−13 * | |||||
| Protected areas | 6.39 × 10−5 (1.56 × 10−5) | 4.21 × 10−5 * | |||||
| C. thous | 1 km | Dense vegetation | 4.20 × 10−3 (2.23 × 10−3) | 5.9 × 10−2 | 9 | 0.0 | 0.7813 |
| Urban infrastructure | −1.66 × 10−2 (6.77 × 10−3) | 1.40 × 10−2 * | |||||
| Interaction | −6.88 × 10−3 (1.78 × 10−3) | 1.14 × 10−4 * | |||||
| Period (dry) | 8.30 × 10−2 (5.91 × 10−2) | 1.60 × 10−1 | |||||
| Protected areas | −3.96 × 10−6 (9.38 × 10−6) | 6.73 × 10−1 | |||||
| P. cancrivorus | 1 km | Exposed soil | 1.55 × 10−1 (3.00 × 10−2) | 2.28 × 10−7 * | 8 | 0.0 | 0.5792 |
| Urban infrastructure | −1.12 (8.05 × 10−1) | 1.64 × 10−1 | |||||
| Period (dry) | −1.36 (2.65 × 10−1) | 2.79 × 10−7 * | |||||
| Protected areas | 1.19 × 10−5 (2.33 × 10−5) | 6.09 × 10−1 | |||||
| Exposed soil | 1.59 × 10−1 (3.01 × 10−2) | 1.23 × 10−7 * | 9 | 1.2 | 0.3166 | ||
| Urban infrastructure | −2.92 × 10−1 (1.07) | 7.85 × 10−1 | |||||
| Interaction | −2.76 × 10−1 (3.90 × 10−1) | 4.80 × 10−1 | |||||
| Period (dry) | −1.36 (2.65 × 10−1) | 2.67 × 10−7 * | |||||
| Protected areas | 1.17 × 10−5 (2.33 × 10−5) | 6.14 × 10−1 | |||||
| E. sexcinctus | 1 km | Sparse vegetation | 2.09 × 10−2 (6.16 × 10−3) | 6.98 × 10−4 * | 8 | 0.0 | 0.5092 |
| Urban infrastructure | −6.24 × 10−1 (5.21 × 10−1) | 2.30 × 10−1 | |||||
| Period (dry) | −1.59 (2.65 × 10−1) | 2.20 × 10−9 * | |||||
| Protected areas | 2.53 × 10−5 (2.04 × 10−5) | 2.16 × 10−1 | |||||
| Sparse vegetation | 1.93 × 10−2 (6.24 × 10−3) | 2 × 10−3 * | 9 | 0.4 | 0.4200 | ||
| Urban infrastructure | −2.95 (2.86) | 3.01 × 10−1 | |||||
| Interaction | 4.44 × 10−2 (4.41 × 10−2) | 3.14 × 10−1 | |||||
| Period (dry) | −1.60 (2.66 × 10−1) | 1.70 × 10−9 * | |||||
| Protected areas | 2.57 × 10−5 (2.03 × 10−5) | 2.06 × 10−1 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Santos, R.; Shimabukuro, A.; Taili, I.; Muriel, R.; Lupinetti-Cunha, A.; Freitas, S.R.; Calabuig, C. Mammalian Roadkill in a Semi-Arid Region of Brazil: Species, Landscape Patterns, Seasonality, and Hotspots. Diversity 2023, 15, 780. https://doi.org/10.3390/d15060780
Santos R, Shimabukuro A, Taili I, Muriel R, Lupinetti-Cunha A, Freitas SR, Calabuig C. Mammalian Roadkill in a Semi-Arid Region of Brazil: Species, Landscape Patterns, Seasonality, and Hotspots. Diversity. 2023; 15(6):780. https://doi.org/10.3390/d15060780
Chicago/Turabian StyleSantos, Raul, Ayko Shimabukuro, Itainara Taili, Roberto Muriel, Artur Lupinetti-Cunha, Simone Rodrigues Freitas, and Cecilia Calabuig. 2023. "Mammalian Roadkill in a Semi-Arid Region of Brazil: Species, Landscape Patterns, Seasonality, and Hotspots" Diversity 15, no. 6: 780. https://doi.org/10.3390/d15060780