
Suitability of Wheel Ruts: Characterization of Overlooked Breeding Habitats of Alpine Newts
Department of Zoology and Fisheries, Faculty of Agrobiology, Food and Natural Resources, Czech University of Life Sciences Prague, 16521 Prague, Czech Republic
Diversity 2023, 15(6), 762; https://doi.org/10.3390/d15060762
Submission received: 20 May 2023
/
Revised: 5 June 2023
/
Accepted: 6 June 2023
/
Published: 9 June 2023
(This article belongs to the Special Issue Ecology and Conservation of Amphibians)
Abstract
:The Alpine newt (Ichthyosaura alpestris) is sensitive to habitat destruction and alteration, and as a result there has been a decline in their populations across Europe. Due to historical landscape changes, the populations of Alpine newts drawn from lowlands and hilly areas have recently been primarily found in artificial habitats, such as wheel ruts. In some areas inhabited by the Alpine newt range, wheel ruts are the only available habitat for newt breeding, and thus the only one allowing the maintenance of their populations. However, our knowledge about this type of habitat is limited. Therefore, 60 wheel ruts in a hilly forested area in the Czech Republic were checked for the presence of Alpine newt larvae. Their characteristics were measured (conductivity, pH, dissolved oxygen, depth, water coverage, plants, presence of prey, hydroperiod). The presence of larvae was positively associated with higher values of pH, coverage of water plants, and the presence of prey. The strongest predictor was the hydrological stability of wheel ruts, and newts were found to prefer non-drying ruts. From the conservation point of view, this means that the suitability of wheel ruts for Alpine newts can be estimated quite easily. This can essentially be performed on the basis of visual inspection if it is carried out through the breeding season of newts.
1. Introduction
Amphibians are decreasing in abundance worldwide [1,2]. This is true for Caudata amphibians [3] and also for Alpine newts (Ichthyosaura alpestris). The Alpine newt species is rated as being at the level of least concern in most of its range. However, a continuously decreasing trend is observed for its populations across Europe [4].
The Alpine newt species is widely distributed across western and central Europe from lowlands to mountains (up to 2500 m a.s.l.) [4,5]. It is very adaptable and usually breeds in lentic habitats of natural or anthropogenic origin that vary greatly in their characteristics [4,5,6]. Nevertheless, most of our knowledge about the ecology and ethology of Alpine newts comes from laboratory studies [7,8,9] or from mountain lake localities [10,11,12]. Due to historical landscape changes, populations of Alpine newts from lowlands and hilly areas have recently been primarily found in artificial habitats such as agricultural ponds, abandoned quarries, garden pools, or even small ruts caused by automobile traffic on unpaved roads [5,13]. These wheel ruts are extreme habitats. This is primarily due to the water small volume and resulting instability with sudden fluctuations in water temperature or turbidity [14,15,16]. In some areas of the Alpine newt range, these are the only available habitats for their breeding and thus allow for the maintenance of their populations [13,17,18]. Our knowledge about this extreme habitat is limited despite the considerable importance of wheel ruts for Alpine newt populations.
Because potential conservation actions focused on amphibians must be based on an understanding of population functioning and their reactions and adaptations to their environment [19], the aim of this study was to determine the primary characteristics of wheel ruts that affect the presence, reproduction, and development of Alpine newts.
2. Materials and Methods
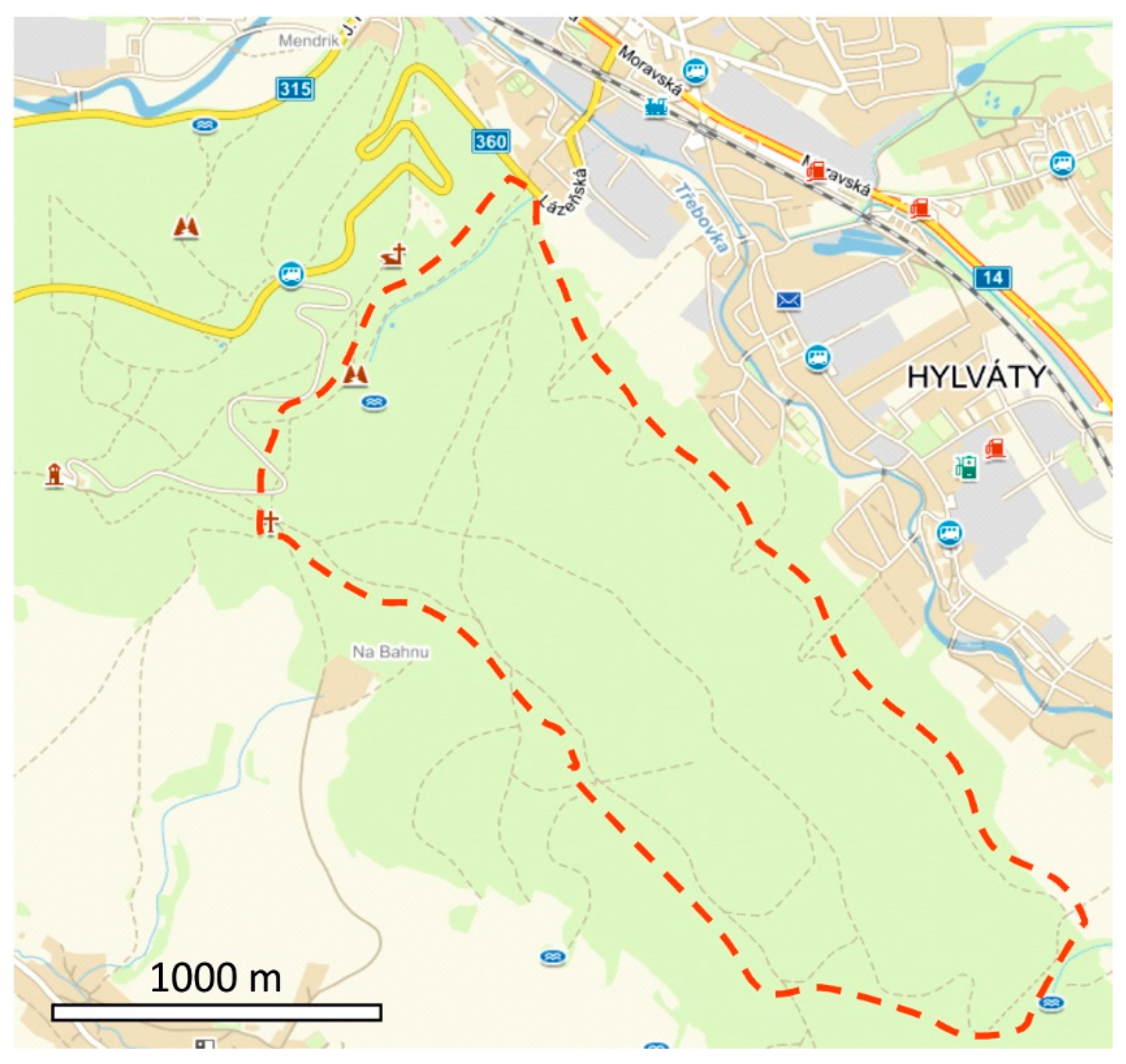
Data about wheel ruts and their occupancy by Alpine newts were collected in Kozlovecký hřbet (GPS: 49.954 N, 16.397 E) on an area of about 108 ha. This hilly area, with a dense network of forest roads, is situated in the west and the southwest direction of the Hylváty settlement and lies in the Czech Republic (Figure 1). It is wholly covered by a spruce forest of different ages, which is used as a source of timber. Water bodies, namely 60 wheel ruts over an altitude range of 393–548 m with possible use by amphibians for breeding, were surveyed (Figure 2). The only reservoirs suitable for amphibian breeding are wheel ruts in this area. The streams here are periodic, have high slopes, and thus dry up very quickly. In addition to the Alpine newt, also several individuals of common frog families (Rana temporaria) were found in the area. However, the reproduction of this species was not recorded during the study period.
From 12 March till 15 October 2011, the characteristics of each wheel rut were measured (some of these characteristics repeatedly) and the presence of Alpine newts was checked. This period corresponds with the breeding and subsequent larval development of Alpine newts in the study area [15,20]. Because the presence of adult newts can be transient, the suitability of wheel ruts for newts was based on the occurrence of larvae. The Alpine newt was the only amphibian species that reproduced in the studied area.
The depth of the ruts (in cm) and the temperature of the water inside them were measured once per month. In the following analysis, the highest depth value was used. The temperature was measured at positions 10 cm under the water surface using the thermometer DETR3 Voltcgraft. Due to low variation among wheel ruts in terms of the overall minimal and maximal temperature value of the difference between min. and max. temperature for each, the wheel rut was used as a factor temperature. Conductivity, pH, and dissolved oxygen were detected once per season on the 13 of August using Greisinger’s GmbH electronic device (GMH 3430, GMH 3530, GMH 3610). Data collection was always conducted between 10.00 and 15.00 h.
The presence of water plants was determined in percent (%) as an estimated portion of the covered bottom based on photographs taken each month. Values were taken from the photo, with the highest coverage being taken for analysis. In the warmest part of the year, some wheel ruts desiccate, therefore I evaluated the hydroperiod of wheel ruts as being stable or unstable. During the whole observation period, I looked for the presence of potential prey and especially searched for earthworms (Lumbricidae), mosquito larvae (Culicidae), and zooplankton (Cladocera, Cyclopoida), which were found in some wheel ruts by a previous study [20]. The presence of prey was confirmed if the prey was found in the wheel rut at least once during the study period.
Then, independent continuous variables were recorded: temperature (difference between min.–max. temperature, in °C), depth (highest value from all measurements of max. depth, in cm), coverage of water plants (highest value from all measurements, in %), conductivity (μS/cm), dissolved oxygen (mg/L) and pH. The independent binary variables were hydroperiod (stable/unstable) and the presence of prey (present/absent). As a dependent variable presence (yes/no) of Alpine newt, larvae from a particular rut were used.
Before the application of the primary model, I checked for partial correlations among all continuous independent variables. Conductivity was in a positive relationship with pH (Pearson correlation coefficient, r = 0.77, p < 0.001) and a negative relationship with dissolved oxygen (Pearson correlation coefficient, r = −0.39, p < 0.01). Therefore, factor conductivity was not used in subsequent analysis. Other correlations were insignificant. The independence of the binary variables, i.e., hydroperiod and presence of prey, was checked by 2 × 2 contingency tables and these were not connected (Yates-corrected χ2 = 0.97, p = 0.326).
I used a generalized linear model with binomial distribution to test the characteristics of wheel ruts that contained larval Alpine newts. The starting maximal model contained all factors and their two-way interactions. In search for the minimal adequate model, I followed the general recommendation for model simplification [21,22] and used function ANOVA with chi-test analysis for comparison of models. Statistical significance was established using α = 0.05. All tests were computed using R statistical software, version 4.1.2 [23].
3. Results
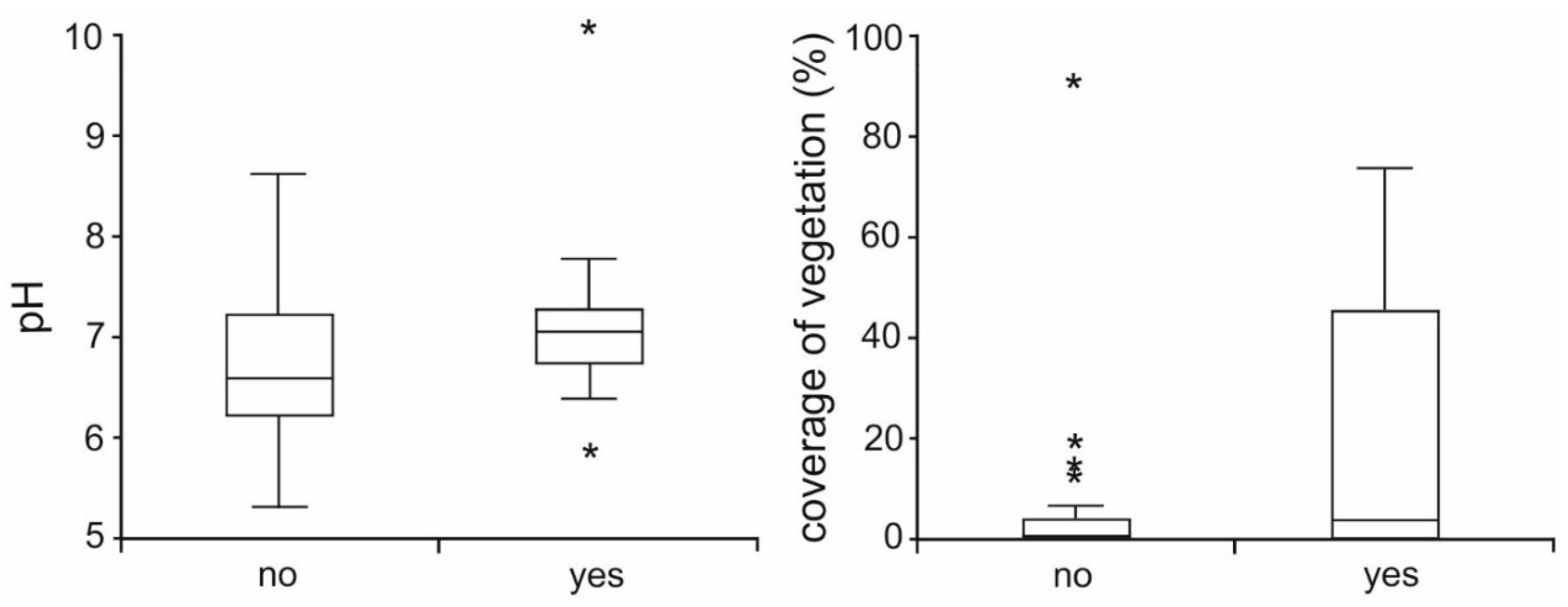
Larval newts were present in 20% of mapped wheel ruts. The minimal adequate model, which explains the presence of newt larvae in wheel ruts, contains pH, coverage of water plants, hydroperiod, and presence of prey as significant factors (Table 1). Alpine newts try to avoid reproducing in drying ruts (i.e., in those with unstable hydroperiod); however, these constitute the majority (68%) of monitored wheel ruts in the studied area. The presence of prey was confirmed at 73% of studied wheel ruts and this factor had a positive effect on the occurrence of larval newts. Both the higher coverage of water plants and higher pH had a positive effect on the presence of larval newts (Figure 3).
4. Discussion
From studied factors, the strongest predictor of Alpine newt larvae absence in wheel ruts was found to be unstable hydroperiod, i.e., if the wheel rut dries out at least once. Hydroperiod is one of the primary factors influencing amphibian species richness in a particular locality and this factor affects the reproduction success of a species [17,24]. Temporary ponds or pools are risky for larval development as the early drying of a rut during the season terminates the egg and larval stages of newts [25]. On the other hand, due to their short hydroperiod, these unstable water bodies host limited numbers of predator species, which is their primary advantage for newts. This is especially true for fishes, which are recognized as the largest threat to the successful reproduction of European newts, and the presence of fish is a strong predictor of the absence or future absence of newts in particular water habitats [26,27,28]. Drying out excludes fishes and some invertebrate predators from these habitats [28,29]. In our study, ruts with stable hydroperiods did not differ from those with unstable hydroperiods in terms of variety and presence of predators. Wheel ruts were too small to host fish and dragonfly larvae were found in both types of ruts. Except for the fish presence of high sludge depth (over 15 cm) on the bottom of the water, habitat was recently recognized as another factor that strongly predicted Alpine newt absence [28]. This factor was neglected at times in our study, but the bottom of the ruts was always clay muddy with a depth of layers up to 5 cm.
For Alpine newts, habitat type (lake, pond, pool, etc.) is one of the strongest predictors of their presence in a particular locality [6]. In this study, all-wheel ruts in the area had the same origin (water-filled vehicle tracks) and the means of their saturation was similar. The highest water levels were in wheel ruts during spring (from the end of March to early April), when wheel ruts were filled from melting snow. During the rest of the season water levels were dependent particularly on rainfall. Desiccation events were most numerous in July. In contrast with the study of Mole salamander (Ambystoma talpoideum) [25], a similar number of Alpine newts and same-sex ratios were found in the studied area during seasons with different amounts of rainfall [16]. This suggests that Alpine newts were able to cope with wheel rut drying. We found that adults were able to move among wheel ruts during breeding seasons [15] and that these movements were sex-specific [16]. Adult females were able to ensure reproduction success by spreading reproduction investments among several wheel ruts. However, this behavior was not just a simple distribution of risk—during data collection for this study, we recorded only a single unstable wheel rut where drying led to the death of the larvae present. Therefore, it seems that the perspective of early wheel rut drying could be estimated by female newts quite successfully, at least in years without extreme drought.
Positive effects on Alpine newt larvae presence also had higher values of pH and coverage of water vegetation. Alpine newts are considered to be the most acid-tolerant European newt species [30] and are able to long-term prosper at pH values from ranging 5.0 to 5.6 [5]. However, Alpine newts in this study preferred reproduction wheel ruts with higher pH values. Larval newts were only found in wheel ruts with pH values of 6.0 and more. Generally, lowering pH means decreasing the number of hatched larvae in European newt species and their earlier hatching [31,32]. Earlier hatched larvae then need a longer time to reach the size and obtain the energy necessary for successful passing through metamorphosis [30].
As proven in earlier studies, aquatic plants in breeding habitats positively affect the presence and abundance of European newt species [26,33]. Submerged vegetation is of primary importance [34]. Dense vegetation enables newts to survive in areas with predatory fish [5]. Submerged vegetation is also crucial for the development of newts. Wrapping eggs individually into the leaves of aquatic plants protects them from predators [7,35] and from UV radiation that has a teratogenic effect on embryos [36]. Up to 90% [7] or 96% [35] of eggs are deposited by females on submerged leaves, and the remainder are placed on the bottom substrate. However, a female’s willingness to lay eggs in tanks without vegetation is significantly lower [37].
The presence of water vegetation is more important for egg-laying females of Alpine newts than water temperature. The optimal temperature for egg laying is 17.5 °C (with a range of 12.5–22.5 °C) for Alpine newts. However, females tend to find a place with submerged vegetation even if they are in the water with optimal temperature [37]. In deeper water habitats larval Alpine newts try to avoid the top water layer below the surface (0–15 cm) and, from the studied species of newts, seem to be most benthic [11]. In the studied area, most of the wheel ruts were not deeper than 15 cm; therefore, larval newts had no option for dealing with water temperature in the water column. The presence of larvae is affected by the decision of adult newts to breed or not in a particular rut. Nevertheless, even significant differences (15.5 °C in seasonal maximum) among particular wheel rut temperatures had no effect on the occurrence of larvae.
Human-induced habitat loss and degradation are the most prominent present stressors for Caudata amphibians [38]. However, sometimes human activity can inadvertently create and maintain new breeding sites, such as wheel ruts. Despite the extremity of such an environment, wheel ruts form specifically suited breeding habitat in European forests [14,17] that are often the only sites available. For this reason, they should not be overlooked. From the point of view of practical nature conservation conclusions of our study means that the suitability of wheel ruts for Alpine newts can be estimated quite easily. Indeed, it may be essentially conducted on the basis of visual inspection if it is carried out through the breeding season of newts (except for pH, the other three factors that significantly influence the presence of larval Alpine news can be estimated by eye).
Funding
This research received no external funding.
Institutional Review Board Statement
Not applicable.
Data Availability Statement
The datasets generated and/or analyzed during the current study are available from the authors upon request.
Acknowledgments
The author thanks Martina Kubečková for her help, patience, and endurance during field data collection.
Conflicts of Interest
The author declares no conflict of interest.
References
- Stuart, S.N.; Chanson, J.S.; Cox, N.A.; Young, B.E.; Rodrigues, A.S.L.; Fischman, D.L.; Waller, R.W. Status and trends of amphibian declines and extinctions worldwide. Science 2004, 306, 1783–1786. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Grant, E.H.C.; Miller, D.A.W.; Muths, E. A synthesis of evidence of drivers of amphibian declines. Herpetologica 2020, 76, 101–107. [Google Scholar] [CrossRef]
- Martel, A.; Blooi, M.; Adriaensen, C.; Van Rooij, P.; Beukema, W.; Fisher, M.C.; Farrer, R.A.; Schmidt, B.R.; Tobler, U.; Goka, K.; et al. Recent introduction of a chytrid fungus endangers Western Palearctic salamanders. Science 2014, 346, 630–631. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Arntzen, J.W.; Denoël, M.; Kuzmin, S.; Ishchenko, V.; Beja, P.; Andreone, F.; Jehle, R.; Nyström, P.; Miaud, C.; Anthony, B.; et al. Ichthyosaura alpestris. The IUCN Red List of Threatened Species. 2009. Available online: www.iucnredlist.org/species/59472/89702213 (accessed on 14 January 2023).
- Roček, Z.; Joly, P.; Grossenbacher, K. 2003: Bergmolch–Triturus alpestris (Laurenti, 1768). In Handbuch der Reptilien und Amphibian Europas; Böhme, W., Ed.; Aula Verlag: Wiebelsheim, Germany, 2003; pp. 607–656. [Google Scholar]
- Cirovic, R.; Radovi, D.; Vukov, T.D. Breeding site traits of European newts (Triturus macedonicus, Lissotriton vulgaris, and Mesotriton alpestris: Salamndridae) in the Montenegrin Karst region. Arch. Biol. Sci. Belgrade 2008, 60, 459–468. [Google Scholar] [CrossRef]
- Miaud, C. Identification of newt (Triturus alpestris and T. helveticus) eggs predators and protective role of oviposition behaviour. J. Zool. 1993, 231, 575–582. [Google Scholar] [CrossRef]
- Denoël, M. Effect of rival males on the courtship of paedomorphic and metamorphic Triturus alpestris (Amphibia: Salamandridae). Copeia 2003, 3, 618–623. [Google Scholar] [CrossRef] [Green Version]
- Denoël, M.; Mathieu, M.; Poncin, P. Effect of water temperature on the courtship behavior of the alpine newt Triturus alpestris. Behav. Ecol. Sociobiol. 2005, 58, 121–127. [Google Scholar] [CrossRef]
- Miaud, C.; Guyetant, R.; Faber, H. Age, size, and growth of the alpine newt, Triturus alpestris (Urodela: Salamandridae), at high altitude and a review of life-history trait variation throughout its range. Herpetologica 2000, 56, 135–144. [Google Scholar]
- Vogrin, M. Micro-habitat use within a guild of newt larvae (Trituridae) in an Alpine lake. Biol. Bratisl. 2006, 61, 579–584. [Google Scholar] [CrossRef]
- Denoël, M.; Schabetsberger, R.; Joly, P. Trophic specialisations in alternative heterochronic morphs. Naturwissenschaften 2004, 91, 81–84. [Google Scholar]
- Denoël, M. Amphibiens et Reptiles de Wallonie, Le Triton alpestre, Tri Tur Us Alpestris (Laurenti, 1768); Aves-Raînne & Région Wallonne: Namur, Belgium, 2007; pp. 62–71. [Google Scholar]
- Joger, U. Wassergefullte Wangenspuren Auf Forstwegen: Synokologische Untersuchungen an Einem Kurzlebigen Okosystem; Chimaira: Frankfurt am Main, Germany, 2000. [Google Scholar]
- Kopecký, O.; Vojar, J.; Denoël, M. Movements of Alpine newts (Mesotriton alpestris) between small aquatic habitats (ruts) during the breeding season. Amphibia-Reptilia 2010, 31, 109–116. [Google Scholar] [CrossRef] [Green Version]
- Kopecký, O.; Vojar, J.; Denoël, M. Sex-specific effect of pool desiccation on the movement of Alpine Newts, Mesotriton alpestris (Laurenti, 1768), among breeding sites. Herpetozoa 2012, 24, 127–134. [Google Scholar]
- Babik, W.; Rafinski, J. Amphibian breeding site characteristics in the Western Carpathians, Poland. Herpetol. J. 2001, 11, 41–51. [Google Scholar]
- Denoël, M.; Demars, B. The benefits of heterospecific oophagy in a top predator. Acta Oecol. 2008, 34, 74–79. [Google Scholar] [CrossRef]
- Semlitsch, R.D. Critical elements for biologically based recovery plans of aquatic-breeding Amphibians. Conserv. Biol. 2002, 16, 619–629. [Google Scholar] [CrossRef] [Green Version]
- Kopecký, O.; Vojar, J. Které vlastnosti vodních biotopů ovlivňují početnost čolků horských (Mesotriton alpestris) za nestálých podmínek prostředí. Sborník Sev. Muz. Lib. 2007, 25, 131–139. [Google Scholar]
- Crawley, M.J. The R Book; John Wiley and Sons: Chichester, UK, 2007. [Google Scholar]
- Pekár, S.; Brabec, M. Moderní Analýza Biologických Dat 1; Scientia: Brno, Czech Republic, 2009. [Google Scholar]
- R Development Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2021. [Google Scholar]
- Baber, M.J.; Fleishman, E.; Babbitt, K.J.; Tarr, T.L. The relationship between wetland hydroperiod and nestedness patterns in assemblages of larval amphibians and predatory macroinvertebrates. Oikos 2004, 107, 16–27. [Google Scholar] [CrossRef]
- Semlitsch, R.D. Density-dependent growth and fecundity in the paedomorphic salamander Ambystoma talpoideum. Ecology 1987, 68, 1003–1008. [Google Scholar] [CrossRef]
- Denoël, M.; Lehmann, A. Multi-scale effect of landscape processes and habitat quality on newt abundance: Implications for conservation. Biol. Conserv. 2006, 130, 495–504. [Google Scholar] [CrossRef] [Green Version]
- Hartel, T.; Nemes, S.; Cogalniceanu, D.; Ollerer, K.; Schweiger, O.; Moga, C.-I.; Demeter, L. The effect of fish and aquatic habitat complexity on amphibians. Hydrobiologia 2007, 583, 173–178. [Google Scholar] [CrossRef]
- DeTroyer, N.; Forio, M.A.E.; Roels, K.; DeMeester, L.; Lemmens, P.; Declerck, S.A.J.; Martens, K.; Goethals, P. Key management rules for agricultural newt breeding ponds based on habitat suitability models. Glob. Ecol. Conserv. 2020, 23, e01086. [Google Scholar] [CrossRef]
- Wellborn, G.A.; Skelly, D.K.; Werner, E.E. Mechanisms creating community structure across a freshwater habitat gradient. Annu. Rev. Ecol. Syst. 1996, 27, 337–363. [Google Scholar] [CrossRef] [Green Version]
- D’Amen, M.; Vignoli, L.; Bologna, M.A. The effects of temperature and pH on the embryonic development of two species of Triturus (Caudata: Salamandridae). Amphib. -Reptil. 2007, 28, 295–300. [Google Scholar]
- Griffiths, R.A.; DeWijer, P.; Brady, L. The effect of pH on embryonic and larval development in smooth and palmate newts, Triturus vulgaris and Triturus helveticus. J. Zool. 1993, 230, 401–409. [Google Scholar] [CrossRef]
- Griffiths, R.A.; DeWijer, P. Differential effects of pH and temperature on embryonic development in the British newts (Triturus). J. Zool. 1994, 234, 613–622. [Google Scholar] [CrossRef]
- Strijbosch, H. Habitat selection of amphibians during their aquatic phase. Oikos 1979, 33, 363–372. [Google Scholar] [CrossRef]
- Pavignano, I.; Giacoma, C.; Castellano, S. A multivariate analysis of amphibian habitat determinants in north western Italy. Amphibia-Reptilia 1990, 11, 311–324. [Google Scholar] [CrossRef]
- Orizaola, G.; Braňa, F. Oviposition behaviour and vulnerability of eggs to predation in four newt species (genus Triturus). Herpetol. J. 2003, 13, 121–124. [Google Scholar]
- Marco, A.; Lizana, M.; Alvarez, A.; Blaustein, A.R. Egg-wrapping behaviour protects newt embryos from UV radiation. Anim. Behav. 2001, 61, 639–644. [Google Scholar] [CrossRef] [Green Version]
- Dvořák, J.; Gvoždík, L. Oviposition preferences in newts: Does temperature matter? Ethology 2009, 115, 533–539. [Google Scholar] [CrossRef]
- Hof, C.; Araújo, M.B.; Jetz, W.; Rahbek, C. Additive threats from pathogens, climate and land-use change for global amphibian diversity. Nature 2011, 480, 516–519. [Google Scholar] [CrossRef]
Figure 1.
Map of the study area (spruce forest in a hilly area near Hylváty settlement, Czech Republic) delineated by red dashed line.
Figure 1.
Map of the study area (spruce forest in a hilly area near Hylváty settlement, Czech Republic) delineated by red dashed line.

Figure 2.
Example of wheel ruts in the studied area of Kozlovecký hřbet (Czech Republic) that were inhabited by larval Alpine newts (Ichthyosaura alpestris).
Figure 2.
Example of wheel ruts in the studied area of Kozlovecký hřbet (Czech Republic) that were inhabited by larval Alpine newts (Ichthyosaura alpestris).

Figure 3.
Presence or absence of larval Alpine newts in wheel ruts with relation to factors pH and percentage of coverage of the bottom of ruts by vegetation (line in the box—median; box 25–75%; whiskers—range of outliers; asterix—extremes).
Figure 3.
Presence or absence of larval Alpine newts in wheel ruts with relation to factors pH and percentage of coverage of the bottom of ruts by vegetation (line in the box—median; box 25–75%; whiskers—range of outliers; asterix—extremes).


{kind=link}
{kind=link}
{kind=link}
Table 1.
A minimal adequate model of GLM with binomial distribution examining factors influencing the occurrence of Alpine newt larvae in wheel ruts.
Table 1.
A minimal adequate model of GLM with binomial distribution examining factors influencing the occurrence of Alpine newt larvae in wheel ruts.
| df | F | p | |
|---|---|---|---|
| coverage of plants | 1 | 4.03 | 0.045 |
| pH | 1 | 4.75 | 0.029 |
| presence of prey | 1 | 6.76 | 0.009 |
| hydroperiod | 1 | 9.13 | 0.003 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the author. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Kopecký, O. Suitability of Wheel Ruts: Characterization of Overlooked Breeding Habitats of Alpine Newts. Diversity 2023, 15, 762. https://doi.org/10.3390/d15060762
AMA Style
Kopecký O. Suitability of Wheel Ruts: Characterization of Overlooked Breeding Habitats of Alpine Newts. Diversity. 2023; 15(6):762. https://doi.org/10.3390/d15060762
Chicago/Turabian StyleKopecký, Oldřich. 2023. "Suitability of Wheel Ruts: Characterization of Overlooked Breeding Habitats of Alpine Newts" Diversity 15, no. 6: 762. https://doi.org/10.3390/d15060762
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.