
What Does “ITS” Say about Hybridization in Lineages of Sarsia (Corynidae, Hydrozoa) from the White Sea?


Abstract
:
1. Introduction
2. Materials and Methods
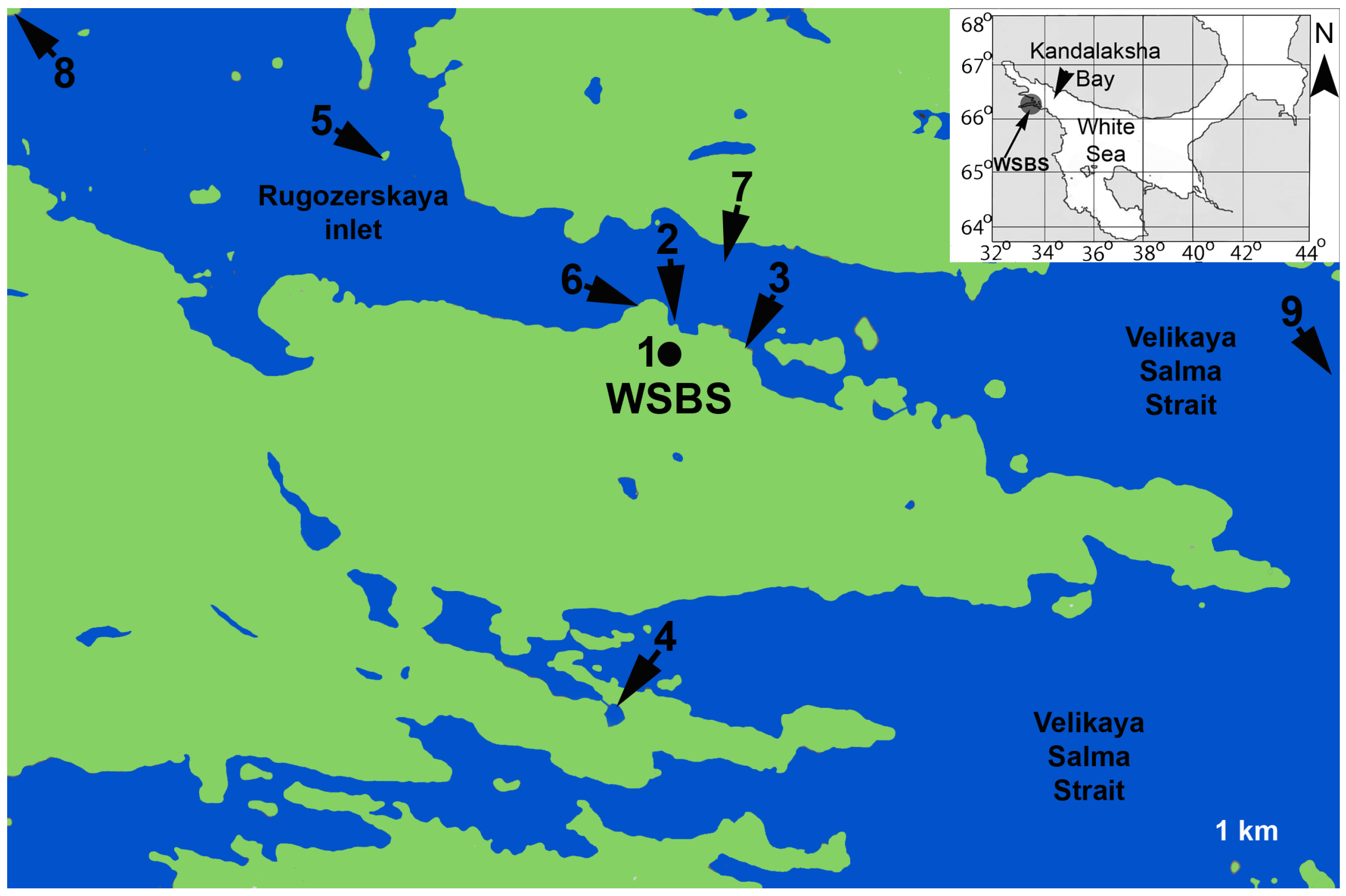
2.1. Sampling and Experimental Cultures
2.2. Morphological Analysis
2.3. Molecular Analysis
2.4. DNA Cloning
2.5. Phylogenetic Analysis
3. Results
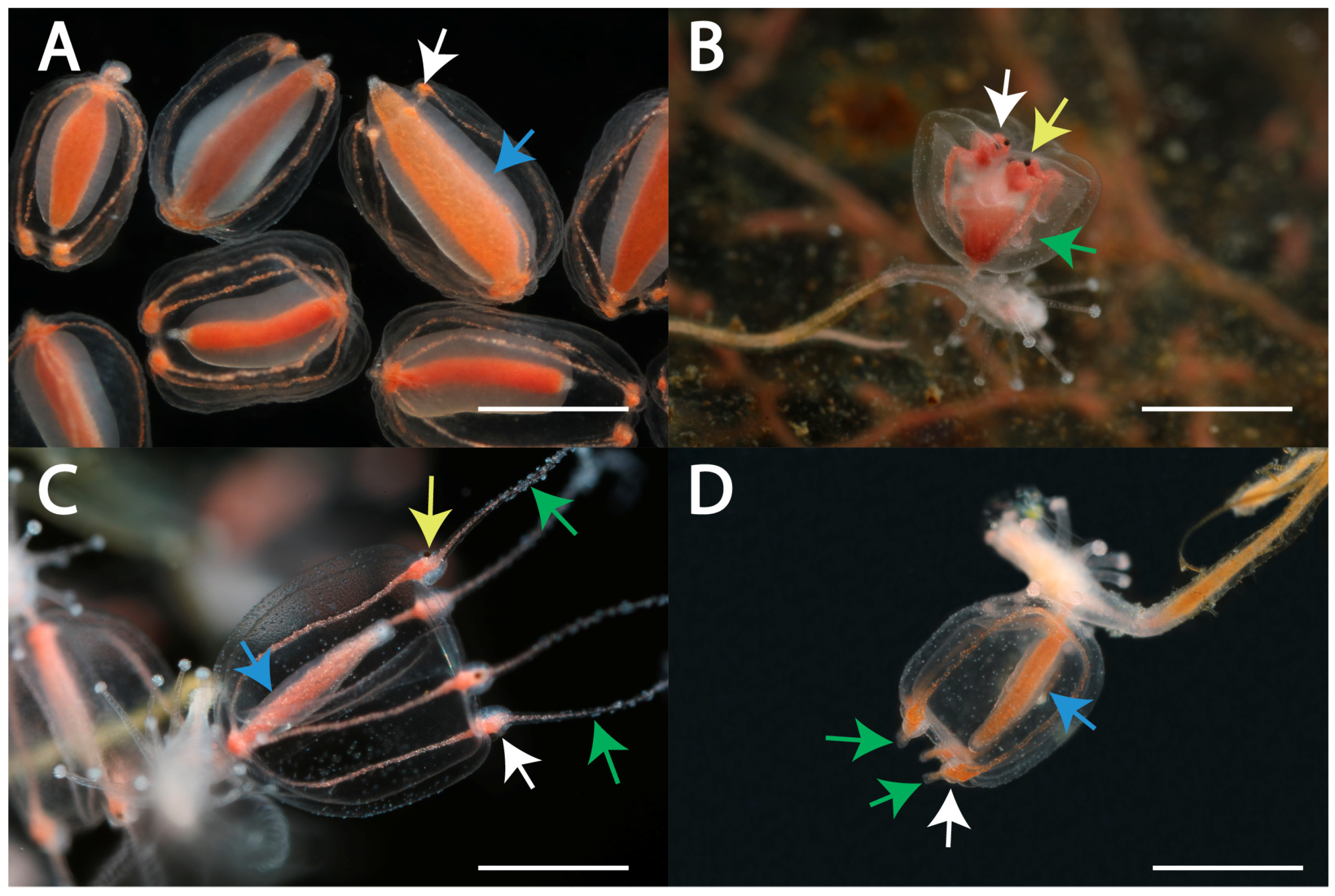
3.1. Morphotypes of Gonophores in Specimens of Sarsia spp.
3.2. Analysis of COI
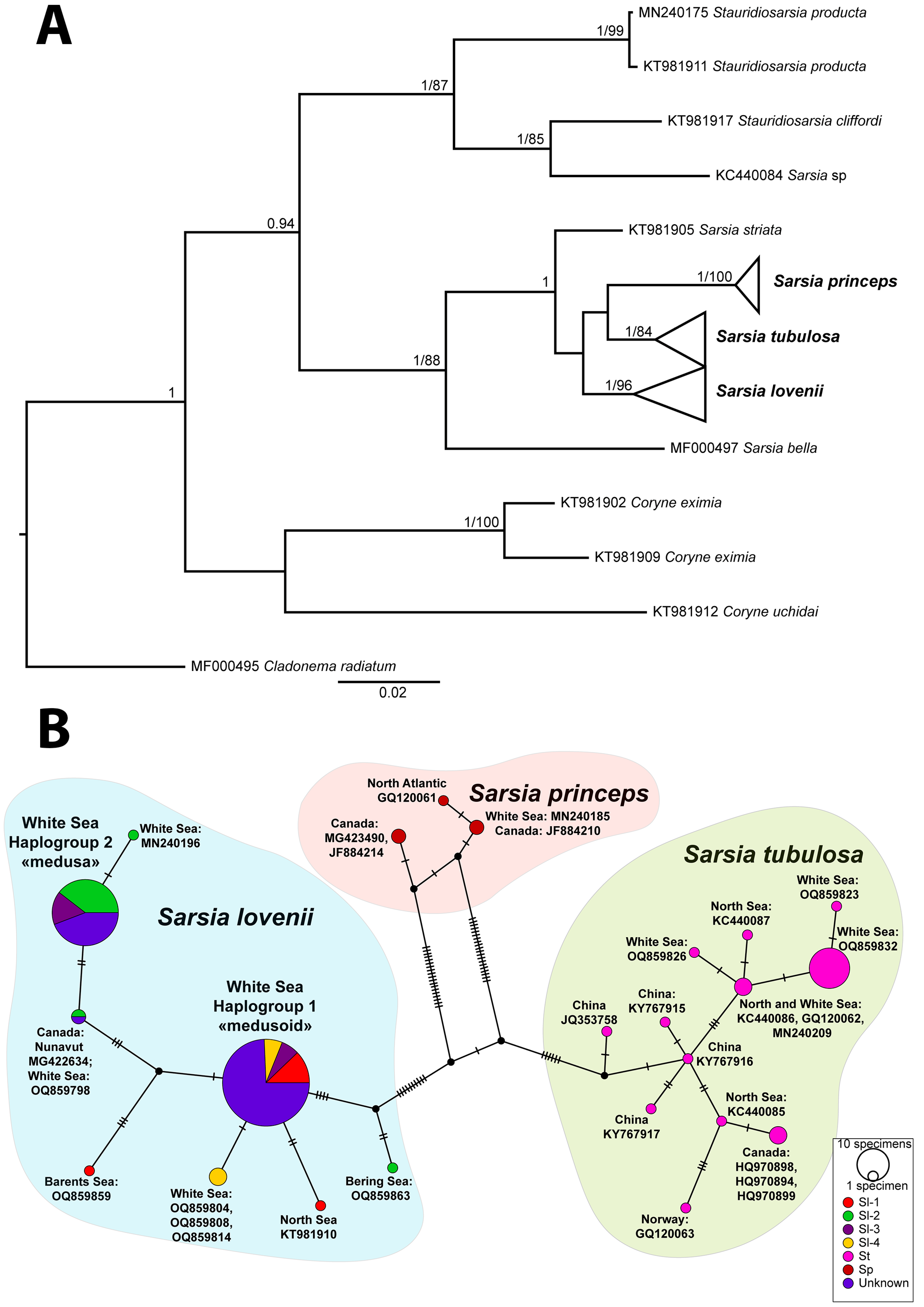
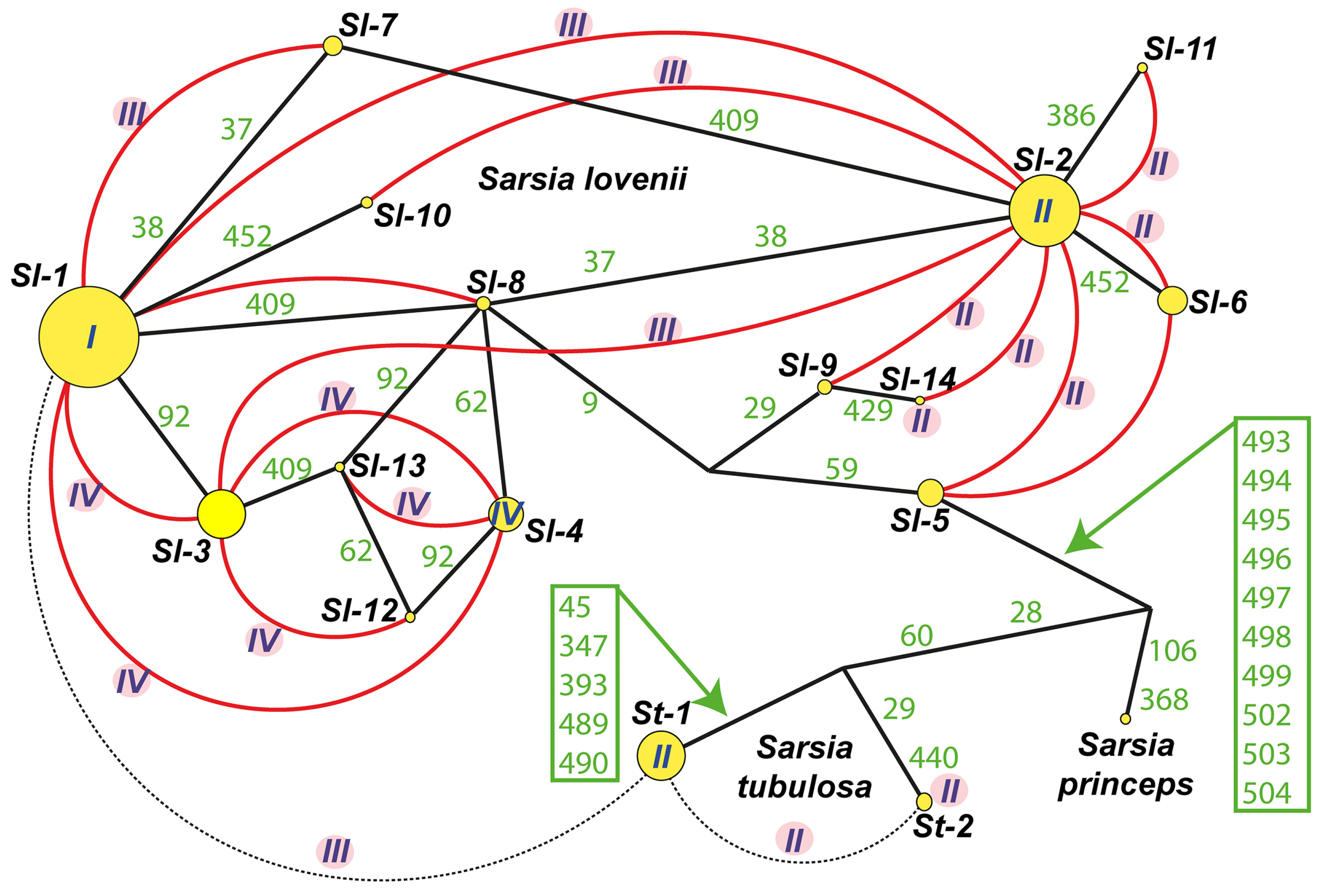
3.3. Analysis of ITS
4. Discussion
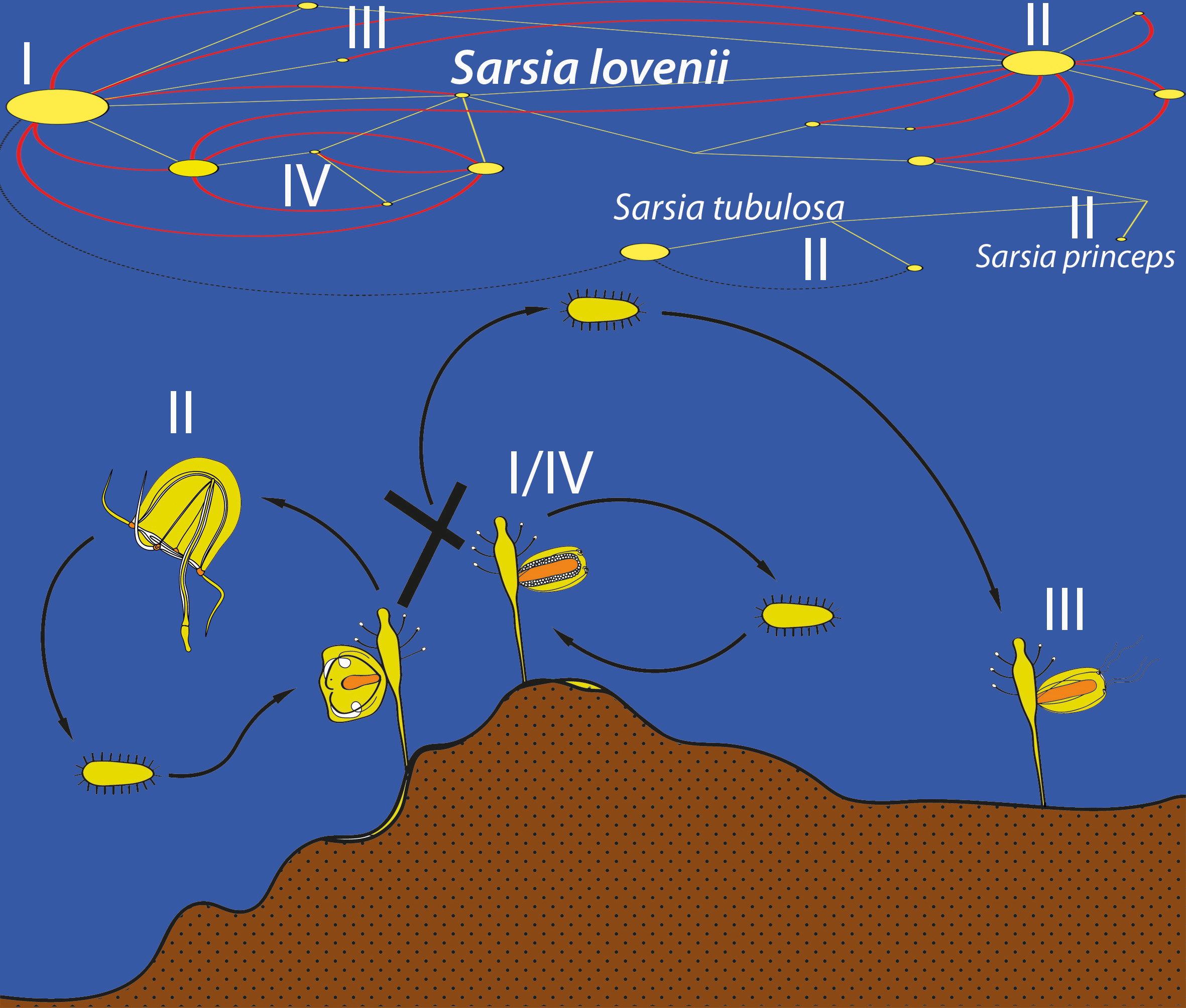
4.1. Morphotypes of Gonophores in Sarsia lovenii in the White Sea and Period of Reproduction
4.2. Crossing Experiments
4.3. Phylogeny, Species and COI Haplogroups of Sarsia spp. in the White Sea
4.4. Hybridization or Intragenomic Polymorphism?
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Barton, N.H.; Hewitt, G.M. Analysis of hybrid zones. Annu. Rev. Ecol. Evol. Syst. 1985, 16, 113–148. Available online: https://www.jstor.org/stable/2097045 (accessed on 19 August 2022). [CrossRef]
- Abbott, R.; Albach, D.; Ansell, S.; Arntzen, J.W.; Baird, S.J.; Bierne, N.; Boughman, J.; Brelsford, A.; Buerkle, C.A.; Buggs, R.; et al. Hybridization and speciation. J. Evol. Biol. 2013, 26, 229–246. [Google Scholar] [CrossRef] [PubMed]
- Gontier, N. Reticulate Evolution; Springer: Cham, Switzerland; Heidelberg, Germany; New York, NY, USA; Dordrecht, The Netherlands; London, UK, 2015; pp. 1–40. ISBN 978-3-319-16344-4. [Google Scholar]
- Richards, Z.T.; van Oppen, M.J.; Wallace, C.C.; Willis, B.L.; Miller, D.J. Some rare Indo-Pacific coral species are probable hybrids. PLoS ONE 2008, 3, e3240. [Google Scholar] [CrossRef] [PubMed]
- Fogarty, N.D.; Vollmer, S.V.; Levitan, D.R. Weak prezygotic isolating mechanisms in threatened Caribbean Acropora corals. PLoS ONE 2012, 7, e30486. [Google Scholar] [CrossRef] [PubMed]
- Richards, Z.T.; Hobbs, J.P.A. Hybridisation on coral reefs and the conservation of evolutionary novelty. Curr. Zool. 2015, 61, 132–145. [Google Scholar] [CrossRef]
- Hobbs, J.P.A.; Richards, Z.T.; Popovic, I.; Lei, C.; Staeudle, T.M.; Montanari, S.R.; DiBattista, J.D. Hybridisation and the evolution of coral reef biodiversity. Coral Reefs 2022, 41, 535–549. [Google Scholar] [CrossRef]
- Miller, R.L. Sperm chemotaxis in the hydromedusae. I. Species-specificity and sperm behavior. Mar. Biol. 1979, 53, 99–113. [Google Scholar] [CrossRef]
- Miller, R.L. Identification of sibling species within the “Sarsia tubulosa complex” at Friday Harbor, Washington (Hydrozoa: Anthomedusae). J. Exp. Mar. Biol. Ecol. 1982, 62, 153–172. [Google Scholar] [CrossRef]
- Kubota, S. Crossing-experiments between Japanese populations of three hydrozoans symbiotic with bivalves. Hydrobiologia 1991, 216, 429–436. [Google Scholar] [CrossRef]
- Soong, K.; Cho, L.C. Synchronized release of medusae from three species of hydrozoan fire corals. Coral Reefs 1998, 17, 145–154. [Google Scholar] [CrossRef]
- Brinckmann-Voss, A. Reproductive barriers and early development from hybridization experiments in two sympatric species of the genus Sarsia (Cnidaria, Hydrozoa, Anthoathecatae, Corynidae). Vie Milieu/Life Environ. 2002, 52, 121–130. Available online: https://hal.sorbonne-universite.fr/hal-03198882 (accessed on 19 January 2017).
- Prudkovsky, A.A.; Ekimova, I.A.; Neretina, T.V. A case of nascent speciation: Unique polymorphism of gonophores within hydrozoan Sarsia lovenii. Sci. Rep. 2019, 9, 15567. [Google Scholar] [CrossRef] [PubMed]
- Schroth, W.; Jarms, G.; Streit, B.; Schierwater, B. Speciation and phylogeography in the cosmopolitan marine moon jelly, Aurelia sp. BMC Evol. Biol. 2002, 2, 1. [Google Scholar] [CrossRef]
- Daly, M.; Brugler, M.R.; Cartwright, P.; Collins, A.G.; Dawson, M.N.; Fautin, D.G.; France, S.C.; McFadden, C.S.; Opresko, D.M.; Rodriguez, E.; et al. The phylum Cnidaria: A review of phylogenetic patterns and diversity 300 years after Linnaeus. In Linnaeus Tercentenary: Progress in Invertebrate Taxonomy, Z.-Q. Zhang and W.A. Shear, editors. Zootaxa 2007, 1668, 127–182. [Google Scholar] [CrossRef]
- Kayal, E.; Bentlage, B.; Sabrina Pankey, M.; Ohdera, A.H.; Medina, M.; Plachetzki, D.C.; Collins, A.G.; Ryan, J.F. Phylogenomics provides a robust topology of the major cnidarian lineages and insights on the origins of key organismal traits. BMC Evol. Biol. 2018, 18, 1–18. [Google Scholar] [CrossRef]
- Mills, C.E.; Marques, A.C.; Migotto, A.E.; Calder, D.R.; Hand, C. Hydrozoa: Polyps, hydromedusae, and siphonophora. In The Light and Smith Manual: Intertidal Invertebrates from Central California to Oregon; Carlton, J.T., Ed.; University of California Press: Berkeley, CA, USA, 2007; ISBN 9780520239395. [Google Scholar]
- Bouillon, J.; Gravili, C.; Gili, J.M.; Boero, F. An Introduction to Hydrozoa; Publications Scientifiques du Muséum: Paris, France, 2006. [Google Scholar]
- Cornelius, P.F. Medusa loss in leptolid Hydrozoa (Cnidaria), hydroid rafting, and abbreviated life-cycles among their remote-island faunae: An interim review. Sci. Mar. 1992, 56, 245–261. [Google Scholar]
- Leclère, L.; Schuchert, P.; Cruaud, C.; Couloux, A.; Manuel, M. Molecular phylogenetics of Thecata (Hydrozoa, Cnidaria) reveals long-term maintenance of life history traits despite high frequency of recent character changes. Syst. Biol. 2009, 58, 509–526. [Google Scholar] [CrossRef]
- Miglietta, M.P.; Cunningham, C.W. Evolution of life cycle, colony morphology, and host specificity in the family Hydractiniidae (Hydrozoa, Cnidaria). Evol. Int. J. Org. Evol. 2012, 66, 3876–3901. [Google Scholar] [CrossRef]
- Schuchert, P. Survey of the family Corynidae (Cnidaria, Hydrozoa). Rev. Suisse Zool. 2001, 108, 739–878. [Google Scholar] [CrossRef]
- Vilgalys, R.; Hester, M. Rapid genetic identification and mapping of enzymatically amplified ribosomal DNA from several Cryptococcus species. J. Bacteriol. Mycol. 1990, 172, 4238–4246. [Google Scholar] [CrossRef]
- Vilgalys, R. Conserved Primer Sequences for PCR Amplification of Fungal rDNA. 2018 Unpubl. Available online: https://sites.duke.edu/vilgalyslab/rdna_primers_for_fungi/ (accessed on 23 March 2019).
- Edgar, R.C. MUSCLE: Multiple sequence alignment with high accuracy and high throughput. Nucleic Acids Res. 2004, 32, 1792–1797. [Google Scholar] [CrossRef] [PubMed]
- Tamura, K.; Stecher, G.; Peterson, D.; Filipski, A.; Kumar, S. MEGA6: Molecular evolutionary genetics analysis version 6.0. Mol. Biol. Evol. 2013, 30, 2725–2729. [Google Scholar] [CrossRef] [PubMed]
- Darriba, D.; Taboada, G.L.; Doallo, R.; Posada, D. jModelTest 2: More models, new heuristics and parallel computing. Nat. Methods 2012, 9, 772. [Google Scholar] [CrossRef] [PubMed]
- Lartillot, N.; Lepage, T.; Blanquart, S. PhyloBayes 3: A Bayesian software package for phylogenetic reconstruction and molecular dating. Bioinformatics 2009, 25, 2286–2288. [Google Scholar] [CrossRef]
- Minh, B.Q.; Schmidt, H.A.; Chernomor, O.; Schrempf, D.; Woodhams, M.D.; Von Haeseler, A.; Lanfear, R. IQ-TREE 2: New models and efficient methods for phylogenetic inference in the genomic era. Mol. Biol. Evol. 2020, 37, 1530–1534. [Google Scholar] [CrossRef]
- Kalyaanamoorthy, S.; Minh, B.Q.; Wong, T.K.; Von Haeseler, A.; Jermiin, L.S. ModelFinder: Fast model selection for accurate phylogenetic estimates. Nat. Methods 2017, 14, 587–589. [Google Scholar] [CrossRef]
- Clement, M.; Snell, Q.; Walke, P.; Posada, D.; Crandall, K. TCS: Estimating gene genealogies. In Proceedings of the 16th International Parallel and Distributed Processing Symposium, Lauderdale, FL, USA, 15–19 April 2002; Volume 3, p. 0184. [Google Scholar] [CrossRef]
- Flot, J.-F. Champuru 1.0: A computer software for unraveling mixtures of two DNA sequences of unequal lengths. Mol. Ecol. Notes 2007, 7, 974–977. [Google Scholar] [CrossRef]
- Librado, P.; Rozas, J. DnaSP v5: A software for comprehensive analysis of DNA polymorphism data. Bioinformatics 2009, 25, 1451–1452. [Google Scholar] [CrossRef] [PubMed]
- Bandelt, H.-J.; Forster, P.; Röhl, A. Median-joining networks for inferring intraspecific phylogenies. Mol. Biol. Evol. 1999, 16, 37–48. [Google Scholar] [CrossRef]
- Dyomin, A.G.; Koshel, E.I.; Kiselev, A.M.; Saifitdinova, A.F.; Galkina, S.A.; Fukagawa, T.; Kostareva, A.A.; Gaginskaya, E.R. Chicken rRNA gene cluster structure. PLoS ONE 2016, 11, e0157464. [Google Scholar] [CrossRef] [PubMed]
- Schlötterer, C. Ribosomal DNA probes and primers. In Molecular Tools for Screening Biodiversity; Karp, A., Isaac, P.G., Ingram, D.S., Eds.; Chapman & Hall: London, UK, 1998; pp. 267–276. ISBN 978-94-010-6496-5. [Google Scholar]
- Ki, J.S.; Kim, I.C.; Lee, J.S. Comparative analysis of nuclear ribosomal DNA from the moon jelly Aurelia sp. 1 (Cnidaria: Scyphozoa) with characterizations of the 18S, 28S genes, and the intergenic spacer (IGS). In Proceedings of the Second International Jellyfish Blooms Symposium, Gold Coast, QLD, Australia, 24–27 June 2007; Springer: Dordrecht, The Netherlands, 2009; pp. 229–239. [Google Scholar]
- Li, X.; Xu, J.; He, Y.; Shen, S.; Zhu, J.; Shen, Z. The complete nuclear ribosomal DNA (nrDNA) cistron sequence of Pyropia yezoensis (Bangiales, Rhodophyta). J. Appl. Phycol. 2016, 28, 663–669. [Google Scholar] [CrossRef]
- Guo, Z.; Han, L.; Ding, Y.; Hou, X.; Liang, Z. Molecular characterisation of the complete nuclear ribosomal DNA sequence of the blacklip abalone Haliotis rubra. New Zealand J. Mar. Freshw. Res. 2018, 52, 430–443. [Google Scholar] [CrossRef]
- Baldwin, B.G.; Sanderson, M.J.; Porter, J.M.; Wojciechowski, M.F.; Campbell, C.S.; Donoghue, M.J. The ITS region of nuclear ribosomal DNA: A valuable source of evidence on angiosperm phylogeny. Ann. Mo. Bot. Gard. 1995, 82, 247–277. [Google Scholar] [CrossRef]
- Hilário, S.; Santos, L.; Phillips, A.J.; Alves, A. Caveats of the internal transcribed spacer region as a barcode to resolve species boundaries in Diaporthe. Fungal Biol. 2022, 126, 54–74. [Google Scholar] [CrossRef]
- Kuninaga, S.; Natsuaki, T.; Takeuchi, T.; Yokosawa, R. Sequence variation of the rDNA ITS regions within and between anastomosis groups in Rhizoctonia solani. Curr. Genet. 1997, 32, 237–243. [Google Scholar] [CrossRef]
- Sang, T.; Crawford, D.J.; Stuessy, T.F. Documentation of reticulate evolution in peonies (Paeonia) using internal transcribed spacer sequences of nuclear ribosomal DNA: Implications for biogeography and concerted evolution. Proc. Natl. Acad. Sci. USA 1995, 92, 6813–6817. [Google Scholar] [CrossRef]
- Forsman, Z.H.; Hunter, C.L.; Fox, G.E.; Wellington, G.M. Is the ITS region the solution to the “species problem” in corals? Intragenomic variation and alignment permutation in Porites, Siderastrea and outgroup taxa. In Proceedings of the 10th International Coral Reef Symposium, Okinawa, Japan, 28 June–2 July 2004; Volume 1, pp. 14–23. [Google Scholar]
- Campbell, R.D.; Iniguez, A.R.; Iniguez, A.J.; Martínez, D.E. Hydra of Hawaii: Phylogenetic relationships with continental species. Hydrobiologia 2013, 713, 199–205. [Google Scholar] [CrossRef]
- Cunha, A.F.; Genzano, G.N.; Marques, A.C. Reassessment of morphological diagnostic characters and species boundaries requires taxonomical changes for the genus Orthopyxis L. Agassiz, 1862 (Campanulariidae, Hydrozoa) and some related campanulariids. PLoS ONE 2015, 10, e0117553. [Google Scholar] [CrossRef] [PubMed]
- Postaire, B.; Magalon, H.; Bourmaud, C.A.F.; Gravier-Bonnet, N.; Bruggemann, J.H. Phylogenetic relationships within Aglaopheniidae (Cnidaria, Hydrozoa) reveal unexpected generic diversity. Zool. Scr. 2016, 45, 103–114. [Google Scholar] [CrossRef]
- Schuchert, P. High genetic diversity in the hydroid Plumularia setacea: A multitude of cryptic species or extensive population subdivision? Mol. Phylogenetics Evol. 2014, 76, 1–9. [Google Scholar] [CrossRef]
- Schuchert, P. DNA barcoding of some Pandeidae species (Cnidaria, Hydrozoa, Anthoathecata). Rev. Suisse Zool. 2018, 125, 101–127. [Google Scholar] [CrossRef]
- Prudkovsky, A.A.; Neretina, T.V. The life cycle of Catablema vesicarium (A. Agassiz, 1862) (Hydrozoa, Pandeidae). Polar Biol. 2016, 39, 533–542. [Google Scholar] [CrossRef]
- Reimer, J.D.; Takishita, K.; Ono, S.; Tsukahara, J.; Maruyama, T. Molecular evidence suggesting interspecific hybridization in Zoanthus spp. (Anthozoa: Hexacorallia). Zool. Sci. 2007, 24, 346–359. [Google Scholar] [CrossRef] [PubMed]
- Liao, D.; Pavelitz, T.; Kidd, J.R.; Kidd, K.K.; Weiner, A.M. Concerted evolution of the tandemly repeated genes encoding human U2 snRNA (the RNU2 locus) involves rapid intrachromosomal homogenization and rare interchromosomal gene conversion. EMBO J. 1997, 16, 588–598. [Google Scholar] [CrossRef]
- Elder Jr, J.F.; Turner, B.J. Concerted evolution of repetitive DNA sequences in eukaryotes. Q. Rev. Biol. 1995, 70, 297–320. [Google Scholar] [CrossRef]
- Hillis, D.M.; Dixon, M.T. Ribosomal DNA: Molecular evolution and phylogenetic inference. Q. Rev. Biol. 1991, 66, 411–453. [Google Scholar] [CrossRef]
- Wang, S.; Zhang, L.; Zhan, A.; Wang, X.; Liu, Z.; Hu, J.; Bao, Z. Patterns of concerted evolution of the rDNA family in a natural population of zhikong scallop, Chlamys farreri. J. Mol. Evol. 2007, 65, 660–667. [Google Scholar] [CrossRef] [PubMed]
- Naidoo, K.; Steenkamp, E.T.; Coetzee, M.P.; Wingfield, M.J.; Wingfield, B.D. Concerted evolution in the ribosomal RNA cistron. PLoS ONE 2013, 8, e59355. [Google Scholar] [CrossRef]
- Harris, D.J.; Crandall, K.A. Intragenomic variation within ITS1 and ITS2 of freshwater crayfishes (Decapoda: Cambaridae): Implications for phylogenetic and microsatellite studies. Mol. Biol. Evol. 2000, 17, 284–291. [Google Scholar] [CrossRef]
- Vollmer, S.V.; Palumbi, S.R. Testing the utility of internally transcribed spacer sequences in coral phylogenetics. Mol. Ecol. 2004, 13, 2763–2772. [Google Scholar] [CrossRef]
- Wei, N.W.V.; Wallace, C.C.; Dai, C.F.; Pillay, K.R.M.; Chen, C.A. Analyses of the Ribosomal Internal Transcribed Spacers (ITS) and the 5.8 S Gene Indicate that Extremely High rDNA Heterogeneity is a Unique Feature in the Scleractinian Coral Genus Acropora (Scleractinia; Acroporidae). Zool. Stud. 2006, 45, 404–418. [Google Scholar]
- Sánchez, J.A.; Dorado, D. Intragenomic ITS2 variation in Caribbean seafans. In Proceedings of the 11th International Coral Reef Symposium, Ft. Lauderdale, FL, USA, 7–11 July 2008; pp. 1383–1387. [Google Scholar]
- Gong, L.; Shi, W.; Yang, M.; Si, L.; Kong, X. Non-concerted evolution in ribosomal ITS2 sequence in Cynoglossus zanzibarensis (Pleuronectiformes: Cynoglossidae). Biochem. Syst. Ecol. 2016, 66, 181–187. [Google Scholar] [CrossRef]
- Xu, B.; Zeng, X.M.; Gao, X.F.; Jin, D.P.; Zhang, L.B. ITS non-concerted evolution and rampant hybridization in the legume genus Lespedeza (Fabaceae). Sci. Rep. 2017, 7, 40057. [Google Scholar] [CrossRef] [PubMed]
- O’Donnell, K.; Cigelnik, E. Two divergent intragenomic rDNA ITS2 types within a monophyletic lineage of the Fungus fusarium are nonorthologous. Mol. Phylogenet. Evol. 1997, 7, 103–116. [Google Scholar] [CrossRef] [PubMed]
- Muir, G.; Fleming, C.C.; Schlötterer, C. Three divergent rDNA clusters predate the species divergence in Quercus petraea (Matt.) Liebl. and Quercus robur L. Mol. Biol. Evol. 2001, 18, 112–119. [Google Scholar] [CrossRef] [PubMed]
- Van Oppen, M.J.H.; Wörheide, G.; Takabayashi, M. Nuclear markers in evolutionary and population genetic studies of scleractinian corals and sponges. In Proceedings of the 9th International Coral Reef Symposium, Bali, Indonesia, 23–27 October 2000; Volume 1, pp. 131–138. [Google Scholar]
- Aguilar, C.; Reimer, J.D. Molecular phylogenetic hypotheses of Zoanthus species (Anthozoa: Hexacorallia) using RNA secondary structure of the internal transcribed spacer 2 (ITS2). Mar. Biodivers. 2010, 40, 195–204. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Schemes of Crossing | DNA Isolates |
|---|---|
| S. lovenii medusa female × S. lovenii medusoid male | H140, H144, H151, H153, H157, H168 |
| S. lovenii medusa male × S. lovenii medusoid female | H149, H150, H158, H159, H170 |
| S. lovenii medusa female × S. lovenii male | H122 |
| S. lovenii medusoid female × S. tubulosa male | H163 |
| S. tubulosa female × S. tubulosa male | H249 |
| S. lovenii medusa female × S. lovenii medusa male | H238, H251 |
| S. lovenii hybrid F2: hybrid F1 males (H159 + H140) × S. lovenii medusa female | H233, H236 |
| Year of Collection | Sampling Location | Number of Specimens | Number of Medusa Specimens/Number of Polyp Specimens with Mature Gonophores |
|---|---|---|---|
| 2015 | Aquarium | 6 | 0/1 [13] |
| 2016 | In the sea, WSBS | 9 | 4/0 [13] |
| 2017 | In the sea, WSBS | 23 | 22/1 [13] |
| 2018 | In the sea, WSBS | 5 | 1/4 [13] |
| 2019 | In the sea, WSBS, Bering Sea, Barents Sea | 43 | 1/10 |
| 2019 | Aquarium | 17 | 0/5 |
| 2019 | Aquarium (crossing experiment) | 13 | 0/11 |
| 2020 | In the sea, WSBS | 43 | 14/8 |
| 2020 | Aquarium (crossing experiment) | 4 | 0/0 |
| 2021 | In the sea, WSBS | 20 | 0/17 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Prudkovsky, A.; Vetrova, A.; Kremnyov, S. What Does “ITS” Say about Hybridization in Lineages of Sarsia (Corynidae, Hydrozoa) from the White Sea? Diversity 2023, 15, 675. https://doi.org/10.3390/d15050675
Prudkovsky A, Vetrova A, Kremnyov S. What Does “ITS” Say about Hybridization in Lineages of Sarsia (Corynidae, Hydrozoa) from the White Sea? Diversity. 2023; 15(5):675. https://doi.org/10.3390/d15050675
Chicago/Turabian StylePrudkovsky, Andrey, Alexandra Vetrova, and Stanislav Kremnyov. 2023. "What Does “ITS” Say about Hybridization in Lineages of Sarsia (Corynidae, Hydrozoa) from the White Sea?" Diversity 15, no. 5: 675. https://doi.org/10.3390/d15050675