
Niche Characteristics of Alternanthera philoxeroide-Invaded Plant Communities in Heterogeneous Habitats and Their Latitudinal Trends
Abstract
:1. Introduction
2. Materials and Methods
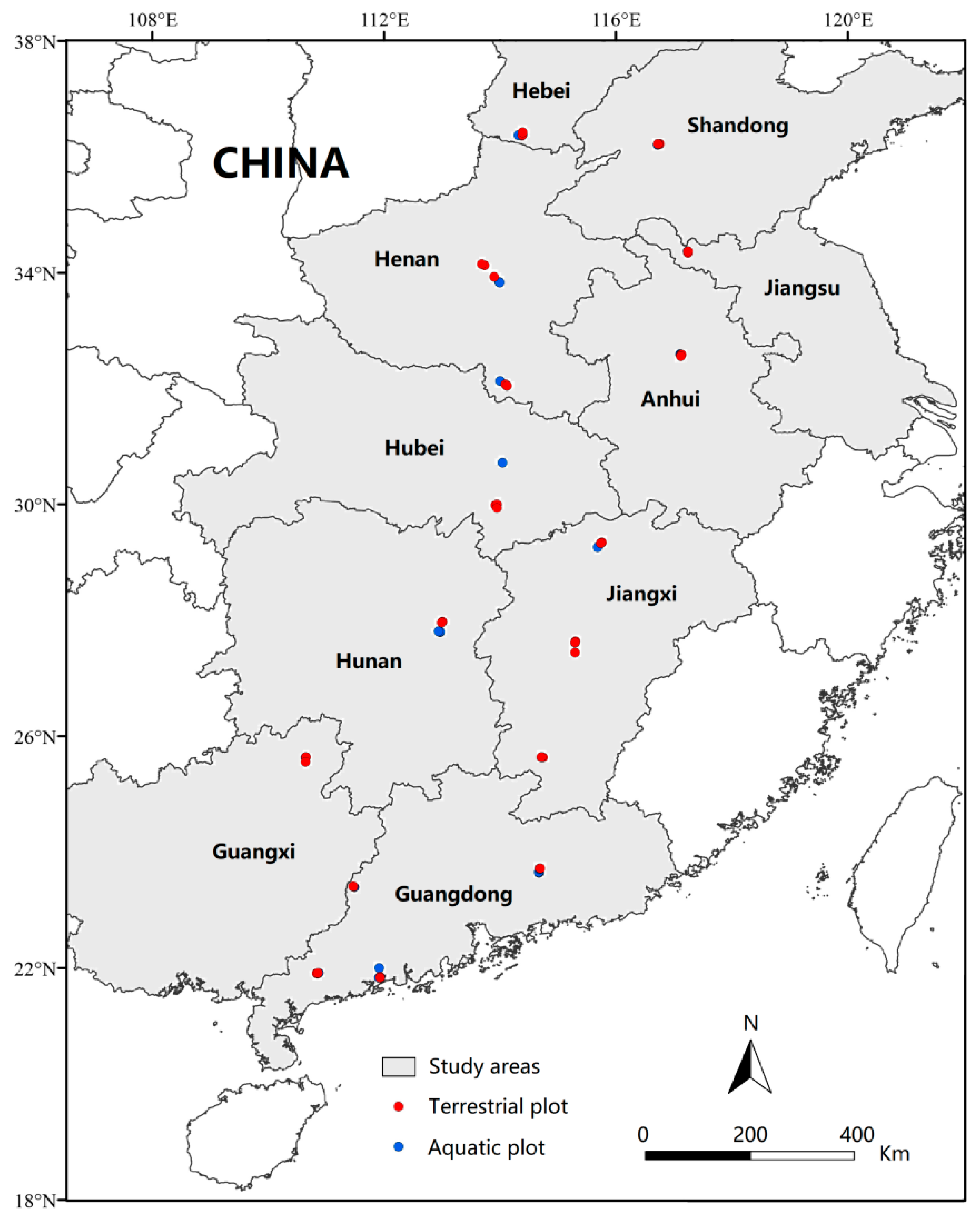
2.1. Site Selection
2.2. Field Survey
2.3. Data Calculation
2.3.1. Importance Value
- IV = (relative abundance + relative height + relative coverage)/3.
- The total IV was the sum of a plant species’ IV in 40 terrestrial or 40 aquatic plots.
2.3.2. Niche Characteristics

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Code | Plant Species | Frequency | IV | B(L) | Code | Plant Species | Frequency | IV | B(L) |
|---|---|---|---|---|---|---|---|---|---|
| Terrestrial Community | Aquatic Community | ||||||||
| 1 | Alternanthera philoxeroides | 40 | 17.360 | 0.960 | 1 | Alternanthera philoxeroides | 40 | 23.752 | 0.943 |
| 2 | Digitaria sanguinalis | 30 | 2.936 | 0.414 | 2 | Paspalum paspaloides | 25 | 5.427 | 0.424 |
| 3 | Echinochloa crusgalli | 28 | 1.112 | 0.314 | 3 | Polygonum hydropiper | 20 | 0.727 | 0.301 |
| 4 | Bidens pilosa | 25 | 0.739 | 0.354 | 4 | Echinochloa crusgalli | 18 | 1.226 | 0.210 |
| 5 | Ipomoea nil | 23 | 0.599 | 0.294 | 5 | Bidens pilosa | 14 | 0.341 | 0.232 |
| 6 | Eleusine indica | 22 | 1.064 | 0.278 | 6 | Commelina communis | 9 | 0.534 | 0.107 |
| 7 | Cyperus votundus | 21 | 0.901 | 0.204 | 7 | Cyperus rotundus | 8 | 0.198 | 0.126 |
| 8 | Setaria viridis | 19 | 0.972 | 0.322 | 8 | Kyllinga brevifolia | 7 | 0.123 | 0.146 |
| 9 | Commelina communis | 18 | 1.301 | 0.285 | 9 | Eclipta prostrata | 7 | 0.217 | 0.086 |
| 10 | Cyperus rotundus | 16 | 0.303 | 0.269 | 10 | Humulus scandens | 7 | 0.444 | 0.099 |
| 11 | Acalypha australis | 16 | 0.198 | 0.287 | 11 | Ipomoea nil | 7 | 0.232 | 0.079 |
| 12 | Humulus scandens | 15 | 1.120 | 0.203 | 12 | Lemna minor | 6 | 0.284 | 0.115 |
| 13 | Ageratum conopids | 15 | 0.680 | 0.160 | 13 | Digitaria sanguinalis | 6 | 0.123 | 0.120 |
| 14 | Erigeron canadensis | 15 | 0.425 | 0.138 | 14 | Acorus calamus | 5 | 0.146 | 0.106 |
| 15 | Artemisia argyi | 15 | 0.342 | 0.278 | 15 | Glycine soja | 5 | 0.309 | 0.075 |
| 16 | Eclipta prostrata | 15 | 0.195 | 0.214 | 16 | Alternanthera sessilis | 5 | 0.184 | 0.074 |
| 17 | Cynodon dactylon | 14 | 0.695 | 0.108 | 17 | Eleusine indica | 5 | 0.190 | 0.096 |
| 18 | Polygonum hydropiper | 14 | 0.677 | 0.129 | 18 | Trigastrotheca stricta | 5 | 0.291 | 0.052 |
| 19 | Amaranthus blitum | 13 | 0.170 | 0.132 | 19 | Cyperus votundus | 5 | 0.169 | 0.111 |
| 20 | Erigeron annuus | 12 | 0.231 | 0.123 | 20 | Polygonum perfoliatum | 4 | 0.061 | 0.092 |
| 21 | Oxalis corniculata | 10 | 0.250 | 0.099 | 21 | Phragmites australis | 4 | 0.187 | 0.083 |
| 22 | Paspalum paspaloides | 9 | 0.435 | 0.155 | 22 | Senna tora | 4 | 0.043 | 0.060 |
| 23 | Broussonetia papyrifera | 9 | 0.296 | 0.112 | 23 | Symphyotrichum subulatum | 4 | 0.197 | 0.053 |
| 24 | Ludwigia prostrata | 9 | 0.168 | 0.113 | 24 | Microstegium nodosum | 4 | 0.146 | 0.041 |
| 25 | Portulaca oleracea | 9 | 0.106 | 0.122 | |||||
2.4. Statistical Analyses
3. Results
3.1. Niche Characteristics at the Level of the Whole Community
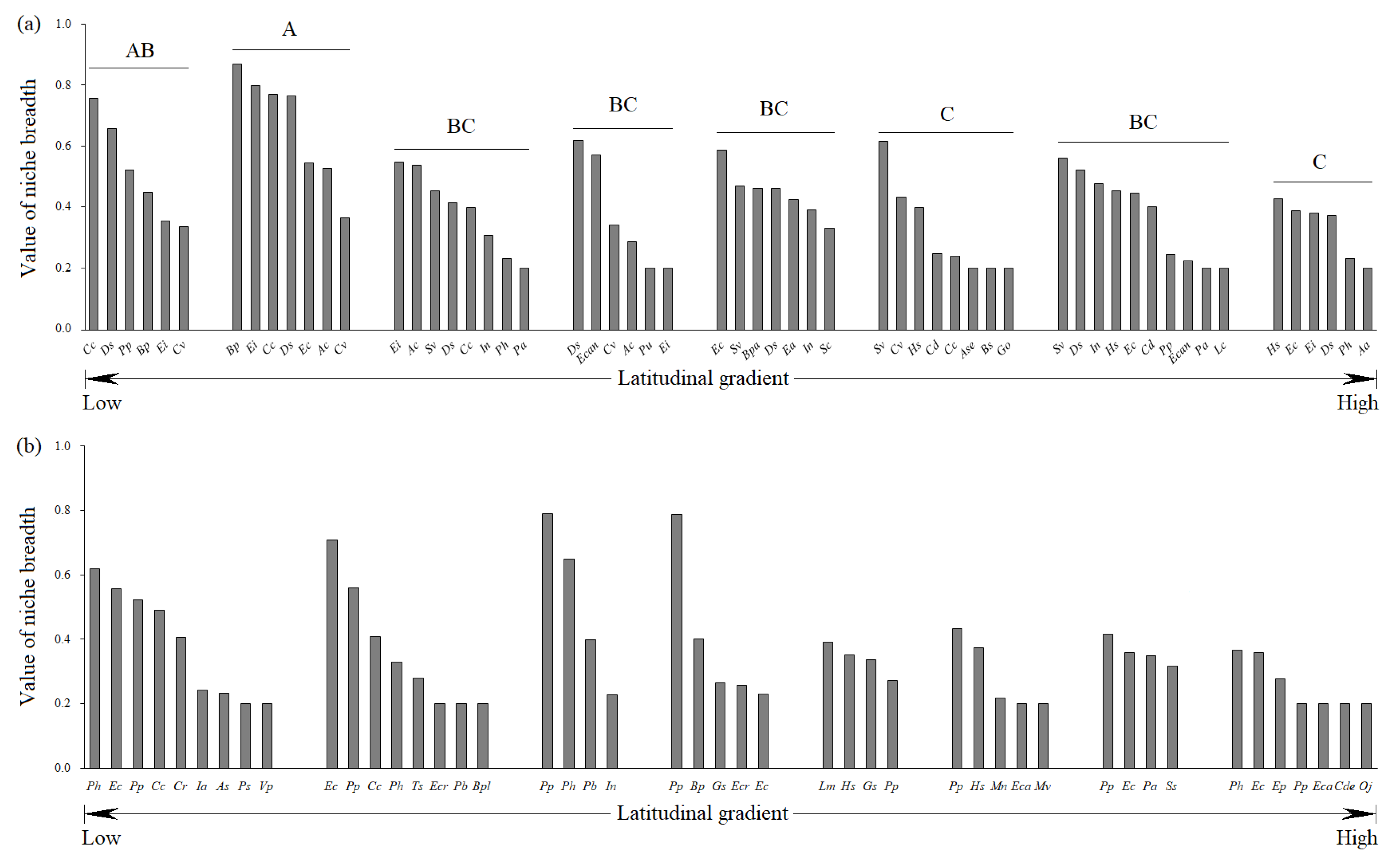
3.1.1. Niche Breadth
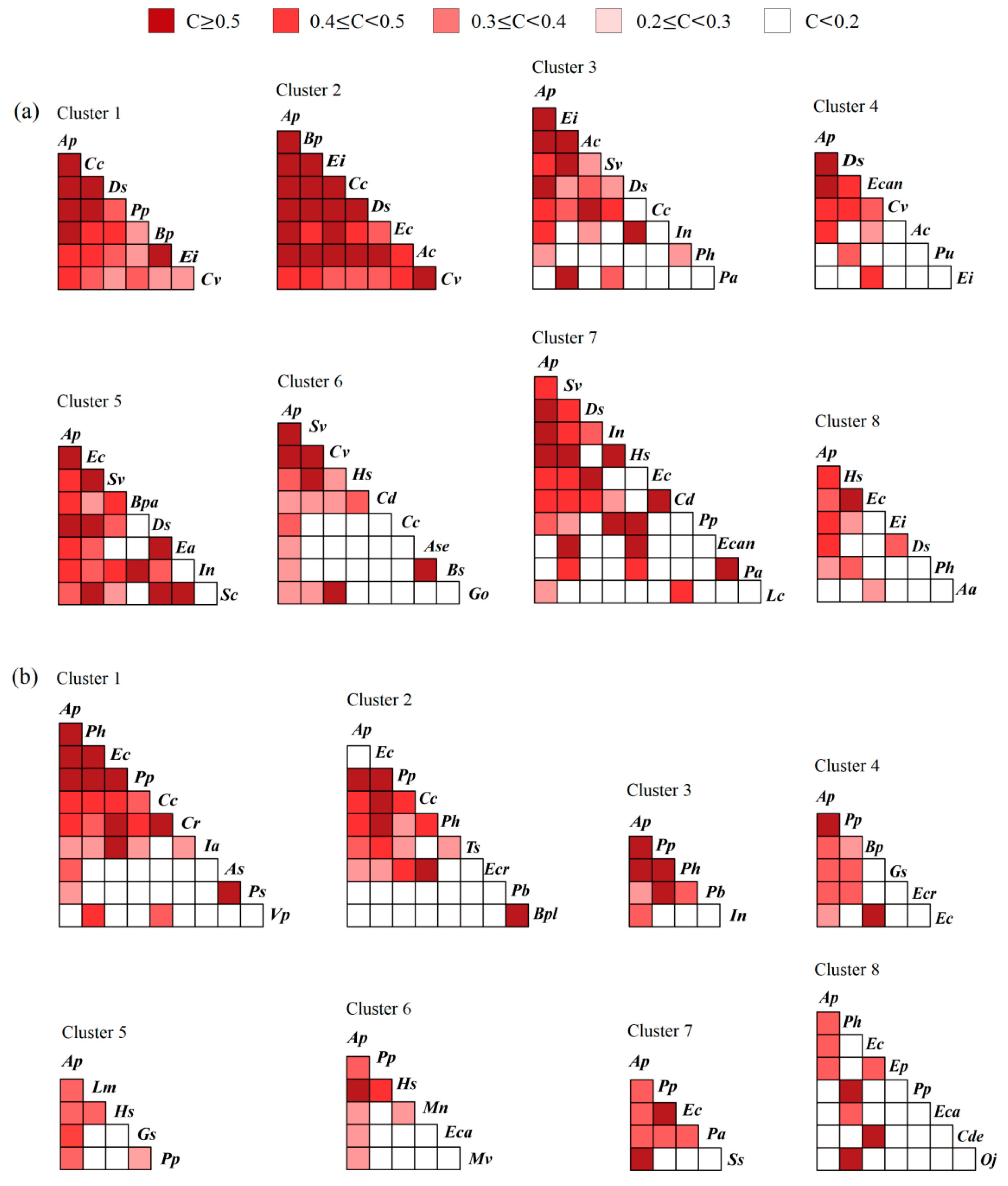
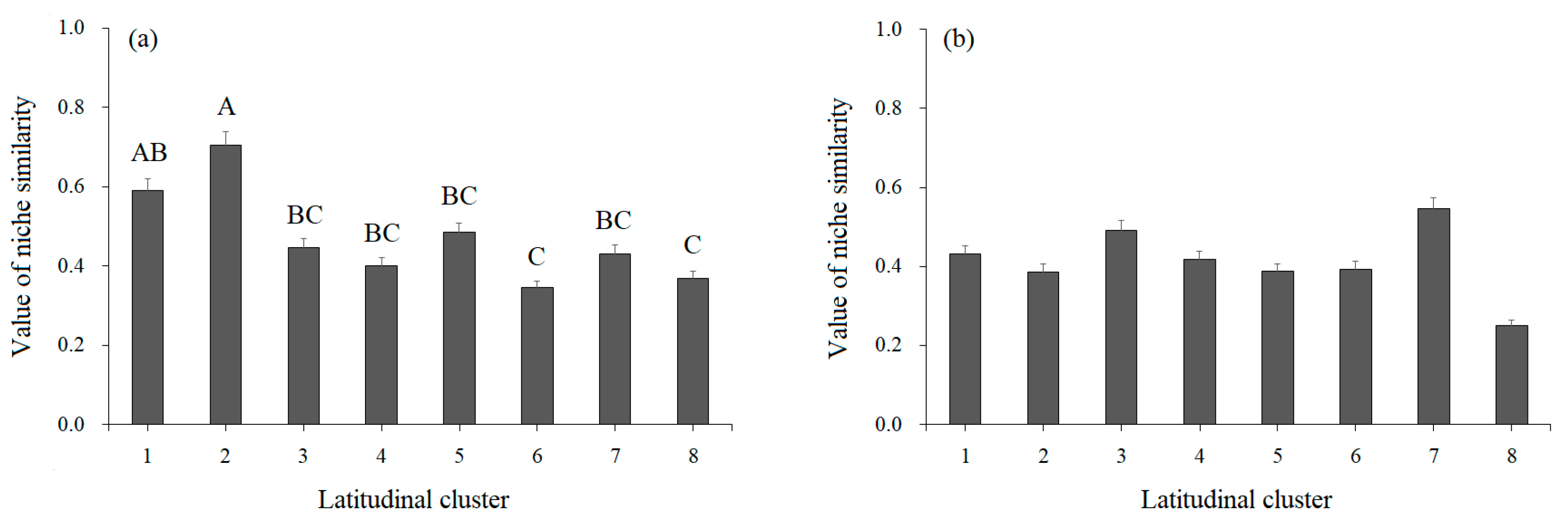
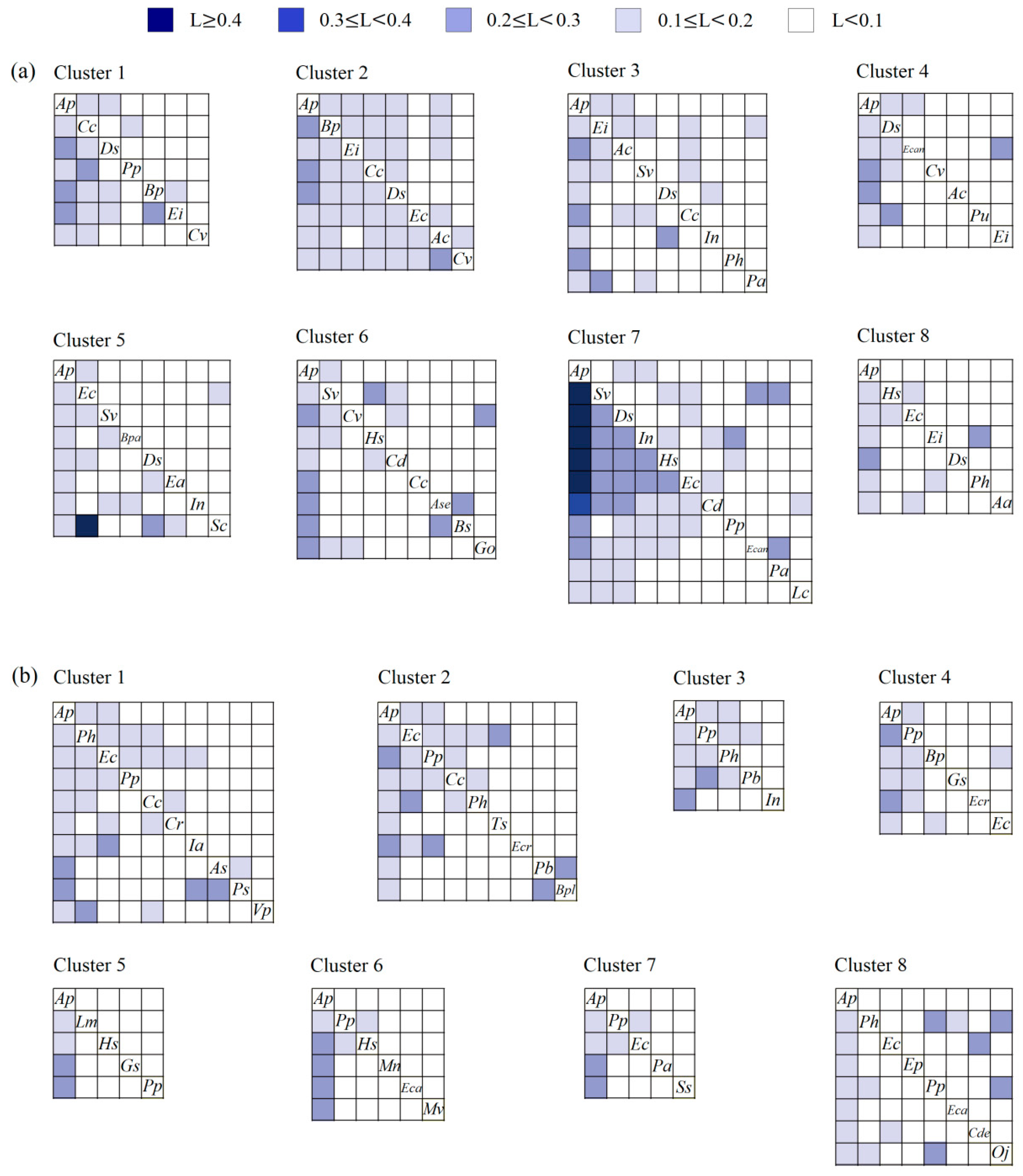
3.1.2. Niche Similarity

3.1.3. Niche Overlap
3.2. Latitudinal Trends of Niche Characteristics
3.2.1. Variations in Niche Breadth along the Latitudinal Gradient

3.2.2. Variations in Niche Similarity along the Latitudinal Gradient

3.2.3. Variations in Niche Overlap along the Latitudinal Gradient

4. Discussion
4.1. Causes of Niche Characteristic Formation in Heterogeneous Habitats
4.2. Causes of Niche Breadth Variations along the Latitudinal Gradient in Heterogeneous Habitats
4.3. Causes of Niche Similarity and Overlap Variations along the Latitudinal Gradient in Heterogeneous Habitats
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Seebens, H.; Blackburn, T.M.; Dyer, E.E.; Genovesi, P.; Hulme, P.E.; Jeschke, J.M.; Pagad, S.; Pyšek, P.; Winter, M.; Arianoutsou, M.; et al. No saturation in the accumulation of alien species worldwide. Nat. Commun. 2017, 8, 14435. [Google Scholar] [CrossRef] [PubMed]
- Carboni, M.; Livingstone, S.W.; Isaac, M.E.; Cadotte, M.W. Invasion drives plant diversity loss through competition and ecosystem modification. J. Ecol. 2021, 109, 3587–3601. [Google Scholar] [CrossRef]
- Pabst, R.; Dias, F.S.; Borda-de-Água, L.; Rodríguez-González, P.M.; Capinha, C. Assessing and predicting the distribution of riparian invasive plants in continental Portugal. Front. Ecol. Evol. 2022, 10, 875578. [Google Scholar] [CrossRef]
- Lebbink, G.; Dwyer, J.; Fensham, R. An invasive grass species has both local and broad-scale impacts on diversity: Potential mechanisms and implications. J. Veg. Sci. 2021, 32, 1240–1253. [Google Scholar] [CrossRef]
- Yue, M.; Shen, H.; Ye, W.; Li, W.; Chen, J. Winter low temperature disturbance in the southern subtropics of China promotes the competitiveness of an invasive plant. Biol. Invasions 2021, 23, 2913–2925. [Google Scholar] [CrossRef]
- Gross, N.; Liancourt, P.; Butters, R.; Duncan, R.; Hulme, P.E. Functional equivalence, competitive hierarchy and facilitation determine species coexistence in highly invaded grasslands. New Phytol. 2015, 206, 175–186. [Google Scholar] [CrossRef]
- Wang, C.; Yu, Y.; Cheng, H.; Du, D. Which factor contributes most to the invasion resistance of native plant communities under the co-invasion of two invasive plant species? Sci. Total Environ. 2022, 813, 152628. [Google Scholar] [CrossRef]
- Wu, H.; Ding, J.Q. Abiotic and biotic determinants of plant diversity in aquatic communities invaded by water hyacinth (Eichhornia crassipes (Mart.) Solms). Front. Plant Sci. 2020, 11, 1306. [Google Scholar] [CrossRef]
- Dubey, D.; Kumar, S.; Dutta, V. In-situ anatomical and elemental response of aquatic macrophytes against nutrient enrichment in freshwater tropical lakes. Appl. Water Sci. 2022, 12, 163. [Google Scholar] [CrossRef]
- Huang, X.; Ke, F.; Li, Q.; Zhao, Y.; Guan, B.; Li, K. Functional traits underlying performance variations in the overwintering of the cosmopolitan invasive plant water hyacinth (Eichhornia crassipes) under climate warming and water drawdown. Ecol. Evol. 2022, 12, e9181. [Google Scholar] [CrossRef]
- Kuebbing, S.E.; Nuñez, M.A. Invasive non-native plants have a greater effect on neighbouring natives than other non-natives. Nat. Plants 2016, 2, 1613. [Google Scholar] [CrossRef]
- Byers, J.E.; Noonburg, E.G. Scale dependent effects of biotic resistance. Ecology 2003, 84, 1428–1433. [Google Scholar] [CrossRef]
- Fridley, J.D.; Stachowicz, J.J.; Naeem, S.; Sax, D.F.; Seabloom, E.W.; Smith, M.D.; Stohlgren, T.J.; Tilman, D.; Von Holle, B. The invasion paradox: Reconciling pattern and process in species invasions. Ecology 2007, 88, 3–17. [Google Scholar] [CrossRef]
- Powell, K.I.; Chase, J.M.; Knight, T.M. Invasive plants have scale-dependent effects on diversity by altering species-area relationships. Science 2013, 339, 316–318. [Google Scholar] [CrossRef]
- Tilman, D. Niche tradeoffs, neutrality, and community structure: A stochastic theory of resource competition, invasion, and community assembly. Proc. Natl. Acad. Sci. USA 2004, 101, 10854–10861. [Google Scholar] [CrossRef]
- Guisan, A.; Petitpierre, B.; Brönnimann, O.; Daehler, C.; Kueffer, C. Unifying niche shift studies: Insights from biological invasions. Trends Ecol. Evol. 2014, 29, 260–269. [Google Scholar] [CrossRef]
- Cai, Q.; Welk, E.; Ji, C.; Fang, W.; Sabatini, F.M.; Zhu, J.; Zhu, J.; Tang, Z.; Attorre, F.; Campos, J.A.; et al. The relationship between niche breadth and range size of beech (Fagus) species worldwide. J. Biogeogr. 2021, 48, 1240–1253. [Google Scholar] [CrossRef]
- Pastore, A.I.; Barabás, G.; Bimler, M.D.; Mayfield, M.M.; Miller, T.E. The evolution of niche overlap and competitive differences. Nat. Ecol. Evol. 2021, 5, 330–337. [Google Scholar] [CrossRef]
- Cadotte, M.W.; Tucker, C.M. Should Environmental Filtering be Abandoned? Trends Ecol. Evol. 2017, 32, 429–437. [Google Scholar] [CrossRef]
- Liu, C.; Wolter, C.; Courchamp, F.; Roura-Pascual, N.; Jeschke, J.M. Biological invasions reveal how niche change affects the transferability of species distribution models. Ecology 2022, 103, e3719. [Google Scholar] [CrossRef]
- Yuan, Y.; Tang, X.; Liu, M.; Liu, X.; Tao, J. Species distribution models of the Spartina alterniflora Loisel in its origin and invasive country reveal an ecological niche shift. Front. Plant Sci. 2021, 12, 738769. [Google Scholar] [CrossRef] [PubMed]
- Fridley, J.D. Extended leaf phenology and the autumn niche in deciduous forest invasions. Nature 2012, 485, 359–362. [Google Scholar] [CrossRef] [PubMed]
- Datta, A.; Schweiger, O.; Ingolf, K. Niche expansion of the invasive plant species Ageratina adenophora despite evolutionary constraints. J. Biogeogr. 2019, 46, 1306–1315. [Google Scholar]
- Enders, M.; Havemann, F.; Ruland, F.; Bernard-Verdier, M.; Catford, J.A.; Gómez-Aparicio, L.; Haider, S.; Heger, T.; Kueffer, C.; Kühn, I.; et al. A conceptual map of invasion biology: Integrating hypotheses into a consensus network. Glob. Ecol. Biogeogr. 2020, 29, 978–991. [Google Scholar] [CrossRef]
- Yang, Y.H.; Bao, L.P. Scale-dependent changes in species richness caused by invader competition. Ecol. Model. 2022, 469, 109996. [Google Scholar] [CrossRef]
- Sexton, J.P.; Montiel, J.; Shay, J.E.; Stephens, M.R.; Slatyer, R.A. Evolution of ecological niche breadth. Annu. Rev. Ecol. Evol. Syst. 2017, 48, 183–206. [Google Scholar] [CrossRef]
- Finch, J.; Walck, J.L.; Hidayati, S.N.; Kramer, A.T.; Lason, V.; Havens, K. Germination niche breadth varies inconsistently among three Asclepias congeners along a latitudinal gradient. Plant Biol. 2019, 21, 425–438. [Google Scholar] [CrossRef]
- Schuster, M.J.; Wragg, P.D.; Reich, P.B. Phenological niche overlap between invasive buckthorn (Rhamnus cathartica) and native woody species. For. Ecol. Manag. 2021, 498, 11956. [Google Scholar] [CrossRef]
- Stotz, G.C.; Pec, G.J.; Cahill, J.F. Is biotic resistance to invaders dependent upon local environmental conditions or primary productivity? A meta-analysis. Basic Appl. Ecol. 2016, 17, 377–387. [Google Scholar] [CrossRef]
- Welshofer, K.B.; Zarnetske, P.L.; Lany, N.K.; Read, Q.D. Short-term responses to warming vary between native vs. exotic species and with latitude in an early successional plant community. Oecologia 2018, 187, 333–342. [Google Scholar] [CrossRef]
- Cirtwill, A.R.; Stouffer, D.B.; Romanuk, T.N. Latitudinal gradients in biotic niche breadth vary across ecosystem types. Proc. R. Soc. B-Biol. Sci. 2015, 282, 20151589. [Google Scholar] [CrossRef]
- Vázquez, D.P.; Stevens, R.D. The latitudinal gradient in niche breadth: Concepts and evidence. Am. Nat. 2004, 164, E1–E19. [Google Scholar] [CrossRef]
- Alahuhta, J.; Virtala, A.; Hjort, J.; Ecke, F.; Johnson, L.B.; Sass, L.; Heino, J. Average niche breadths of species in lake macrophyte communities respond to ecological gradients variably in four regions on two continents. Oecologia 2017, 184, 219–235. [Google Scholar] [CrossRef]
- Wu, H.; Dong, S.J.; Rao, B.Q. Latitudinal trends in the structure, similarity and beta diversity of plant communities invaded by Alternanthera philoxeroides in heterogeneous habitats. Front. Plant Sci. 2022, 13, 1021337. [Google Scholar] [CrossRef]
- Wu, H.; Carrillo, J.; Ding, J.Q. Species diversity and environmental determinants of aquatic and terrestrial communities invaded by Alternanthera philoxeroides. Sci. Total Environ. 2017, 581, 666–675. [Google Scholar] [CrossRef]
- Lu, X.M.; Siemann, E.; Shao, X.; Wei, H.; Ding, J. Climate warming affects biological invasions by shifting interactions of plants and herbivores. Glob. Chang. Biol. 2013, 19, 2339–2347. [Google Scholar] [CrossRef]
- Wu, H.; Ismail, M.; Ding, J.Q. Global warming increases the interspecific competitiveness of the invasive plant alligator weed, Alternanthera philoxeroides. Sci. Total Environ. 2017, 575, 1415–1422. [Google Scholar] [CrossRef]
- Wang, T.; Hu, J.; Wang, R.; Liu, C.; Yu, D. Tolerance and resistance facilitate the invasion success of Alternanthera philoxeroides in disturbed habitats: A reconsideration of the disturbance hypothesis in the light of phenotypic variation. Environ. Exp. Bot. 2018, 153, 135–142. [Google Scholar] [CrossRef]
- Wu, H.; Carrillo, J.; Ding, J.Q. Invasion by alligator weed, Alternanthera philoxeroides, is associated with decreased species diversity across the latitudinal gradient in China. J. Plant Ecol. 2016, 93, 311–319. [Google Scholar] [CrossRef]
- Lu, X.M.; Siemann, E.; He, M.Y.; Wei, H.; Shao, X.; Ding, J. Climate warming increases biological control agent impact on a non-target species. Ecol. Lett. 2015, 18, 48–56. [Google Scholar] [CrossRef]
- Ma, Y.; Li, Q.; Pan, S.; Liu, C.; Han, M.; Brancelj, A. Niche and interspecific associations of Pseudoanabaena limnetica-Exploring the influencing factors of its succession stage. Ecol. Indic. 2022, 138, 108806. [Google Scholar] [CrossRef]
- Broennimann, O.; Fitzpatrick, M.C.; Pearman, P.B.; Petitpierre, B.; Pellissier, L.; Yoccoz, N.G.; Thuiller, W.; Fortin, M.-J.; Randin, C.; Zimmermann, N.E.; et al. Measuring ecological niche overlap from occurrence and spatial environmental data. Glob. Ecol. Biogeogr. 2012, 21, 481–497. [Google Scholar] [CrossRef]
- Martorell, C.; Almanza-Celis, C.A.; Pérez-García, E.A.; Sánchez-Ken, J.G. Co-existence in a species-rich grassland: Competition, facilitation and niche structure over a soil depth gradient. J. Veg. Sci. 2015, 26, 674–685. [Google Scholar] [CrossRef]
- Luo, L.; Kong, X.; Gao, Z.; Zheng, Y.; Yang, Y.; Li, X.; Yang, D.; Geng, Y.; Yang, Y. Comparative transcriptome analysis reveals ecological adaption of cold tolerance in northward invasion of Alternanthera philoxeroides. BMC Genom. 2020, 21, 532. [Google Scholar] [CrossRef] [PubMed]
- Stotz, G.C.; Salgado-Luarte, C.; Escobedo, V.M.; Valladares, F.; Gianoli, E. Global trends in phenotypic plasticity of plants. Ecol. Lett. 2021, 24, 2267–2281. [Google Scholar] [CrossRef]
- Müller, C.; Junker, R.R. Chemical phenotype as important and dynamic niche dimension of plants. New Phytol. 2022, 234, 1168–1174. [Google Scholar] [CrossRef]
- Lancaster, L.T. On the macroecological significance of eco-evolutionary dynamics: The range shift-niche breadth hypothesis. Philos. Trans. R. Soc. B-Biol. Sci. 2022, 377, 20210013. [Google Scholar] [CrossRef]
- Bar-Massada, A. Complex relationships between species niches and environmental heterogeneity affect species co-occurrence patterns in modelled and real communities. Proc. R. Soc. B-Biol. Sci. 2015, 282, 20150927. [Google Scholar] [CrossRef]
- Segura, A.M.; Calliari, D.; Kruk, C.; Conde, D.; Bonilla, S.; Fort, H. Emergent neutrality drives phytoplankton species coexistence. Proc. R. Soc. B-Biol. Sci. 2011, 278, 2355–2361. [Google Scholar] [CrossRef]
- Pennington, R.T.; Richardson, J.E.; Lavin, M. Insights into the historical construction of species-rich biomes from dated plant phylogenies, neutral ecological theory and phylogenetic community structure. New Phytol. 2006, 172, 605–616. [Google Scholar] [CrossRef]
- dos Santos Ribas, L.G.; de Cassia-Silva, C.; Petsch, D.K.; Silveira, M.L.; Lima-Ribeiro, M.S. The potential invasiveness of an aquatic macrophyte reflects founder effects from native niche. Biol. Invasions 2018, 20, 3347–3355. [Google Scholar] [CrossRef]
- He, D.; Chen, Y.; Zhao, K.; Cornelissen, J.H.C.; Chu, C. Intra- and interspecific trait variations reveal functional relationships between specific leaf area and soil niche within a subtropical forest. Ann. Bot. 2018, 121, 1173–1182. [Google Scholar] [CrossRef]
- van Boheemen, L.A.; Atwater, D.Z.; Hodgins, K.A. Rapid and repeated local adaptation to climate in an invasive plant. New Phytol. 2019, 222, 614–627. [Google Scholar] [CrossRef]
- Yang, Y.; Liu, M.; Pan, Y.; Huang, H.; Pan, X.; Sosa, A.; Hou, Y.; Zhu, Z.; Li, B. Rapid evolution of latitudinal clines in growth and defence of an invasive weed. New Phytol. 2021, 230, 845–856. [Google Scholar] [CrossRef]
- Morin, X.; Chuine, I. Niche breadth, competitive strength and range size of tree species: A trade-off based framework to understand species distribution. Ecol. Lett. 2006, 9, 185–195. [Google Scholar] [CrossRef]
- Foster, S.L.; Kharouba, H.M.; Kharouba, H.M. Testing the assumption of environmental equilibrium in an invasive plant species over a 130 year history. Ecography 2022, 2022, e06284. [Google Scholar] [CrossRef]
- Golivets, M.; Wallin, K.F. Neighbour tolerance, not suppression, provides competitive advantage to non-native plants. Ecol. Lett. 2018, 21, 745–759. [Google Scholar] [CrossRef]
- MacDougall, A.S.; Gilbert, B.; Levine, J.M. Plant invasions and the niche. J. Ecol. 2009, 97, 609–615. [Google Scholar] [CrossRef]
- Carscadden, K.A.; Emery, N.C.; Arnillas, C.A.; Cadotte, M.W.; Afkhami, M.E.; Gravel, D.; Livingstone, S.W.; Wiens, J.J. Niche breadth: Causes and consequences for ecology, evolution, and conservation. Q. Rev. Biol. 2020, 95, 179–214. [Google Scholar] [CrossRef]
- Anufriieva, E.V.; Shadrin, N.V. Extreme hydrological events destabilize aquatic ecosystems and open doors for alien species. Quat. Int. 2018, 475, 11–15. [Google Scholar] [CrossRef]
- Wu, H.; Ding, J.Q. Global change sharpens the double-edged sword of aquatic alien plants in China and beyond. Front. Plant Sci. 2019, 10, 787. [Google Scholar] [CrossRef]
- Kriticos, D.J.; Brunel, S. Assessing and managing the current and future pest risk from water hyacinth, (Eichhornia crassipes), an invasive aquatic plant threatening the environment and water security. PLoS ONE 2016, 11, e0120054. [Google Scholar] [CrossRef] [PubMed]
- Jawed, A.; Kar, P.; Verma, R.; Shukla, K.; Hemanth, P.; Thakur, V.K.; Pandey, L.M.; Gupta, R.K. Integration of biological control with engineered heterojunction nano-photocatalysts for sustainable and effective management of water hyacinth weed. J. Environ. Chem. Eng. 2022, 10, 106976. [Google Scholar] [CrossRef]
- Banerjee, A.K.; Liang, X.R.; Harms, N.E.; Tan, F.; Lin, Y.; Feng, H.; Wang, J.; Li, Q.; Jia, Y.; Lu, X.; et al. Spatio-temporal pattern of cross-continental invasion: Evidence of climatic niche shift and predicted range expansion provide management insights for smooth cordgrass. Ecol. Indic. 2022, 140, 109052. [Google Scholar] [CrossRef]
- Jia, X.; Pan, X.-Y.; Sosa, A.; Li, B.; Chen, J. Differentiation in growth and biomass allocation among three native Alternanthera philoxeroides varieties from Argentina. Plant Species Biol. 2010, 25, 85–92. [Google Scholar] [CrossRef]
- Jiang, P.; Xu, G.; He, Y.; Sun, T.; Liu, C.; Chen, C.; Zuo, R.; Sun, C. Complete chloroplast genome of Alternanthera philoxeroides by de novo sequencing. Mitochondrial DNA Part B-Resour. 2021, 6, 1826–1828. [Google Scholar] [CrossRef]



Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wu, H.; Dong, S.; Wang, Y.; Wang, L.; Rao, B. Niche Characteristics of Alternanthera philoxeroide-Invaded Plant Communities in Heterogeneous Habitats and Their Latitudinal Trends. Diversity 2023, 15, 651. https://doi.org/10.3390/d15050651
Wu H, Dong S, Wang Y, Wang L, Rao B. Niche Characteristics of Alternanthera philoxeroide-Invaded Plant Communities in Heterogeneous Habitats and Their Latitudinal Trends. Diversity. 2023; 15(5):651. https://doi.org/10.3390/d15050651
Chicago/Turabian StyleWu, Hao, Sijin Dong, Yanyan Wang, Li Wang, and Benqiang Rao. 2023. "Niche Characteristics of Alternanthera philoxeroide-Invaded Plant Communities in Heterogeneous Habitats and Their Latitudinal Trends" Diversity 15, no. 5: 651. https://doi.org/10.3390/d15050651