
Faunistic and Structural Changes in Shallow Coastal Benthic Communities of the Ebre Delta (NW Mediterranean Sea)

 , ,
, ,
Abstract
:1. Introduction
2. Materials and Methods
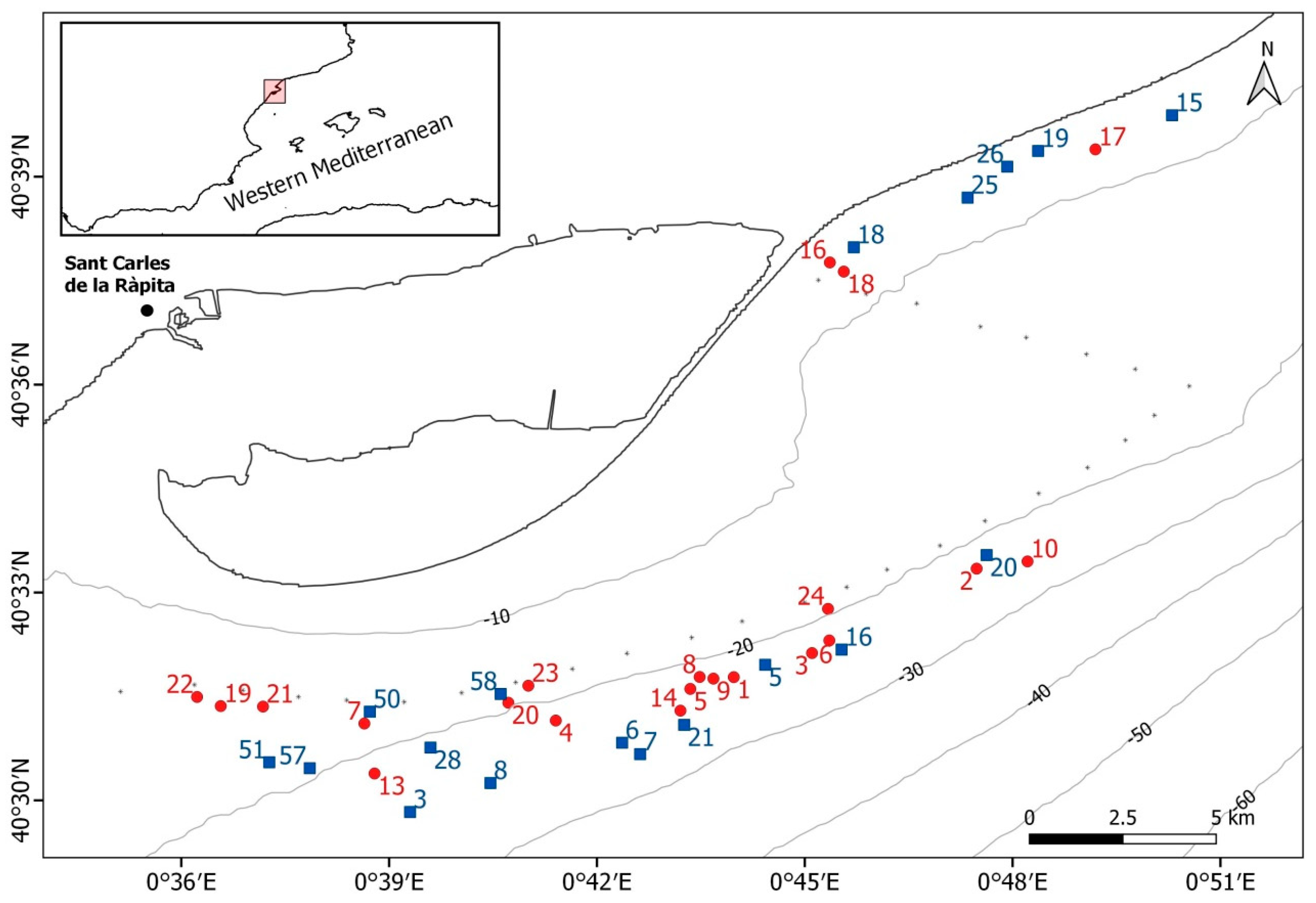
2.1. Study Area and Sampling Methods
2.2. Data Analysis
3. Results
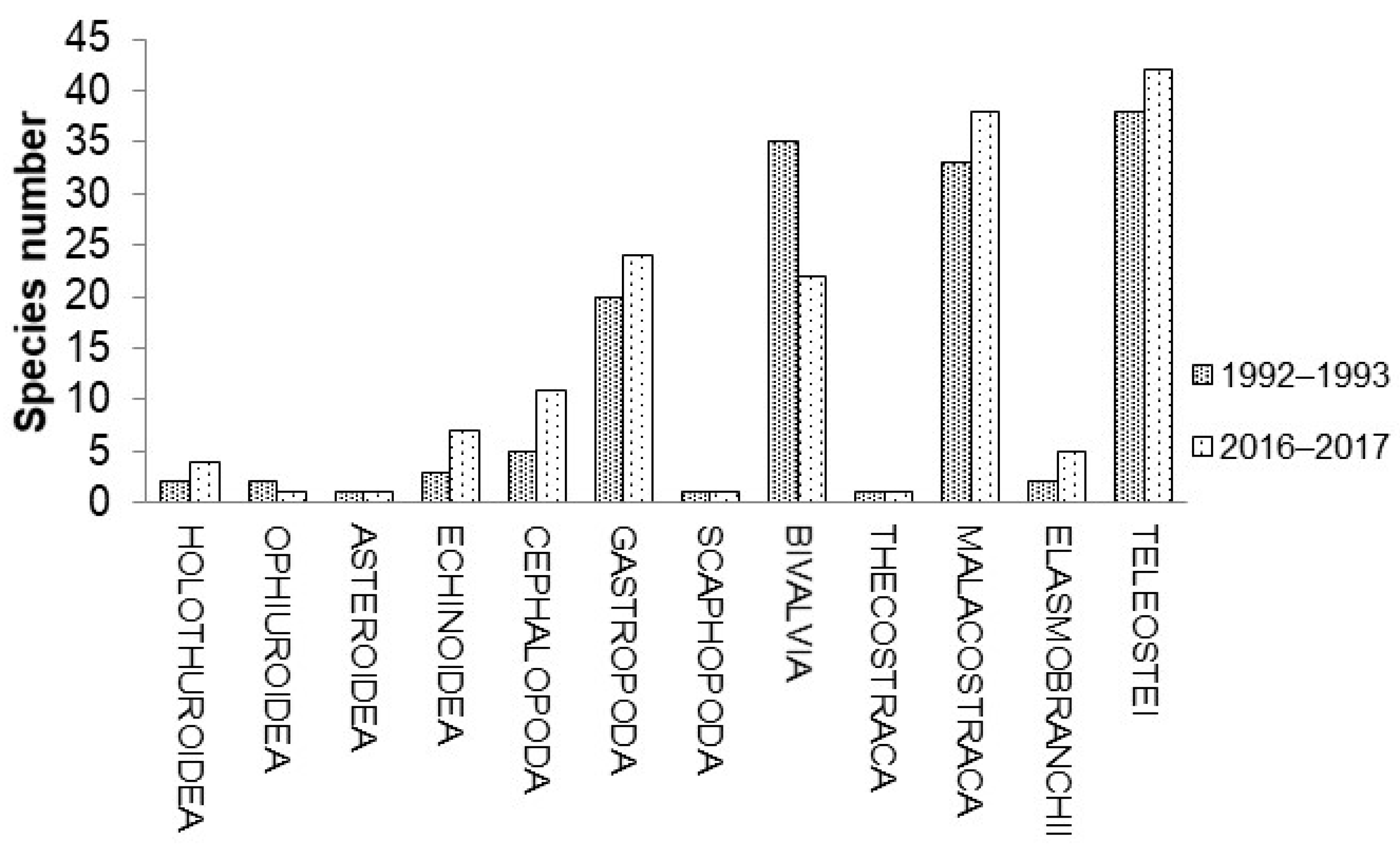
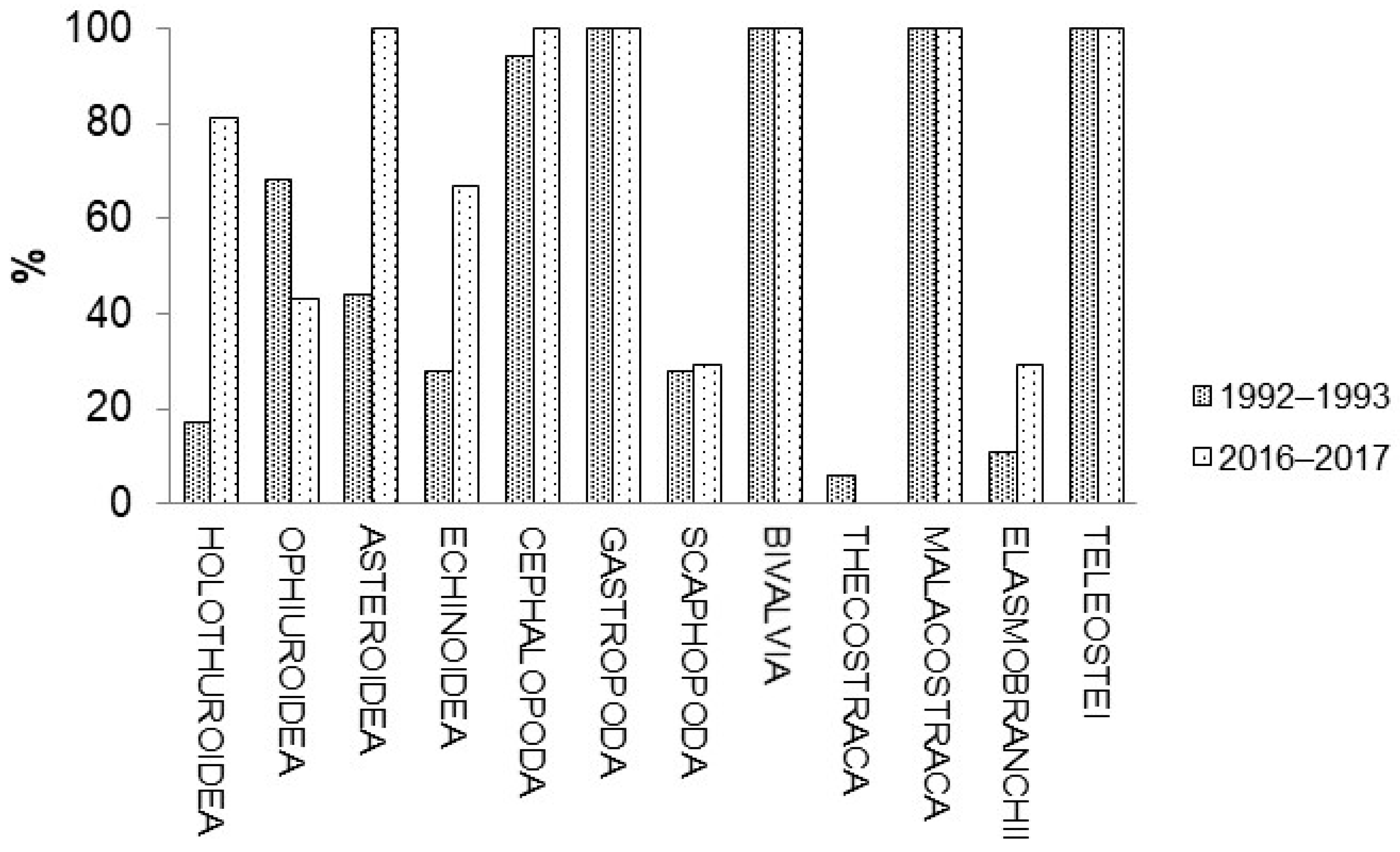
3.1. Faunistic Composition
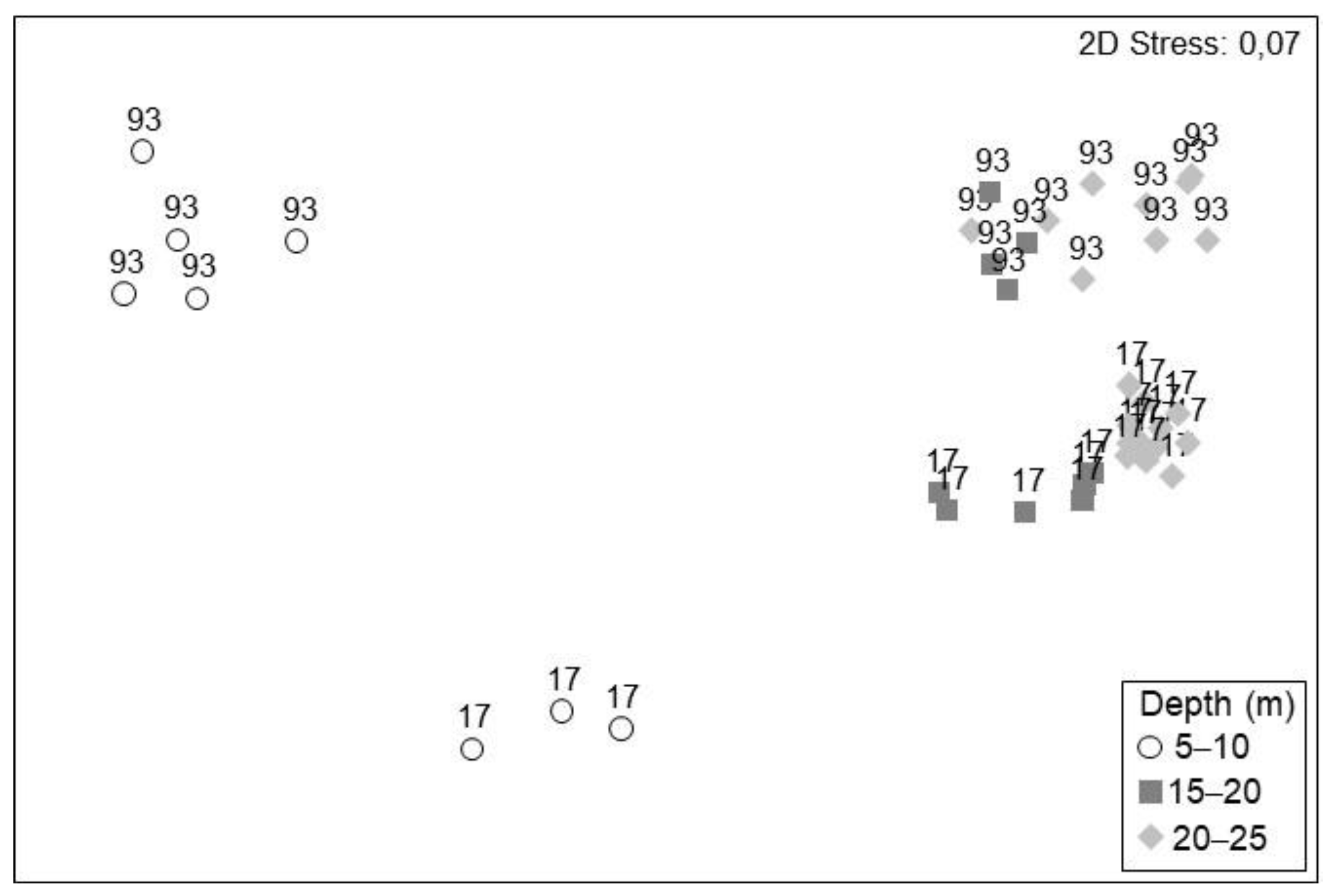
3.2. Characterization of Assemblages
3.3. Biodiversity
3.4. Biomass
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Coll, M.; Piroddi, C.; Steenbeek, J.; Kaschner, K.; Ben Rais Lasram, F.; Aguzzi, J.; Ballesteros, E.; Bianchi, C.N.; Corbera, J.; Dailianis, T.; et al. The biodiversity of the Mediterranean Sea: Estimates, patterns, and threats. PLoS ONE 2010, 5, e11842. [Google Scholar] [CrossRef] [PubMed]
- Dauvin, J.C. Paradox of estuarine quality: Benthic indicators and indices, consensus or debate for the future. Mar. Pollut. Bull. 2007, 55, 271–281. [Google Scholar] [CrossRef]
- Dauvin, J.C.; Gomez Gesteira, J.L.; Salvande Fraga, M. Taxonomic sufficiency: An overview of its use in the monitoring of sublittoral benthic communities after oil spills. Mar. Pollut. Bull. 2003, 46, 552–555. [Google Scholar] [CrossRef]
- Labrune, C.; Grémare, A.; Amouroux, J.M.; Sardá, R.; Gil, J.; Taboada, S. Structure and diversity of shallow soft bottom benthic macrofauna in the Gulf of Lions (NW Mediterranean). Helgol. Mar. Res. 2008, 62, 201–214. [Google Scholar] [CrossRef]
- Labrune, C.; Gauthier, O.; Conde, A.; Grall, J.; Blomqvist, M.; Bernard, G.; Gallon, G.; Dannheim, J.; van Hoey, G.; Grémare, A. A general-purpose biotic index to measure changes in benthic habitat quality across several pressure gradients. J. Mar. Sci. Eng. 2021, 9, 654. [Google Scholar] [CrossRef]
- Pinedo, S.; Jordana, E.; Manzanera, M.; Ballesteros, E. Using MEDOCC (MEDiterranean OCCidental) index to evaluate the ecological status of Catalan coastal waters (Northwestern Mediterranean Sea) over time and depths. Handb. Environ. Chem. 2016, 43, 201–225. [Google Scholar]
- Rufino, M.M.; Pereira, A.M.; Pereira, F.; Moura, P.; Vasconcelos, P.; Gaspar, M.B. Habitat structure shaping megabenthic communities inhabiting subtidal soft bottoms along the Algarve coast (Portugal). Hydrobiologia 2017, 784, 249–264. [Google Scholar] [CrossRef]
- Beukema, J.J.; Dekker, R.; Drent, J. Parallel changes of Limecola (Macoma) balthica populations in the Dutch Wadden Sea. Mar. Ecol. Prog. Ser. 2017, 585, 71–79. [Google Scholar] [CrossRef]
- Kröncke, I.; Zeiss, B.; Rensing, C. Long-term variability in macrofauna species composition off the island of Norderney (East Frisia, Germany) in relation to changes in climatic and environmental conditions. Senckenb. Marit. 2001, 31, 65–82. [Google Scholar] [CrossRef]
- Sagarin, R.D.; Barry, J.P.; Gilman, S.E.; Baxter, C.H. Climate related changes in an intertidal community over short and long time scales. Ecol. Monogr. 1999, 69, 465–490. [Google Scholar] [CrossRef]
- Lamela-Silvarrey, C.; Fernández, C.; Anadón, R.; Arrontes, J. Fucoid assemblages on the north coast of Spain: Past and present (1977–2007). Bot. Mar. 2012, 55, 199–207. [Google Scholar] [CrossRef]
- Grémare, A.; Amouroux, J.M.; Vétion, G. Long-term comparison of macrobenthos within the soft bottoms of the Bay of Banyuls-sur-mer (northwestern Mediterranean Sea). J. Sea Res. 1998, 40, 281–302. [Google Scholar] [CrossRef]
- Labrune, C.; Grémare, A.; Guizien, K.; Amouroux, J.M. Long-term comparison of soft bottom macrobenthos in the Bay of Banyuls-sur-Mer (north-western Mediterranean Sea): A reappraisal. J. Sea Res. 2007, 58, 125–143. [Google Scholar] [CrossRef]
- Brind’Amour, A.; Laffargue, P.; Morin, J.; Vaz, S.; Foveau, A.; Le Bris, H. Morphospecies and taxonomic sufficiency of benthic megafauna in scientific bottom trawl surveys. Cont. Shelf Res. 2014, 72, 1–9. [Google Scholar] [CrossRef]
- Bertrand, J.A.; De Sola, L.G.; Papaconstantinou, C.; Relini, G.; Souplet, A. The general specifications of the MEDITS surveys. Sci. Mar. 2002, 66, 9–17. [Google Scholar] [CrossRef]
- Spedicato, M.T.; Massutí, E.; Mérigot, B.; Tserpes, G.; Jadaud, A.; Relini, G. The MEDITS trawl survey specifications in an ecosystem approach to fishery management. Sci. Mar. 2019, 83, 9–20. [Google Scholar] [CrossRef]
- Pubill, E.; Abelló, P.; Ramón, M.; Baeta, M. Faunistic assemblages of a sublittoral coarse sand habitat of the northwestern Mediterranean. Sci. Mar. 2011, 75, 189–196. [Google Scholar]
- Santelli, A.; Cvitković, I.; Despalatović, M.; Fabi, G.; Grati, F.; Marčeta, B.; Punzo, E.; Raicevich, S.; Strafella, P.; Spagnolo, A.; et al. Spatial persistence of megazoobenthic assemblages in the Adriatic Sea. Mar. Ecol. Prog. Ser. 2017, 566, 31–48. [Google Scholar] [CrossRef]
- Raaijmakers, R.; Krywkow, J.; Van der Veen, A. Flood risk perceptions and spatial multi-criteria analysis: An exploratory research for hazard mitigation. Nat. Hazards 2008, 46, 307–322. [Google Scholar] [CrossRef]
- Stratmann, T.; van Oevelen, D.; Martínez Arbizu, P.; Wei, C.-L.; Liao, J.-X.; Cusson, M.; Scrosati, R.A.; Archambault, P.; Snelgrove, P.V.R.; Ramey-Balci, P.A.; et al. The BenBioDen database, a global database for meio-, macro- and megabenthic biomass and densities. Sci. Data 2020, 7, 206. [Google Scholar] [CrossRef]
- Lyons, D.A.; Benedetti-Cecchi, L.; Frid, C.L.J.; Vinebrooke, R.D. Modifiers of impacts on marine ecosystems: Disturbance regimes, multiple stressors and receiving environments. In Marine Ecosystems: Human Impacts on Biodiversity, Functioning and Services; Crowe, T.P., Frid, C.L.J., Eds.; Cambridge University Press: Cambridge, UK, 2015; pp. 73–110. [Google Scholar]
- Borja, A.; Elliott, M.; Carstensen, J.; Heiskanen, A.-S.; Bund, W. Marine management—Towards an integrated implementation of the European Marine Strategy Framework and the Water Framework Directives. Mar. Pollut. Bull. 2010, 60, 2175–2186. [Google Scholar] [CrossRef] [PubMed]
- Jennings, S.; Smith, A.D.; Fulton, E.A.; Smith, D.C. The ecosystem approach to fisheries: Management at the dynamic interface between biodiversity conservation and sustainable use. Ann. N. Y. Acad. Sci. 2014, 1322, 48–60. [Google Scholar] [CrossRef]
- Palanques, A.; Puig, P.; Guillén, J.; Jiménez, J.; Gracia, V.; Sánchez-Arcilla, A.; Madsen, O. Near-bottom suspended sediment fluxes on the microtidal low-energy Ebro continental shelf (NW Mediterranean). Cont. Shelf Res. 2002, 22, 285–303. [Google Scholar] [CrossRef]
- Guillen, J.; Palanques, A. A shoreface zonation in the Ebro Delta based on grain size distribution. J. Coast. Res. 1997, 13, 867–878. [Google Scholar]
- Palanques, A.; Guillén, J. Coastal changes in the Ebro delta: Natural and human factors. J. Coast. Conserv. 1998, 4, 17–26. [Google Scholar]
- Martín, P.; Sánchez, P.; Ramón, M. Population structure and exploitation of Bolinus brandaris (Mollusca: Gastropoda) off the Catalan coast (northwestern Mediterranean). Fish. Res. 1995, 23, 319–331. [Google Scholar] [CrossRef]
- Clarke, K.R.; Warwick, R.M. Change in Marine Communities: An Approach to Statistical Analysis and Interpretation; PRIMER-E, Ed.; Plymouth Marine Laboratory: Plymouth, UK, 2001. [Google Scholar]
- Anderson, M.J.; Gorley, R.N.; Clarke, K.R. PERMANOVA+for PRIMER: Guide to Software and Statistical Methods; PRIMER-E: Plymouth, UK, 2008; p. 218. [Google Scholar]
- Marina, P.; Rueda, J.L.; Urra, J.; Salas, C.; Gofas, S.; García Raso, J.E.; Moya, F.; García, T.; López-González, N.; Laiz-Carrión, R.; et al. Sublittoral soft bottom assemblages within a Marine Protected Area of the northern Alboran Sea. J. Mar. Biolog. Assoc. UK 2015, 95, 871–884. [Google Scholar] [CrossRef]
- Urra, J.; Gofas, S.; Rueda, J.L.; Marina, P. Molluskan assemblages in littoral soft bottoms of the Alboran Sea (Western Mediterranean Sea). Mar. Biol. Res. 2011, 7, 27–42. [Google Scholar] [CrossRef]
- Bedulli, D.; Bassignani, F.; Bruschi, A. Use of biodiversity hotspots for conservation of marine molluscs: A regional approach. Med. Mar. Sci. 2002, 3, 113–121. [Google Scholar] [CrossRef]
- Gladstone, W. The potential value of indicator groups in the selection of marine reserves. Biol. Conserv. 2002, 104, 211–220. [Google Scholar] [CrossRef]
- Nerlović, V.; Doğan, A.; Hrs-Brenko, M. Response to oxygen deficiency (depletion): Bivalve assemblages as an indicator of ecosystem instability in the northern Adriatic Sea. Biologia 2011, 66, 1114–1126. [Google Scholar] [CrossRef]
- Borja, A.; Franco, J.; Pérez, V. A marine biotic index to establish the ecological quality of soft-bottom benthos within European estuarine and coastal environments. Mar. Pollut. Bull. 2000, 40, 1100–1114. [Google Scholar] [CrossRef]
- Baeta, M.; Ramón, M.; Galimany, E. Decline of a Callista chione (Bivalvia: Veneridae) bed in the Maresme coast (northwestern Mediterranean Sea). Ocean Coast. Manag. 2014, 93, 15–25. [Google Scholar] [CrossRef]
- Dang, C.; de Montaudouin, X.; Gam, M.; Paroissin, C.; Bru, N.; Caill-Milly, N. The Manila clam population in Arcachon Bay (SW France): Can it be kept sustainable? J. Shellfish Res. 2010, 63, 108–118. [Google Scholar] [CrossRef]
- Delgado, M.; Silva, L.; Gómez, S.; Masferrer, E.; Cojan, M.; Gaspar, M.B. Population and production parameters of the wedge clam Donax trunculus (Linnaeus, 1758) in intertidal areas on the southwest Spanish coast: Considerations in relation to protected areas. Fish. Res. 2017, 193, 232–241. [Google Scholar] [CrossRef]
- Peharda, M.; Ezgeta Balic, D.; Radman, M.; Sinjkević, N. Age, growth and population structure of Acanthocardia tuberculata (Bivalvia: Cardiidae) in the eastern Adriatic Sea. Sci. Mar. 2012, 76, 59–66. [Google Scholar]
- Ramón, M.; Cano, J.; Peña, J.B.; Campos, M.J. Current status and perspectives of mollusc (bivalves and gastropods) culture in the Spanish Mediterranean. Bol. Inst. Esp. Oceanogr. 2005, 21, 361–373. [Google Scholar]
- Demestre, M.; Sartor, P.; Garcia-de-Vinuesa, A. Ecological importance of survival of unwanted invertebrates discarded in different NW Mediterranean trawl fisheries. Sci. Mar. 2018, 82, 189–198. [Google Scholar] [CrossRef]
- Cloern, J.E.; Abreu, P.C.; Carstensen, J.; Chauvaud, L.; Elmgren, R.; Grall, J.; Greening, H.; Johansson, J.O.R.; Kahru, M.; Sherwood, E.T.; et al. Human activities and climate variability drive fast-paced change across the world’s estuarine–coastal ecosystems. Glob. Chang. Biol. 2016, 22, 513–529. [Google Scholar] [CrossRef]
- Collie, J.S.; Hall, S.J.; Kaiser, M.J.; Poiner, I.R. A quantitative analysis of fishing impacts on shelf-sea benthos. J. Anim. Ecol. 2000, 69, 785–798. [Google Scholar] [CrossRef]
- Kaiser, M.J.; Clarke, K.R.; Hinz, H.; Austen, M.C.V.; Somerfield, P.J.; Karakassis, I. Global analysis and prediction of the response of benthic biota and habitats to fishing. Mar. Ecol. Prog. Ser. 2006, 311, 1–14. [Google Scholar] [CrossRef]
- Thrush, S.F.; Dayton, P.K. Disturbance to marine benthic habitats by trawling and dredging; implications for marine biodiversity. Annu. Rev. Ecol. Evol. Syst. 2002, 33, 449–473. [Google Scholar] [CrossRef]
- Lambert, G.I.; Jennings, S.; Kaiser, M.J.; Hinz, H.; Hiddink, J.G. Quantification and prediction of the impact of fishing on epifaunal communities. Mar. Ecol. Prog. Ser. 2011, 430, 71–86. [Google Scholar] [CrossRef]
- Hunt, H.L.; Scheibling, R.E. Role of early post-settlement mortality in recruitment of benthic marine invertebrates. Mar. Ecol. Prog. Ser. 1997, 155, 269–301. [Google Scholar] [CrossRef]
- Morris, A.W.; Howarth, M.J. Bed stress induced sediment resuspension. Cont. Shelf Res. 1998, 18, 1203–1213. [Google Scholar] [CrossRef]
- Thistle, D. Natural physical disturbances and communities of marine soft bottoms. Mar. Ecol. Prog. Ser. 1981, 6, 223–228. [Google Scholar] [CrossRef]
- Jennings, S.; Kaiser, M.J. The effects of fishing on marine ecosystems. Adv. Mar. Biol. 1998, 34, 201–352. [Google Scholar]
- Kaiser, M.J. Significance of bottom-fishing disturbance. Conserv. Biol. 1998, 12, 1230–1235. [Google Scholar] [CrossRef]
- De Juan, S.; Thrush, S.F.; Demestre, M. Functional changes as indicators of trawling disturbance on a benthic community located in a fishing ground (NW Mediterranean Sea). Mar. Ecol. Prog. Ser. 2007, 334, 117–129. [Google Scholar] [CrossRef]
- DARP. ORDRE ARP/114/2018, de 6 de Juliol, per la Qual S’aprova el Pla de Gestió per a les Embarcacions de Marisqueig Amb Dragues per a Embarcació; Departament d’Agricultura, Ramaderia, Pesca i Alimentació, Diari Oficial de la Generalitat de Catalunya: Barcelona, Spain, 2018; p. 7. [Google Scholar]
- Rodríguez-Santalla, I.; Navarro, N. Main threats in Mediterranean coastal wetlands. The Ebro Delta case. J. Mar. Sci. Eng. 2021, 9, 1190. [Google Scholar] [CrossRef]
- MedECC. Climate and Environmental Change in the Mediterranean Basin—Current Situation and Risks for the Future. First Mediterranean Assessment report; Cramer, W., Guiot, J., Marini, K., Eds.; Union for the Mediterranean, Plan Bleu, UNEP/MAP: Marseille, France, 2020; p. 632. [Google Scholar]
- Anadón, R.A.; Danovaro, R.; Dippner, J.W.; Drinkwater, K.F.; Hawkins, S.J.; O’Sullivan, G.; Oguz, T.; Reid, P.C. Impacts of Climate Change on the European Marine and Coastal Environment; European Science Foundation: Strasbourg, France, 2007; pp. 1–84. [Google Scholar]
- Prado, P.; Peñas, A.; Ibáñez, C.; Cabanes, P.; Jornet, L.; Álvarez, N.; Caiola, N. Prey size and species preferences in the invasive blue crab, Callinectes sapidus: Potential effects in marine and freshwater ecosystems. Estuar. Coast. Shelf Sci. 2020, 245, 106997. [Google Scholar] [CrossRef]
- Amores, A.; Marcos, M.; Carrió, D.S.; Gómez-Pujol, L. Coastal impacts of Storm Gloria (January 2020) over the north-western Mediterranean. Nat. Hazards Earth Syst. Sci. 2020, 20, 1955–1968. [Google Scholar] [CrossRef]
- Abelló, P.; Carbonell, A.; Torres, P. Biogeography of epibenthic crustaceans on the shelf and upper slope off the Iberian Peninsula Mediterranean coasts: Implications for the establishment of natural management areas. Sci. Mar. 2002, 66, 183–198. [Google Scholar] [CrossRef]
- Abelló, P.; Valladares, F.J.; Castellón, A. Analysis of the structure of decapod crustacean assemblages off the Catalan coast (North West Mediterranean). Mar. Biol. 1988, 98, 39–49. [Google Scholar] [CrossRef]
- DeLaHoz, M.V.; Sardà, F.; Coll, M.; Sáez, R.; Mechó, A.; Oliva, F.; Ballesteros, M.; Palomera, I. Biodiversity patterns of megabenthic non-crustacean invertebrates from an exploited ecosystem of the Northwestern Mediterranean Sea. Reg. Stud. Mar. Sci. 2018, 19, 47–68. [Google Scholar] [CrossRef]
- Cartes, J.; Sardà, F. Zonation of deep-sea decapod fauna in the Catalan Sea (Western Mediterranean). Mar. Ecol. Prog. Ser. 1993, 94, 27–34. [Google Scholar] [CrossRef]
- Ramón, M.; Abelló, P.; Ordines, F.; Massutí, E. Deep epibenthic communities in two contrasting areas of the Balearic Islands (western Mediterranean). J. Mar. Syst. 2014, 138, 182–193. [Google Scholar] [CrossRef]
- Jiménez, J.A.; Sánchez-Arcilla, A.; Bou, J.; Ortiz, M.A. Analysing short-term shoreline changes along the Ebro Delta (Spain) using aerial photographs. J. Coast. Res. 1997, 13, 1256–1266. [Google Scholar]
- Rufino, M.M.; Gaspar, M.B.; Pereira, A.M.; Maynou, F.; Monteiro, C.C. Ecology of megabenthic bivalve communities from sandy beaches on the south coast of Portugal. Sci. Mar. 2010, 74, 163–178. [Google Scholar] [CrossRef]
- Collen, B.; Böhm, M.; Kemp, R.; Baillie, J. Spineless: Status and Trends of the World’s Invertebrates; Zoological Society of London: London, UK, 2012; p. 86. [Google Scholar]
- Dirzo, R.; Young, H.S.; Galetti, M.; Ceballos, G.; Isaac, N.J.B.; Collen, B. Defaunation in the Anthropocene. Science 2014, 345, 401–406. [Google Scholar] [CrossRef]
- Eisenhauer, N.; Bonn, A.; Guerra, C.A. Recognizing the quiet extinction of invertebrates. Nat. Commun. 2019, 10, 50. [Google Scholar] [CrossRef] [PubMed]
- Hallmann, C.A.; Sorg, M.; Jongejans, E.; Siepel, H.; Hofland, N.; Schwan, H.; Stenmans, W.; Müller, A.; Sumser, H.; Hörren, T.; et al. More than 75 percent decline over 27 years in total flying insect biomass in protected areas. PLoS ONE 2017, 12, e0185809. [Google Scholar] [CrossRef]
- Ceballos, G.; Ehrlich, P.R.; Dirzo, R. Biological annihilation via the ongoing sixth mass extinction signaled by vertebrate population losses and declines. Proc. Natl. Acad. Sci. USA 2017, 114, E6089–E6096. [Google Scholar] [CrossRef] [PubMed]
- Rife, G.S. Ecosystem services provided by benthic macroinvertebrate assemblages in marine coastal zones. In Ecosystem Services and Global Ecology; Hufnagel, L., Ed.; InTech Open: Rijeka, Croatia, 2018; pp. 61–79. [Google Scholar]
- Dame, R.F. Ecology of Marine Bivalves: An Ecosystem Approach; CRC Press: Boca Raton, FL, USA, 1996; p. 272. [Google Scholar]
- Newell, R.I.E. Ecosystem influences of natural and cultivated populations of suspension-feeding bivalve molluscs: A review. J. Shellfish Res. 2004, 23, 51–61. [Google Scholar]
- IPBES. Regional Assessment Report on Biodiversity and Ecosystem Services for Europe and Central Asia; IPBES: Bonn, Germany, 2018; p. 48. [Google Scholar]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Overall PERMANOVA | |||||
|---|---|---|---|---|---|
| Source of Variation | df | SS | MS | Pseudo-F | p (Perm) |
| Depth strata | 2 | 28,469 | 14,234 | 22.387 | 0.0001 * |
| Year | 1 | 10,077 | 10,077 | 15.848 | 0.0001 * |
| Depth strata x Year | 2 | 9805.8 | 4902.9 | 7.711 | 0.0001 * |
| Residual | 33 | 20,983 | 635.8 | - | - |
| Pairwise test for pairs of levels of factor ‘Depth strata’ | |||||
| p (Perm) | p (Perm) | ||||
| Contrast | 1992–1933 | 2016–2017 | |||
| 0–5 m vs. 15–20 m | 0.0078 * | 0.0122 * | |||
| 0–5 m vs. 20–25 m | 0.0004 * | 0.002 * | |||
| 15–20 m vs. 20–25 m | 0.0014 * | 0.0001 * | |||
| Pairwise test for pairs of levels of factor ‘Year’ | |||||
| p (Perm) | p (Perm) | p (Perm) | |||
| Contrast | 0–5 m | 15–20 m | 20–25 m | ||
| 93 vs. 17 | 0.0177 * | 0.005 * | 0.0001 * | ||
| 5–10 m | 5–10 m | ||||
|---|---|---|---|---|---|
| 1992–1993 | 2016–2017 | ||||
| Average similarity: 51.10 | Average similarity: 64.20 | ||||
| Species | Av. D | Contr% | Species | Av. D | Contr% |
| Chamelea gallina | 3252.11 | 27.67 | Liocarcinus vernalis | 285.65 | 22.29 |
| Spisula subtruncata | 5624.11 | 18.19 | Mactra stultorum | 214.6 | 12.05 |
| Donax semistriatus | 1183.83 | 8.16 | Diogenes pugilator | 145.8 | 9.09 |
| Liocarcinus vernalis | 916.45 | 7.7 | Chamelea gallina | 100.27 | 6.43 |
| Diogenes pugilator | 849.58 | 7.66 | Penaeus kerathurus | 75.17 | 6.41 |
| Acanthocardia tuberculata | 758.6 | 7.33 | Spisula subtruncata | 112.73 | 6.19 |
| Tritia mutabilis | 711.23 | 6.05 | Tritia mutabilis | 78.91 | 5.95 |
| Cum% 82.75 | Cum% 68.41 | ||||
| 15–20 m | 15–20 m | ||||
| 1992–1993 | 2016–2017 | ||||
| Average similarity: 59.40 | Average similarity: 61.72 | ||||
| Species | Av. D | Contr% | Species | Av. D | Contr% |
| Turritellinella tricarinata | 533.54 | 23.66 | Liocarcinus depurator | 136.25 | 13.61 |
| Spisula subtruncata | 333.8 | 16.07 | Bolinus brandaris | 97.04 | 9.8 |
| Bolinus brandaris | 277.27 | 13.35 | Astropecten irregularis | 69.13 | 7.5 |
| Aporrhais pespelecani | 130.58 | 6.26 | Goneplax rhomboides | 57.7 | 6.77 |
| Bivetiella cancellata | 124.17 | 5.99 | Aporrhais pespelecani | 65.83 | 6.09 |
| Goneplax rhomboides | 83.47 | 4.4 | Penaeus kerathurus | 46 | 5.87 |
| Astropecten irregularis | 68 | 4.23 | Medorippe lanata | 34.24 | 4.4 |
| Cum% 73.96 | Cum% 54.04 | ||||
| 20–25 m | 20–25 m | ||||
| 1992–1993 | 2016–2017 | ||||
| Average similarity: 51.58 | Average similarity: 70.66 | ||||
| Species | Av. D | Contr% | Species | Av. D | Contr% |
| Turritellinella tricarinata | 748.07 | 28.07 | Bolinus brandaris | 354 | 33.5 |
| Bolinus brandaris | 145.46 | 14.29 | Turritellinella tricarinata | 148.96 | 10.89 |
| Goneplax rhomboides | 94.78 | 7.85 | Astropecten irregularis | 78.48 | 7.84 |
| Medorippe lanata | 63.38 | 5.95 | Medorippe lanata | 43.4 | 4.59 |
| Liocarcinus depurator | 75.15 | 5.33 | Arnoglossus laterna | 36.98 | 3.75 |
| Bivetiella cancellata | 64.14 | 4.98 | Alpheus glaber | 38.82 | 3.3 |
| Aporrhais pespelecani | 54.85 | 3.48 | Goneplax rhomboides | 37.62 | 3.24 |
| Cum% 69.94 | Cum% 67.12 |
| Groups 5–10 m | |||
|---|---|---|---|
| Average dissimilarity = 98.30 | |||
| 1992–1993 | 2016–2017 | ||
| Species | Av. D | Av. D | Contr% |
| Spisula subtruncata | 5624.11 | 112.73 | 22.21 |
| Chamelea gallina | 3252.11 | 100.27 | 20.36 |
| Donax semistriatus | 1183.83 | 9.61 | 8.27 |
| Acanthocardia tuberculata | 758.6 | 0 | 5.2 |
| Ophiura ophiura | 1326.38 | 88.78 | 5.17 |
| Tritia mutabilis | 711.23 | 78.91 | 4.47 |
| Diogenes pugilator | 849.58 | 145.8 | 4.38 |
| Liocarcinus vernalis | 916.45 | 285.65 | 3.77 |
| Mactra stultorum | 609.88 | 214.6 | 2.92 |
| Cum.% 76.76 | |||
| Groups 15–20 m | |||
| Average dissimilarity = 80.10 | |||
| 1992–1993 | 2016–2017 | ||
| Species | Av. D | Av. D | Contr% |
| Turritellinella tricarinata | 533.54 | 51.68 | 19.55 |
| Spisula subtruncata | 333.8 | 44.65 | 12.54 |
| Bolinus brandaris | 277.27 | 97.04 | 7.84 |
| Liocarcinus depurator | 23.98 | 136.25 | 4.81 |
| Bivetiella cancellata | 124.17 | 10.4 | 4.63 |
| Liocarcinus vernalis | 4.8 | 87.92 | 3.38 |
| Aporrhais pespelecani | 130.58 | 65.83 | 3 |
| Paguristes eremita | 48.14 | 1.06 | 2.16 |
| Cum.% 57.91 | |||
| Groups 20–25 m | |||
| Average dissimilarity = 59.95 | |||
| 1992–1993 | 2016–2017 | ||
| Species | Av. D | Av. D | Contr% |
| Turritellinella tricarinata | 748.07 | 148.96 | 29.84 |
| Bolinus brandaris | 145.46 | 354 | 11.49 |
| Astropecten irregularis | 9.31 | 78.48 | 3.81 |
| Goneplax rhomboides | 94.78 | 37.62 | 3.48 |
| Liocarcinus depurator | 75.15 | 14.01 | 3.45 |
| Bivetiella cancellata | 64.14 | 16.78 | 2.75 |
| Aporrhais pespelecani | 54.85 | 41.23 | 2.21 |
| Cum.% 55.75 | |||
| 5–10 m | 15–20 m | 20–25 m | |||||||
|---|---|---|---|---|---|---|---|---|---|
| CLASS | 1992–1993 | 2016–2017 | % | 1992–1993 | 2016–2017 | % | 1992–1993 | 2016–2017 | % |
| Holothuroidea | 2.2 | 4.4 | 0.2 | −94.8 | 2.6 | 0.2 | −93.6 | ||
| Ophiuroidea | 7912.7 | 11.7 | −99.9 | 2.2 | 0.8 | −62.5 | 0.3 | 0.1 | −73.1 |
| Asteroidea | 46.3 | 28.6 | −38.3 | 10.7 | 18.2 | 70.4 | 8.7 | 22 | 152.3 |
| Echinoidea | 1.1 | 2.3 | 101.6 | 2.6 | 0.5 | −81.6 | |||
| Cephalopoda | 999.5 | 7.4 | −99.3 | 16.2 | 2.9 | −82 | 8.6 | 8.4 | −2.2 |
| Gastropoda | 392.2 | 5.9 | −98.5 | 91.5 | 16.4 | −82.1 | 177 | 77.4 | −56.3 |
| Bivalvia | 11,692.9 | 22.7 | −99.8 | 9.2 | 2 | −78.3 | 1.7 | 1.1 | −36.3 |
| Malacostraca | 799.6 | 45 | −94.4 | 13.6 | 15.1 | 10.8 | 12.8 | 3.3 | −74.2 |
| Elasmobranchii | 140.4 | 5 | −96.5 | 0.1 | 0.6 | 0.1 | −82.4 | ||
| Teleostei | 178.8 | 2.7 | −98.5 | 6.6 | 2.3 | −65.2 | 3.7 | 1.8 | −50.5 |
| Total | 121,672 | 856.4 | −99.3 | 1238.0 | 413.5 | −66.6 | 1607.4 | 759.9 | −52.7 |
| Source of Variation | df | SS | MS | F | p |
|---|---|---|---|---|---|
| Species richness | |||||
| Year | 1 | 530.93 | 530.93 | 12.105 | <0.05 |
| Depth | 2 | 500.103 | 250.05 | 5.701 | <0.05 |
| Year x Depth | 2 | 132.68 | 66.34 | 1.513 | >0.05 |
| Error | 33 | 1447.45 | 43.86 | ||
| Shannon–Wiener | |||||
| Year | 1 | 1.488 | 1.488 | 5.203 | < 0.05 |
| Depth | 2 | 2.830 | 1.415 | 4.948 | < 0.05 |
| Year x Depth | 2 | 1.779 | 0.890 | 3.110 | <0.05 |
| Error | 33 | 9.437 | 0.286 | ||
| Evenness | |||||
| Year | 1 | 0.071 | 0.071 | 3.519 | > 0.05 |
| Depth | 2 | 0.199 | 0.099 | 4.941 | < 0.05 |
| Year x Depth | 2 | 0.114 | 0.057 | 2.830 | > 0.05 |
| Error | 33 | 0.663 | 0.020 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ramón, M.; Marco-Herrero, E.; Galimany, E.; Recasens, L.; Abelló, P. Faunistic and Structural Changes in Shallow Coastal Benthic Communities of the Ebre Delta (NW Mediterranean Sea). Diversity 2023, 15, 623. https://doi.org/10.3390/d15050623
Ramón M, Marco-Herrero E, Galimany E, Recasens L, Abelló P. Faunistic and Structural Changes in Shallow Coastal Benthic Communities of the Ebre Delta (NW Mediterranean Sea). Diversity. 2023; 15(5):623. https://doi.org/10.3390/d15050623
Chicago/Turabian StyleRamón, Montserrat, Elena Marco-Herrero, Eve Galimany, Laura Recasens, and Pere Abelló. 2023. "Faunistic and Structural Changes in Shallow Coastal Benthic Communities of the Ebre Delta (NW Mediterranean Sea)" Diversity 15, no. 5: 623. https://doi.org/10.3390/d15050623