
Crambe insularis sp. nov. (Crambeidae: Poecilosclerida) a New Crambeid from the Eastern Tropical North Pacific: Morphological, Molecular and Ontogenetic Approach


Abstract
:1. Introduction
2. Materials and Methods
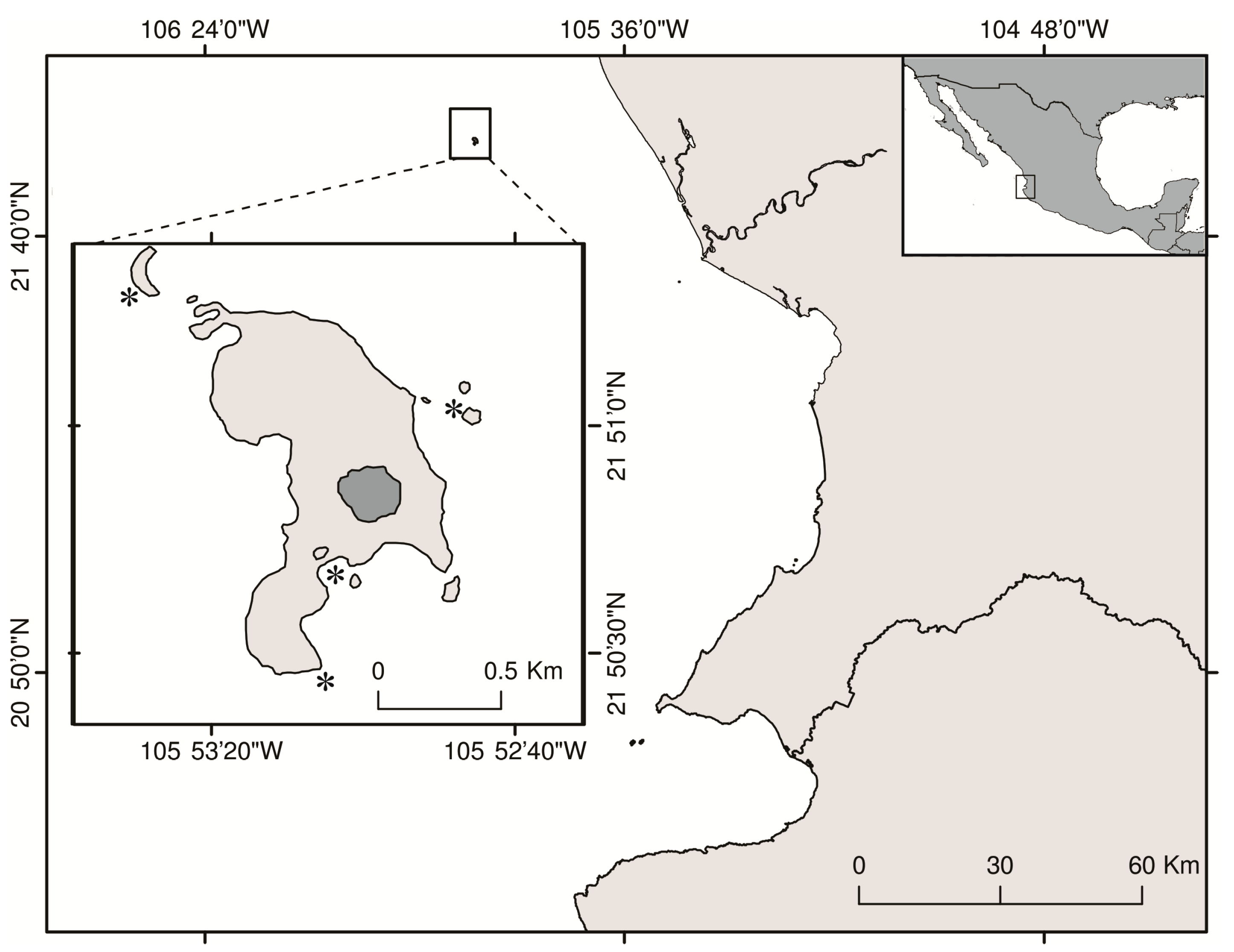
2.1. Sampling
2.2. Morphological Description
2.3. Molecular Identification
2.4. Sequence Alignment and Phylogenetic Analysis
3. Results
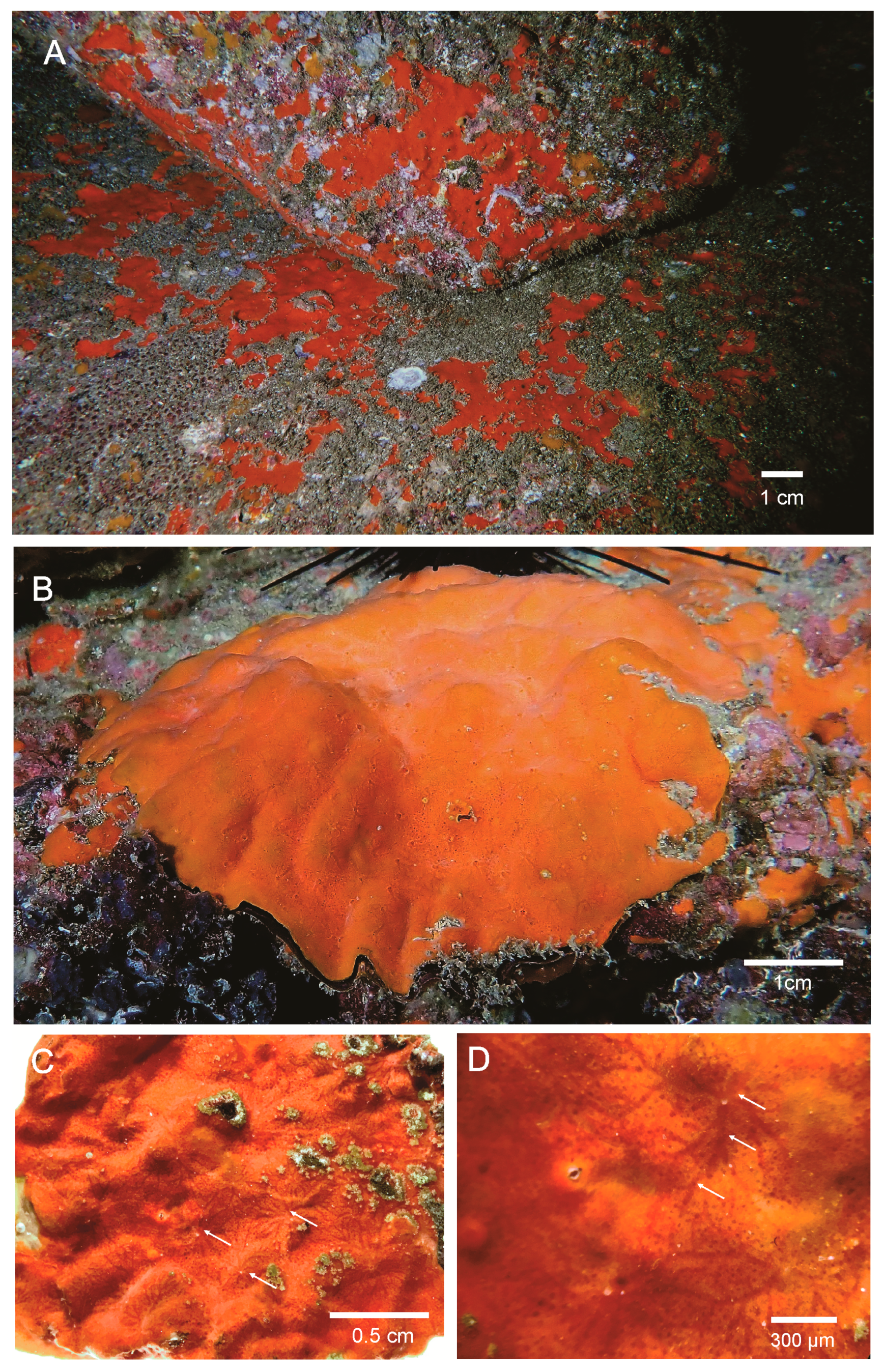
3.1. Taxonomic Descriptions
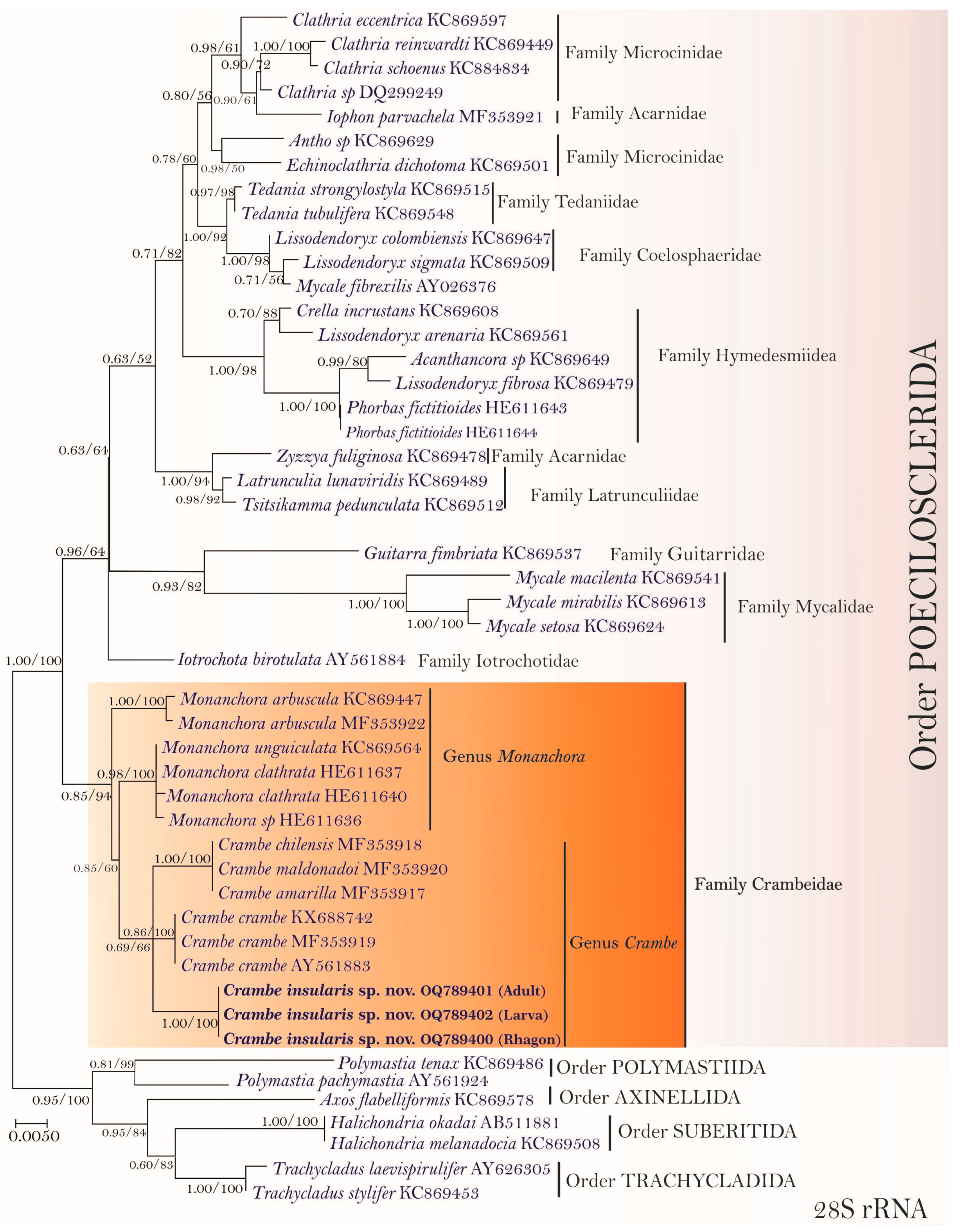
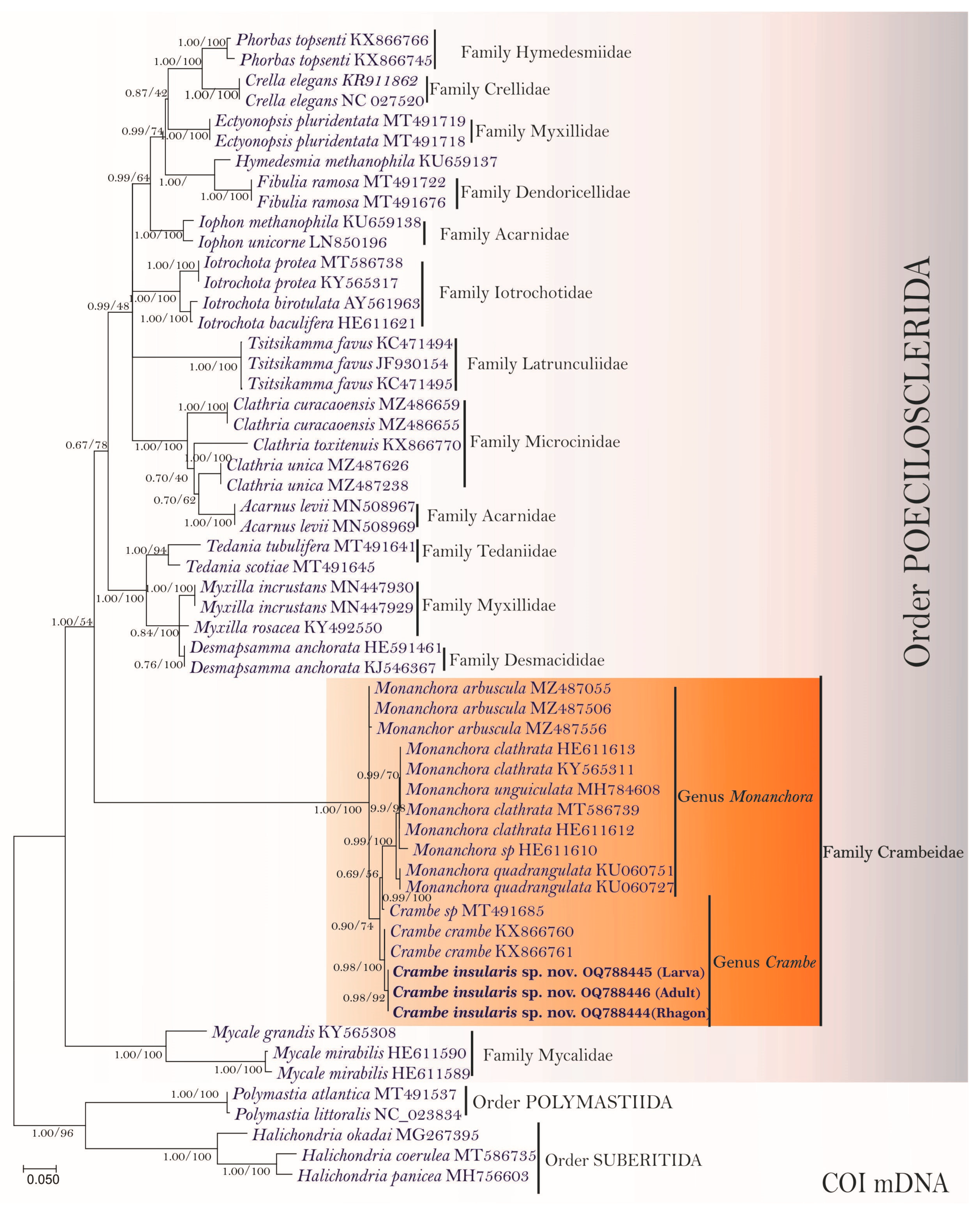
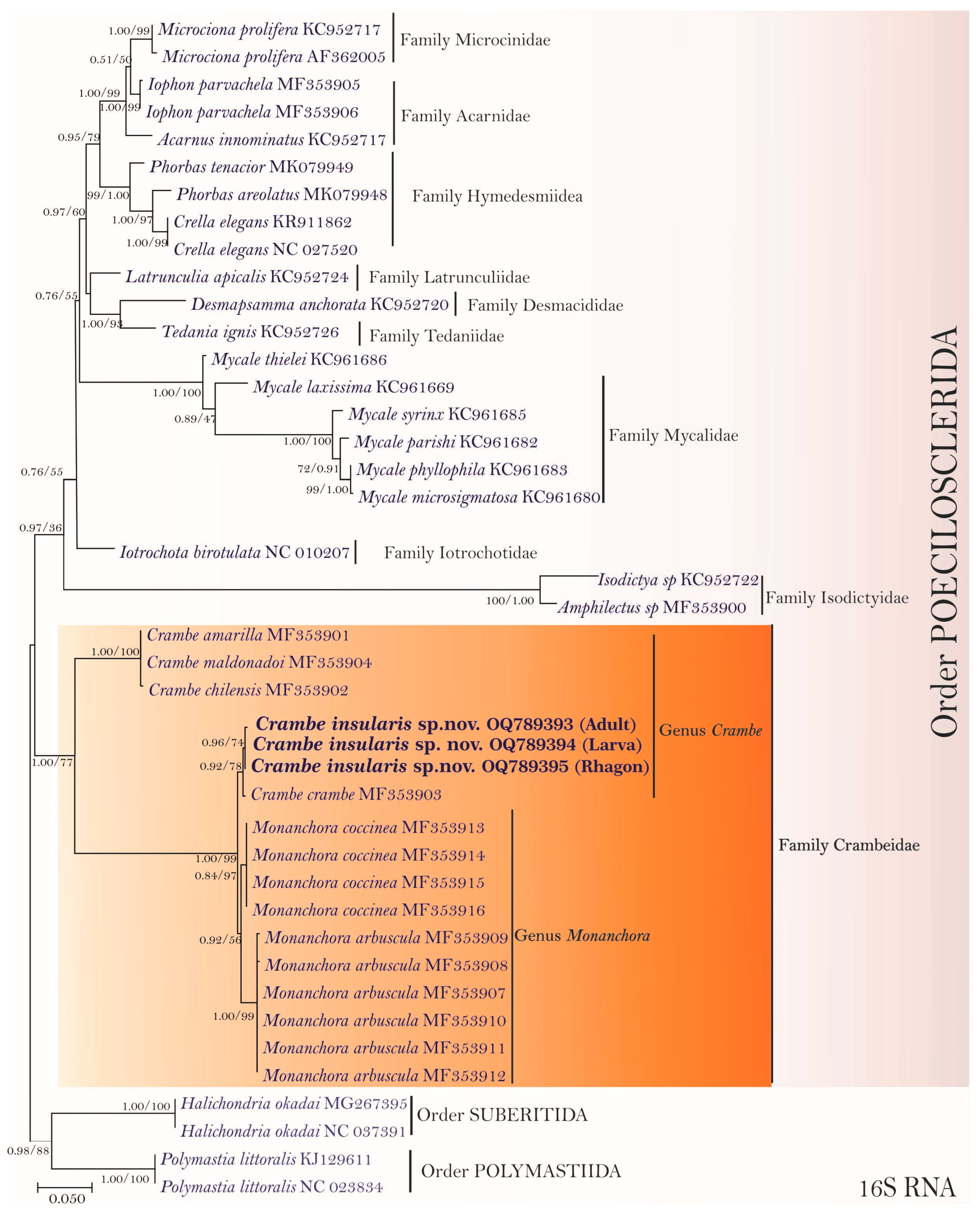
3.2. Molecular Phylogenetic Analyses and Pairwise Genetics Distance Inferences
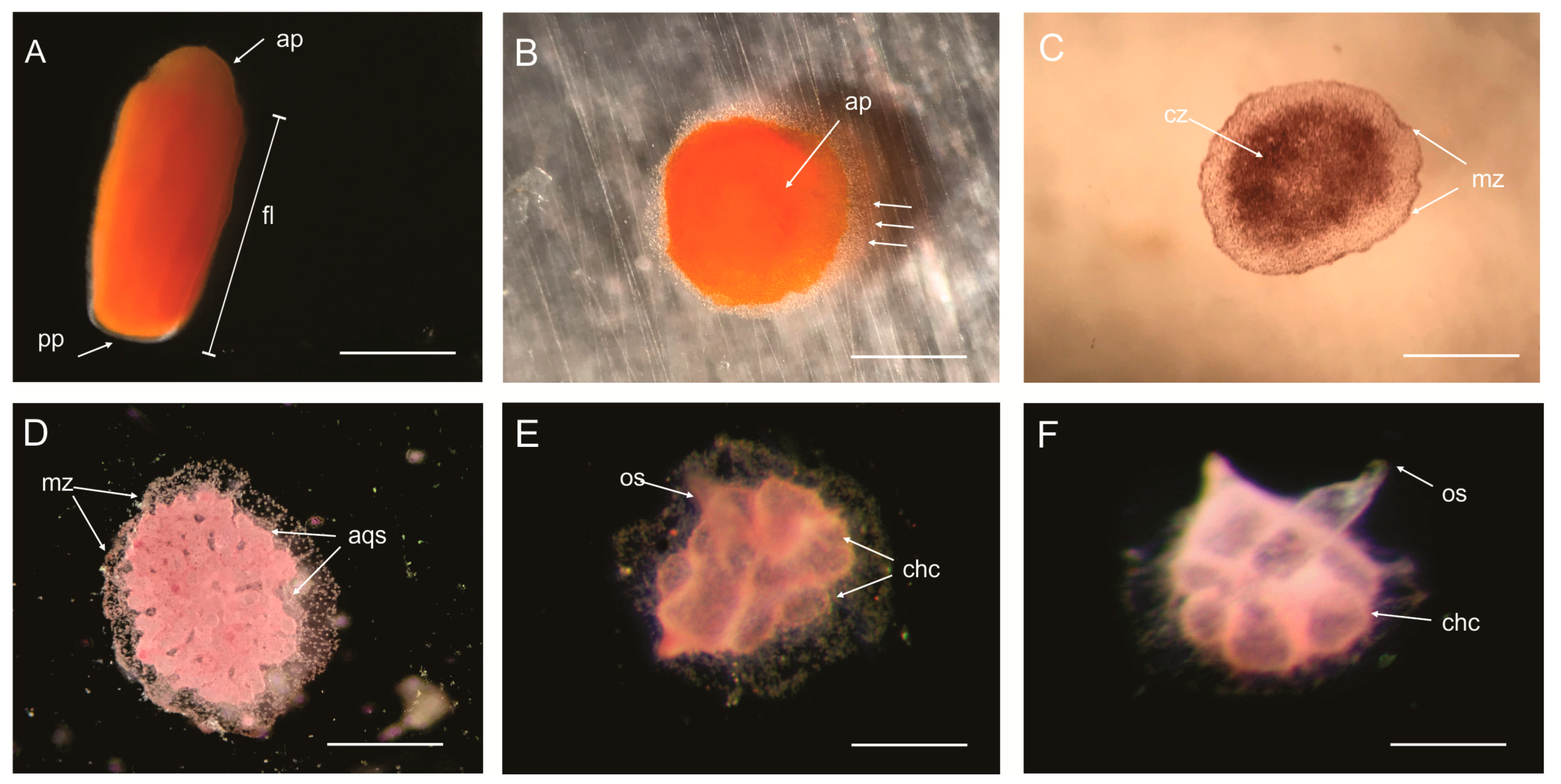
3.3. External Morphology of the Larva
3.4. Rhagon Phase
4. Discussion
Parenchymella Larva
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Van Soest, R.W.M. Family Crambeidae Lévi, 1963. In Systema Porifera: A Guide to the Classification of Sponges; Hooper, J.N.A., Van Soest, R.W.M., Eds.; Kluwer Academic/Plenum Publishers: New York, NY, USA, 2002; pp. 547–555. ISBN 0-306-47260-0. [Google Scholar]
- Esteves, E.L.; Lerner, C.; Lôbo-Hajdu, G.; Hajdu, E. Monanchora brasiliensis sp. nov. (Poecilosclerida: Demospongiae), new crambeid from the Brazilian coast, south-western Atlantic, with monodentate anchorate chelae. J. Mar. Biol. Assoc. UK 2012, 92, 869–876. [Google Scholar] [CrossRef]
- Van Soest, R.W.M.; Boury-Esnault, N.; Hooper, J.N.A.; Rützler, K.; de Voogd, N.J.; Alvarez, B.; Hajdu, E.; Pisera, A.B.; Manconi, R.; Schönberg, C.; et al. World Porifera Database. 2019. Available online: http://www.marinespecies.org/porifera (accessed on 29 April 2019).
- Esteves, E.L.; de Paula, T.S.; Lerner, C.; Lôbo-Hajdu, G.; Hajdu, E. Morphological and molecular systematics of the ‘Monanchora arbuscula complex’ (Poecilosclerida: Crambeidae), with the description of five new species and a biogeographic discussion of the genus in the Tropical Western Atlantic. Invertebr. Syst. 2018, 32, 457–503. [Google Scholar] [CrossRef]
- Arevabini, C.; Crivelenti, Y.D.; de Abreu, M.H.; Bitencourt, T.A.; Santos, M.F.C.; Berlinck, R.G.S.; Hajdu, E.; Beleboni, R.E.; Fachin, A.L.; Marins, M. Antifungal activity of metabolites from the marine sponges Amphimedon sp. and Monanchora arbuscula against Aspergillus flavus strains isolated from peanuts (Arachis hypogaea). Nat. Prod. Commun. 2014, 9, 33–36. [Google Scholar] [CrossRef] [PubMed]
- Santos, M.F.; Harper, P.M.; Williams, D.E.; Mesquita, J.T.; Pinto, E.G.; da Costa-Silva, T.A.; Hajdu, E.; Ferreira, A.G.; Santos, R.A.; Murphy, P.J.; et al. Anti-parasitic guanidine and pyrimidine alkaloids from the marine sponge Monanchora arbuscula. J. Nat. Prod. 2015, 78, 1101–1112. [Google Scholar] [CrossRef]
- Uriz, M.J.; Maldonado, M. A reconsideration of the relationship between polyaxonid and monaxonid spicules in Demospongiae: New data from the genera Crambe and Discorhabdella (Porifera). Biol. J. Linn. Soc. 1995, 55, 1–15. [Google Scholar] [CrossRef]
- Maldonado, M.; Carmen Carmona, M.; Van Soest, R.W.; Pomponi, S.A. First record of the sponge genera Crambe and Discorhabdella for the eastern Pacific, with description of three new species. J. Nat. Hist. 2001, 35, 1261–1276. [Google Scholar] [CrossRef]
- Leví, C. Résultats scientifiques des campagnes de la ‘Calypso’. V. Campagne en Mer Rouge (1951–1952). 11. Spongiaires de Mer Rouge. Ann. Inst. Océanographique Monaco. 1958, 34, 3–46. [Google Scholar]
- Boury-Esnault, N. Spongiaires de la zone rocheuse de Banyuls-sur-Mer. II. Systématique. Vie Milieu 1971, 22, 287–349. [Google Scholar]
- Vacelet, J.; Vasseur, P.; Lévi, C. Spongiaires de la Pente Externe des Récifs coralliens de Tulear (sud-ouest de Madagascar). Mémoires Muséum Natl. D’Histoire Nat. Série A Zool. 1976, 99, 1–116. [Google Scholar]
- Maldonado, M.; Uriz, M.J. Skeletal morphology of two controversial poecilosclerid genera (Porifera, Demospongiae): Discorhabdella and Crambe. Helgoländer Meeresunters 1996, 50, 369–390. [Google Scholar] [CrossRef]
- Maldonado, M.; Carmona, M.C.; Uriz, M.J.; Cruzado, A. Decline in Mesozoic reef-building sponges explained by silicon limitation. Nature 1999, 401, 785–788. [Google Scholar] [CrossRef]
- Lévi, C. Résultats scientifiques des campagnes de la ‘Calypso’. XIV. Campagne 1954 dans l’Océan Indien. 2. Les spongiaires de l’Ile Aldabra. Ann. Inst. Océanographique Monaco 1961, 39, 3–32. [Google Scholar]
- Lévi, C. Les Spongiaires à desmes astéroïdes. Bull. Inst. Océanographique Monaco 1960, 57, 1–8. [Google Scholar]
- Lévi, C. Spongiaires d’Afrique du Sud (1). Poecilosclerides. Trans. R. Soc. South Afr. 1963, 37, 1–71. [Google Scholar] [CrossRef]
- Uriz, M.J. Deep-water sponges from the continental shelf and slope of Namibia (South-West Africa): Classes Hexactinellida and Demospongiae. Monogr. Zool. Mar. 1988, 3, 9–157. [Google Scholar]
- Maldonado, M.; Benito, J. Crambe tuberosa n. sp. (Demospongiae, Poecilosclerida): A new Mediterranean poecilosclerid with lithistid affinities. Cah. Biol. Mar. 1991, 32, 323–332. [Google Scholar]
- Esteves, E.L.; Lôbo-Hajdu, G.; Hajdu, E. Three new species of Crambe (Crambeidae: Poecilosclerida: Demospongiae) from the south-eastern Pacific, with a review of morphological characters for the genus. J. Mar. Biol. Assoc. UK 2007, 87, 1367–1378. [Google Scholar] [CrossRef]
- de Voogd, N.J.; Alvarez, B.; Boury–Esnault, N.; Carballo, J.L.; Cárdenas, P.; Díaz, M.-C.; Dohrmann, M.; Downey, R.; Hajdu, E.; Hooper, J.N.A.; et al. World Porifera Database. Crambeidae Lévi. 1963. Available online: https://www.marinespecies.org/porifera/porifera.php?p=taxdetails&id=1316 (accessed on 11 January 2023).
- Gómez, P.; Bakus, G. Aplysina gerardogreeni and Aplysina aztecus (Porifera: Demospongiae) new species from the Mexican Pacific. An. Inst. Cienc. Mar Limnol. Univ. Autónoma México 1992, 19, 175–180. [Google Scholar]
- Gómez, P. First record and new species of Gastrophanella (Porifera: Demospongiae: Lithistida) from the central East Pacific. Proc. Biol. Soc. Wash. 1998, 111, 774–780. Available online: https://biostor.org/reference/69064 (accessed on 11 January 2023).
- Cruz-Barraza, J.A.; Carballo, J.L. Taxonomy of sponges (Porifera) associated with corals from the Mexican Pacific Ocean. Zool. Stud. 2008, 47, 741–758. [Google Scholar]
- Aguilar-Camacho, J.M.; Carballo, J.L.; Cruz-Barraza, J.A. Acarnidae (Porifera: Demospongiae: Poecilosclerida) from the Mexican Pacific Ocean with the description of six new species. Sci. Mar. 2013, 77, 677–696. [Google Scholar] [CrossRef]
- Carballo, J.L.; Gómez, P.; Cruz-Barraza, J.A. Biodiversidad de Porifera en México. Rev. Mex. Biodivers. 2014, 85, 143–153. [Google Scholar] [CrossRef]
- Boury-Esnault, N.; Rützler, K. Thesaurus of Sponge Morphology. Smithson. Contrib. Zool. 1997, 596, 1–55. [Google Scholar] [CrossRef]
- Folmer, O.; Black, M.; Hoeh, W.; Lutz, R.; Vrijenhoek, R. DNA primers for amplification of mitochondrial cytochrome c oxidase subunit I from diverse metazoan invertebrates. Mol. Mar. Biol. Biotechnol. 1994, 3, 294–299. [Google Scholar]
- Nichols, S.A. An evaluation of support for order-level monophyly and interrelationships within the class Demospongiae using partial data from the large subunit rDNA and cytochrome oxidase subunit I. Mol. Phylogenet. Evol. 2005, 34, 81–96. [Google Scholar] [CrossRef]
- Lavrov, D.V.; Wang, X.; Kelly, M. Reconstructing ordinal relationships in the Demospongiae using mitochondrial genomic data. Mol. Phylogenet. Evol. 2008, 49, 111–124. [Google Scholar] [CrossRef]
- Kumar, S.; Stecher, G.; Li, M.; Knyaz, C.; Tamura, K. MEGA X: Molecular evolutionary genetics analysis across computing platforms. Mol. Phylogenet. Evol. 2018, 35, 1547–1549. [Google Scholar] [CrossRef]
- Posada, D. jModelTest: Phylogenetic model averaging. Mol. Biol. Evol. 2008, 25, 1253–1256. [Google Scholar] [CrossRef]
- Ronquist, F.; Teslenko, M.; van der, P.M.; Ayres, D.L.; Darling, A.; Höhna, S.; Larget, B.; Liu, L.; Suchard, M.A.; Huelsenbeck, J.P. MrBayes 3.2: Efficient Bayesian Phylogenetic Inference and Model Choice Across a Large Model Space. Syst. Biol. 2012, 61, 539–542. [Google Scholar] [CrossRef]
- Stone, R.P.; Lehnert, H.; Reiswig, H. A guide to the deep-water sponges of the Aleutian Island Archipelago. In NOAA Professional Paper NMFS12; NOAA: Silver Spring, MA, USA, 2011; Volume 12, 187p. Available online: http://hdl.handle.net/1834/30422 (accessed on 3 September 2021).
- Carter, H.J. New Genus of Sponges. Ann. Mag. Nat. Hist. 1883, 11, 369–370. [Google Scholar] [CrossRef]
- Hechtel, G.J. New species and records of shallow water Demospongiae from Barbados, West Indies. Postilla 1969, 132, 1–38. [Google Scholar] [CrossRef]
- Van Soest, R.W.M. Monanchora stocki n. sp. (Porifera, Poecilosclerida) from the Mid-Atlantic islands. Bijdr. Dierkd. 1990, 60, 249–255. [Google Scholar]
- Pulitzer-Finali, G. A collection of marine sponges from East Africa. Ann. Mus. Civ. Stor. Nat. Giacomo Doria 1993, 89, 247–350. Available online: https://biostor.org/reference/100342 (accessed on 1 January 2022).
- Vacelet, J.; Boury-Esnault, N. Une nouvelle éponge du genre Crambe (Demospongiae, Poecilosclerida) de Méditerraneée, C. tailliezi n. sp. Trav. Sci. Parc. Nation Port-Cros Fr. 1982, 8, 107–113. [Google Scholar]
- Spalding, M.D.; Fox, G.R.; Allen, N.; Davidson, Z.A.; Ferdaña, M.; Finlayson, B.S.; Halpern, M.A.J.; Al, L.; Sara, A.L.; Kirsten, D.M.; et al. Marine Ecoregions of the World: A Bioregionalization of Coastal and Shelf Areas. BioScience 2007, 57, 573–583. [Google Scholar] [CrossRef]
- Redmond, N.E.; Morrow, C.C.; Thacker, R.W.; Diaz, M.C.; Boury-Esnault, N.; Cárdenas, P.; Hajdu, E.; Lôbo-Hajdu, G.; Picton, B.E.; Pomponi, S.A.; et al. Phylogeny and systematics of Demospongiae in light of new small-subunit ribosomal DNA (18S) sequences. Integr. Comp. Biol. 2013, 53, 388–415. [Google Scholar] [CrossRef]
- Hill, M.S.; Hill, A.L.; Lopez, J.; Perterson, K.J.; Pomponi, S.; Diaz, M.C.; Thacker, R.W.; Adamska, M.; Boury-Esnault, N.; Cárdenas, P.; et al. Reconstruction of family level phylogenetic relationships within Demospongiae (Porifera) using nuclear encoded housekeeping genes. PLoS ONE 2013, 8, e50437. [Google Scholar] [CrossRef]
- Vargas, S.; Erpenbeck, D.; Göcke, C.; Hall, K.A.; Hooper, J.N.A.; Janussen, D.; Wörheide, G. Molecular phylogeny of Abyssocladia (Cladorhizidae: Poecilosclerida) and Phelloderma (Phellodermidae: Poecilosclerida) suggests a diversification of chelae microscleres in cladorhizid sponges. Zool. Scr. 2013, 42, 106–116. [Google Scholar] [CrossRef]
- Thacker, R.W.; Hill, A.L.; Hill, M.S.; Redmond, N.E.; Collins, A.G.; Morrow, C.C.; Spicer, L.; Carmack, C.A.; Zappe, M.E.; Pohlmann, D.; et al. Nearly complete 28S rRNA gene sequences confirm new hypotheses of sponge evolution. Integr. Comp. Biol. 2013, 53, 373–387. [Google Scholar] [CrossRef]
- Ereskovsky, A.V. Typization of Sponge Development and Its Significance for Phylogeny. In The Comparative Embryology of Sponges; Springer: Dordrecht, The Netherlands, 2010; pp. 209–230. [Google Scholar] [CrossRef]
- Uriz, M.J.; Turon, X.; Becerro, M.A. Morphology and ultrastructure of the swimming larvae of Crambe crambe (Demospongiae, Poecilosclerida). Invertebr. Biol. 2001, 120, 295–307. [Google Scholar] [CrossRef]
- Boury-Esnault, N. Ultrastructure de la larve parenchymella d’Hamigera hamigera (Schmidt) (Démosponge, Poecilosclerida). Origine des cellules grises. Cah. Biol. Mar. 1976, 17, 9–20. [Google Scholar]
- Bergquist, P.R.; Sinclair, C.R.; Green, C.R.; Silyn-Roberts, H. Comparative morphology and behavior of larvae of Demospongiae. In Biologie des Spongiaires; Colloq. Int. CNRS n. 291; Lévi, C., Boury-Esnault, N., Eds.; CNRS: Paris, France, 1979; pp. 103–111. [Google Scholar]
- Maldonado, M.; George, S.B.; Young, C.M.; Vaquerizo, I. Depth regulation in parenchymella larvae of a demosponge: Relative roles of skeletogenesis, biochemical changes, and behavior. Mar. Ecol. Progr. Ser. 1997, 148, 115–124. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Material Examined | Choanosomal Styles/Tylotes | Ectosomal Subtylostyles |
|---|---|---|
| Holotype LEB–ICML–(3187) | 200–(241.4)–275 × 5.0–(6.8)–10 | 155–(190.1)–225 × 2.5–(2.9)–5 |
| Paratype LEB–ICML–(3188) | 178–(245.5)–325 × 5.0–(9.4)–10.5 | 175–(221.6)–264 × 3.5–(6.5)–6 |
| Paratype LEB–ICML–(3189) | 198–(231.5)–287 × 5.0–(6.9)–9.8 | 169–(210.6)–256 × 3.6–(5.7)–7.5 |
| Species | Locality | Ectosomal (Sub) (Tylo) Styles | Choanosomal (Acantho) (Tylo) Styles | Desmas | Anchorate Isochelea | Additional Microscleres | Skeletal Arrangement | References | ||
|---|---|---|---|---|---|---|---|---|---|---|
| Crambe insularis sp. nov. | Pacific coast of Mexico | 211–411 | 5.5–6.8 | 275–483 | 12–16 | Absent | Absent | Absent | Plumose | [*] |
| C. crambe Schmidt, 1862 | Mediterranean Sea | 316–281 | 7 | 423–539 | 11–17 | Poorly developed Astroclone or Sphaeroclone | Unguiferate with reduced shortened three-toothed I: 28–43 II: 8–11 | Absent | Plumose | [1,13] |
| C. erecta Pulitzer-Finali, 1993 | Adriatic Sea | 230–340 | 6–9 | 350–500 | 14–38 | Astroclone 120–180 | Anchorate I: 33 II:16–19 | Absent | Plumose | [1,8,12,37] |
| C. paramensis Maldonado, Carmona, van Soest & Pomponi, 2001 | Pacific coast of Panama | 117–208 | 2.5–3.5 | 180–300 | 6.5–12 | Astroclone 26–65 | Spatulate Three/five-toothed 20–22 | Absent | Hymedesmioid | [1,8] |
| C. acuata Lévi, 1958 | Red Sea and Aldabra southern Madagascar | 180–220 | 2–7 | 120–175 | 8–10 | Astroclone 15–75 | Three/five-toothed I: 20–22 II: 8–10 | Spined microxeas 16–90, rare raphides | – | [11,15] |
| C. acuata Lévi, 1958 | South Atlantic, East coasts of Africa | 267.7 | 10.5 | 315 | 10.75 | Astro/sphaeroclone 105 | Anchorate 35 | Spined microxeas, rare raphides | – | [15,16] |
| C. chilensis Esteves et al.,2007 | South–east Pacific, coast of Chile | 186–387 | 4–14 | 245–833 | 10–42 | Astroclone (4–7 arms) 50–232 | Spatuliferous– five–toothed 21–37 | Spined microxeas. 30–67 | Hymedesmioid | [19] |
| C. amarilla Esteves et al.,2007 | South–east Pacific, coast of Chile | 189–402 | 4–10 | 315–877 | 9–24 | Astroclone (6 arms) 43–178 | Unguiferate/Spatuliferous five–toothed 21–33 | Rare or even absent spined microxeas 45–58, Raphides | Hymedesmioid | [19] |
| C. maldonadoi Esteves et al.,2007 | South–east Pacific, coast of Chile | 400–785 | 4–15 | 255–769 | 12–34 | Sphaeroclone (5–6 arms) 94–211 | Spatuliferous–three/five–toothed 22–31 | Spined microxeas 22–43 | Hymedesmioid | [19] |
| C. tailliezi Vacelet & Boury–Esnault, 1982 | Western Mediterranean | Microspiny point 205–330 | 2.5–4 | 350–460 | 6–8 | Astro/sphaeroclone 25–120 | Unguiferate three toothed 23–26 | Spined microxeas 7.8–15.6 Desmoi | – | [38] |
| C. tuberosa Maldonado & Benito, 1991 | Mediterranean Alboran Island | 87–104 | 4.6–5.8 | Tuberose tyle I: 500–700 II: 150–300 | I: 20–30 II: 5–10 | Sphaeroclone (4–7 arms) 94–134 | Five–toothed isoanchorae 27–30 | Special asters | Hymedesmioid | [18] |
| 28s | |||||||||
| 1 | 2 | 3 | 4 | 5 | 6 | 7 | |||
| 1 | Crambe insularis sp. nov. | – | 0.004 | 0.005 | 0.004 | 0.004 | 0.005 | 0.005 | |
| 2 | Crambe amarilla | 0.017 | – | 0.000 | 0.000 | 0.004 | 0.005 | 0.004 | |
| 3 | Crambe maldonadoi | 0.017 | 0.000 | – | 0.000 | 0.005 | 0.005 | 0.004 | |
| 4 | Crambe chilensis | 0.017 | 0.000 | 0.000 | – | 0.004 | 0.005 | 0.004 | |
| 5 | Crambe crambe | 0.014 | 0.012 | 0.012 | 0.012 | – | 0.004 | 0.005 | |
| 6 | Monanchora arbuscula | 0.022 | 0.025 | 0.025 | 0.025 | 0.017 | – | 0.004 | |
| 7 | Monanchora clathrata | 0.024 | 0.020 | 0.020 | 0.020 | 0.018 | 0.006 | – | |
| 16S | |||||||||
| 1 | 2 | 3 | 4 | 5 | 6 | 7 | |||
| 1 | Crambe insularis sp. nov. | – | 0.021 | 0.021 | 0.021 | 0.003 | 0.007 | 0.006 | |
| 2 | Crambe amarilla | 0.156 | – | 0.000 | 0.002 | 0.022 | 0.023 | 0.022 | |
| 3 | Crambe maldonadoi | 0.156 | 0.000 | – | 0.002 | 0.022 | 0.023 | 0.022 | |
| 4 | Crambe chilensis | 0.159 | 0.002 | 0.002 | – | 0.022 | 0.023 | 0.023 | |
| 5 | Crambe crambe | 0.006 | 0.160 | 0.160 | 0.163 | – | 0.007 | 0.005 | |
| 6 | Monanchora arbuscula | 0.027 | 0.172 | 0.172 | 0.175 | 0.026 | – | 0.007 | |
| 7 | Monanchora coccinea | 0.015 | 0.166 | 0.166 | 0.169 | 0.013 | 0.020 | – | |
| COI | |||||||||
| 1 | 2 | 3 | 4 | 5 | 6 | 7 | 8 | ||
| 1 | Crambe insularis sp. nov. | – | 0.003 | 0.005 | 0.008 | 0.007 | 0.007 | 0.006 | 0.007 |
| 2 | Crambe crambe | 0.005 | – | 0.004 | 0.008 | 0.007 | 0.006 | 0.005 | 0.007 |
| 3 | Crambe sp. | 0.016 | 0.011 | – | 0.008 | 0.006 | 0.006 | 0.005 | 0.006 |
| 4 | Monanchora sp. | 0.043 | 0.037 | 0.036 | – | 0.004 | 0.004 | 0.008 | 0.005 |
| 5 | Monanchora uningulata | 0.032 | 0.027 | 0.026 | 0.013 | – | 0.000 | 0.007 | 0.004 |
| 6 | Monanchora clathrata | 0.032 | 0.027 | 0.025 | 0.012 | 0.000 | – | 0.007 | 0.004 |
| 7 | Monanchora arbuscula | 0.025 | 0.020 | 0.018 | 0.047 | 0.036 | 0.036 | – | 0.007 |
| 8 | Monanchora quuadrangulata | 0.034 | 0.029 | 0.024 | 0.021 | 0.011 | 0.011 | 0.035 | – |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bautista-Guerrero, E.; Carballo, J.L.; Rodríguez-Troncoso, A.P. Crambe insularis sp. nov. (Crambeidae: Poecilosclerida) a New Crambeid from the Eastern Tropical North Pacific: Morphological, Molecular and Ontogenetic Approach. Diversity 2023, 15, 608. https://doi.org/10.3390/d15050608
Bautista-Guerrero E, Carballo JL, Rodríguez-Troncoso AP. Crambe insularis sp. nov. (Crambeidae: Poecilosclerida) a New Crambeid from the Eastern Tropical North Pacific: Morphological, Molecular and Ontogenetic Approach. Diversity. 2023; 15(5):608. https://doi.org/10.3390/d15050608
Chicago/Turabian StyleBautista-Guerrero, Eric, José Luis Carballo, and Alma Paola Rodríguez-Troncoso. 2023. "Crambe insularis sp. nov. (Crambeidae: Poecilosclerida) a New Crambeid from the Eastern Tropical North Pacific: Morphological, Molecular and Ontogenetic Approach" Diversity 15, no. 5: 608. https://doi.org/10.3390/d15050608