
Major Cuban Green Turtle Rookeries: Identification of Management Units and Their Genetic Relationships with Other Greater Caribbean Populations
 , , ,
, , ,
Abstract
:1. Introduction

2. Materials and Methods
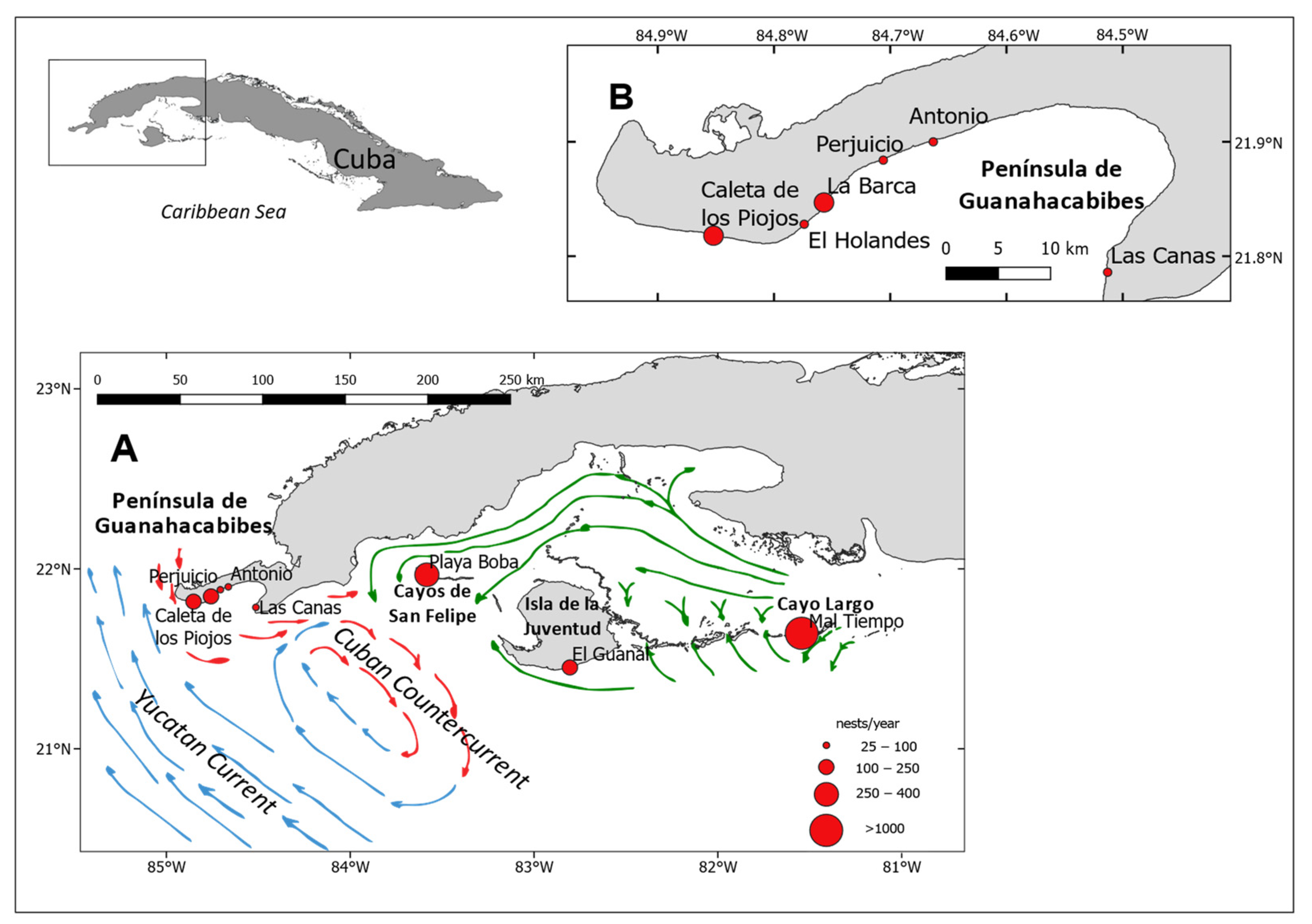
2.1. Sampling and Study Site
2.2. Laboratory Analyses
2.3. Data Analysis
3. Results
3.1. Genetic Diversity
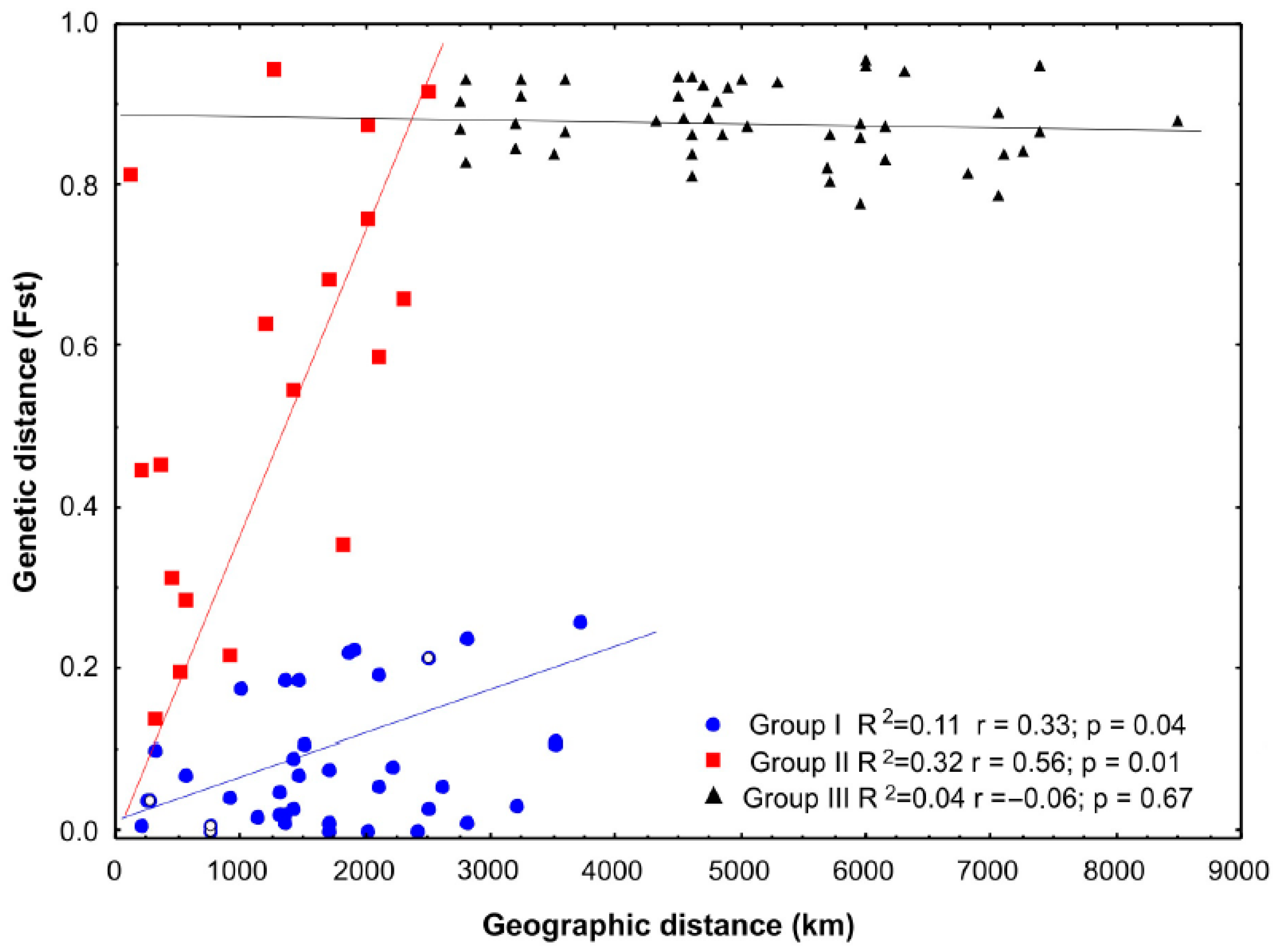
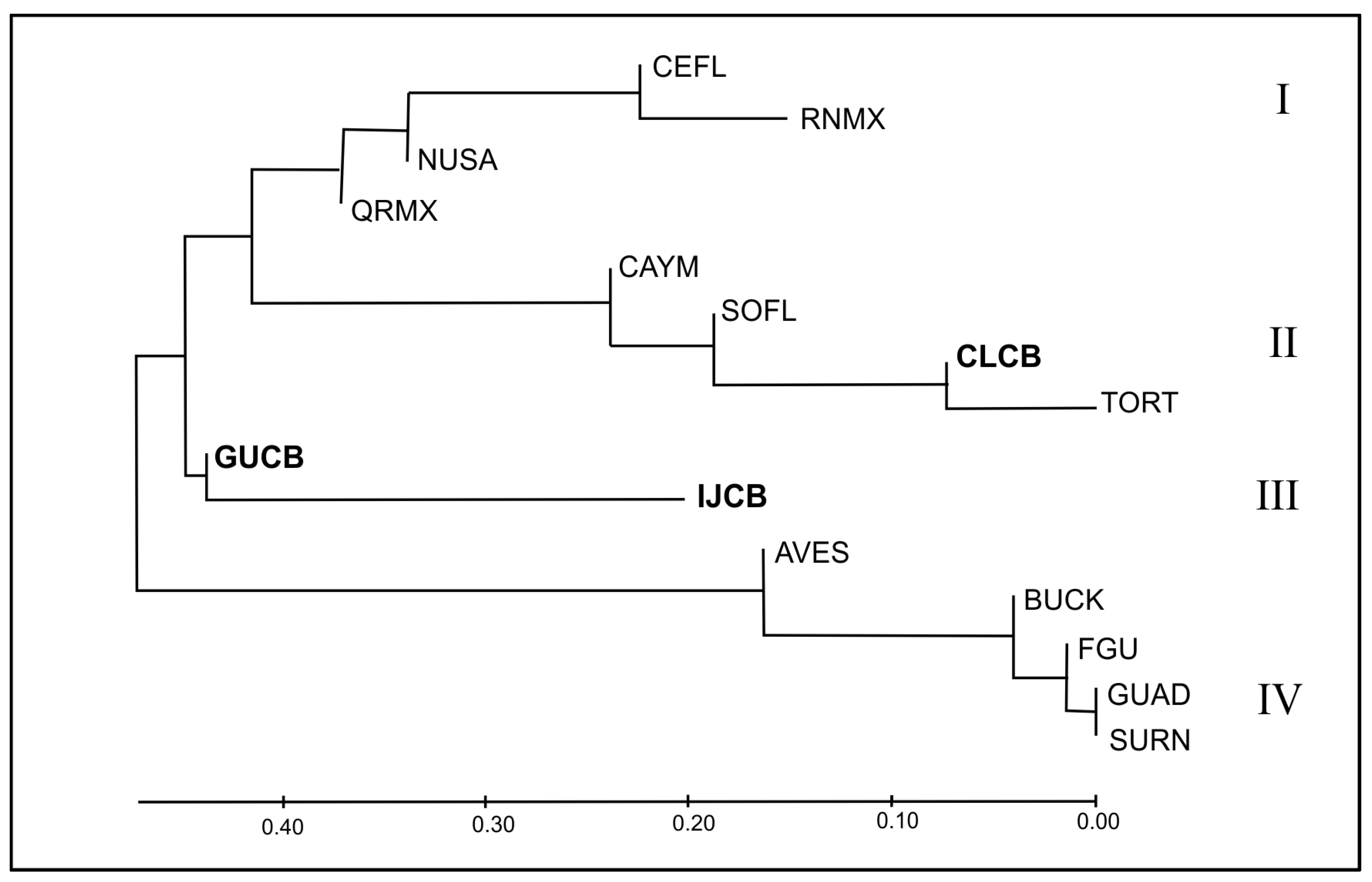
3.2. Identification of Cuban Management Units
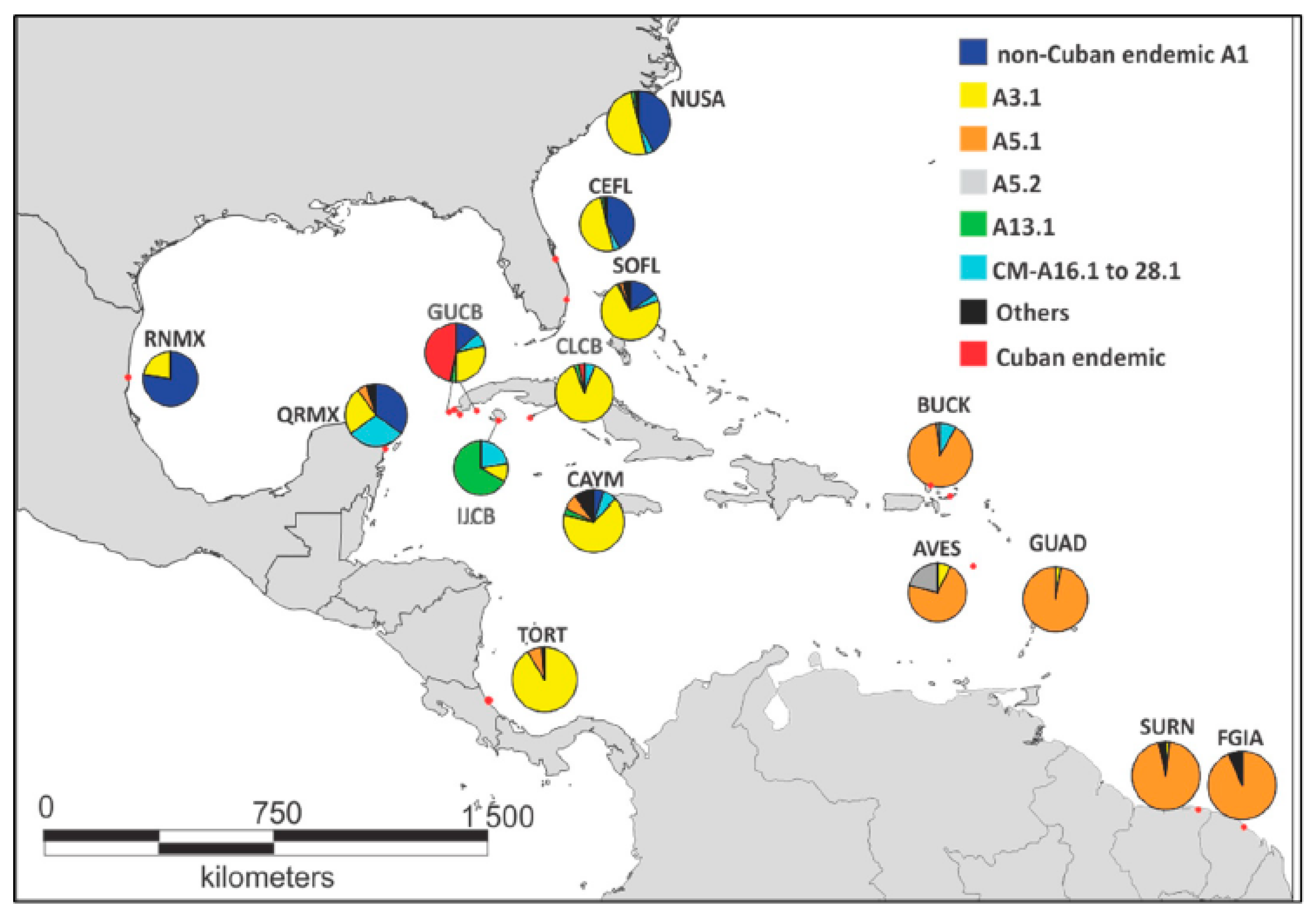
3.3. Cuban Rookeries in the Wider Caribbean Context

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| A | NUSA | SOFL | CEFL | RNMX | QRMX | GUCB | IJCB | CLCB | CAYM | TORT | BUCK | AVES | GUAD | SURN | FGU |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| NUSA | 0.004 | 0.006 | 0.005 | 0.012 | 0.000 | 0.000 | 0.000 | 0.007 | 0.000 | 0.000 | 0.000 | 0.000 | 0.000 | 0.000 | |
| SOFL | 0.043 | 0.000 | 0.000 | 0.001 | 0.000 | 0.000 | 0.055 | 0.020 | 0.000 | 0.000 | 0.000 | 0.000 | 0.000 | 0.000 | |
| CEFL | 0.038 | 0.181 | 0.327 | 0.005 | 0.000 | 0.000 | 0.000 | 0.000 | 0.000 | 0.000 | 0.000 | 0.000 | 0.000 | 0.000 | |
| RNMX | 0.100 | 0.207 | 0.001 | 0.009 | 0.000 | 0.000 | 0.000 | 0.000 | 0.000 | 0.000 | 0.000 | 0.000 | 0.000 | 0.000 | |
| QRMX | 0.072 | 0.157 | 0.099 | 0.072 | 0.001 | 0.000 | 0.000 | 0.007 | 0.004 | 0.000 | 0.000 | 0.000 | 0.000 | 0.000 | |
| GUCB | 0.091 | 0.155 | 0.171 | 0.115 | 0.060 | 0.000 | 0.000 | 0.000 | 0.000 | 0.000 | 0.000 | 0.000 | 0.000 | 0.000 | |
| IJCB | 0.634 | 0.599 | 0.662 | 0.760 | 0.299 | 0.362 | 0.000 | 0.000 | 0.000 | 0.000 | 0.000 | 0.000 | 0.000 | 0.000 | |
| CLCB | 0.193 | 0.022 | 0.330 | 0.569 | 0.271 | 0.200 | 0.800 | 0.000 | 0.113 | 0.000 | 0.000 | 0.000 | 0.000 | 0.000 | |
| CAYM | 0.049 | 0.028 | 0.191 | 0.148 | 0.037 | 0.104 | 0.370 | 0.059 | 0.007 | 0.020 | 0.000 | 0.000 | 0.007 | 0.000 | |
| TORT | 0.112 | 0.035 | 0.252 | 0.239 | 0.188 | 0.225 | 0.547 | 0.022 | 0.003 | 0.000 | 0.000 | 0.000 | 0.000 | 0.000 | |
| BUCK | 0.871 | 0.866 | 0.909 | 0.878 | 0.781 | 0.789 | 0.834 | 0.887 | 0.777 | 0.820 | 0.170 | 0.205 | 0.095 | 0.255 | |
| AVES | 0.857 | 0.858 | 0.904 | 0.860 | 0.778 | 0.788 | 0.826 | 0.869 | 0.774 | 0.816 | 0.015 | 0.179 | 0.030 | 0.098 | |
| GUAD | 0.925 | 0.893 | 0.924 | 0.950 | 0.854 | 0.819 | 0.930 | 0.951 | 0.824 | 0.839 | 0.008 | 0.025 | 0.771 | 0.754 | |
| SURN | 0.930 | 0.900 | 0.926 | 0.950 | 0.880 | 0.834 | 0.937 | 0.952 | 0.848 | 0.845 | 0.026 | 0.044 | −0.015 | 0.642 | |
| FGU | 0.921 | 0.893 | 0.923 | 0.954 | 0.858 | 0.821 | 0.922 | 0.944 | 0.829 | 0.839 | 0.001 | 0.018 | −0.018 | −0.009 | |
| B | NUSA | SOFL | CEFL | RNMX | QRMX | GUCB | IJCB | CLCB | CAYM | TORT | BUCK | AVES | GUAD | SURN | FGU |
| NUSA | 0.004 | 0.006 | 0.0008 | 0.012 | 0.001 | 0.000 | 0.000 | 0.007 | 0.000 | 0.000 | 0.000 | 0.000 | 0.000 | 0.000 | |
| SOFL | 0.050 | 0.000 | 0.000 | 0.001 | 0.000 | 0.000 | 0.074 | 0.124 | 0.000 | 0.000 | 0.000 | 0.000 | 0.000 | 0.000 | |
| CEFL | 0.043 | 0.198 | 0.327 | 0.005 | 0.000 | 0.000 | 0.000 | 0.000 | 0.000 | 0.000 | 0.000 | 0.000 | 0.000 | 0.000 | |
| RNMX | 0.109 | 0.225 | 0.000 | 0.013 | 0.000 | 0.000 | 0.000 | 0.000 | 0.000 | 0.000 | 0.000 | 0.000 | 0.000 | 0.000 | |
| QRMX | 0.081 | 0.176 | 0.107 | 0.071 | 0.004 | 0.000 | 0.000 | 0.002 | 0.004 | 0.000 | 0.000 | 0.000 | 0.000 | 0.000 | |
| GUCB | 0.054 | 0.139 | 0.089 | 0.074 | 0.040 | 0.000 | 0.000 | 0.000 | 0.000 | 0.000 | 0.000 | 0.000 | 0.000 | 0.000 | |
| IJCB | 0.660 | 0.629 | 0.685 | 0.760 | 0.312 | 0.448 | 0.000 | 0.000 | 0.000 | 0.000 | 0.000 | 0.000 | 0.000 | 0.000 | |
| CLCB | 0.214 | 0.020 | 0.354 | 0.589 | 0.285 | 0.217 | 0.814 | 0.060 | 0.117 | 0.000 | 0.000 | 0.000 | 0.000 | 0.000 | |
| CAYM | 0.056 | 0.010 | 0.222 | 0.192 | 0.070 | 0.101 | 0.454 | 0.037 | 0.078 | 0.000 | 0.000 | 0.000 | 0.000 | 0.000 | |
| TORT | 0.112 | 0.032 | 0.258 | 0.239 | 0.186 | 0.185 | 0.548 | 0.022 | 0.0192 | 0.000 | 0.000 | 0.000 | 0.000 | 0.000 | |
| BUCK | 0.878 | 0.877 | 0.916 | 0.880 | 0.789 | 0.841 | 0.837 | 0.891 | 0.814 | 0.820 | 0.296 | 0.194 | 0.104 | 0.235 | |
| AVES | 0.865 | 0.868 | 0.911 | 0.865 | 0.778 | 0.833 | 0.829 | 0.876 | 0.803 | 0.811 | −0.002 | 0.397 | 0.139 | 0.226 | |
| GUAD | 0.931 | 0.904 | 0.931 | 0.950 | 0.860 | 0.872 | 0.930 | 0.954 | 0.863 | 0.839 | 0.009 | 0.007 | 0.769 | 0.748 | |
| SURN | 0.935 | 0.909 | 0.932 | 0.950 | 0.884 | 0.882 | 0.936 | 0.954 | 0.881 | 0.845 | 0.0268 | 0.027 | −0.016 | 0.599 | |
| FGU | 0.926 | 0.903 | 0.930 | 0.940 | 0.862 | 0.872 | 0.920 | 0.945 | 0.864 | 0.839 | 0.011 | 0.011 | −0.018 | −0.008 |
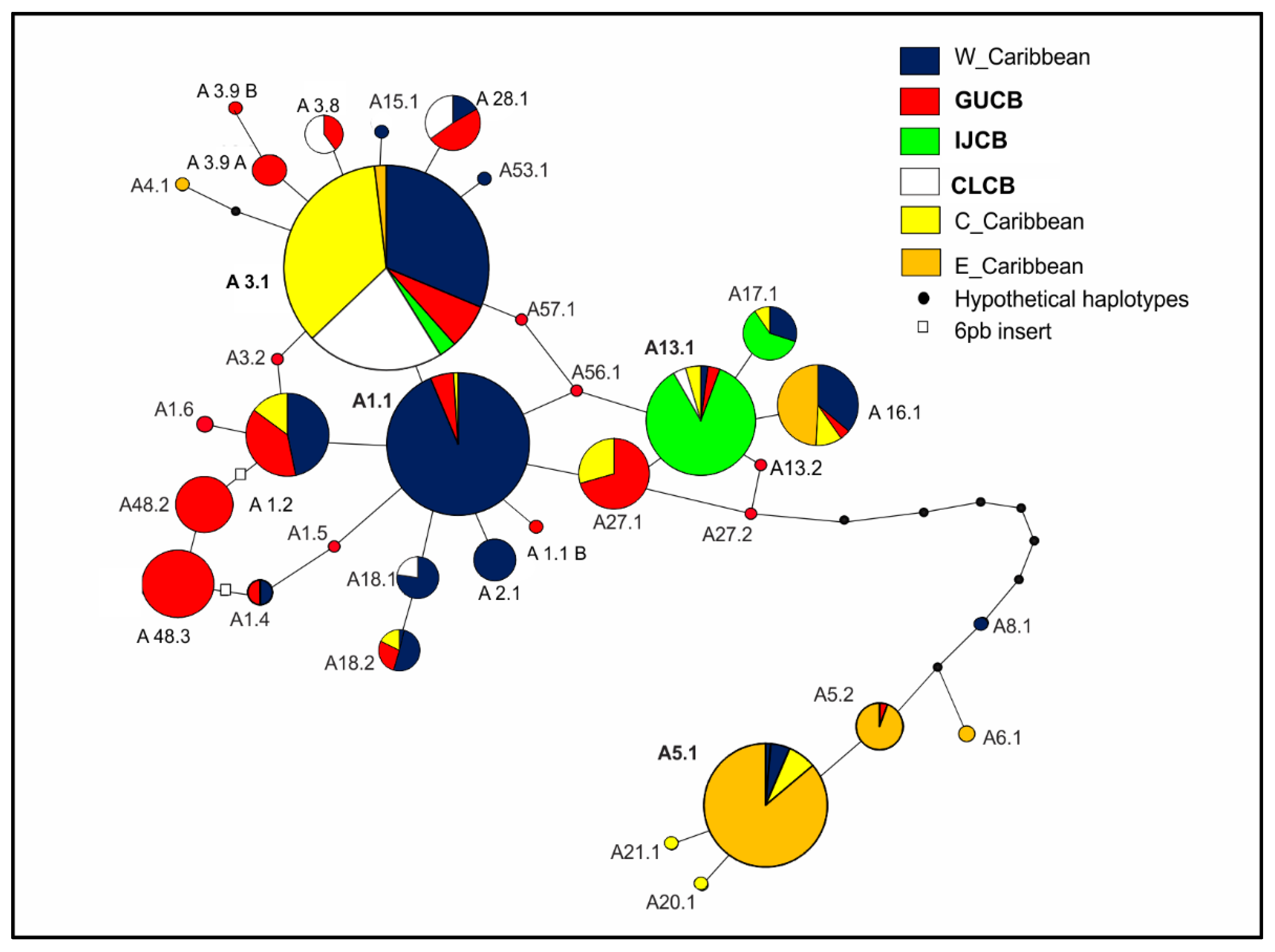
3.4. Evolutionary Relationships
4. Discussion
4.1. Genetic Diversity in Cuban Rookeries
4.2. Evolutionary Perspectives and Management Units

4.3. Cuban Rookeries in the Caribbean Context
4.4. Conservation Implications and Future Studies
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Kohn, M.H.; Murphy, W.J.; Ostrander, E.A.; Wayne, R.K. Genomics and Conservation Genetics. Trends Ecol. Evol. 2006, 21, 629–637. [Google Scholar] [CrossRef] [PubMed]
- Velez-Zuazo, X.; Ramos, W.D.; van Dam, R.P.; Diez, C.E.; Abreu Grobois, F.A.; Mcmillan, W.O. Dispersal, Recruitment and Migratory Behaviour in a Hawksbill Sea Turtle Aggregation. Mol. Ecol. 2008, 17, 839–853. [Google Scholar] [CrossRef] [PubMed]
- Bolker, B.M.; Okuyama, T.; Bjorndal, K.A.; Bolten, A.B. Incorporating Multiple Mixed Stocks in Mixed Stock Analysis: ‘Many-to-Many’Analyses. Mol. Ecol. 2007, 16, 685–695. [Google Scholar] [CrossRef]
- Bass, A.L.; Epperly, S.P.; Braun-McNeill, J. Green Turtle (Chelonia mydas) Foraging and Nesting Aggregations in the Caribbean and Atlantic: Impact of Currents and Behavior on Dispersal. J. Hered. 2006, 97, 346–354. [Google Scholar] [CrossRef] [PubMed]
- Moncada, F.; Abreu-Grobois, F.A.; Muhlia-Melo, A.; Bell, C.; Tröeng, S.; Bjorndal, K.A.; Bolten, A.B.; Meylan, A.B.; Zurita, J.; Espinosa, G. Movement Patterns of Green Turtles (Chelonia mydas) in Cuba and Adjacent Caribbean Waters Inferred from Flipper Tag Recaptures. J. Herpetol. 2006, 40, 22–34. [Google Scholar] [CrossRef]
- Jensen, M.P.; FitzSimmons, N.N.; Dutton, P.H. Molecular Genetics of Sea Turtles. In The Biology of Sea Turtles; Wyneken, J., Lohmann, K.J., Musick, J.A., Eds.; CRC Press: Boca Raton, FL, USA, 2013; Volume 3, pp. 135–161. ISBN 978-1-4398-7308-3. [Google Scholar]
- Chaloupka, M.; Musick, J.A.; Lutz, P.L. Age, Growth, and Population Dynamics. Biol. Sea Turt. 1997, 2, 233–276. [Google Scholar]
- Komoroske, L.M.; Jensen, M.P.; Stewart, K.R.; Shamblin, B.M.; Dutton, P.H. Advances in the Application of Genetics in Marine Turtle Biology and Conservation. Front. Mar. Sci. 2017, 4, 156. [Google Scholar] [CrossRef]
- Bowen, B.W.; Meylan, A.B.; Ross, J.P.; Limpus, C.J.; Balazs, G.H.; Avise, J.C. Global Population Structure and Natural History of the Green Turtle (Chelonia mydas) in Terms of Matriarchal Phylogeny. Evolution 1992, 46, 865–881. [Google Scholar]
- Formia, A.; Broderick, A.C.; Glen, F.; Godley, B.J.; Hays, G.C.; Bruford, M.W. Genetic Composition of the Ascension Island Green Turtle Rookery Based on Mitochondrial DNA: Implications for Sampling and Diversity. Endanger. Species Res. 2007, 3, 145–158. [Google Scholar] [CrossRef]
- Reece, J.S.; Castoe, T.A.; Parkinson, C.L. Historical Perspectives on Population Genetics and Conservation of Three Marine Turtle Species. Conserv. Genet. 2005, 6, 235–251. [Google Scholar] [CrossRef]
- Bjorndal, K.A.; Bolten, A.B.; Moreira, L.; Bellini, C.; Marcovaldi, M.Â. Population Structure and Diversity of Brazilian Green Turtle Rookeries Based on Mitochondrial DNA Sequences. Chelonian Conserv. Biol. 2006, 5, 262–268. [Google Scholar] [CrossRef]
- Seminoff, J.A.; Allen, C.D.; Balazs, G.H.; Dutton, P.H.; Eguchi, T.; Haas, H.; Hargrove, S.A.; Jensen, M.; Klemm, D.L.; Lauritsen, A.M.; et al. Status Review of the Green Turtle (Chelonia mydas) under the Endangered Species Act; SWFSC (Southwest Fisheries Science Center): La Jolla, CA, USA, 2015. [Google Scholar]
- Dow, W.; Eckert, K.; Palmer, M.; Kramer, P. An Atlas of Sea Turtle Nesting Habitat for the Wider Caribbean Region; Wider Caribbean Sea Turtle Conservation Network (WIDECAST): Beaufort, NC, USA, 2007. [Google Scholar]
- Moncada Gavilán, F.; Azanza-Ricardo, J.; Forneiro, Y.; Gerhartz, J.L.; Nodarse, G.; Cruz, Y. Programa de Monitoreo de Tortugas Marinas. In Estado Actual de la Biodiversidad Marino-Costera, en la Región de los Archipiélagos del Sur de Cuba; Hernández Ávila, A., Ed.; Impresos Dominicanos s.r.l: La Habana, Cuba, 2014; pp. 130–141. [Google Scholar]
- Nodarse Andreu, G.; Moncada Gavilán, F.; Medina Cruz, Y.; Rodríguez Castillo, C.; Hernández Orozco, F.; Blanco López, R.; Escobar González, E. Comportamiento de La Anidación de Tortugas Marinas en los Cayos San Felipe y Archipiélago de los Canarreos, Cuba (2001–2006). Rev. Cuba. Investig. Pesq. 2010, 27, 66–71. [Google Scholar]
- Moncada, F.; Abreu-Grobois, F.A.; Bagley, D.; Bjorndal, K.A.; Bolten, A.B.; Camiñas, J.A.; Ehrhart, L.; Muhlia-Melo, A.; Nodarse, G.; Schroeder, B.A. Movement Patterns of Loggerhead Turtles Caretta Caretta in Cuban Waters Inferred from Flipper Tag Recaptures. Endanger. Species Res. 2010, 11, 61–68. [Google Scholar] [CrossRef]
- Shamblin, B.M.; Dutton, P.H.; Shaver, D.J.; Bagley, D.A.; Putman, N.F.; Mansfield, K.L.; Ehrhart, L.M.; Peña, L.J.; Nairn, C.J. Mexican Origins for the Texas Green Turtle Foraging Aggregation: A Cautionary Tale of Incomplete Baselines and Poor Marker Resolution. J. Exp. Mar. Biol. Ecol. 2017, 488, 111–120. [Google Scholar] [CrossRef]
- Ruiz, U.A.; Vega, P.M.; Riveron, G.F.B.; Abreu, G.A.F.; Solano, A.G.; Pérez, M.T.; Pérez, B.E.; Azanza, R.J.; Frías, S.R.; Díaz, F.R.; et al. Estructura Genética de Poblaciones de Caretta Caretta En El Gran Caribe y La Costa Atlántica de Estados Unidos, con Enfasis en Colonias de Anidacion del Suroeste Cubano. Rev. Investig. Mar. 2008, 29, 151–160. [Google Scholar]
- Ruiz-Urquiola, A.; Riverón-Giró, F.B.; Pérez-Bermúdez, E.; Abreu-Grobois, F.A.; González-Pumariega, M.; James-Petric, B.L.; Díaz-Fernández, R.; Álvarez-Castro, J.M.; Jager, M.; Azanza-Ricardo, J.; et al. Population Genetic Structure of Greater Caribbean Green Turtles (Chelonia mydas) Based on Mitochondrial DNA Sequences, with an Emphasis on Rookeries from Southwestern Cuba. Rev. Investig. Mar. 2010, 31, 33–52. [Google Scholar]
- Shamblin, B.M.; Bagley, D.A.; Ehrhart, L.M.; Desjardin, N.A.; Martin, R.E.; Hart, K.M.; Naro-Maciel, E.; Rusenko, K.; Stiner, J.C.; Sobel, D.; et al. Genetic Structure of Florida Green Turtle Rookeries as Indicated by Mitochondrial DNA Control Region Sequences. Conserv. Genet. 2015, 16, 673–685. [Google Scholar] [CrossRef]
- Abreu-Grobois, F.A.; Horrocks, J.A.; Formia, A.; Dutton, P.H.; LeRoux, R.; Vélez-Zuazo, X.; Soares, L.; Meylan, P. New MtDNA D-Loop Primers Which Work for a Variety of Marine Turtle Species May Increase the Resolution of Mixed Stock Analysis. In Proceedings of the 26th Annual Symposium on Sea Turtle Biology, ISTS, Athens, Greece, 3–8 April 2006; Frick, M., Panagopoulou, A., Rees, A.F., Williams, K., Eds.; International Sea Turtle Society: Island of Crete, Greece, 2006; p. 200. [Google Scholar]
- Azanza Ricardo, J.; Moncada Gavilán, F.; Martin-Viaña, Y.F.; Gerhartz Muro, J.L. Chapter 3: Cuba. In North Atlantic and Wider Caribbean: MTSG Regional Report; Nalovic, M., Cuevas, E., Godfrey, M., Eds.; IUCN-SSC Marine Turtle Specialist Group: Ross, CA, USA, 2019; pp. 48–67. [Google Scholar]
- Hillis, D.M.; Moritz, C.; Mable, B.K. Molecular Systematics; Sinauer: Sunderland, MA, USA, 1996; Volume 2. [Google Scholar]
- Hall, T. BioEdit, Version 5.0.6; North Carolina State University, Department of Microbiology: Raleigh, NC, USA, 2001.
- Kumazawa, Y.; Nishida, M. Complete Mitochondrial DNA Sequences of the Green Turtle and Blue-Tailed Mole Skink: Statistical Evidence for Archosaurian Affinity of Turtles. Mol. Biol. Evol. 1999, 16, 784–792. [Google Scholar] [CrossRef]
- Lahanas, P.N.; Miyamoto, M.M.; Bjorndal, K.A.; Bolten, A.B. Molecular Evolution and Population Genetics of Greater Caribbean Green Turtles (Chelonia mydas) as Inferred from Mitochondrial DNA Control Region Sequences. Genetica 1994, 94, 57–66. [Google Scholar] [CrossRef]
- Shamblin, B.M.; Dutton, P.H.; Bjorndal, K.A.; Bolten, A.B.; Naro-Maciel, E.; Santos, A.J.B.; Bellini, C.; Baptistotte, C.; Marcovaldi, M.Â.; Nairn, C.J. Deeper Mitochondrial Sequencing Reveals Cryptic Diversity and Structure in Brazilian Green Turtle Rookeries. Chelonian Conserv. Biol. 2015, 14, 167–172. [Google Scholar] [CrossRef]
- Shamblin, B.M.; Godfrey, M.H.; Pate, S.M.; Thompson, W.P.; Sutton, H.; Altman, J.; Fair, K.; McClary, J.; Wilson, A.M.; Milligan, B.; et al. Green Turtles Nesting at Their Northern Range Limit in the United States Represent a Distinct Subpopulation. Chelonian Conserv. Biol. 2018, 17, 314–319. [Google Scholar] [CrossRef]
- Encalada, S.E.; Lahanas, P.N.; Bjorndal, K.A.; Bolten, A.B.; Miyamoto, M.M.; Bowen, B.W. Phylogeography and Population Structure of the Atlantic and Mediterranean Green Turtle Chelonia mydas: A Mitochondrial DNA Control Region Sequence Assessment. Mol. Ecol. 1996, 5, 473–483. [Google Scholar] [CrossRef] [PubMed]
- Bjorndal, K.A.; Bolten, A.B.; Troeng, S. Population Structure and Genetic Diversity in Green Turtles Nesting at Tortuguero, Costa Rica, Based on Mitochondrial DNA Control Region Sequences. Mar. Biol. 2005, 147, 1449–1457. [Google Scholar] [CrossRef]
- Shamblin, B.M.; Bjorndal, K.A.; Bolten, A.B.; Hillis-Starr, Z.M.; Lundgren, I.A.N.; Naro-Maciel, E.; Nairn, C.J. Mitogenomic Sequences Better Resolve Stock Structure of Southern Greater Caribbean Green Turtle Rookeries. Mol. Ecol. 2012, 21, 2330–2340. [Google Scholar] [CrossRef]
- Costa Jordao, J.; Bondioli, A.C.V.; de Almeida-Toledo, L.F.; Bilo, K.; Berzins, R.; Le Maho, Y.; Chevallier, D.; De Thoisy, B. Mixed-Stock Analysis in Green Turtles Chelonia mydas: MtDNA Decipher Current Connections among West Atlantic Populations. Mitochondrial DNA Part A 2017, 28, 197–207. [Google Scholar] [CrossRef]
- Librado, P.; Rozas, J. DnaSP v5: A Software for Comprehensive Analysis of DNA Polymorphism Data. Bioinformatics 2009, 25, 1451–1452. [Google Scholar] [CrossRef] [PubMed]
- Excoffier, L.; Lischer, H.E.L. Arlequin Suite Ver 3.5: A New Series of Programs to Perform Population Genetics Analyses under Linux and Windows. Mol. Ecol. Resour. 2010, 10, 564–567. [Google Scholar] [CrossRef] [PubMed]
- Manni, F.; Guérard, E.; Heyer, E. Geographic Patterns of (Genetic, Morphologic, Linguistic) Variation: How Barriers Can Be Detected by Using Monmonier’s Algorithm. Hum. Biol. 2004, 76, 173–190. [Google Scholar] [CrossRef]
- Kumar, S.; Stecher, G.; Li, M.; Knyaz, C.; Tamura, K. MEGA X: Molecular Evolutionary Genetics Analysis across Computing Platforms. Mol. Biol. Evol. 2018, 35, 1547–1549. [Google Scholar] [CrossRef]
- Clement, M.; Posada, D.; Crandall, K.A. TCS: A Computer Program to Estimate Gene Genealogies. Mol. Ecol. 2000, 9, 1657–1659. [Google Scholar] [CrossRef]
- Múrias dos Santos, A.; Cabezas, M.P.; Tavares, A.I.; Xavier, R.; Branco, M. TcsBU: A Tool to Extend TCS Network Layout and Visualization. Bioinformatics 2016, 32, 627–628. [Google Scholar] [CrossRef] [PubMed]
- Benjamini, Y.; Yekutieli, D. False Discovery Rate–Adjusted Multiple Confidence Intervals for Selected Parameters. J. Am. Stat. Assoc. 2005, 100, 71–81. [Google Scholar] [CrossRef]
- Naro-Maciel, E.; Hart, K.M.; Cruciata, R.; Putman, N.F. DNA and Dispersal Models Highlight Constrained Connectivity in a Migratory Marine Megavertebrate. Ecography 2017, 40, 586–597. [Google Scholar] [CrossRef]
- Anderson, J.D.; Shaver, D.J.; Karel, W.J. Genetic Diversity and Natal Origins of Green Turtles (Chelonia mydas) in the Western Gulf of Mexico. J. Herpetol. 2013, 47, 251–257. [Google Scholar] [CrossRef]
- Maggs, C.A.; Castilho, R.; Foltz, D.; Henzler, C.; Jolly, M.T.; Kelly, J.; Olsen, J.; Perez, K.E.; Stam, W.; Väinölä, R.; et al. Evaluating Signatures of Glacial Refugia for North Atlantic Benthic Marine Taxa. Ecology 2008, 89, S108–S122. [Google Scholar] [CrossRef] [PubMed]
- Jensen, M.P.; FitzSimmons, N.N.; Bourjea, J.; Hamabata, T.; Reece, J.; Dutton, P.H. The Evolutionary History and Global Phylogeography of the Green Turtle (Chelonia mydas). J. Biogeogr. 2019, 46, 860–870. [Google Scholar] [CrossRef]
- van der Zee, J.P.; Christianen, M.J.; Nava, M.; Velez-Zuazo, X.; Hao, W.; Bérubé, M.; van Lavieren, H.; Hiwat, M.; Berzins, R.; Chevalier, J.; et al. Population Recovery Changes Population Composition at a Major Southern Caribbean Juvenile Developmental Habitat for the Green Turtle, Chelonia mydas. Sci. Rep. 2019, 9, 14392. [Google Scholar] [CrossRef] [PubMed]
- Bowen, B.W.; Rocha, L.A.; Toonen, R.J.; Karl, S.A. The Origins of Tropical Marine Biodiversity. Trends Ecol. Evol. 2013, 28, 359–366. [Google Scholar] [CrossRef]
- Dutton, P.H.; Jensen, M.P.; Frey, A.; LaCasella, E.; Balazs, G.H.; Zárate, P.; Chassin-Noria, O.; Sarti-Martinez, A.L.; Velez, E. Population Structure and Phylogeography Reveal Pathways of Colonization by a Migratory Marine Reptile (C Helonia Mydas) in the Central and Eastern Pacific. Ecol. Evol. 2014, 4, 4317–4331. [Google Scholar] [CrossRef]
- Jackson, J.B. Reefs since Columbus. Coral Reefs 1997, 16, S23–S32. [Google Scholar] [CrossRef]
- Patrício, A.R.; Vélez-Zuazo, X.; van Dam, R.P.; Diez, C.E. Genetic Composition and Origin of Juvenile Green Turtles Foraging at Culebra, Puerto Rico, as Revealed by MtDNA. Lat. Am. J. Aquat. Res. 2017, 45, 506–520. [Google Scholar] [CrossRef]
- Jensen, M.P.; Dalleau, M.; Gaspar, P.; Lalire, M.; Jean, C.; Ciccione, S.; Mortimer, J.A.; Quillard, M.; Taquet, C.; Wamukota, A.; et al. Seascape Genetics and the Spatial Ecology of Juvenile Green Turtles. Genes 2020, 11, 278. [Google Scholar] [CrossRef] [PubMed]
- Dethmers, K.E.; Broderick, D.; Moritz, C.; Fitzsimmons, N.N.; Limpus, C.J.; Lavery, S.; Whiting, S.; Guinea, M.; Prince, R.I.; Kennett, R.O.D. The Genetic Structure of Australasian Green Turtles (Chelonia mydas): Exploring the Geographical Scale of Genetic Exchange. Mol. Ecol. 2006, 15, 3931–3946. [Google Scholar] [CrossRef] [PubMed]
- Browne, D.C.; Horrocks, J.A.; Abreu-Grobois, F.A. Population Subdivision in Hawksbill Turtles Nesting on Barbados, West Indies, Determined from Mitochondrial DNA Control Region Sequences. Conserv. Genet. 2010, 11, 1541–1546. [Google Scholar] [CrossRef]
- Dutton, P.H.; Jensen, M.P.; Frutchey, K.; Frey, A.; LaCasella, E.; Balazs, G.H.; Cruce, J.; Tagarino, A.; Farman, R.; Tatarata, M. Genetic Stock Structure of Green Turtle (Chelonia mydas) Nesting Populations across the Pacific Islands. Pac. Sci. 2014, 68, 451–464. [Google Scholar] [CrossRef]
- García-Machado, E.; Ulmo-Díaz, G.; Castellanos-Gell, J.; Casane, D. Patterns of Population Connectivity in Marine Organisms of Cuba. Bull. Mar. Sci. 2018, 94, 193–211. [Google Scholar] [CrossRef]
- Stiebens, V.A.; Merino, S.E.; Roder, C.; Chain, F.J.; Lee, P.L.; Eizaguirre, C. Living on the Edge: How Philopatry Maintains Adaptive Potential. Proc. R. Soc. B Biol. Sci. 2013, 280, 20130305. [Google Scholar] [CrossRef]
- Gaspar, P.; Candela, T.; Shillinger, G.L. Dispersal of Juvenile Leatherback Turtles from Different Caribbean Nesting Beaches: A Model Study. Front. Mar. Sci. 2022, 9, 959366. [Google Scholar] [CrossRef]
- Allard, M.W.; Miyamoto, M.M.; Bjorndal, K.A.; Bolten, A.B.; Bowen, B.W. Support for Natal Homing in Green Turtles from Mitochondrial DNA Sequences. Copeia 1994, 1994, 34–41. [Google Scholar] [CrossRef]
- Carreras, C.; Pont, S.; Maffucci, F.; Pascual, M.; Barcelo, A.; Bentivegna, F.; Cardona, L.; Alegre, F.; SanFelix, M.; Fernandez, G.; et al. Genetic Structuring of Immature Loggerhead Sea Turtles (Caretta Caretta) in the Mediterranean Sea Reflects Water Circulation Patterns. Mar. Biol. 2006, 149, 1269–1279. [Google Scholar] [CrossRef]
- Bowen, B.W.; Meylan, A.B.; Avise, J.C. An Odyssey of the Green Sea Turtle: Ascension Island Revisited. Proc. Natl. Acad. Sci. USA 1989, 86, 573–576. [Google Scholar] [CrossRef]
- Gaspar, P.; Georges, J.-Y.; Fossette, S.; Lenoble, A.; Ferraroli, S.; Le Maho, Y. Marine Animal Behaviour: Neglecting Ocean Currents Can Lead Us up the Wrong Track. Proc. R. Soc. B Biol. Sci. 2006, 273, 2697–2702. [Google Scholar] [CrossRef] [PubMed]
- Luschi, P.; Hays, G.C.; Papi, F. A Review of Long-Distance Movements by Marine Turtles, and the Possible Role of Ocean Currents. Oikos 2003, 103, 293–302. [Google Scholar] [CrossRef]
- Hays, G.C. Ocean Currents and Marine Life. Curr. Biol. 2017, 27, R470–R473. [Google Scholar] [CrossRef] [PubMed]
- Blumenthal, J.M.; Abreu-Grobois, F.A.; Austin, T.J.; Broderick, A.C.; Bruford, M.W.; Coyne, M.S.; Ebanks-Petrie, G.; Formia, A.; Meylan, P.A.; Meylan, A.B.; et al. Turtle Groups or Turtle Soup: Dispersal Patterns of Hawksbill Turtles in the Caribbean. Mol. Ecol. 2009, 18, 4841–4853. [Google Scholar] [CrossRef]
- Gaspar, P.; Benson, S.R.; Dutton, P.H.; Réveillère, A.; Jacob, G.; Meetoo, C.; Dehecq, A.; Fossette, S. Oceanic Dispersal of Juvenile Leatherback Turtles: Going beyond Passive Drift Modeling. Mar. Ecol. Prog. Ser. 2012, 457, 265–284. [Google Scholar] [CrossRef]
- Putman, N.F.; Scott, R.; Verley, P.; Marsh, R.; Hays, G.C. Natal Site and Offshore Swimming Influence Fitness and Long-Distance Ocean Transport in Young Sea Turtles. Mar. Biol. 2012, 159, 2117–2126. [Google Scholar] [CrossRef]
- Shamblin, B.M.; Hart, K.M.; Martin, K.J.; Ceriani, S.A.; Bagley, D.A.; Mansfield, K.L.; Ehrhart, L.M.; Nairn, C.J. Green Turtle Mitochondrial Microsatellites Indicate Finer-Scale Natal Homing to Isolated Islands than to Continental Nesting Sites. Mar. Ecol. Prog. Ser. 2020, 643, 159–171. [Google Scholar] [CrossRef]
- Arriaza, L.; Rodas, L.; Simanca, J.; Lorenzo, S.L.; Milian, D.E.; Romero, P. Contribución a la Gestión Ambiental del Golfo de Batabanó, Cuba: Modelación Numérica de Corrientes Marinas. Rev. Investig. Mar. 2011, 29, 89–99. [Google Scholar]
- Moncada, F.; Coppenrath, C.M.; Hirsch, S.; Nodarse, G.; Page-Karjian, A.; Reeves, A.M.; Perrault, J.R. Nesting Green Turtle Tagged in Cuba, Recaptured in Florida. Mar. Turt. Newsl. 2019, 156, 1–2. [Google Scholar]
- Claro, R.; Reshetnikov, Y.S.; Alcolado, P.M. Physical Attributes of Coastal Cuba. In Ecology of the Marine Fishes of Cuba; Smithsonian Institution Press: Washington, DC, USA, 2001; pp. 1–20. [Google Scholar]
- Espinosa, G.; Díaz, R.; Matos, J.; Becquer, U.; Romo, J.; Borrell, Y. Variación Aloenzimática en Poblaciones Cubanas del Camarón Blanco Litopenaeus Schmitti. Rev. Investig. Mar. 2003, 24, 11–19. [Google Scholar]
- Barcia, A.R.; Lopez, G.E.; Hernandez, D.; García-Machado, E. Temporal Variation of the Population Structure and Genetic Diversity of Farfantepenaeus Notialis Assessed by Allozyme Loci. Mol. Ecol. 2005, 14, 2933–2942. [Google Scholar] [CrossRef] [PubMed]
- Castellanos-Gell, J.; Robainas-Barcia, A.; Pina-Amargós, F.; Chevalier-Monteagudo, P.; Metcalfe, C.; Molina, W.F.; Casane, D.; García-Machado, E. Genetic Diversity of Reef Fishes around Cuba: A Multispecies Assessment. Mar. Biol. 2016, 163, 1–16. [Google Scholar] [CrossRef]
- Chaves, J.A.; Peña, M.; Valdés-Uribe, J.A.; Muñoz-Pérez, J.P.; Vallejo, F.; Heidemeyer, M.; Torres-Carvajal, O. Connectivity, Population Structure, and Conservation of Ecuadorian Green Sea Turtles. Endanger. Species Res. 2017, 32, 251–264. [Google Scholar] [CrossRef]
- Barbanti, A.; Martin, C.; Blumenthal, J.M.; Boyle, J.; Broderick, A.C.; Collyer, L.; Ebanks, G.; Godley, B.J.; Mustin, W.; Ordonez, V.; et al. How many came home? Evaluating ex-situ conservation of green turtles in the Cayman Islands. Mol. Ecol. 2019, 28, 1637–1651. [Google Scholar] [CrossRef] [PubMed]
- Cremades, C.; Lefèvre, S. Chapter 2: Guadeloup. In Sea Turtles in the North-West Atlantic & Caribbean Region MTSG Regional Report; Nalovic, M., Cuevas, E., Godfrey, M., Eds.; IUCN-SSC Marine Turtle Specialist Group: Ross, CA, USA, 2018; pp. 40–49. [Google Scholar]
- Berzins, R.; Chevalier, J.; Chevalier, D.; De Thoisy, B.; Kelle, L.; Lankester, M.C.; Nalovic, M. Chapter 1: French Guiana. In Sea Turtles in the North-West Atlantic & Caribbean Region MTSG Regional Report; Nalovic, M., Cuevas, E., Godfrey, M., Eds.; IUCN-SSC Marine Turtle Specialist Group: Ross, CA, USA, 2018; pp. 28–39. [Google Scholar]



| C. Piojos | La Barca | Perjuicio | Antonio | San Felipe | I. Juventud | Cayo Largo | |
|---|---|---|---|---|---|---|---|
| C. Piojos | - | 38.89 | 31.74 | 36.44 | 28.48 | 41.24 | 58.36 |
| La Barca | 0.073 | - | 14.45 | 32.90 | 16.63 | 26.19 | 40.05 |
| Perjuicio | 0.068 | 0.012 | - | 21.03 | 9.60 | 16.31 | 21.03 |
| Antonio | 0.040 | 0.037 | 0.005 | - | 5.68 | 45.38 | 26.64 |
| San Felipe | 0.121 | 0.043 | 0.000 | 0.000 | - | 16.41 | 10.80 |
| I. Juventud | 0.316 | 0.430 | 0.448 | 0.430 | 0.610 | - | 31.97 |
| Cayo Largo | 0.326 | 0.292 | 0.269 | 0.213 | 0.200 | 0.800 | - |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Azanza-Ricardo, J.; Abreu-Grobois, F.A.; Oyama, K.; Chassin-Noria, O.; Espinosa-López, G.; González-Sansón, G. Major Cuban Green Turtle Rookeries: Identification of Management Units and Their Genetic Relationships with Other Greater Caribbean Populations. Diversity 2023, 15, 586. https://doi.org/10.3390/d15050586
Azanza-Ricardo J, Abreu-Grobois FA, Oyama K, Chassin-Noria O, Espinosa-López G, González-Sansón G. Major Cuban Green Turtle Rookeries: Identification of Management Units and Their Genetic Relationships with Other Greater Caribbean Populations. Diversity. 2023; 15(5):586. https://doi.org/10.3390/d15050586
Chicago/Turabian StyleAzanza-Ricardo, Julia, F. Alberto Abreu-Grobois, Ken Oyama, Omar Chassin-Noria, Georgina Espinosa-López, and Gaspar González-Sansón. 2023. "Major Cuban Green Turtle Rookeries: Identification of Management Units and Their Genetic Relationships with Other Greater Caribbean Populations" Diversity 15, no. 5: 586. https://doi.org/10.3390/d15050586