

How Common Are Lesions on the Tails of Sauropods? Two New Pathologies in Titanosaurs from the Late Cretaceous of Argentine Patagonia
 , and
, and
Abstract
:1. Introduction
2. Material and Methods
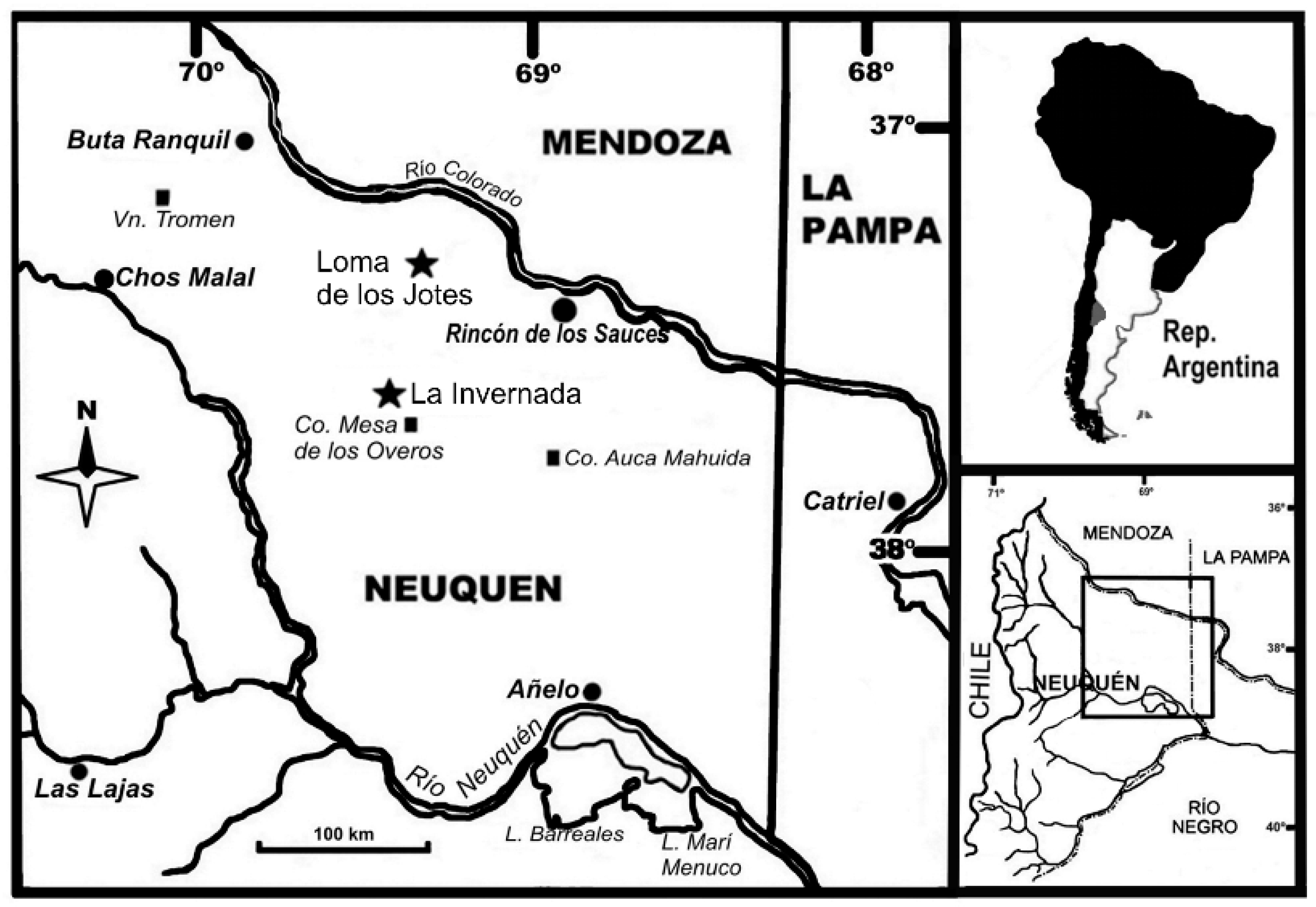
2.1. Institutional Abbreviations
2.2. Material
2.3. CT Scan
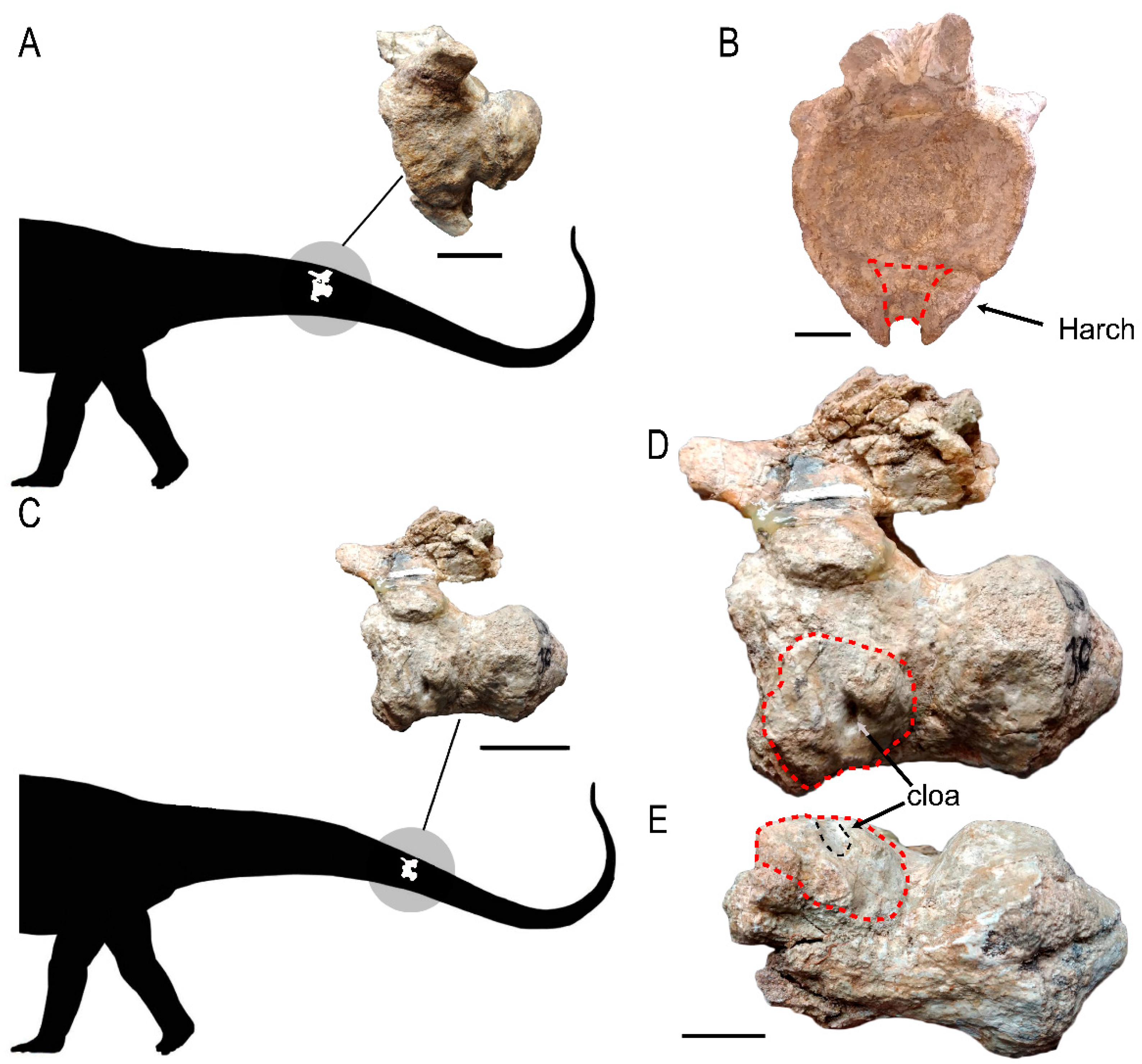
3. Description
3.1. External Morphology
3.2. Internal Morphology (CT Data)
4. Differential Diagnosis
4.1. Infections
4.2. Spondyloarthropathy
4.3. MAU-Pv-LI-601
4.4. MAU-Pv-LJ-472/1
5. Discussion
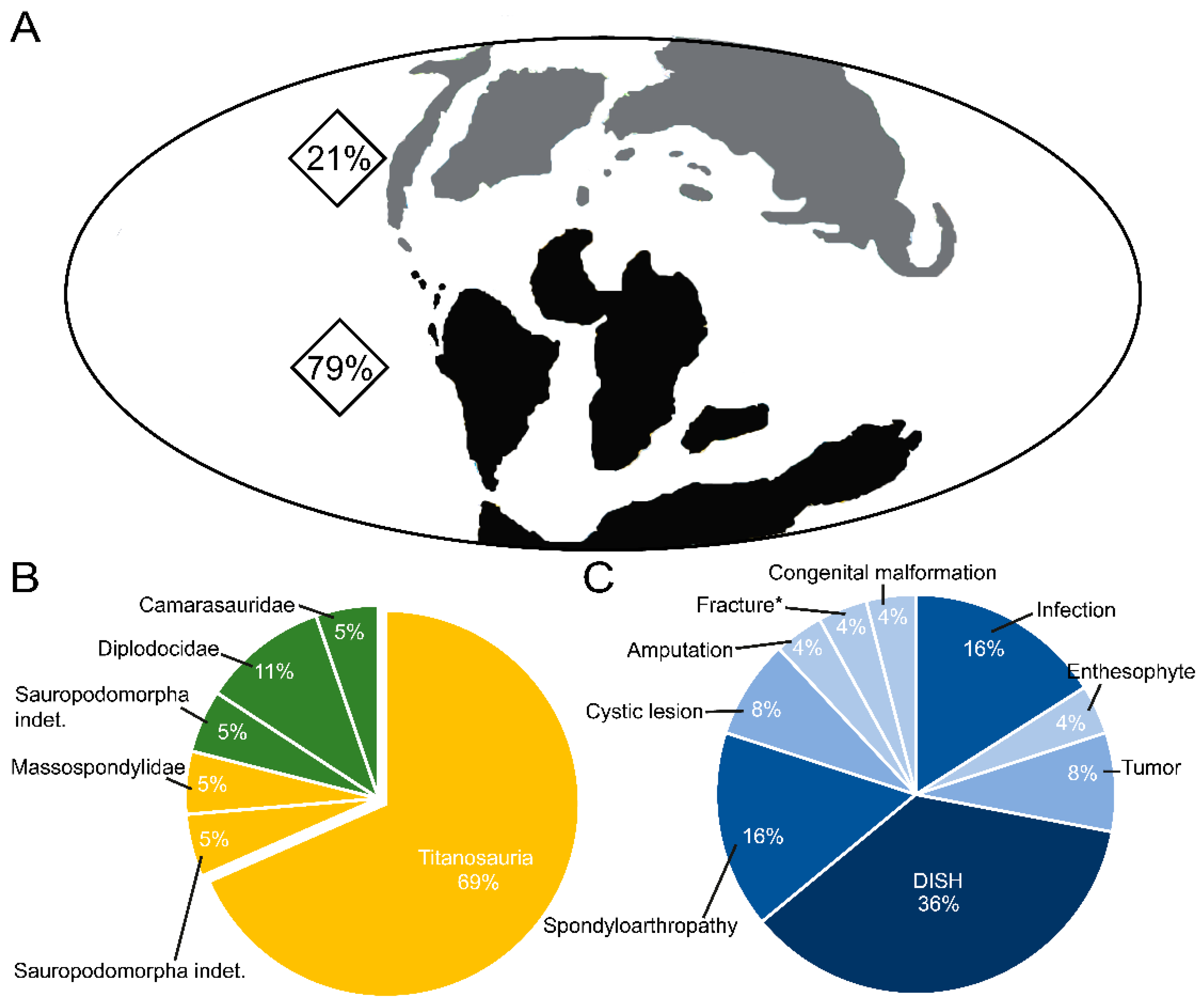
5.1. The Paleopathological Record of Sauropod Tails
5.2. Paleobiological Implications of the Presence of Paleopathologies on Sauropod Tails
6. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Sander, P.M.; Christian, A.; Clauss, M.; Fechner, R.; Gee, C.T.; Griebeler, E.M.; Gunga, H.C.; Hummel, J.; Mallison, H.; Perry, S.F.; et al. Biology of the sauropod dinosaurs: The evolution of gigantism. Biol. Rev. Camb. Philos. Soc. 2011, 86, 117–155. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cerda, I.; Paulina Carabajal, A.; Salgado, L.; Coria, R.A.; Reguero, M.A.; Tambussi, C.P.; Moly, J.J. The first record of a sauropod dinosaur from Antarctica. Sci. Nat. 2012, 99, 83–87. [Google Scholar] [CrossRef] [PubMed]
- Upchurch, P.; Barrett, P.M.; Dodson, P. Sauropoda. In The Dinosauria, 2nd ed.; Weishampel, D.B., Dodson, P., Osmólska, H., Eds.; University of California Press: Berkeley, CA, USA, 2004; pp. 295–322. [Google Scholar]
- Barrett, P.M.; Benson, R.B.J.; Upchurch, P. Dinosaur of Dorset: Part II, the Sauropod dinosaur (Saurischia, Sauropoda) with additional comments on the theropods. Proc. Dorset Nat. Hist. Archaeol. Soc. 2010, 131, 113–126. [Google Scholar]
- Bonaparte, J.F. The early radiation and phylogenetic relationships of the Jurassic sauropod dinosaurs, based on vertebral anatomy. In The Beginning of the Age of Dinosaurs; Padian, K., Ed.; Cambridge University Press: Cambridge, UK, 1986; pp. 247–258. [Google Scholar]
- Powell, J.E. Revision of South American Titanosaurid dinosaururs: Palaeobiological, palaeobiogeographical and phylogenetic aspects. In Records of the Queen Victoria Museum; Queen Victoria Museum and Art Gallery: Launceston, Australia, 2003; 173p. [Google Scholar]
- Mannion, P.D.; Upchurch, P.; Jin, X.; Zheng, W. New information on the Cretaceous sauropod dinosaurs of Zhejiang Province, China: Impact on Laurasian titanosauriform phylogeny and biogeography. R. Soc. Open Sci. 2019, 6, 191057. [Google Scholar] [CrossRef] [Green Version]
- Filippi, L.S.; Salgado, L.; Garrido, A.C. A new giant basal titanosaur sauropod in the Upper Cretaceous (Coniacian) of the Neuquén Basin, Argentina. Cretac. Res. 2019, 100, 61–81. [Google Scholar] [CrossRef]
- Hechenleitner, E.M.; Leuzinger, L.; Martinelli, A.G.; Rocher, S.; Fiorelli, L.E.; Taborda, J.R.A.; Salgado, L. Two Late Cretaceous sauropods reveal titanosaurian dispersal across South America. Commun. Biol. 2020, 3, 622. [Google Scholar] [CrossRef]
- Gallina, P.A.; Canale, J.L.; Carballido, J.L. The earliest known titanosaur sauropod dinosaur. Ameghiniana 2021, 58, 35–51. [Google Scholar] [CrossRef]
- Pérez-Moreno, A.; Carballido, J.l.; Otero, A.; Salgado, L.; Calvo, J.O. The axial skeleton of Rinconsaurus caudamirus (Sauropoda: Titanosauria) form the Late Cretaceous of Patagonia, Argentina. Ameghiniana 2022, 59, 1–46. [Google Scholar]
- Paulina-Carabajal, A. Neuroanatomy of titanosaurid dinosaurs from the Upper Cretaceous of Patagonia, with comments on endocranial variability within Sauropoda. Anat. Rec. 2012, 295, 2141–2156. [Google Scholar] [CrossRef]
- Paulina-Carabajal, A.; Filippi, L.S.; Knoll, F. Neuroanatomy of the titanosaurian sauropod Narambuenatitan palomoi from the Upper Cretaceous of Patagonia, Argentina. Publicación De La Electrónica De La Asoc. Paleontológica Argent. 2020, 20, 1–9. [Google Scholar]
- García, R.A.; Salgado, L.; Fernández, M.S.; Cerda, I.A.; Paulina Carabajal, A.; Otero, A.; Coria, R.A.; Fiorelli, L.E. Paleobiology of Titanosaurs: Reproduction, development, histology, pneumaticity, locomotion and neuroanatomy from the South American fossil record. Ameghiniana 2015, 52, 29–68. [Google Scholar] [CrossRef] [Green Version]
- Cerda, I.; Chinsamy, A.; Pol, D.; Apaldetti, C.; Otero, A.; Powell, J.E.; Martínez, R.N. Novel insight into the origin of the gorwth dynamic of saurpod dinosaurs. PLoS ONE 2017, 12, e0179707. [Google Scholar] [CrossRef] [Green Version]
- González, R.; Cerda, I.A.; Filippi, L.S.; Salgado, L. Early growth dynamics of titanosaur sauropods inferred from bone histology. Palaeogeogr. Palaeoclimatol. Palaeoecol. 2019, 537, 109404. [Google Scholar] [CrossRef]
- da Silva Vidal, L.; Luiz Gomes da Costa Pereira, P.V.; Tavares, S.; Brusatte, S.L.; Paglarelli Bergqvist, L.; dos Anjos Candeiro, C.R. Investigating the enigmatic Aeolosaurini clade: The caudal biomechanics of Aeolosaurus maximus (Aeolosaurini/Sauropoda) using the neutral pose method and the first case of protonic tail condition in Sauropoda. Hist. Biol. 2020, 33, 1836–1856. [Google Scholar] [CrossRef]
- Rothschild, B.M. Scientifically rigorous reptile and amphibian osseous pathology: Lessons for forensic herpetology from comparative and paleoepathology. Appl. Herpetol. 2009, 6, 47–79. [Google Scholar] [CrossRef]
- Arbour, V.M.; Currie, P.J. Tail and pelvis pathologies of ankylosaurian dinosaurs. Hist. Biol. 2011, 23, 375–390. [Google Scholar] [CrossRef]
- Rothschild, B.M.; Schultze, H.P.; Pellegrini, R. (Eds.) Herpetological Osteopathology: Annotated Bibliography of Amphibians and Reptiles; Springer Science: New York, NY, USA, 2012. [Google Scholar]
- Peterson, J.E.; Vittore, C.P. Cranial Pathologies in a Specimen of Pachycephalosaurus. PLoS ONE 2012, 7, e36227. [Google Scholar] [CrossRef] [Green Version]
- Tanke, D.H.; Rothschild, B.M. Paleopathology in Late Cretaceous Hadrosauridae from Alberta, Canada with comments on a putative Tyrannosaurus bite injury on an Edmontosaurus tail. In Hadrosaurs; Eberth, D.A., Evans, D.C., Eds.; Indiana University Press: Bloomington, IN, USA, 2014; pp. 540–572. [Google Scholar]
- Kappelman, J.; Ketcham, R.A.; Pearce, S.; Todd, L.; Akins, W.; Colbert, M.W.; Feseha, M.; Maisano, J.A.; Witzel, A. Perimortem fractures in Lucy suggest mortality from fall out of tall tree. Nature 2016, 537, 503. [Google Scholar] [CrossRef]
- Dumbrava, M.D.; Rothschild, B.M.; Weishampel, D.B.; Csiki-Sava, Z.; Andrei, R.A.; Acheson, K.A.; Codrea, V.A. A dinosaurian facial deformity and the first occurrence of ameloblastoma in the fossil record. Sci. Rep. 2016, 6, 29271. [Google Scholar] [CrossRef]
- Hearn, L.; Williams, A.C. Pain in dinosaurs: What is the evidence? Philos. Trans. R. Soc. B 2019, 374, 20190370. [Google Scholar] [CrossRef]
- Cruzado-Caballero, P.; Díaz-Martínez, I.; Rothschild, B.; Bedell, M.; Pereda-Suberbiola, X. A limping dinosaur in the Late: Pathologies in the pes of the neornithischian Othnielosaurus consors from the Morrison Formation (Upper Jurassic, USA). Hist. Biol. 2020, 33, 1753–1759. [Google Scholar] [CrossRef]
- Cruzado-Caballero, P.; Lecuona, A.; Cerda, I.A.; Díaz-Martínez, I. Osseous paleopathologies of Bonapartesaurus rionegrensis (Ornithopoda, Hadrosauridae) from Allen Formation (Upper Cretaceous) of Patagonia Argentina. Cretac. Res. 2021, 124, 104800. [Google Scholar] [CrossRef]
- Martinelli, A.G.; Teixeira, V.P.A.; Marinho, T.S.; Fonseca, P.H.M.; Cavellani, C.L.; Araujo, A.J.G.; Ribeiro, L.C.B.; Ferraz, M.L.F. Fused mid-caudal vertebrae in the titanosaur Uberabatitan ribeiroi from the Late Cretaceous of Brazil and other bone lesions. Lethaia 2015, 48, 456–462. [Google Scholar] [CrossRef]
- García, R.A.; Cerda, I.A.; Heller, M.; Rothschild, B.M.; Zurriaguz, V. The first evidence of osteomyelitis in a sauropod dinosaur. Lethaia 2016, 50, 227–236. [Google Scholar] [CrossRef]
- González, R.; Gallina, P.A.; Cerda, I.A. Multiple paleopathologies in the dinosaur Bonitasaura salgadoi (Sauropoda: Titanosauria) from the Upper Cretaceous of Patagonia, Argentina. Cretac. Res. 2017, 79, 159–170. [Google Scholar] [CrossRef]
- Clayton, R.J. Description of unusual pathological disorders on pubes and associated left femur from a Diplodocus specimen. Paludicola 2018, 11, 179–187. [Google Scholar]
- Tschopp, E.; Wings, O.; Frauenfelder, T.; Rothschild, B.M. Pathological phalanges in a camarasaurid sauropod dinosaur and implications on behaviour. Acta Palaeontol. Pol. 2016, 61, 125–134. [Google Scholar] [CrossRef] [Green Version]
- Brum, A.S.; Bandeira, K.L.; Holgado, B.; Souza, L.G.; Pêgas, R.V.; Sayão, J.M.; Campos, D.A.; Kellner, A.W. Palaeohistology and palaeopatology of an Aeolosaurini (Sauropoda: Titanosauria) from Morro do Cambambe (Upper Cretaceous, Brazil). [Paleohistología y paleopatología de un Aelosaurini (Sauropoda: Titanosauria) de Morro do Cambambe (Cretácico Superior, Brasil)]. Span. J. Palaeontol. 2021, 36, 1–17. [Google Scholar]
- Garrido, A. Estratigrafía del Grupo Neuquén, Cretácico Superior de la Cuenca Neuquina (Argentina): Nueva propuesta de ordenamiento litoestratigráfico. Rev. Del Mus. Argent. De Cienc. Nat. 2010, 12, 121–177. [Google Scholar] [CrossRef] [Green Version]
- Leanza, H.A.; Hugo, C.A. Cretaceous red beds from southern Neuquén Basin (Argentina): Age, distribution and stratigraphic discontinuities. Publicación Electrónica De La Asoc. Paleontológica Argent. 2001, 7, 117–122. [Google Scholar]
- Filippi, L.S.; García, R.A.; Garrido, A. A new titanosaur sauropod dinosaur from Upper Cretaceous of North Patagonia, Argentina. Acta Palaeontol. Pol. 2011, 56, 505–520. [Google Scholar] [CrossRef] [Green Version]
- Filippi, L.S.; Garrido, A. Pitekunsaurus macayai gen. et sp. nov., nuevo titanosaurio (Saurischia, Sauropoda) del Cretácico Superior de la Cuenca Neuquina, Argentina. Ameghiniana 2008, 45, 575–590. [Google Scholar]
- Gallina, P.A.; Apesteguía, S. Postcranial anatomy of Bonitasaura salgadoi (Sauropoda, Titanosauria) from the Late Cretaceous of Patagonia. J. Vertebr. Paléontol. 2015, 35, e924957. [Google Scholar] [CrossRef]
- Mahajan Mahajan, T.; Ganguly, S.; Para, P.A. Fracture Management in Animals: A Review. J. Chem. Biol. Phys. Sci. 2015, 5, 4053–4057. [Google Scholar]
- Rothschild, B.M.; Martin, L.D. Skeletal Impact of Disease: Bulletin 33. In New Mexico Museum of Natural History and Science; Authority of the State of New Mexico: Alburquerque, NM, USA, 2006; Volume 33. [Google Scholar]
- de Sousa Barbosa, F.H.; Porpino, K.D.O.; Rothschild, B.M.; Silva, R.C.D.; Capone, D. First cancer in an extinct Quaternary non-human mammal. Hist. Biol. 2020, 33, 2878–2882. [Google Scholar] [CrossRef]
- Jacobson, E.R. (Ed.) Infectious Diseases and Pathology of Reptiles. In Viruses and Viral Diseases of Reptiles; CRC Press: Boca Raton, FL, USA, 2007; pp. 395–460. [Google Scholar]
- Anné, J.; Garwood, R.J.; Lowe, T.; Withers, P.J.; Manning, P.L. Interpreting pathologies in extant and extinct archosaurs using micro-CT. PeerJ 2015, 3, e1130. [Google Scholar] [CrossRef] [Green Version]
- Hanna, R.R. Multiple injury and infection in a sub-adult theropod dinosaur Allosaurus fragilis with comparisons to Allosaurus pathology in the Clevland-Lloyd Quarry collection. J. Vertebr. Paleontol. 2002, 22, 76–90. [Google Scholar] [CrossRef]
- Ramírez-Velasco, A.A.; Morales-Salinas, E.; Hernández-Rivera, R.; Tanke, D.H. Spinal and rib osteopathy in Huehuecanauhtlus tiquichensis (Ornithopoda: Hadrosauroidea) from the Late Cretaceous in Mexico. Hist. Biol. 2017, 29, 208–222. [Google Scholar] [CrossRef]
- Resnick, D.; Niwayama, G. Diagnosis of Bone and Joint Disorders; WB Saunders Co.: Philadelphia, PA, USA, 1981; Volume 3, pp. 2058–2079. [Google Scholar]
- de Souza Barbosa, F.H.; da Costa Ribeiro, I.; da Costa, P.V.L.G.; Bergqvist, L.P. Vertebral lesions in a titanosaurian dinosaur from the Lower-Upper Cretaceous of Brazil. Geobios 2018, 51, 385–389. [Google Scholar] [CrossRef]
- Mader, R.; Sarzi-Puttini, P.; Atzeni, F.; Olivieri, I.; Pappone, N.; Verlaan, J.J.; Buskila, D. Extraspinal manifestations of diffuse idiopathic skeletal hyperostosis. Rheumatology 2009, 48, 1478–1481. [Google Scholar] [CrossRef] [Green Version]
- Cisneros, J.C.; Gomes Cabral, U.; de Beer, F.; Damiani, R.; Costa Fortier, D. Spondarthritis in the Triassic. PLoS ONE 2010, 5, e13425. [Google Scholar] [CrossRef] [PubMed]
- Witzmann, F.; Schwarz-Wings, D.; Hampe, O.; Fritsch, G.; Asbach, P. Evidence of spondyloarthropathy in the spine of a phytosaur (Reptilia: Archosauriformes) from the Late Triassic of Halberstadt, Germany. PLoS ONE 2014, 9, e85511. [Google Scholar] [CrossRef] [PubMed]
- Gallina, P.A. Histología ósea del titanosaurio Bonitasaura salgadoi (Dinosauria: Sauropoda) del Cretácico superior de Patagonia. Ameghiniana 2012, 49, 289–302. [Google Scholar] [CrossRef]
- Rothschild, B.M.; Berman, D.S. Fusion of caudal vertebrae in Late Jurassic sauropods. J. Vertebr. Paleontol. 1991, 11, 29–36. [Google Scholar] [CrossRef]
- Conti, S.; Tschopp, E.; Mateus, O.; Zanoni, A.; Masarati, P.; Sala, G. Multibody analysis and soft tissue strength refute supersonic dinosaur tail. Sci. Rep. 2022, 12, 19245. [Google Scholar] [CrossRef] [PubMed]
- Butler, R.J.; Yates, A.M.; Rauhut, O.W.; Foth, C. A pathological tail in a basal sauropodomorph dinosaur from South Africa: Evidence of traumatic amputation? J. Vertebr. Paleontol. 2013, 33, 224–228. [Google Scholar] [CrossRef]
- Xing, L.; Rothschild, B.M.; Ran, H.; Miyashita, T.; Persons, W.S.; Sekiya, T.; Zhang, J.; Tao Wang, T.; Dong, Z. Vertebral fusion in two Early Jurassic sauropodomorph dinosaurs from the Lufeng Formation of Yunnan, China. Acta Palaeontol. Pol. 2015, 60, 643–649. [Google Scholar] [CrossRef] [Green Version]
- Weston, D.A. Nonspecific infection in paleopathology: Interpreting periosteal reactions. In A Companion to Paleopathology; Grauer, A.L., Ed.; Wiley-Blackwell: Oxford, UK, 2012; pp. 492–512. [Google Scholar]
- Díez Díaz, V.; Demuth, O.E.; Schwarz, D.; Mallison, H. The tail of the Late Jurassic sauropod Giraffatitan brancai: Digital reconstruction of its epaxial and hypaxial musculature, and implications for tail biomechanics. Front. Earth Sci. 2020, 8, 160. [Google Scholar] [CrossRef]
- Ibiricu, L.M.; Lamanna, M.C.; Lacovara, K.J. The influence of caudofemoral musculature on the titanosaurian (Saurischia: Sauropoda) tail skeleton: Morphological and phylogenetic implications. Hist. Biol. 2014, 26, 454–471. [Google Scholar] [CrossRef]
- Otero, A.; Gallina, P.A.; Canale, J.I.; Haluza, A. Sauropod haemal arches: Morphotypes, new classification and phylogenetic aspects. Hist. Biol. 2012, 24, 243–256. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| DISEASE | TOTAL |
|---|---|
| Gondwana | |
| Infection | 4 |
| Enthesophyte | 1 |
| Tumor | 2 |
| DISH | 1 |
| Spondyloarthropathy | 3 |
| Cystic lesion | 2 |
| Traumatism | 1 |
| Fracture * | 1 |
| Total | 16 |
| Laurasia | |
| Spondylloartrhopathy | 1 |
| DISH | 8 |
| Congenital malformation | 1 |
| Total | 10 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Cruzado-Caballero, P.; Filippi, L.S.; González-Dionis, J.; Canudo, J.I. How Common Are Lesions on the Tails of Sauropods? Two New Pathologies in Titanosaurs from the Late Cretaceous of Argentine Patagonia. Diversity 2023, 15, 464. https://doi.org/10.3390/d15030464
Cruzado-Caballero P, Filippi LS, González-Dionis J, Canudo JI. How Common Are Lesions on the Tails of Sauropods? Two New Pathologies in Titanosaurs from the Late Cretaceous of Argentine Patagonia. Diversity. 2023; 15(3):464. https://doi.org/10.3390/d15030464
Chicago/Turabian StyleCruzado-Caballero, Penélope, Leonardo Sebastián Filippi, Javier González-Dionis, and José Ignacio Canudo. 2023. "How Common Are Lesions on the Tails of Sauropods? Two New Pathologies in Titanosaurs from the Late Cretaceous of Argentine Patagonia" Diversity 15, no. 3: 464. https://doi.org/10.3390/d15030464