
One of the Deepest Genera of Antipatharia: Taxonomic Position Revealed and Revised

 , , ,
, , ,
Abstract
:1. Introduction
2. Materials and Methods
3. Results
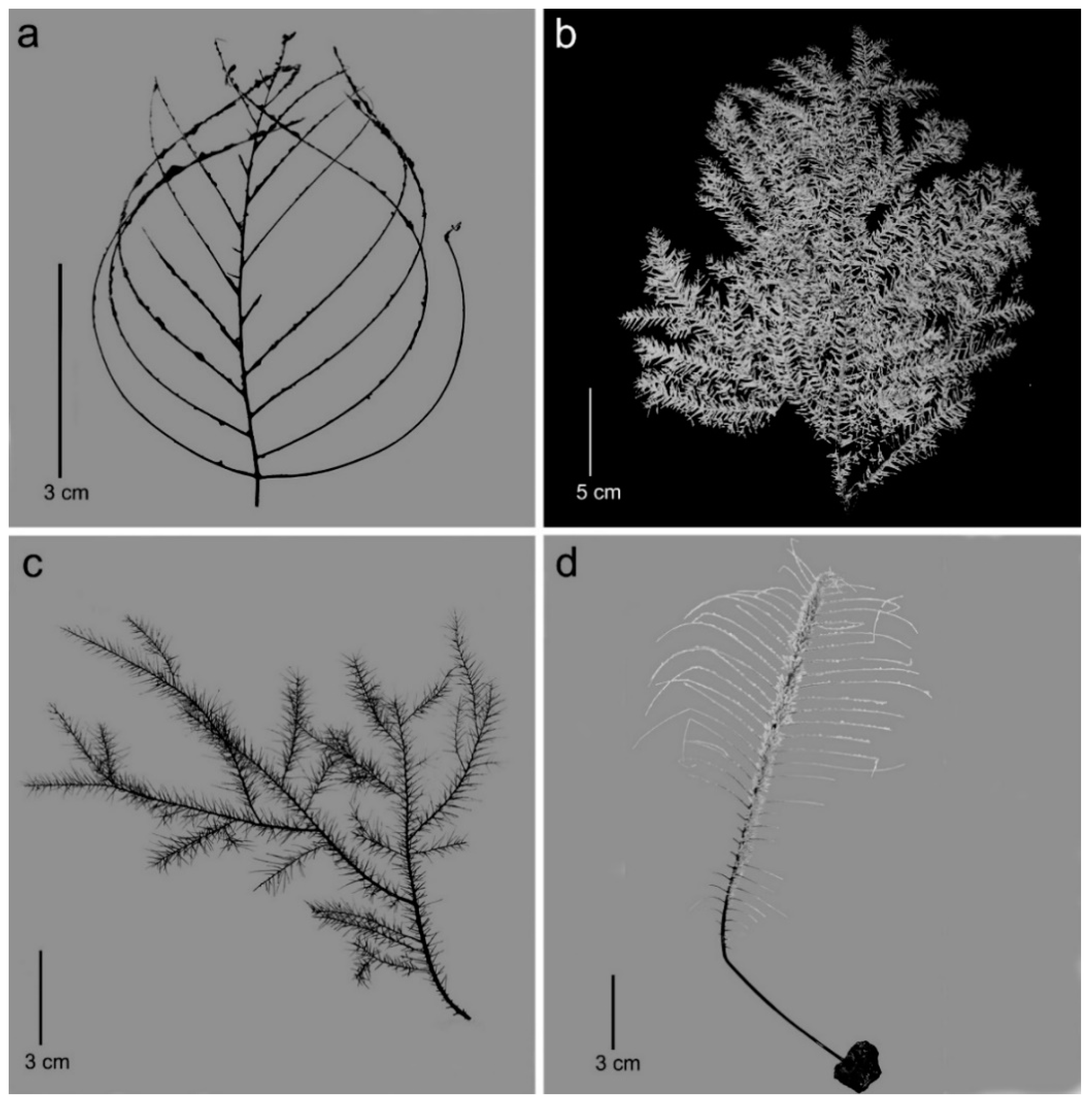
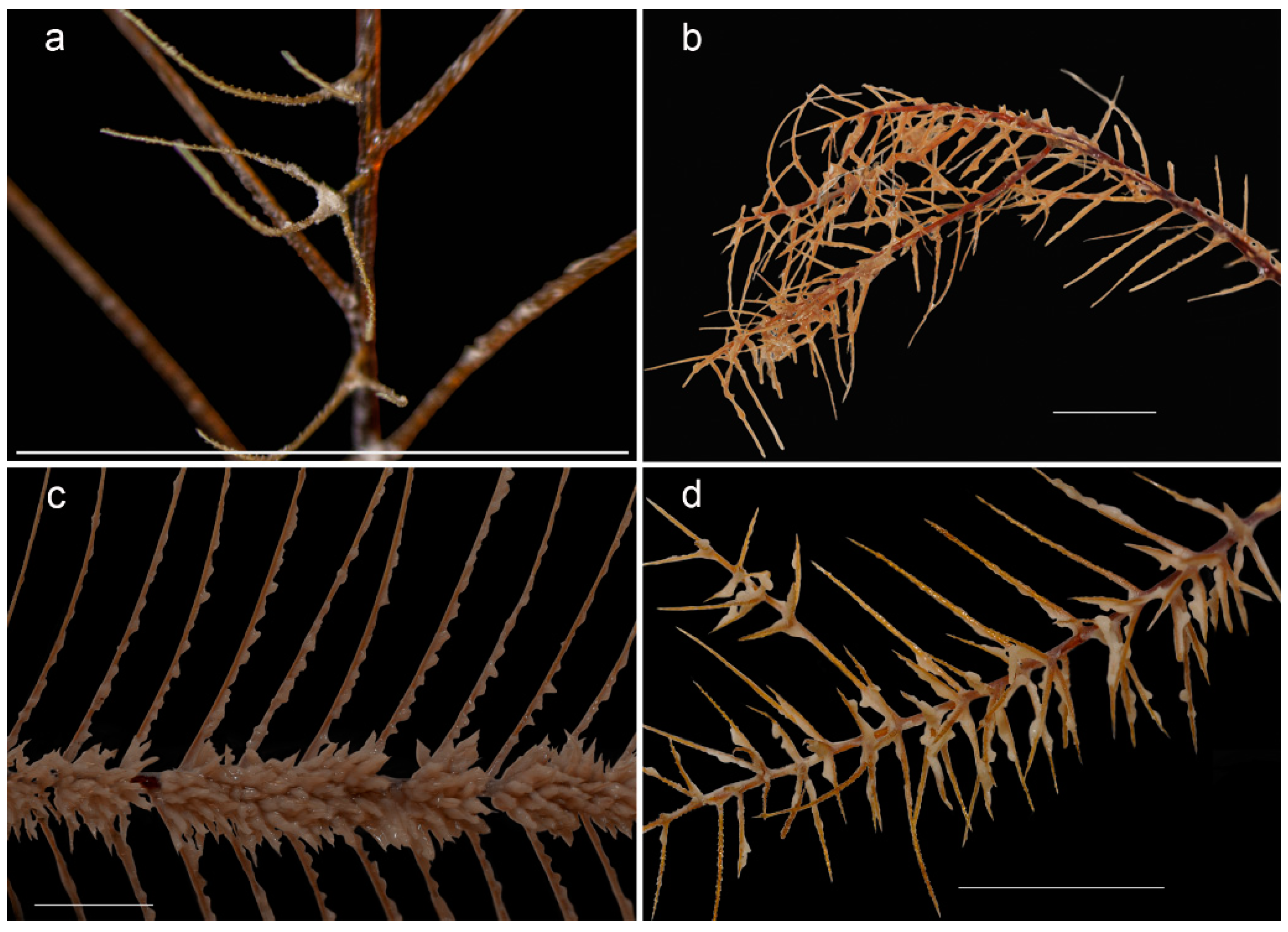
3.1. Morphological Analysis
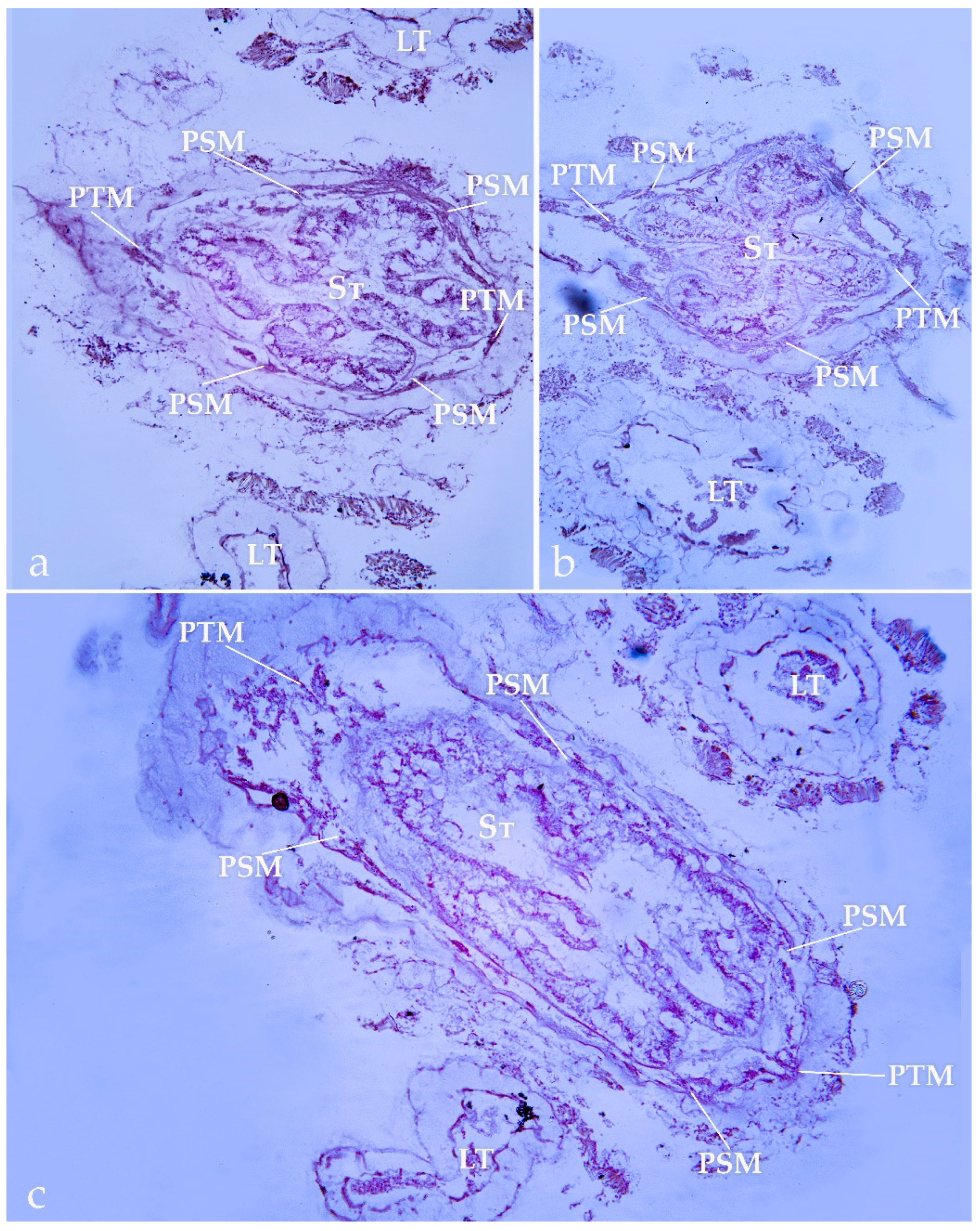
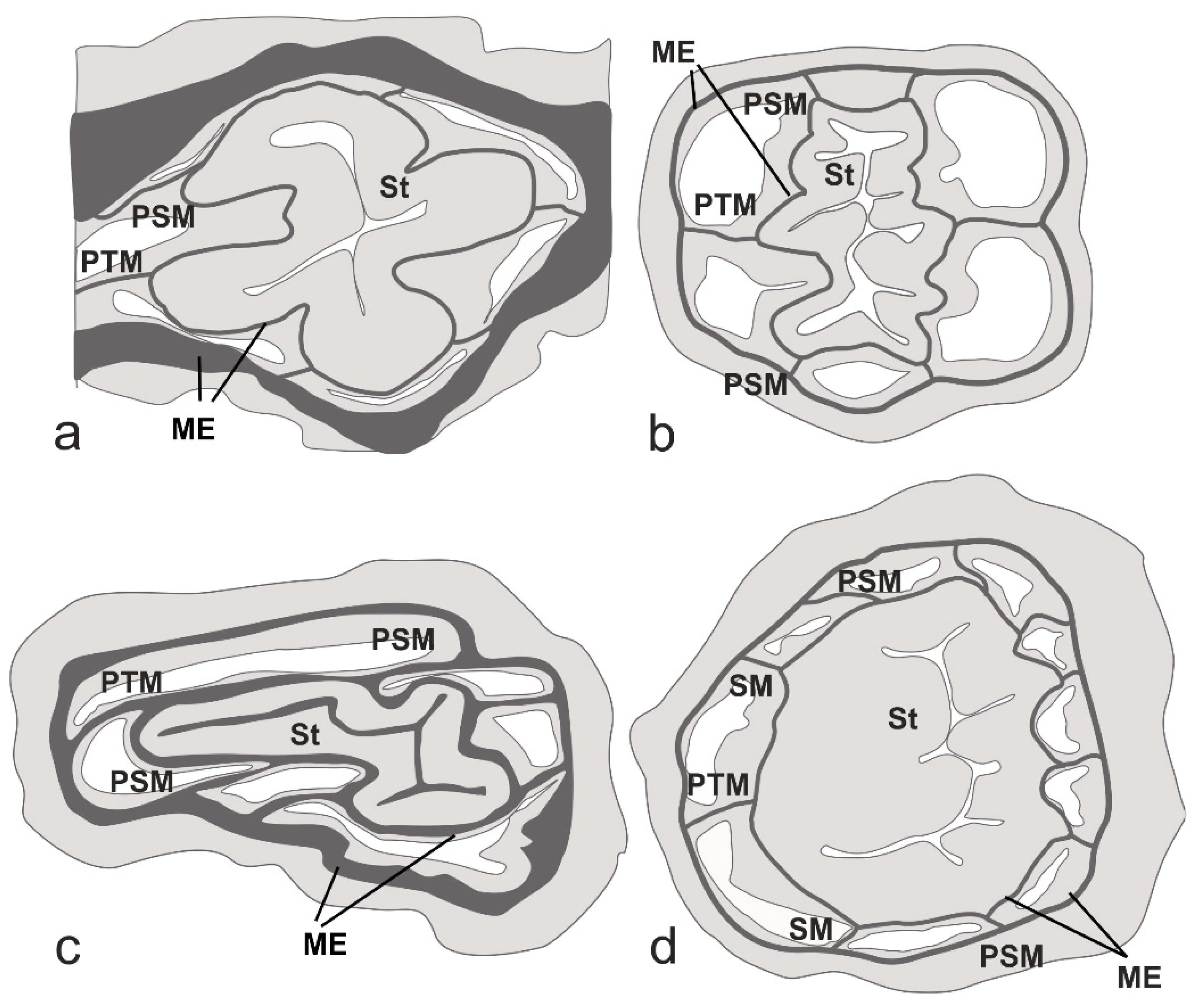
3.2. Histological Analysis
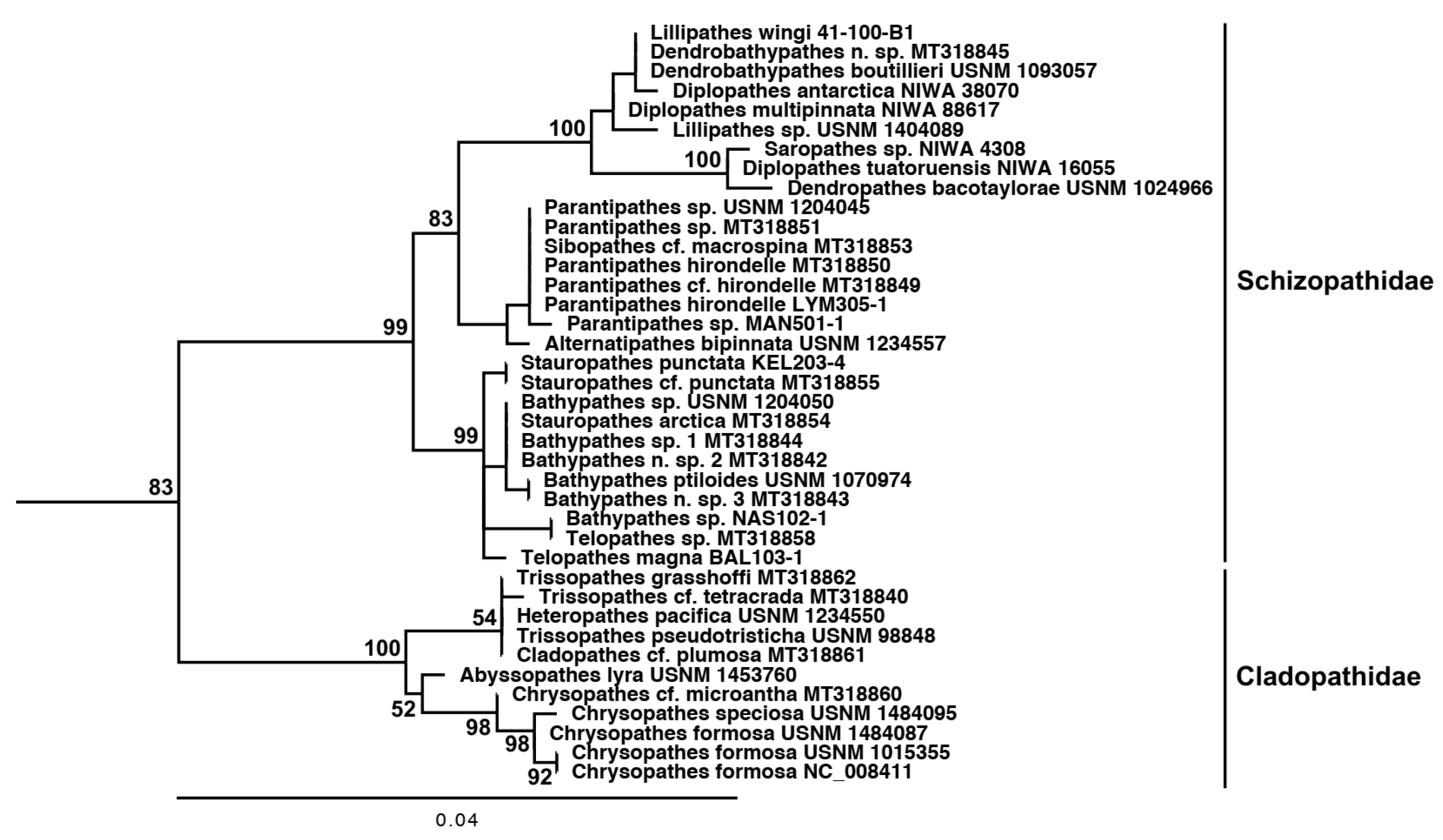
3.3. Molecular Analysis
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Cairns, S.D. Deep-water corals: An overview with special reference to diversity and distribution of deep-water scleractinian corals. Bull. Mar. Sci. 2007, 81, 311–322. [Google Scholar]
- Wagner, D.; Luck, D.G.; Toonen, R.J. The biology and ecology of black corals (Cnidaria: Anthozoa: Hexacorallia: Antipatharia). Adv. Mar. Biol. 2012, 63, 67–132. [Google Scholar] [CrossRef] [PubMed]
- Molodtsova, T.N.; Opresko, D.M. Black corals (Anthozoa: Antipatharia) of the Clarion-Clipperton Fracture Zone. Mar. Biodivers. 2017, 47, 349–365. [Google Scholar] [CrossRef]
- Bo, M.; Montgomery, A.D.; Opresko, D.M.; Wagner, D.; Bavestrello, G. Antipatharians of the mesophotic zone: Four case studies; Coral Reefs of the World. In Mesophotic Coral Ecosystems; Loya, Y.P., Bridge, T.K., Eds.; Springer: Cham, Switzerland, 2019; pp. 683–708. [Google Scholar]
- Molodtsova, T.N.; Opresko, D.M.; Wagner, D. Description of a new and widely distributed species of Bathypathes (Cnidaria: Anthozoa: Antipatharia: Schizopathidae) previously misidentified as Bathypathes alternata Brook, 1889. PeerJ 2022, 10, e12638. [Google Scholar] [CrossRef] [PubMed]
- Brugler, M.R.; Opresko, D.M.; France, S.C. The evolutionary history of the order Antipatharia (Cnidaria: Anthozoa: Hexacorallia) as inferred from mitochondrial and nuclear DNA: Implications for black coral taxonomy and systematics. Zool. J. Linn. Soc. 2013, 169, 312–361. [Google Scholar] [CrossRef] [Green Version]
- Barrett, N.J.; Hogan, R.I.; Allcock, A.L.; Molodtsova, T.; Hopkins, K.; Wheeler, A.J.; Yesson, C. Phylogenetics and mitogenome organisation in black corals (Anthozoa: Hexacorallia: Antipatharia): An order-wide survey inferred from complete mitochondrial genomes. Front. Mar. Sci. 2020, 7, 440. [Google Scholar] [CrossRef]
- Opresko, D.M. A new species of Sibopathes (Cnidaria: Anthozoa: Antipatharia: Antipathidae) from the Gulf of Mexico. Proc. Biol. Soc. Wash. 1993, 106, 195–203. [Google Scholar]
- Chimienti, G.; Terraneo, T.I.; Vicario, S.; Marchese, F.; Purkis, S.J.; Abdulla Eweida, A.; Rodrigue, M.; Benzoni, F. A new species of Bathypathes (Cnidaria, Anthozoa, Antipatharia, Schizopathidae) from the Red Sea and its phylogenetic position. ZooKeys 2022, 1116, 1–22. [Google Scholar] [CrossRef]
- Opresko, D.M. Review of the genus Schizopathes (Cnidaria: Antipatharia: Schizopathidae) with a description of a new species from the Indian Ocean. Proc. Biol. Soc. Wash. 1997, 110, 157–166. [Google Scholar]
- Brook, G. Report on the Antipatharia collected by HMS Challenger during the years 1873–1876. In Report on the Scientific Results of the Voyage of HMS “Challenger” during the Years 1873–76 under the Command of Captain George S. Nares, R.N., F.R.S. and the Late Captain Frank Tourle Thomson, R.N., Zoology. XXII; Thomson, S.C.W., Murray, J., Eds.; HM Stationery Office: London, UK; Edinburgh, UK; Dublin, UK, 1889. [Google Scholar]
- Opresko, D.M. Revision of the Antipatharia (Cnidaria: Anthozoa). Part II. Schizopathidae. Zool. Meded. 2002, 76, 411–422. [Google Scholar]
- Horowitz, J.; Opresko, D.M.; Bridge, T.C.L. Black corals (Anthozoa: Antipatharia) from the deep (916 m-2542 m) Coral Sea, north-eastern Australia. Zootaxa 2018, 4472, 307–326. [Google Scholar] [CrossRef]
- Pasternak, F.A. Antipatharia; Elsevier: Copenhagen, Denmark, 1977; pp. 164–167. [Google Scholar]
- Kennedy, B.R.C.; Cantwell, K.; Malik, M.; Kelley, C.; Potter, J.; Elliott, K.; Lobecker, E.; Gray, L.M.; Sowers, D.; White, M.P.; et al. The unknown and the unexplored: Insights into the Pacific Deep-Sea following NOAA CAPSTONE expeditions. Front. Mar. Sci. 2019, 6, 480. [Google Scholar] [CrossRef] [Green Version]
- Opresko, D.M. Revision of the Antipatharia. (Cnidaria: Anthozoa). Part III. Cladopathidae. Zool. Meded. 2003, 77, 495–536. [Google Scholar]
- Opresko, D.M. New genera and species of antipatharian corals (Cnidaria: Anthozoa) from the North Pacific. Zool. Meded. 2005, 79, 129–165. [Google Scholar]
- Molodtsova, T.N. New species of Hexapathes Kinoshita, 1910 (Anthozoa, Antipatharia, Cladopathidae) from the South-West Pacific. Zoosystema 2006, 28, 597–606. [Google Scholar]
- Opresko, D.M.; Loyola, L.L. Two new species of Chrysopathes (Cnidaria: Anthozoa: Antipatharia) from the Western Atlantic. Zootaxa 2008, 1707, 49–59. [Google Scholar]
- Opresko, D.M.; Molodtsova, T.N. New species of deep-sea Antipatharians from the North Pacific (Cnidaria: Anthozoa: Antipatharia), Part 2. Zootaxa 2021, 4999, 401–422. [Google Scholar] [CrossRef]
- Opresko, D.M.; Stewart, R.; Voza, T.; Tracey, D.; Brugler, M.R. New genus and species of black coral from the SW Pacific and Antarctica (Cnidaria: Anthozoa: Antipatharia: Schizopathidae). Zootaxa 2022, 5169, 31–48. [Google Scholar] [CrossRef]
- Opresko, D.; Tracey, D.M.; Mackay, E. Antipatharia (Black Corals) for the New Zealand Region: A Field Guide of Commonly Sampled New Zealand Black corals INCLUDING Illustrations Highlighting Technical Terms and Black Coral Morphology; Ministry for Primary Industries: Wellington, New Zeeland, 2014; Volume 131. [Google Scholar]
- Miller, M.A.; Pfeiffer, W.; Schwartz, T. The CIPRES science gateway: A community resource for phylogenetic analyses. In Proceedings of the 2011 TeraGrid Conference: Extreme Digital Discovery, Salt Lake, UT, USA, 18–21 July 2011; pp. 1–8. [Google Scholar]
- Minh, B.Q.; Schmidt, H.A.; Chernomor, O.; Schrempf, D.; Woodhams, M.D.; Von Haeseler, A.; Lanfear, R. IQ-TREE 2: New models and efficient methods for phylogenetic inference in the genomic era. Mol. Biol. Evol. 2020, 37, 1530–1534. [Google Scholar] [CrossRef] [Green Version]
- Kumar, S.; Stecher, G.; Li, M.; Knyaz, C.; Tamura, K. MEGA X: Molecular evolutionary genetics analysis across computing platforms. Mol. Biol. Evol. 2018, 35, 1547. [Google Scholar] [CrossRef]
- Carlisle, D.B. Softening chitin for histology. Nature 1960, 187, 1132–1133. [Google Scholar] [CrossRef] [PubMed]
- Barbosa, P.; Berry, D.; Kary, C.K. Insect Histology: Practical Laboratory Techniques, 1st ed.; John Wiley & Sons, Ltd.: Hoboken, NJ, USA, 2014; p. 368. [Google Scholar]
- van Pesch, A.J. The Antipatharia of the Siboga Expedition. In Siboga-Expaditie XVII; Weber, M., Ed.; E.J. Brill: Leyden, The Netherlands, 1914; pp. 1–258. [Google Scholar]
- Wagner, D.; Waller, R.G.; Toonen, R.J. Sexual reproduction of Hawaiian black corals, with a review of the reproduction of antipatharians (Cnidaria: Anthozoa: Hexacorallia). Invertebr. Biol. 2011, 130, 211–225. [Google Scholar] [CrossRef]
- Kinoshita, K. On a new antipatharian Hexapathes heterosticha, n.g. et n.sp. Annot. Zool. Jpn. 1910, 7, 231–234. [Google Scholar]
- Pax, F. Die Antipatharien und Madreporarien des arktischen Gebietes. Fauna Arctica 1932, 6, 267–280. [Google Scholar]
- Pasternak, F. Deep-sea antipatharians of Kuril-Kamchatka Trench. Tr. Inst. Oceanol. AN SSSR. 1958, 27, 180–191. [Google Scholar]
- Molodtsova, T.N. Black corals (Antipatharia: Anthozoa: Cnidaria) of North-East Atlantic. In Biogeography of the North Atlantic seamounts; Mironov, A.N., Gebruk, A.V., Southward, A.J., Eds.; KMK Press: Moscow, Russia, 2006; pp. 141–151. [Google Scholar]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Genus | Mature Corallum Growth Form | Striatum | Primary Pinnulation | Typical Number of Rows of Primary Pinnules | Lateral or Posterolateral Pinnules | Anterior or Anterolateral Pinnules |
|---|---|---|---|---|---|---|
| Abyssopathes | monopodial | present | laterals regular; anteriors irregular | 3 to 4 | simple; basal subopposite others alternating | absent, simple or subpinnulate |
| Cladopathidae | ||||||
| Chrysopathes | branched | present | regular; in biserial groups | 5 to 6 | subpinnulate; basal subopposite, others alternating | subpinnulate |
| Cladopathes | densely branched | no data | Irregular | 4 to 5 | simple;alternating | subpinnulate |
| Heteropathes | monopodial | present | laterals regular; anteriors irregular | 3 to 4 | simple; basal subopposite, others alternating | subpinnulate |
| Hexapathes | monopodial | present | laterals regular; anteriors irregular | 3 to 4 | simple; basal subopposite others alternating | simple or subpinnulate |
| Trissopathes | laxly or densely branched | at least in some species | regular; in biserial groups | 3 to 4 | usually simple; basal subopposite others alternating | subpinnulate |
| Sibopathes | laxly branched | present | regular; in biserial groups | 4 to 6 | simple; basal subopposite others alternating | simple |
| Schizopathidae | ||||||
| Alternatipathes | monopodial or rarely branched | absent | regular | 2 | simple; all alternating | absent |
| Bathypathes | monopodial; rarely branched | present or absent | regular | 2 | simple; all subopposite or all alternating | absent |
| Dendrobathypathes | sparsely to densely branched | no data | regular | 2 | subpinnulate; all alternating | absent |
| Dendropathes | densely branched | no data | regular; in biserial groups | 4 | simple; all alternating | simple |
| Diplopathes | laxly branched | no data | regular | 2 | simple; all alternating | absent |
| Lillipathes | usually laxly branched | present | regular; in biserial groups | 4 | simple; all alternating | simple |
| Parantipathes | monopodial or laxly branched | present or absent | regular; in biserial groups | 6 to 12 | simple; all alternating | simple |
| Saropathes | monopodial? | no data | regular; in biserial groups | 4 | subpinnulate; all alternating | subpinnulate |
| Schizopathes | monopodial | absent | regular | 2 | simple; all alternating | absent |
| Stauropathes | sparsely or densely branched | at least in some species | regular | 2 | simple; all subopposite | absent |
| Taxipathes | sparsely branched | no data | regular; in biserial groups | 6 | simple; all alternating | simple |
| Telopathes | sparsely branched | no data | regular | 2 | simple; alternating or subopposite | absent |
| Umbellapathes | monopodial-like stalked with branched crown | absent | regular | 2 | simple or subpinnulate; all alternating | absent |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Molodtsova, T.N.; Opresko, D.M.; O’Mahoney, M.; Simakova, U.V.; Kolyuchkina, G.A.; Bledsoe, Y.M.; Nasiadka, T.W.; Ross, R.F.; Brugler, M.R. One of the Deepest Genera of Antipatharia: Taxonomic Position Revealed and Revised. Diversity 2023, 15, 436. https://doi.org/10.3390/d15030436
Molodtsova TN, Opresko DM, O’Mahoney M, Simakova UV, Kolyuchkina GA, Bledsoe YM, Nasiadka TW, Ross RF, Brugler MR. One of the Deepest Genera of Antipatharia: Taxonomic Position Revealed and Revised. Diversity. 2023; 15(3):436. https://doi.org/10.3390/d15030436
Chicago/Turabian StyleMolodtsova, Tina N., Dennis M. Opresko, Michael O’Mahoney, Ulyana V. Simakova, Galina A. Kolyuchkina, Yessenia M. Bledsoe, Teresa W. Nasiadka, Rachael F. Ross, and Mercer R. Brugler. 2023. "One of the Deepest Genera of Antipatharia: Taxonomic Position Revealed and Revised" Diversity 15, no. 3: 436. https://doi.org/10.3390/d15030436