
Evidence of Non-Random Social Interactions between Pairs of Bait-Attracted White Sharks in Gansbaai (South Africa)

 ,
,  , ,
, ,
Abstract
:1. Introduction
2. Materials and Methods
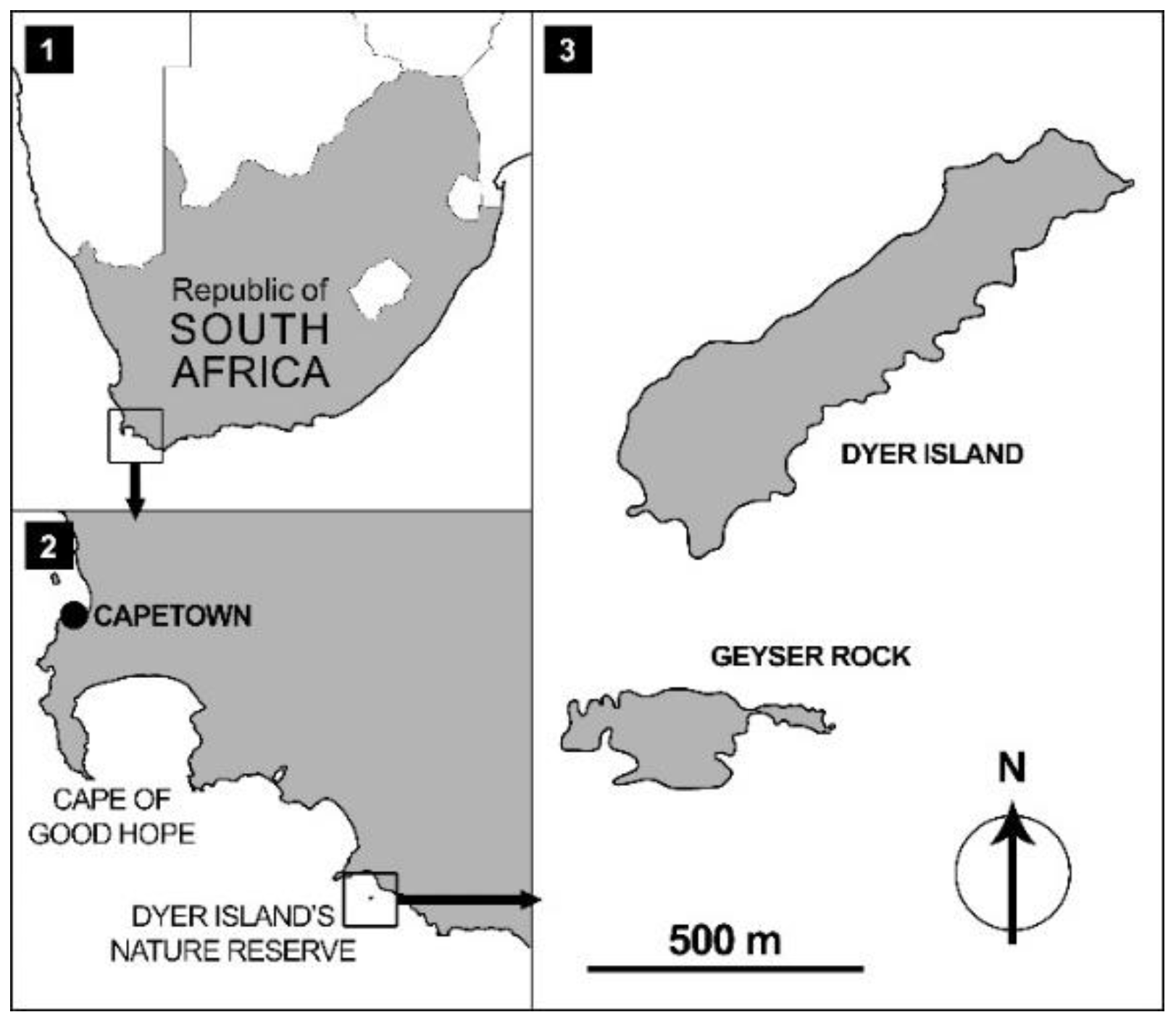
2.1. Study Area
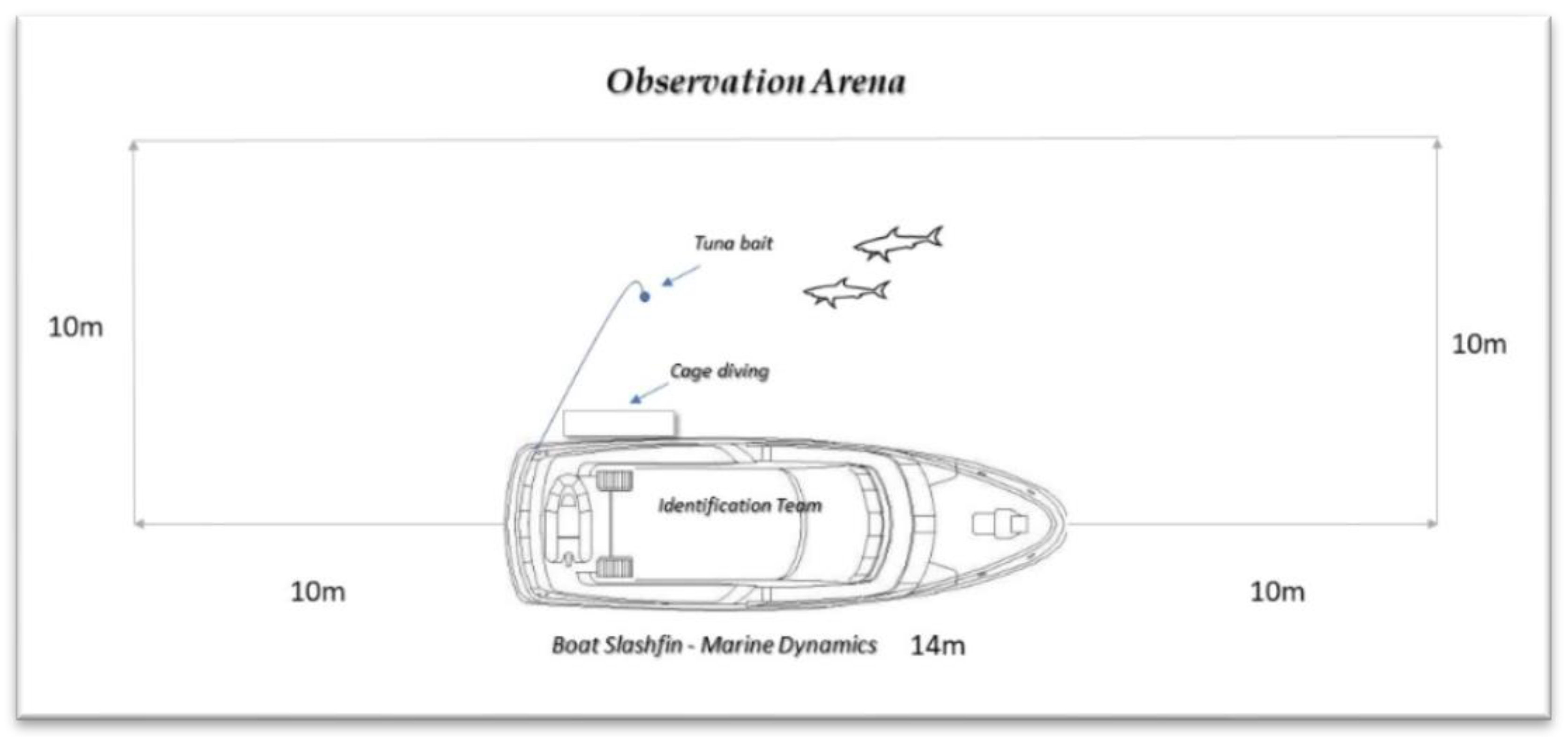
2.2. Data Collection
2.3. Statistical Analysis
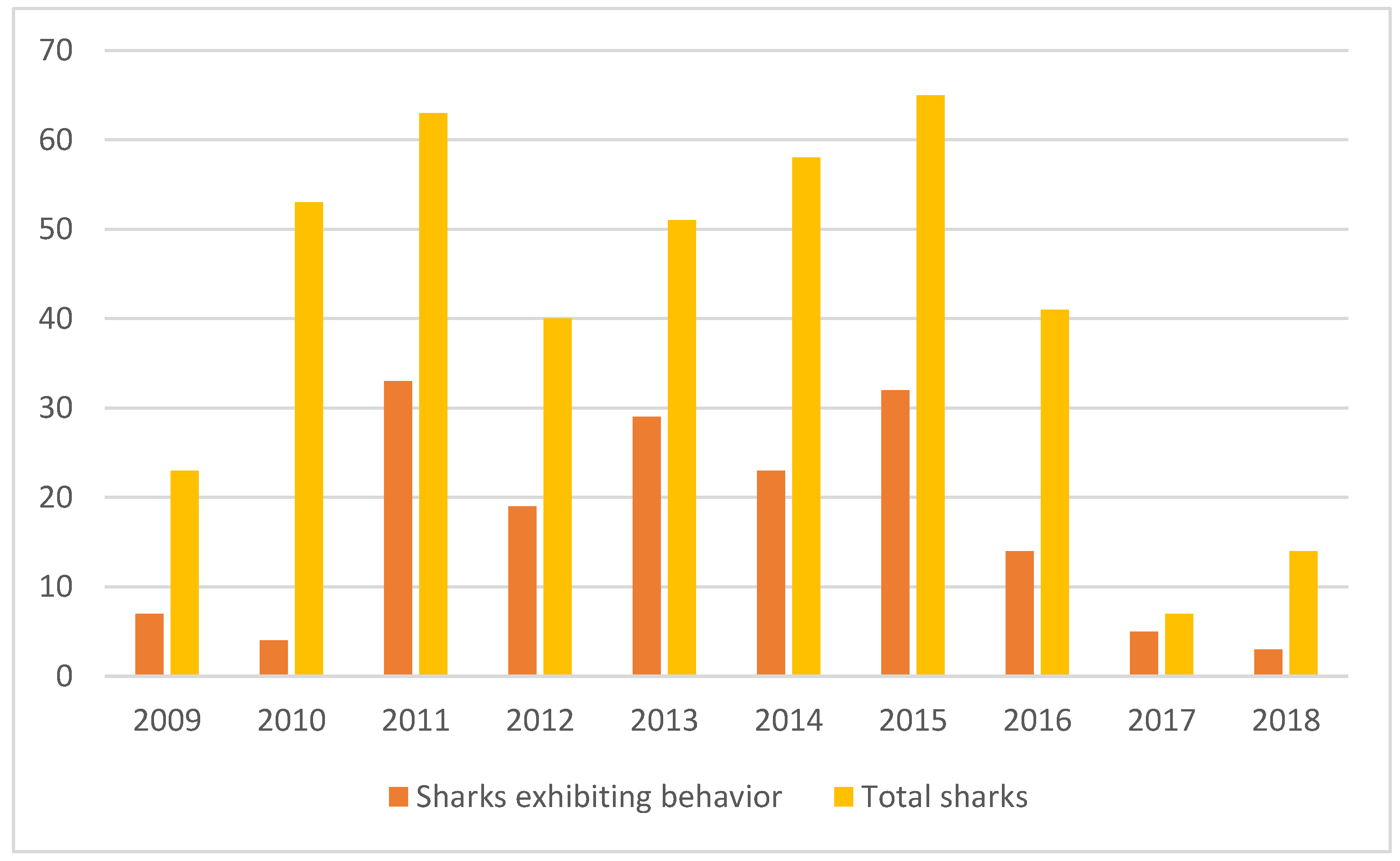
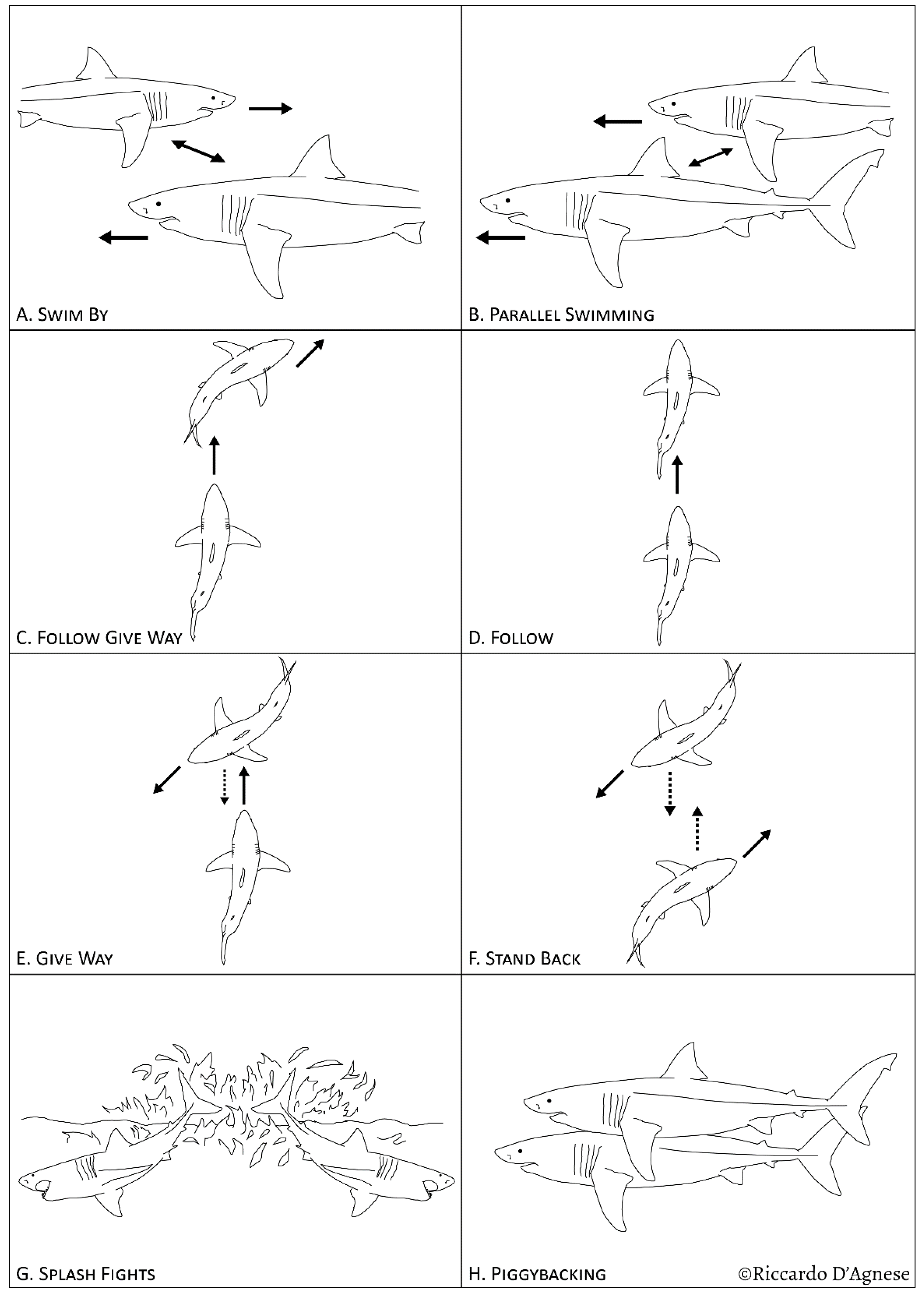
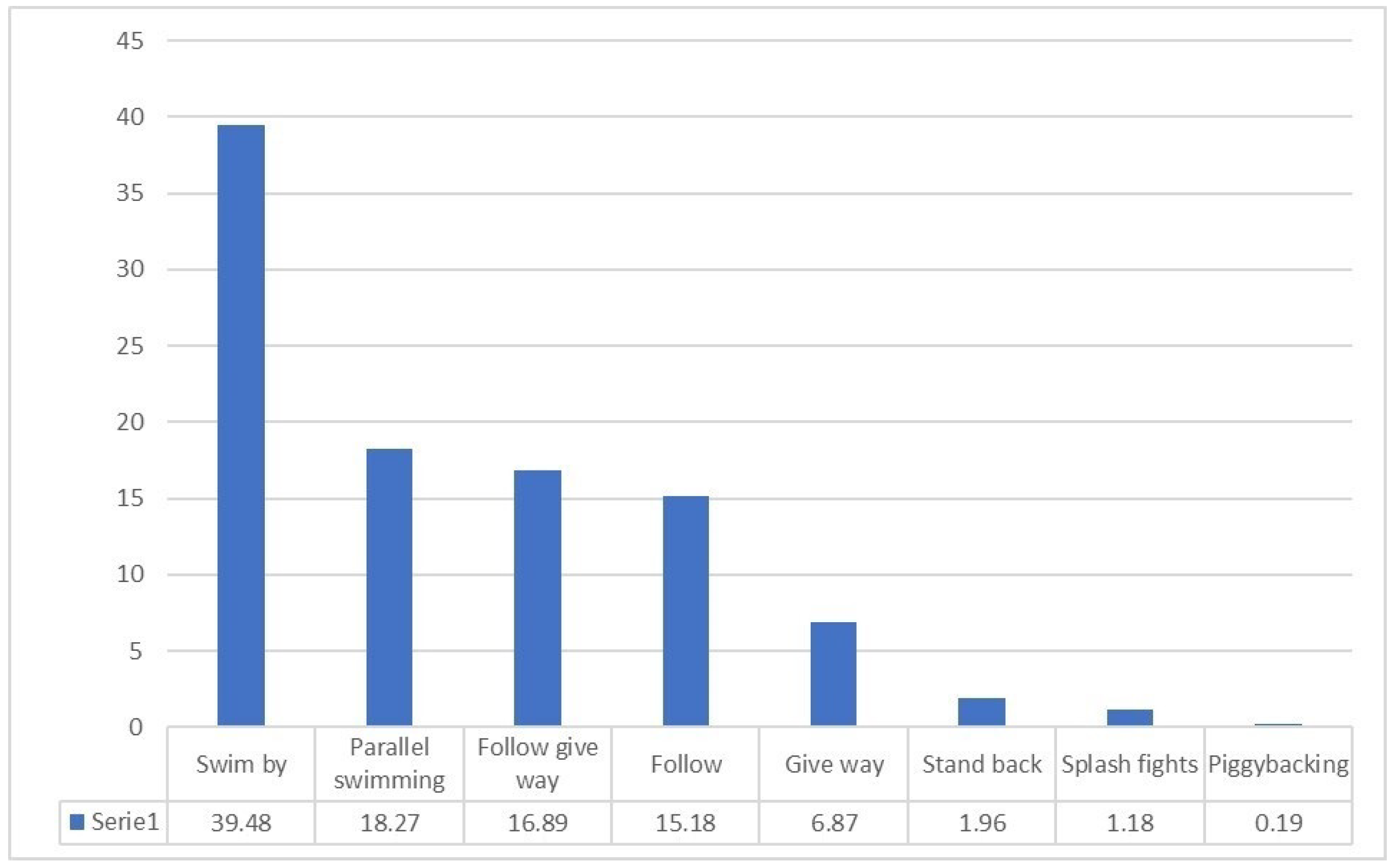
3. Results
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Baldaccini, N.E.; Mainardi, D.; Papi, F. “Introduzione Alla Etologia” Book—by Editoriale Grasso, Bologna. 1990. Available online: https://elearning.unite.it/pluginfile.php/247583/mod_resource/content/1/Testo_Didattica.pdf (accessed on 1 March 2023).
- Nowak, M.A. Five rules for the evolution of cooperation. Science 2006, 314, 1560–1563. [Google Scholar] [CrossRef] [PubMed]
- Lorenz, K. On Aggression; Psychology Press: London, UK, 1966. [Google Scholar]
- Silber, G.K. The relationship of social vocalizations to surface behaviour and aggression in the Hawaiian humpback whale (Megaptera novaeangliae). Can. J. Zool. 1986, 64, 2075–2080. [Google Scholar] [CrossRef]
- Higley, J.D.; King, S.T., Jr.; Hasert, M.F.; Champoux, M.; Suomi, S.J.; Linnoila, M. Stability of interindividual differences in serotonin function and its relationship to severe aggression and competent social behavior in rhesus macaque females. Neuropsychopharmacology 1996, 14, 67–76. [Google Scholar] [CrossRef] [PubMed]
- Connor, R.C.; Mann, J.; Tyack, P.L.; Whitehead, H. Social evolution in toothed whales. Trends Ecol. Evol. 1998, 13, 228–232. [Google Scholar] [CrossRef] [PubMed]
- Oliveira, R.F.; Simões, J.M.; Teles, M.C.; Oliveira, C.R.; Becker, J.D.; Lopes, J.S. Assessment of fight outcome is needed to activate socially driven transcriptional changes in the zebrafish brain. Proc. Natl. Acad. Sci. USA 2016, 113, E654–E661. [Google Scholar] [CrossRef] [PubMed]
- Wilson, D.S. A theory of group selection. Proc. Natl. Acad. Sci. USA 1975, 72, 143–146. [Google Scholar] [CrossRef]
- Burghardt, G.M. Defining “communication”. Commun. Chem. Signals 1970, 1, 5–18. [Google Scholar]
- Hinde, R.A. Interactions, relationships and social structure. Man 1976, 11, 1–17. [Google Scholar] [CrossRef]
- Krause, J.; Ruxton, G. Living in Groups; Oxford University Press: Oxford, UK, 2002. [Google Scholar]
- Wey, T.; Blumstein, D.T.; Shen, W.; Jordan, F. Social network analysis of animal behaviour: A promising tool for the study of sociality. Anim. Behav. 2008, 75, 33344. [Google Scholar] [CrossRef]
- Allee, W.C.; Dickinson, J.C. Dominance and subordination in the smooth dogfish, Mustelus canis (Mitchell). Physiol. Zool. 1954, 27, 356–364. [Google Scholar] [CrossRef]
- Myrberg, A.A., Jr.; Gruber, S.H. The behaviour of the bonnethead shark, Sphyrna tiburo. Copeia 1974, 1974, 358–374. [Google Scholar] [CrossRef]
- Klimley, A.P. Social Organization of Schools of the Scalloped Hammerhead Shark, Sphyrna lewini (Griffith and Smith), in the Gulf of California. Scripps Institution of Oceanography Technical Report, San Diego, CA, USA. 1983. Available online: https://escholarship.org/uc/item/2qg6s9t5 (accessed on 1 March 2023).
- Martin, R.A. Field Guide to the Great White Shark; ReefQuest Centre for Shark Research: Vancouver, BC, USA, 2003; 185p. [Google Scholar]
- Sperone, E.; Micarelli, P.; Andreotti, S.; Spinetti, S.; Andreani, A.; Serena, F.; Brunelli, E.; Tripepi, S. Social interactions among bait attracted white sharks at Dyer Island (South Africa). Mar. Bio Res. 2010, 6, 408–414. [Google Scholar] [CrossRef]
- Becerril-García, E.E.; Hoyos-Padilla, E.M.; Micarelli, P.; Galván-Magaña, F.; Sperone, E. The surface behaviour of white sharks during ecotourism: A baseline for monitoring this threatened species around Guadalupe Island, Mexico. Aquat. Conserv. Mar. Freshw. Ecosyst. 2019, 29, 773–782. [Google Scholar] [CrossRef]
- Schilds, A.; Mourier, J.; Huveneers, C.; Nazimi, L.; Fox, A.; Leu, S.T. Evidence for non-random co-occurrences in a white shark aggregation. Behav. Ecol. Sociobiol. 2019, 73, 138. [Google Scholar] [CrossRef]
- Compagno, L. Sharks of the World: An Annotated and Illustrated Catalogue of Shark Species Known to Date. Vol. 2 Bullhead, Mackerel, and Carpet Sharks (Heterodontiformes, Lamniformes and Orectolobiformes); FAO Species Catalogue for Fishery Purposes, no. 1; FAO: Rome, Italic, 2001; Volume 2. [Google Scholar]
- Klimley, A.P.; Le Bouef, B.J.; Cantara, K.M.; Richert, J.E.; Davis, S.F.; Van Sommeran, S.; Kelly, J.T. The hunting strategy of white sharks (Carcharodon carcharias) near a seal colony. Mar. Biol. 2001, 138, 617–636. [Google Scholar] [CrossRef]
- Klimley, A.P.; Anderson, S.D. Residency Patterns of White Sharks at the South Farallon Islands, California. In Great White Sharks. The Biology of Carcharodon carcharias; Klimley, A.P., Ainley, D., Eds.; Academic Press: Cambridge, MA, USA, 1996. [Google Scholar]
- Long, D.J.; Jones, R.E. White shark predation and scavenging on cetaceans in the Eastern North Pacific Ocean. In Great White Sharks. The Biology of Carcharodon carcharias; Klimley, A.P., Ainley, D., Eds.; Academic Press: Cambridge, MA, USA, 1996. [Google Scholar]
- Chase, I.D. Cooperative and noncooperative behavior in animals. Am. Nat. 1980, 115, 827–857. [Google Scholar] [CrossRef]
- De Vries, H.; Stevens, J.M.G.; Vervaecke, H. Measuring and testing the steepness of dominance hierarchies. Anim. Behav. 2006, 71, 585–592. [Google Scholar] [CrossRef]
- Findlay, R.; Gennari, E.; Cantor, M.; Tittensor, D.P. How solitary are white sharks: Social interactions or just spatial proximity? Behav. Ecol. Sociobiol. 2016, 70, 1735–1744. [Google Scholar] [CrossRef]
- Towner, A.V. Great White Sharks Carcharodon carcharias in Gansbaai, South Africa: Environmental Influences and Changes over Time, 2007–2011. Master’s Thesis, University of Cape Town (UCT), Cape Town, South Africa, 2012. [Google Scholar]
- Martin, R.A.; Hammerschlag, N.; Collier, R.S.; Fallows, C. Predatory behaviour of white sharks (Carcharodon carcharias) at Seal Island, South Africa. J. Mar. Biol. Assoc. UK 2005, 85, 1121–1135. [Google Scholar] [CrossRef]
- Hammerschlag, N.; Martin, R.A.; Fallows, C. Effects of environmental conditions on predator-prey interactions between white sharks (Carcharodon carcharias) and Cape fur seals (Arctocephalus pusillus pusillus) at Seal Island, South Africa. Environ. Biol. Fishes 2006, 76, 341–350. [Google Scholar] [CrossRef]
- Kirkman, S.P.; Oosthuizen, W.H.; Meÿer, M.A. The seal population of Seal Island, False Bay. In Finding a Balance: White Shark Conservation and Recreational Safety in the Inshore Waters of Cape Town, South Africa; Nel, D.C., Peschak, T.P., Eds.; Proceedings of a Specialist Workshop. WWF South Africa Report Series–2006/Marine/001 Annexure 1; YUMPU: Diepoldsau, Switzerland, 2006; pp. 83–94. [Google Scholar]
- Towner, A.V.; Underhill, L.G.; Jewell, O.J.; Smale, M.J. Environmental influences on the abundance and sexual composition of white sharks Carcharodon carcharias in Gansbaai, South Africa. PLoS ONE 2013, 8, e71197. [Google Scholar] [CrossRef] [PubMed]
- Micarelli, P.; Bonsignori, D.; Compagno, L.J.V.; Pacifico, A.; Romano, C.; Reinero, F.R. Analysis of sightings of white sharks in Gansbaai (South Africa). Eur. Zool. J. 2021, 88, 363–374. [Google Scholar] [CrossRef]
- Griffiths, C.L.; Robinson, T.B.; Lange, L.; Mead, A. Marine biodiversity in South Africa: An evaluation of current states of knowledge. PLoS ONE 2010, 5, e12008. [Google Scholar] [CrossRef] [PubMed]
- Towner, A.V.; Wcisel, M.A.; Reisinger, R.R.; Edwards, D.; Jewell, O.J.D. Gauging the threat: The first population estimate for white sharks in South Africa using photo identification and automated software. PLoS ONE 2013, 8, e66035. [Google Scholar] [CrossRef]
- Micarelli, P.; Chieppa, F.; Pacifico, A.; Rabboni, E.; Reinero, F.R. Passive Prey Discrimination in Surface Predatory Behaviour of Bait-Attracted White Sharks from Gansbaai, South Africa. Animals 2021, 11, 2583. [Google Scholar] [CrossRef] [PubMed]
- Ferreira, C.A.; Ferreira, T.P. Population dynamics of white sharks in South Africa. In Great White Sharks: The Biology of Carcharodon carcharias; Klimley, A.P., Ainley, D.G., Eds.; Academic Press: San Diego, CA, USA, 1996; pp. 381–391. [Google Scholar]
- Laroche, R.K.; Kock, A.A.; Dill, L.M.; Oosthuizen, W.H. Effects of provisioning ecotourism activity on the behaviour of white sharks Carcharodon carcharias. Mar. Ecol. Prog. Ser. 2007, 338, 199–209. [Google Scholar] [CrossRef]
- Compagno, L.; Dando, M.; Fowler, S. Sharks of the World; HarperCollins Publishers Ltd.: London, UK, 2005; 368p. [Google Scholar]
- Sperone, E.; Micarelli, P.; Andreotti, S.; Brandmayr, P.; Bernabò, I.; Brunelli, E.; Tripepi, S. Surface behaviour of bait-attracted white sharks at Dyer Island (South Africa). Mar. Biol. Res. 2012, 8, 982–991. [Google Scholar] [CrossRef]
- Springer, S. Social organization of shark populations. In Sharks, Skates and Rays; Gilbert, P.W., Mathewson, R.F., Rall, D.P., Eds.; Johns Hopkins Press: Baltimore, MA, USA, 1967; 644p. [Google Scholar]
- Jacoby, D.M.P.; Croft, D.P.; Sims, D.W. Social behaviour in sharks and rays: Analysis, patterns and implications for conservation. Fish Fish. 2011, 13, 399–417. [Google Scholar] [CrossRef]
- Micarelli, P.; Pieraccini, F.; Reinero, F.R.; Sperone, E. Influence of male presence on the social structure of lesser spotted dogfish (Scyliorhinus canicula) female groups. Int. J. Oceanogr. Aquac. 2020, 4. [Google Scholar] [CrossRef]
- Powter, D.M.; Gladstone, W. The reproductive biology and ecology of the Port Jackson shark Heterodontus portusjacksoni in the coastal waters of eastern Australia. J. Fish Biol. 2008, 72, 2615–2633. [Google Scholar] [CrossRef]
- Ebert, D.A. Observations on the predatory behaviour of the sevengill shark Notorynchus cepedianus. S. Afr. J. Mar. Sci. 1991, 11, 455–465. [Google Scholar] [CrossRef]
- Standora, E.A.; Nelson, D.R. A telemetric study of the behavior of free-swimming angel sharks Squatina californica. Bull. South. Calif. Acad. Sci. 1977, 76, 193–201. [Google Scholar]
- Klimley, A.P.; Nelson, D.R. Schooling of the scalloped hammerhead sharks, Sphyrna lewini, in the Gulf of California. Fish. Bull. 1981, 79, 356–360. [Google Scholar]
- McKibben, J.N.; Nelson, D.R. Patterns of movement and grouping of gray reef sharks, Carcharhinus amblyrhynchos, at Enewetak, Marshall Islands. Bull. Mar. Sci. 1986, 38, 89–110. [Google Scholar]
- Meekan, M.G.; Bradshaw, C.J.A.; Press, M.; Mc Lean, C.; Richards, A.; Quasnichka, S.; Taylor, J. Population size and structure of whale sharks Rhincodon typus at Ningaloo Reef, Western Australia. Mar. Ecol. Prog. Ser. 2006, 319, 275–285. [Google Scholar] [CrossRef]
- Domeier, M.L.; Nasby-Lucas, N. Annual re-sightings of photographically identified white sharks (Carcharodon carcharias) at an eastern Pacific aggregation site (Guadalupe Island, Mexico). Mar. Biol. 2007, 150, 977–984. [Google Scholar] [CrossRef]
- Heupel, M.R.; Simpfendorfer, C.A. Quantitative analysis of aggregation behavior in juvenile blacktip sharks. Mar. Biol. 2005, 147, 1239–1249. [Google Scholar] [CrossRef]
- Stevens, J.D. Biological observations on sharks caught by sport fisherman of New South Wales. Mar. Freshw. Res. 1984, 35, 573–590. [Google Scholar] [CrossRef]
- Economakis, A.E.; Lobel, P.S. Aggregation behavior of the grey reef shark, Carcharhinus amblyrhynchos, at Johnson Atoll, Central Pacific Ocean. Environ. Biol. Fishes 1998, 51, 129–139. [Google Scholar] [CrossRef]
- Speed, C.W.; Meekan, M.G.; Field, I.C.; Mcmahon, C.R.; Stevens, J.D.; Mcgregor, F.; Huveneers, C.; Berger, Y.; Bradshaw, C.J.A. Spatial and temporal movement patterns of a multi-species coastal reef shark aggregation. Mar. Ecol. Prog. Ser. 2011, 429, 261–275. [Google Scholar] [CrossRef]
- Wearmouth, V.J.; Sims, D.W. Sexual segregation in marine fish, reptiles, birds and mammals: Behaviour patterns, mechanisms and conservation implications. Adv. Mar. Biol. 2008, 54, 107–170. [Google Scholar] [PubMed]
- Simpfendorfer, C.A. The Biology of Sharks of the Family Carcharhinidae from the Nearshore Waters of Cleveland Bay, with particular reference to Rhizoprionodon taylori. Doctoral Dissertation, James Cook University, Brisbane, Australia, 1993. [Google Scholar]
- Hight, B.H.; Lowe, C.G. Elevated body temperatures of adult female leopard sharks, Triakis semifasciata, while aggregating in shallow nearshore embayments: Evidence for behavioral thermoregulation? J. Exp. Mar. Biol. Ecol. 2007, 352, 114–128. [Google Scholar] [CrossRef]
- Jacoby, D.M.P.; Busawon, D.S.; Sims, D.W. Sex and social networking: The influence of male presence on social structure of female shark groups. Behav. Ecol. 2010, 21, 808–818. [Google Scholar] [CrossRef]
- Elisio, M.; Colonello, J.H.; Cortés, F.; Jaureguizar, A.J.; Somoza, G.M.; Macchi, G.J. Aggregations and reproductive events of the narrownose smooth-hound shark (Mustelus schmitti) in relation to temperature and depth in coastal waters of the south-western Atlantic Ocean (38–42 S). Mar. Freshw. Res. 2016, 68, 732–742. [Google Scholar] [CrossRef]
- Perry, C.T.; Clingham, E.; Webb, D.H.; de la Parra, R.; Pierce, S.J.; Beard, A.; Henry, L.; Taylor, B.; Andrews, K.; Hobbs, R.; et al. St. Helena: An important reproductive habitat for whale sharks (Rhincodon typus) in the central south Atlantic. Front. Mar. Sci. 2020, 7, 576343. [Google Scholar] [CrossRef]
- Kock, A.; O’Riain, M.J.; Mauff, K.; Meÿer, M.; Kotze, D.; Griffiths, C. Residency, habitat use and sexual segregation of white sharks, Carcharodon carcharias in False Bay, South Africa. PLoS ONE 2013, 8, e55048. [Google Scholar] [CrossRef]
- Becerril-García, E.E.; Hoyos-Padilla, E.M.; Micarelli, P.; Galván-Magaña, F.; Sperone, E. Behavioural responses of white sharks to specific baits during cage diving ecotourism. Sci. Rep. 2020, 10, 11152. [Google Scholar] [CrossRef]
- Duffy, C.; Francis, M.P.; Manning, M.J.; Bonfil, R. Chapter 21: Regional population connectivity, oceanic habitat, and return migration revealed by satellite tagging of white sharks, Carcharodon carcharias, at New Zealand aggregation sites. In Global Perspectives on the Biology and Life History of the White Shark; Domeier, M.L., Ed.; CRC Press: Boca Raton, FL, USA, 2012; pp. 301–318. [Google Scholar]
- Skomal, G.B.; Chisholm, J.; Correia, S.J. Implications of increasing pinniped populations on the diet and abundance of white sharks off the coast of Massachusetts. In Global Perspectives on the Biology and Life History of the White Shark; Domeier, M.L., Ed.; CRC Press: Boca Raton, FL, USA, 2012; pp. 405–418. [Google Scholar]
- Robbins, R.L. Environmental variables affecting the sexual segregation of great white sharks Carcharodon carcharias at the Neptune Islands South Australia. J. Fish Biol. 2007, 70, 1350–1364. [Google Scholar] [CrossRef]
- Semmens, J.; Payne, N.; Huveneers, C.; Sims, D.W.; Bruce, B. Feeding requirements of white sharks may be higher than originally thought. Sci. Rep. 2013, 3, 1471. [Google Scholar] [CrossRef]
- Goldman, K.J.; Anderson, S.D. Space utilization and swimming depth of white sharks, Carcharodon carcharias, at the South Farallon Islands, central California. Env. Biol. Fish. 1999, 56, 351–364. [Google Scholar] [CrossRef]
- Weng, K.C.; Boustany, A.M.; Pyle, P.; Anderson, S.D.; Brown, A.; Block, B.A. Migration and habitat of white sharks (Carcharodon carcharias) in the eastern Pacific Ocean. Mar Biol 2007, 152, 877–894. [Google Scholar] [CrossRef]
- Dunbar, R.; Shultz, S. Evolution in the social brain. Science 2007, 317, 1344–1347. [Google Scholar] [CrossRef]
- Clark, E. Instrumental conditioning of lemon sharks. Science 1959, 130, 217–218. [Google Scholar] [CrossRef]
- Guttridge, T. The Social Organization and Behaviour of the Juvenile Lemon Shark Negaprion breviostris. Ph.D. Thesis, University of Leeds, Institute of Integrative and Comparative, Leeds, UK, 2009. [Google Scholar]
- Northcutt, R.G. Elasmobranch central nervous system organization and its possible evolutionary significance. Am. Zool. 1977, 17, 411–429. [Google Scholar] [CrossRef]
- Yopak, K.E.; Lisney, T.J.; Collin, S.P.; Montgomery, J.C. Variation in brain organization and cerebellar foliation in chondrichthyans: Sharks and holocephalans. Brain Behav. Evol. 2007, 69, 280–300. [Google Scholar] [CrossRef]
- Striedter, G.F.; Northcutt, R.G. Head size constrains forebrain development and evolution in ray-finned fishes. Evol. Dev. 2006, 8, 215–222. [Google Scholar] [CrossRef] [PubMed]
- Holland, K.N.; Meyer, C.G.; Dagorn, L.C. Inter-animal telemetry: Results from first deployment of acoustic ‘business card’ tags. Endanger. Species Res. 2009, 10, 287–293. [Google Scholar] [CrossRef]
- Guttridge, T.L.; Gruber, S.H.; Krause, J.; Sims, D.W. Novel acoustic technology for studying free-ranging shark social behaviour by recording individuals’ interactions. PLoS ONE 2010, 5, e9324. [Google Scholar] [CrossRef] [PubMed]
- Krause, J.; James, R.; Croft, D.P. Personality in the context of social networks. Philos. Trans. R. Soc. B Biol. Sci. 2010, 365, 4099–4106. [Google Scholar] [CrossRef]
- Klimley, A.P.; Nelson, D.R. Diel movement pattern of the scalloped hammerhead shark Sphyrna lewini in relation to el Bajo Espiritu Santo: A refuging central position social system. Behav. Ecol. Sociobiol. 1984, 15, 45–54. [Google Scholar] [CrossRef]
- Sims, D.W.; Southall, E.J.; Quayle, V.A.; Fox, M. Annual social behaviour of basking sharks associated with coastal front areas. Proc. R. Soc. Lond. Ser. B Biol. Sci. 2000, 267, 1897–1904. [Google Scholar] [CrossRef] [PubMed]
- Guttridge, T.L.; Gruber, S.H.; Gledhill, K.S.; Croft, D.P.; Sims, D.W.; Krause, J. Social preferences of juvenile lemon 543 sharks, Negaprion brevirostris. Anim. Behav. 2009, 78, 543–548. [Google Scholar] [CrossRef]
- Jacoby, D.M.P.; Fear, L.N.; Sims, D.W.; Croft, D.P. Shark personalities? Repeatability of social network traits in a widely distributed predatory fish. Behav. Ecol. Sociobiol. 2014, 68, 1995–2003. [Google Scholar] [CrossRef]
- Anderson, J.M.; Clevenstine, A.J.; Stirling, B.S.; Burns, E.S.; Meese, E.N.; White, C.F.; Logan, R.K.; O’Sullivan, J.; Rex, P.T.; May, J., III; et al. Non-random Co-occurrence of Juvenile White Sharks (Carcharodon carcharias) at Seasonal Aggregation Sites in Southern California. Front. Mar. Sci. 2021, 8, 688505. [Google Scholar] [CrossRef]
- Papastamatiou, Y.P.; Mourier, J.; TinHan, T.; Luongo, S.; Hosoki, S.; Santana-Morales, O.; Hoyos-Padilla, M. Social dynamics and individual hunting tactics of white sharks revealed by biologging. Biol. Lett. 2022, 18, 20210599. [Google Scholar] [CrossRef]
- Klimley, A.P.; Le Boeuf, B.J.; Cantara, K.M.; Richert, J.E. Automated tracking of white shark Carcharodon carcharias by radio-acoustic positioning system. Mar. Biol. 2000, 4, 182–193. [Google Scholar]
- Hsu, Y.; Earley, R.L.; Wolf, L.L. Modulating Aggression through Experience. Fish Cognition and Behaviour; Blackwell: Oxford, UK, 2006; pp. 96–118. [Google Scholar]
- Fallows, C.; Gallagher, A.J.; Hammerschlag, N. White sharks (Carcharodon carcharias) scavenging on whales and its potential role in further shaping the ecology of an apex predator. PLoS ONE 2013, 8, e60797. [Google Scholar] [CrossRef]
- Bromilov, M. Feeding Behaviour of White Sharks Carcharodon carcharias around a Cage Diving Vessel and the Implications for Conservation. Ph.D. Thesis, University of Michigan, Department of Ecology & Evolutionary Biology, Ann Arbor, MI, USA, 2014; pp. 6–8. [Google Scholar]
- Strong, W.R.; Murphy, R.C.; Bruce, D.B.; Nelson, D.R. Movements and associated observations of bait attracted white sharks Carcharodon carcharias: A preliminary report. Mar. Freshw. Res. 1992, 43, 13–20. [Google Scholar] [CrossRef]
- Strong, W.R. Repetitive aerial gaping: A thwart-induced behaviour in white sharks. In Great White Sharks; Academic Press: Cambridge, MA, USA, 1996; pp. 207–215. [Google Scholar]
- Lisney, T.J.; Bennet, M.B.; Collin, S.P. Volumetric analysis of sensory brain areas indicates ontogenetic shifts in the relative importance of sensory systems in elasmobranchs. Raffles Bull. Zool. 2007, 55, 7–15. [Google Scholar]
- Reinero, F.R.; Sperone, E.; Giglio, G.; Pacifico, A.; Mahrer, M.; Micarelli, P. Influence of Environmental Factors on Prey Discrimination of Bait-Attracted White Sharks from Gansbaai, South Africa. Animals 2022, 12, 3276. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Sex/Maturity (X) | Patterns/Behavior (Y) |
|---|---|
| Sex | p-Value 0.00 |
| Maturity | p-Value 0.00 |
| Sex/Maturity | p-Value 0.00 |
| Chi–Square on social behavior (p-Value) | |
| Total specimen | 0.00 *** |
| Males | 0.00 *** |
| Females | 0.00 *** |
| Adults | 0.00 *** |
| Immatures | 0.00 *** |
| Test Statistic | 175,98 |
| Chi-squared distribution (critical value) | 3,840 |
| df | 1 |
| p-Value (one-tailed) | 0,027 |
| significance level | 0,050 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Micarelli, P.; Reinero, F.R.; D’Agnese, R.; Pacifico, A.; Giglio, G.; Sperone, E. Evidence of Non-Random Social Interactions between Pairs of Bait-Attracted White Sharks in Gansbaai (South Africa). Diversity 2023, 15, 433. https://doi.org/10.3390/d15030433
Micarelli P, Reinero FR, D’Agnese R, Pacifico A, Giglio G, Sperone E. Evidence of Non-Random Social Interactions between Pairs of Bait-Attracted White Sharks in Gansbaai (South Africa). Diversity. 2023; 15(3):433. https://doi.org/10.3390/d15030433
Chicago/Turabian StyleMicarelli, Primo, Francesca Romana Reinero, Riccardo D’Agnese, Antonio Pacifico, Gianni Giglio, and Emilio Sperone. 2023. "Evidence of Non-Random Social Interactions between Pairs of Bait-Attracted White Sharks in Gansbaai (South Africa)" Diversity 15, no. 3: 433. https://doi.org/10.3390/d15030433