
The Early History of Giant Cockroaches: Gyroblattids and Necymylacrids (Blattodea) of the Late Carboniferous

Abstract
:1. Introduction
2. Material and Methods
2.1. Repositories and Institutional Abbreviations
2.2. Abbreviations
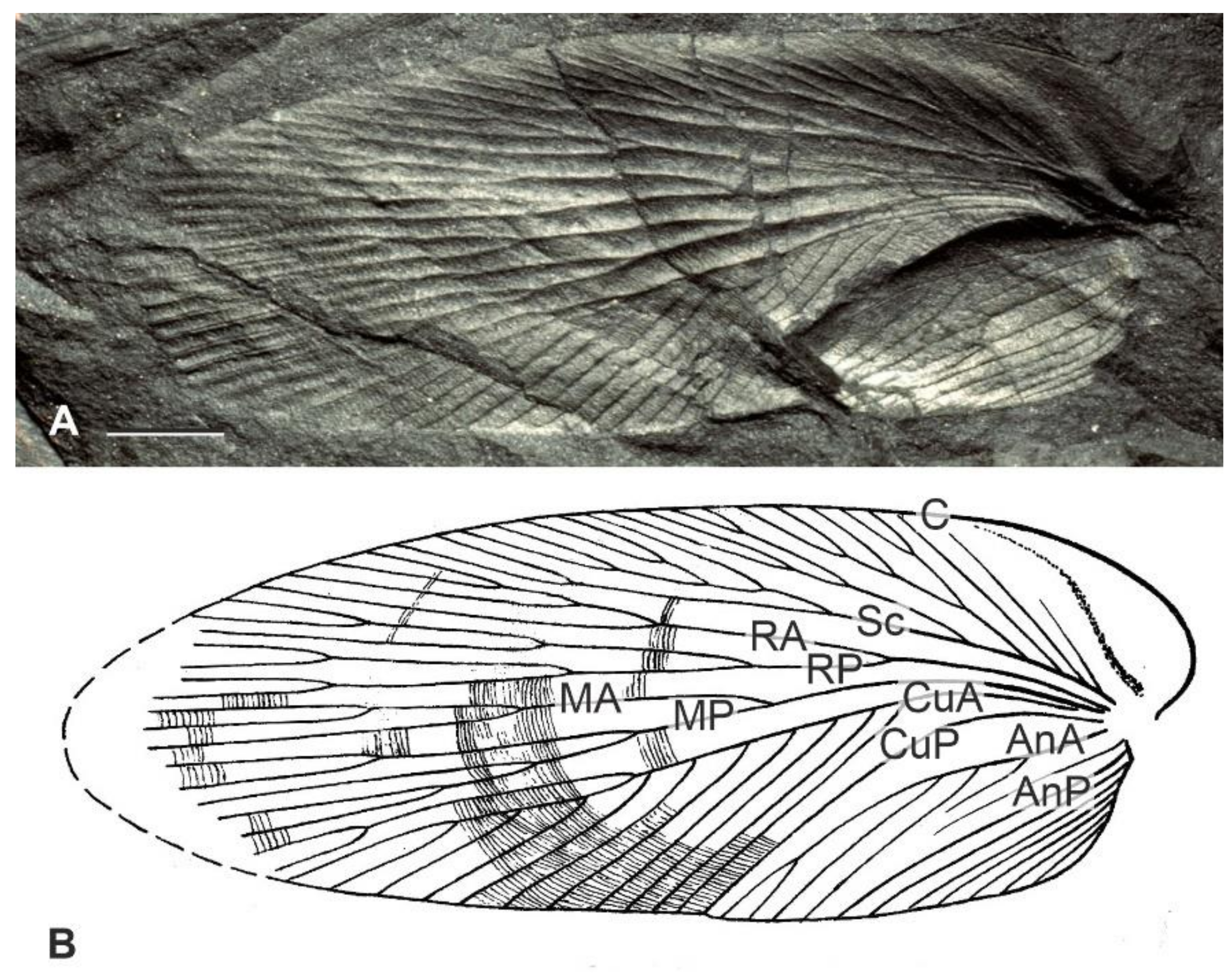
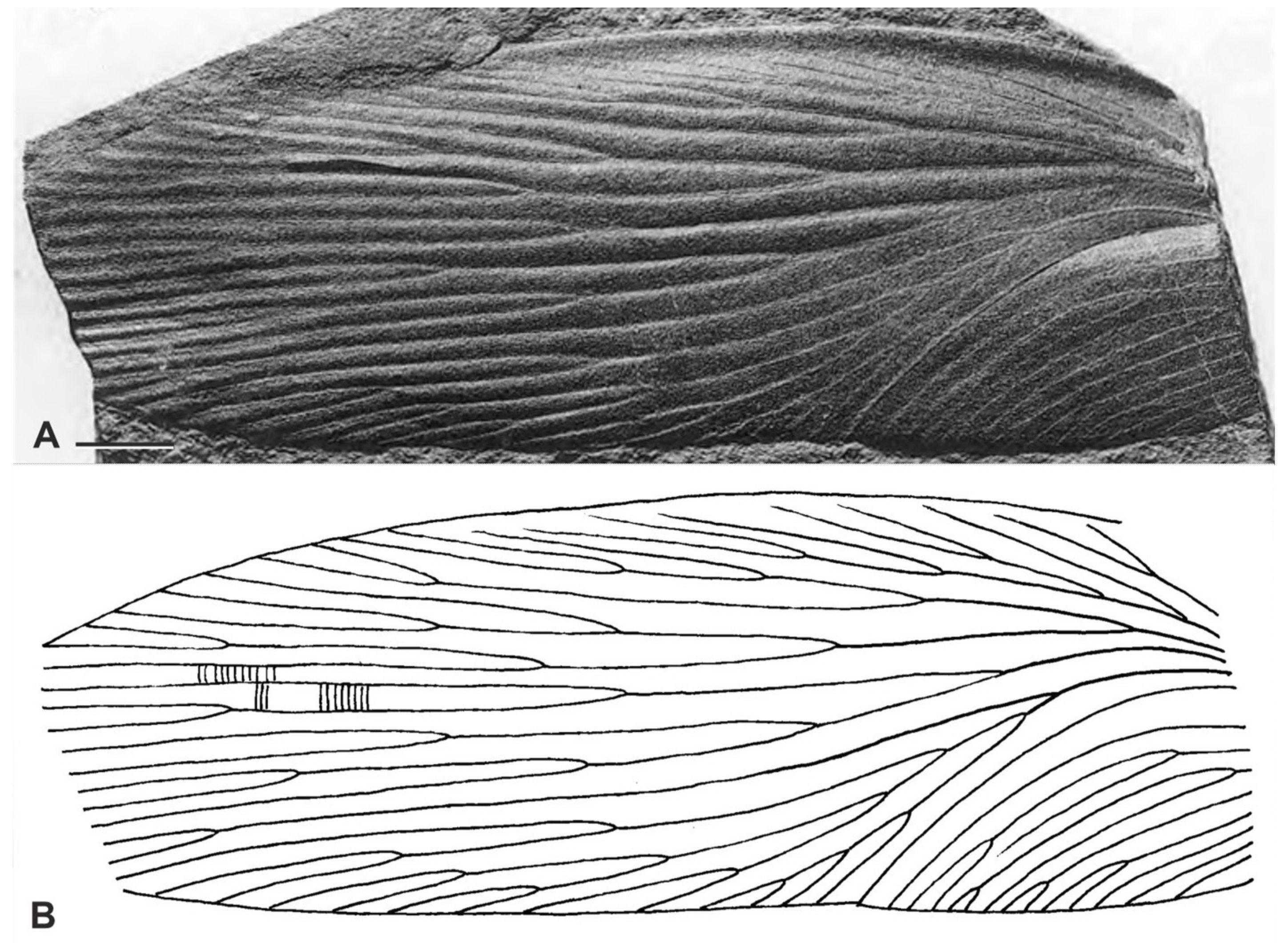
3. Systematic Paleontology



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
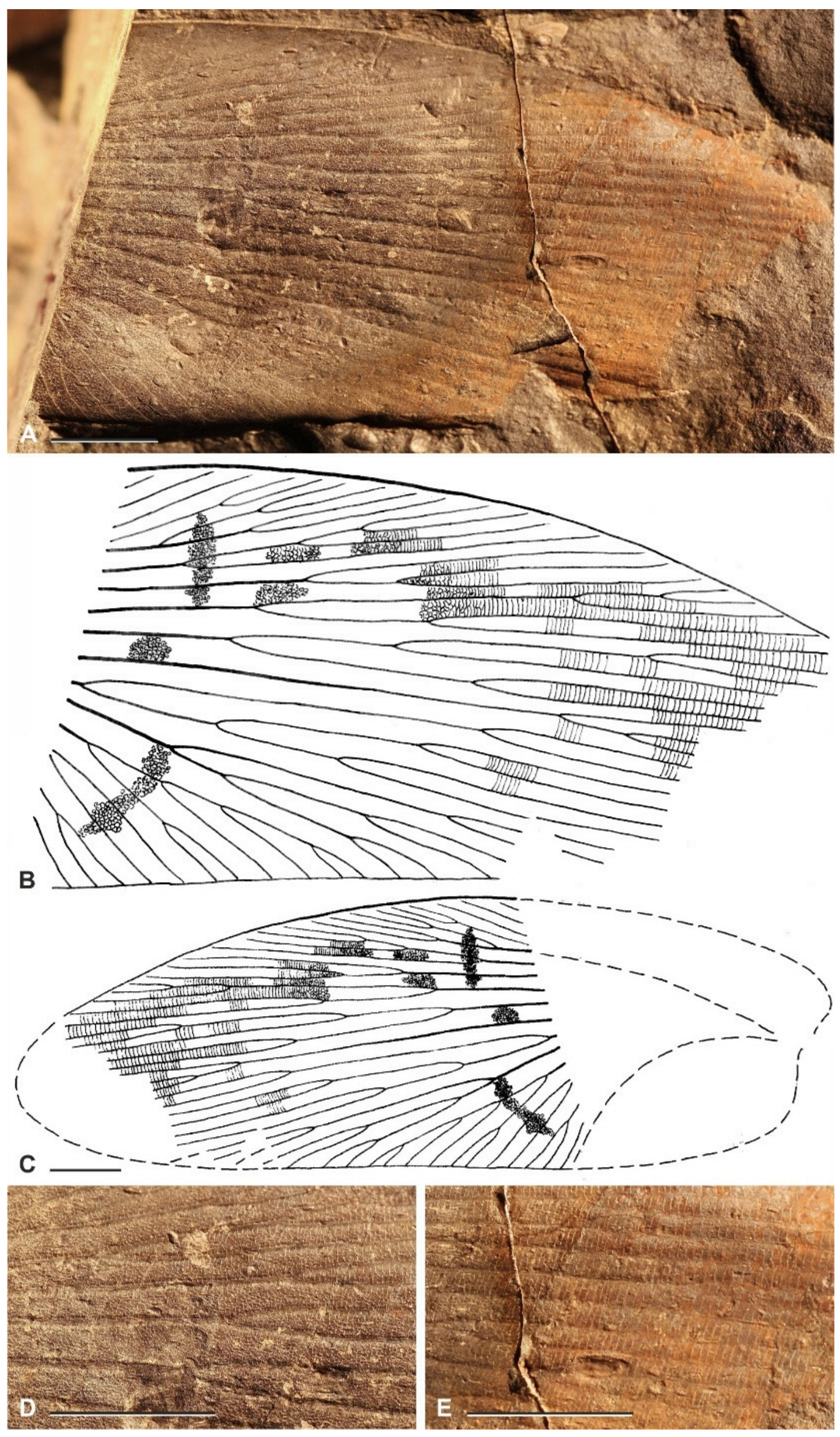{kind=link}
{kind=link}
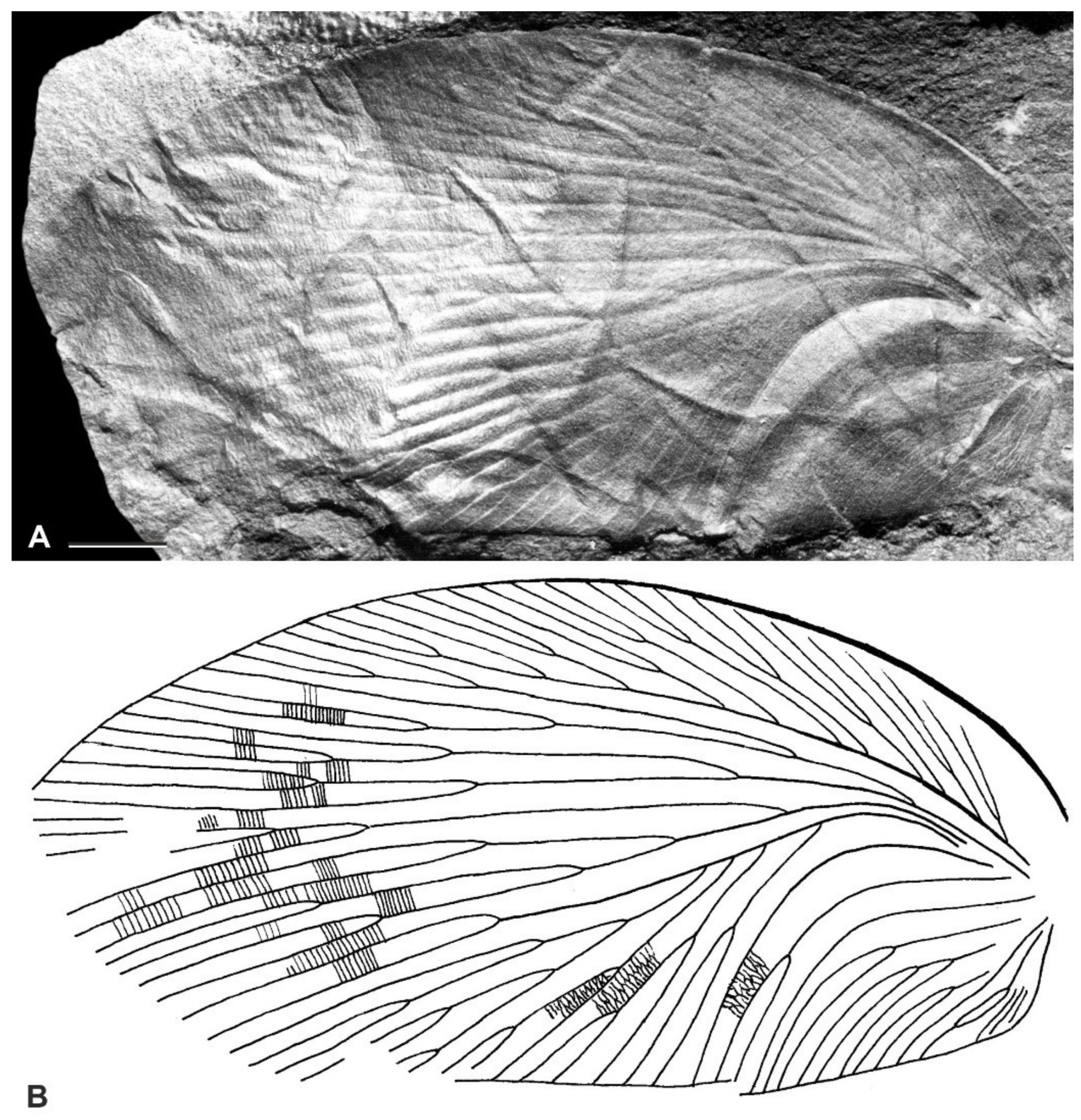{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
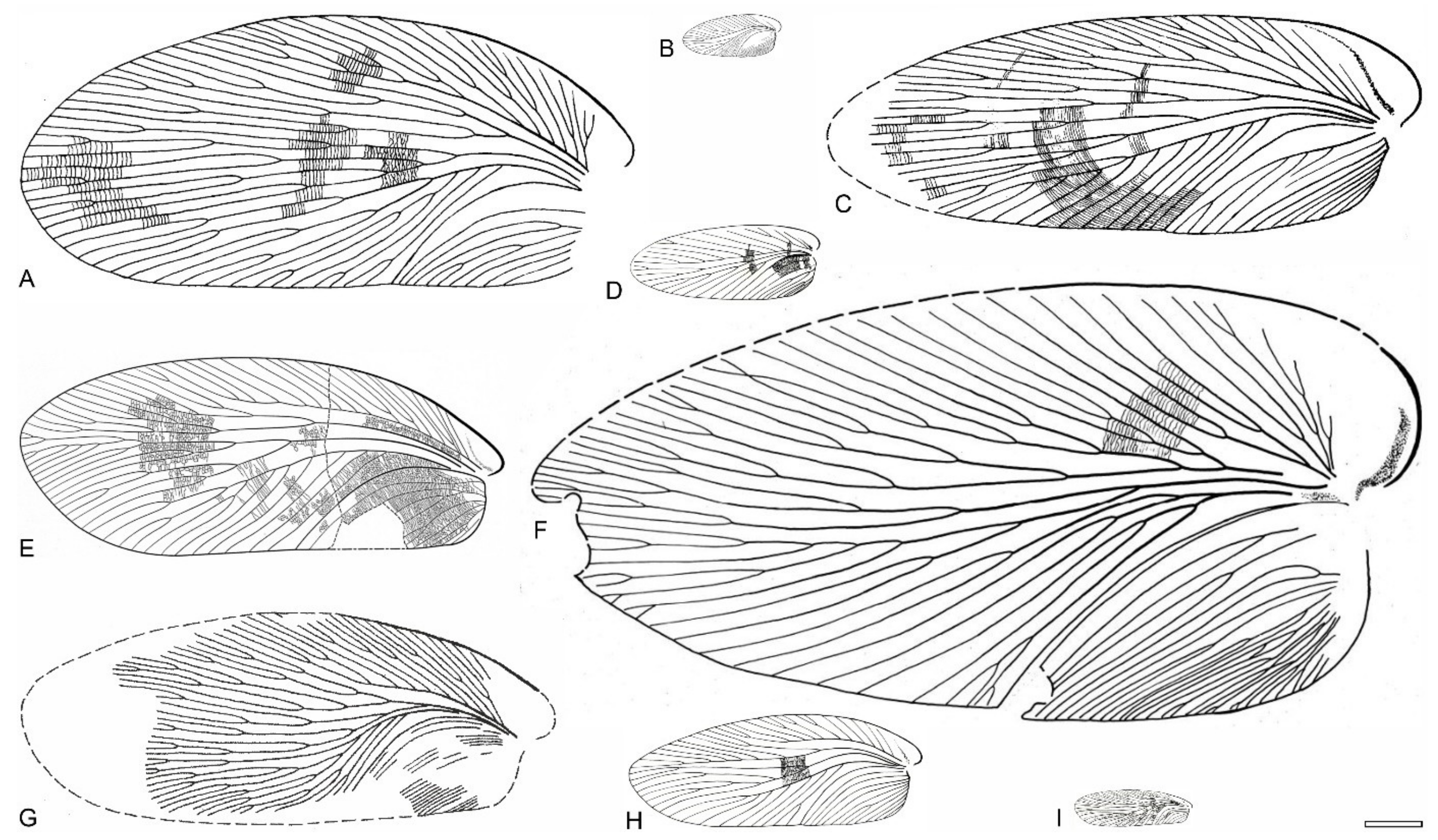{kind=link}
{kind=link}
| Specimen | Locality and Lithostratigraphy | Chronostratigraphy |
|---|---|---|
| Progonoblattina clarkii | ||
| ‘Etoblattina’ clarkii Scudder, 1893 | Pawtucket, Rhode Island, Narragansett Basin, USA; lower Rhode Island Fm. | WC to ?WD, early (to ?late) Moscovian |
| ‘Etoblattina illustris’ Scudder, 1893 | Pawtucket, Rhode Island, Narragansett Basin, USA; lower Rhode Island Fm. | WC to ?WD, early (to ?late) Moscovian |
| ‘Gerablattina scapularis’ Scudder, 1893 | Pawtucket, Rhode Island, Narragansett Basin, USA; lower Rhode Island Fm. | WC to ?WD, early (to ?late) Moscovian |
| ‘Phoberoblatta grandis’ Handlirsch, 1906a | Fishing Creek Gap, Sharp Mountain, Pennsylvania, Appalachian Basin, USA; Pottsville Fm., Sharp Mountain Mb. | WD, late Moscovian |
| ‘Cebenniblatta marcellini’ Laurentiaux, 1950 | Molières-sur-Cèze, Gard Basin, France | WD, late Moscovian |
| Progonoblattina helvetica | ||
| ‘Blatta’ helvetica Heer, 1864 | Lower quarry of Erbignon, Valais, Salvan-Dorenaz Basin, Switzerland; Zone Houillère | StB, Kasimovian |
| ‘Necymylacris boulei’ Agnus, 1903 | Commentry, Commentry Basin, France; Grande Couche | StB/C, Gzhelian |
| ‘Etoblattina steinmannii’ Sterzel, 1904 | Hinterohlsbach near Oppenau, Oppenau Basin, Germany; Oppenau Fm. | StA/B, Kasimovian |
| ‘Etoblattina fontanensis’ Meunier, 1906 | Fontanes, Gard Basin, France; 4th coal seam | StA, Kasimovian |
| ‘Progonoblattina heeri’ Handlirsch, 1906b | Lower quarry of Erbignon, Valais, Salvan-Dorénaz Basin, Switzerland; Zone Houillère | StB, Kasimovian |
| ‘Necymylacris sp.’ Meunier, 1921 | Commentry, Commentry Basin, France; Grande Couche | StB/C, Gzhelian |
| ‘Bertrandiblatta inexpectata’ Laurentiaux, 1950 | Malpertus pass, Pontil road, Gard Basin, France; Le Pin coal seam | StA, Kasimovian |
| ‘Platyblattina maxima’ Laurentiaux, 1950 | Fontanes, Gard Basin, France; 4th coal seam | StA, Kasimovian |
| ‘Platyblattina cf. maxima’ Laurentiaux, 1950 | La Grand’Combe, Gard Basin, France; Le Pin coal seam | StA, Kasimovian |
| ‘Platyblattina ampla’ Laurentiaux, 1950 | Fontanes, Gard Basin, France; 4th coal seam | StA, Kasimovian |
| ‘Livetiblatta incerta’ Laurentiaux, 1950 | La Grand’Combe, Gard Basin, France; Le Pin coal seam | StA, Kasimovian |
| ‘Hispanoblatta cantabrica’ sp. nov. Laurentiaux, 1958, nomen nudum | San Felices de Castilleria, Central Asturian Coalfield, Province Palencia, Spain; San Felices de Castilleria coal seam | StA, Kasimovian |
| ‘Sterzelia lamurensis’ Haudour et al., 1960 | La Mure (Isère) Basin, French Alps; “groupe des Trois-Bancs, Cinquième pendage, niveau 12” | early Stephanian, Kasimovian |
| ‘aff. Necymylacris’ (Eumorphoblatta) boulei Haudour et al., 1960 | La Mure (Isère) Basin, French Alps; “groupe des Trois-Bancs, Cinquième pendage, niveau 12” | early Stephanian, Kasimovian |
| ‘Guichenbachia gigantea’ Guthörl, 1963 | Coal mine Guichenbach near Heusweiler-Saar, Saar-Nahe Basin, Germany; roof of seam Wahlschied, Dilsburg Fm., Ottweiler Subgroup | StA, Kasimovian |
| ‘Dysmenes idoneus’ Laurentiaux, 1966 | Saint-Jaques, well B, Bosmoreau-les-Mines Basin (Creuse), France; level “chez Lamé” | early Stephanian, Kasimovian |
| ‘Anthracoblattina meganalis’ Brauckmann and Hahn, 1983 | Brandeck-Lindle, NE of Hinterohlsbach near Oppenau, Oppenau Basin, Germany; Oppenau Fm. | StA/B, Kasimovian |
| ‘Necymylacris sp.’ Schneider et al., 2005 | Left bank of the Zwickauer Mulde river at Cainsdorf Bridge, Zwickau, Zwickau Subbasin, central Germany; roof of Tiefes Planitzer coal seam, Zwickau Fm. | WD, late Moscovian |
| ‘Anthracoblattina ensifera-gigantea group’, specimen CDUE TaI-13d-25, Belahmira et al., 2019 | Tanamert village at Qued Issene Canyon, locality Tanamert I (TAI), Western High Atlas Mountains, Souss Basin, Ida Ou Ziki Subbasin, Morocco; Oued Issene Fm. | StA/B, Kasimovian |
| Progonoblattina sp. indet. (nomina dubia) | ||
| ‘Mylacris packardi’ Scudder, 1893 | Bristol, Rhode Island, Narragansett Basin, USA; Rhode Island Fm. | WC to WD, early to late Moscovian |
| ‘Archoblattina beecheri’ (Sellards, 1903) | Mazon Creek, Illinois, Illinois Basin, USA; Francis Creek shale, Desmoinesian | WD, late Moscovian |







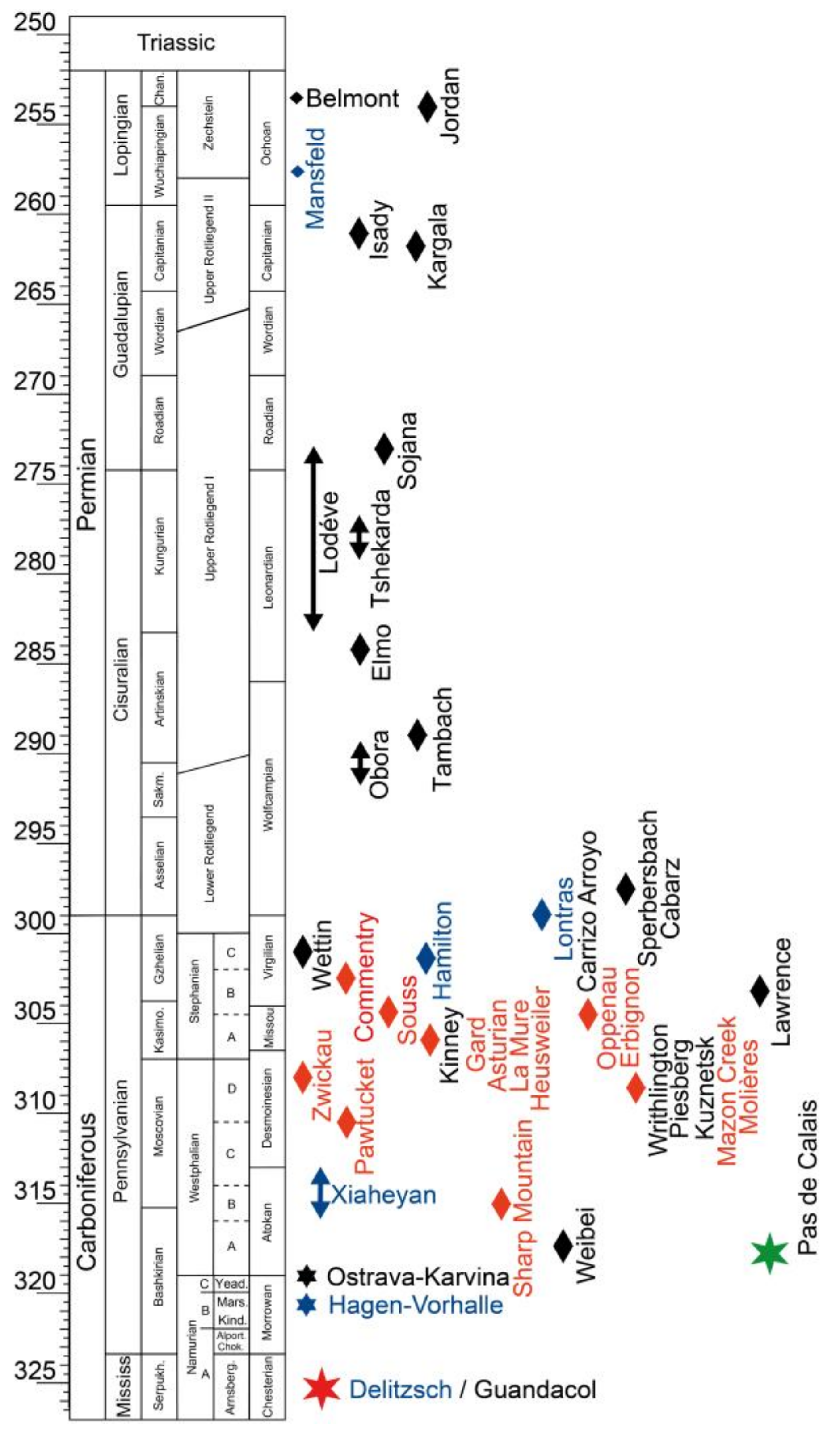
4. Implications on Stratigraphy, Paleobiogeography and Paleoecology
4.1. Stratigraphy
4.2. Paleobiogeography and Paleoecology

5. Discussion
Implications on the Large-Winged Late Paleozoic Blattoids

6. Conclusions
- Large-winged blattoid insects appear suddenly in the Euramerican fossil record in the Middle Pennsylvanian (Moscovian, middle to late Atokan) without any so far known forerunners. The families Necymylacridae Durden, 1969, and Gyroblattidae Durden, 1969, comprise the largest pre-Cenozoic blattoids, with forewing lengths of up to 7 cm, besides scarce Late Pennsylvanian/early Permian species of the Mylacridae genus Opsiomylacris, which reaches forewing lengths of up to 7.5 cm.
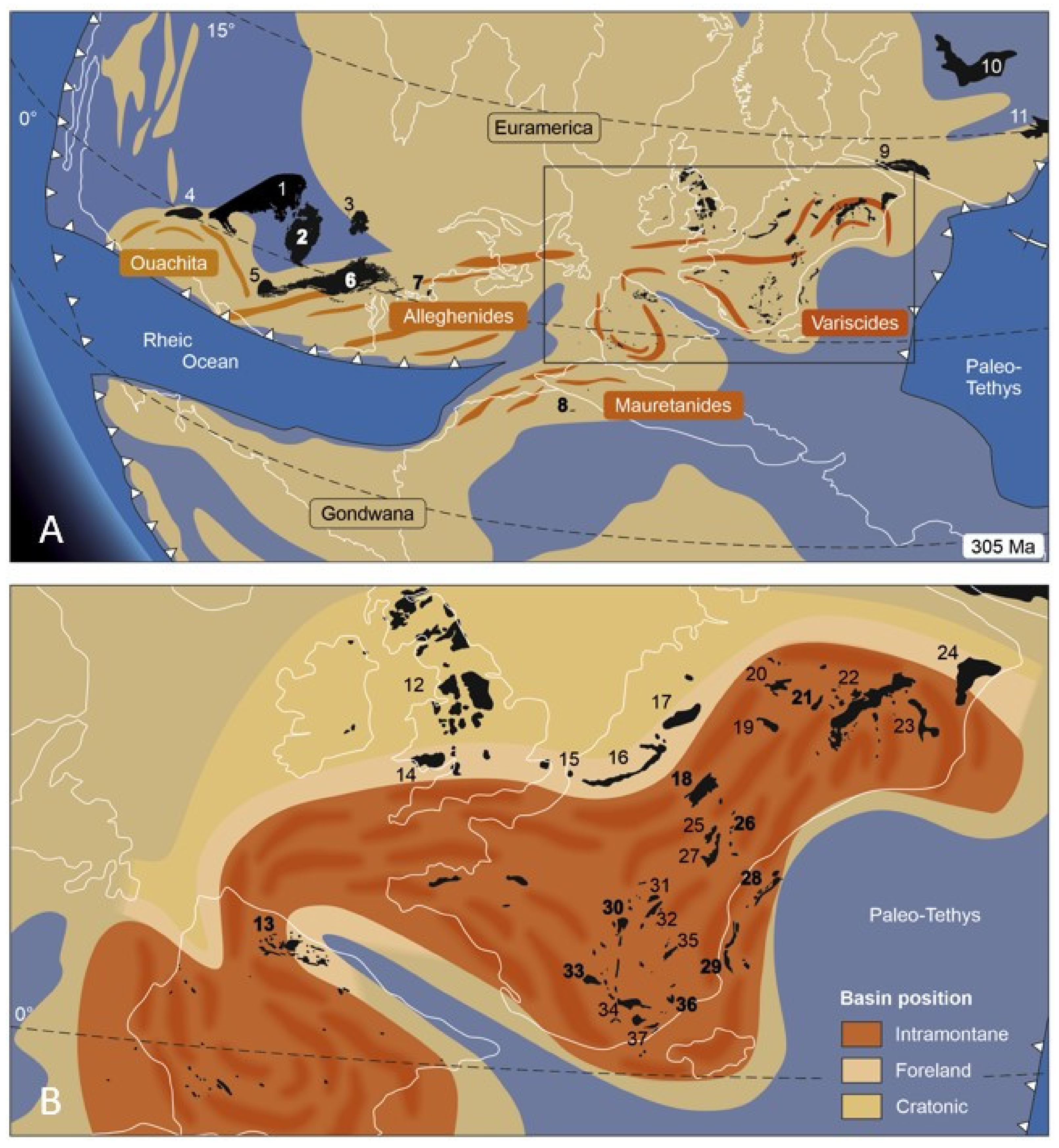
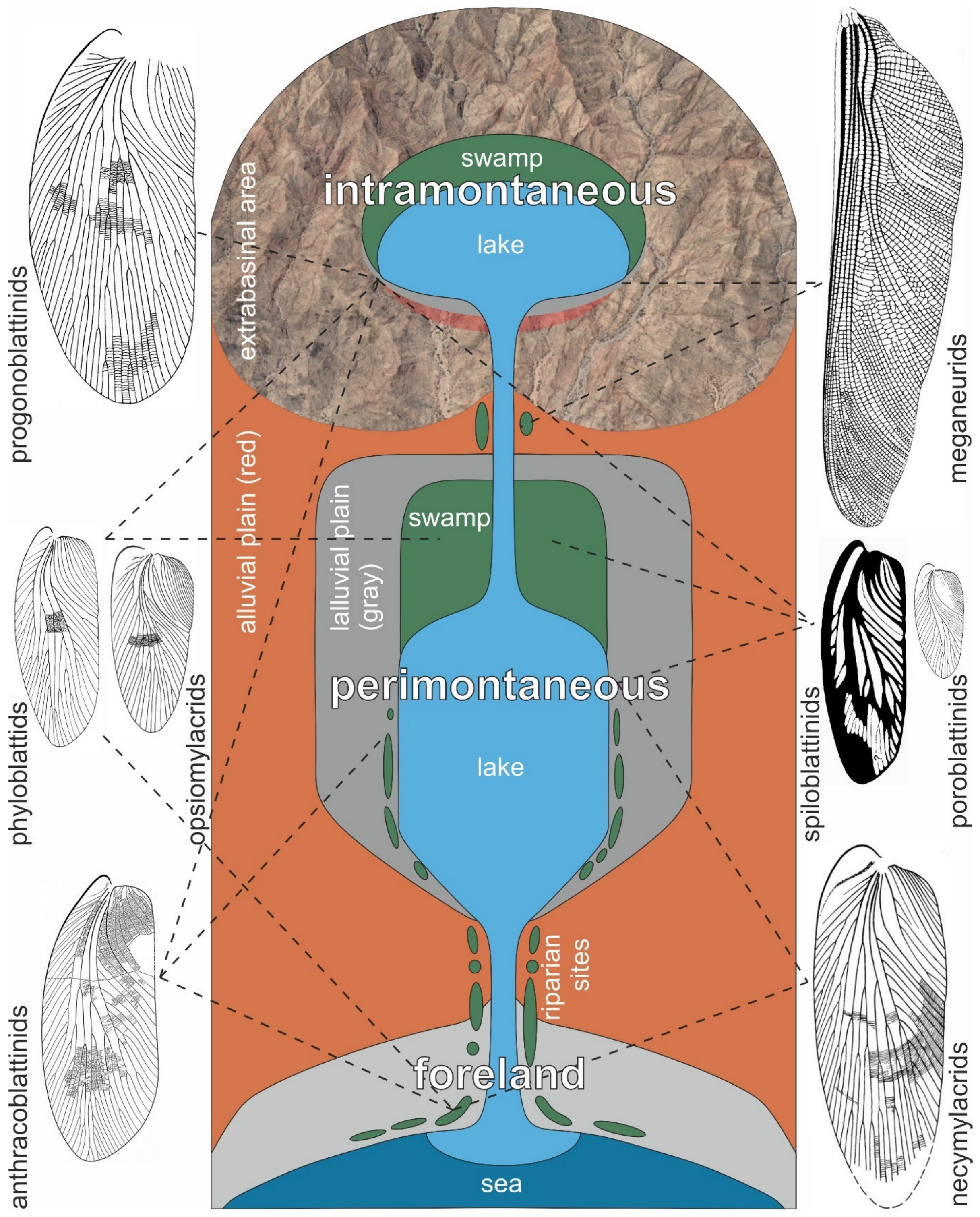
- In contrast to necymylacrids, the gyroblattids display a particular habitat preference, as concluded from their occurrences in North America, North Africa, and Europe. They are found in coal-bearing sequences, particularly in inter-seam fine clastics, roof shales, and lacustrine laminites. As a rule, they are associated with meso- to xerophilous gymnospermous plants, such as Dicranophyllum, Lesleya, Taeniopteris, and Pterophyllum, and several walchian conifers, peltasperms, or cordaitaleans as well. These relationships with floral elements are reflected in the vast North American basins, e.g., the Narragansett, Illinois, and Appalachian basins, where gyroblattids first occurred, but also in the tiny basins of the European and North African Variscids, where they persisted until the Stephanian B/C (middle Gzhelian).
- We hypothesize that gyroblattids lived mainly outside the basin centers that usually maintained ever-wet conditions. Instead, they may have existed in well-drained hinterland areas from where they immigrated with increasing seasonality during the Late Pennsylvanian. Gyroblattids likely followed meso- to xerophilous vegetational elements and preferably colonized relatively small basins, which offered more diverse edaphic conditions caused by the spatial tightness of erosional and depositional areas on a small scale.
- The large-winged gyroblattids and necymylacrids disappeared in the fossil record already in the latest Pennsylvanian (middle Gzhelian, middle Virgilian). At that time, more drought-resistant floras started to spread and increasingly dominated biomes with stronger seasonality and longer periods of dryness. Other blattoid insects adapted to these climate conditions with increasing elytrification, especially species of the genus Opsiomylacris in the early Permian. The increasing size reduction in phyloblattids is possibly linked to these environmental processes.
- To understand the insect‘s interactions with their environment and insect evolution in the permanently developing biosphere, more than venation-pattern descriptions and sophisticated statistics centered on taxonomic data are needed. The thorough revision of the fossil record is crucial to improving the reliability of any popular biodiversity estimation based on fossil taxa. In respect thereof, we revise all gyroblattid species and reduce the genera number from 12 to 1 and the species number from almost 20 to only 2. Concerning several historical finds, we provide photographs combined with appropriate drawings for the first time.
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Zhang, Z.; Schneider, J.W.; Hong, Y. The most ancient roach (Blattida): A new genus and species from the earliest Late Carboniferous (Namurian) of China, with discussion on the phylomorphogeny of early blattids. J. Syst. Palaeontol. 2012, 11, 27–40. [Google Scholar] [CrossRef]
- Wei, D.; Béthoux, O.; Guo, Y.; Schneider, J.W.; Ren, D. New data on the singularly rare ‘cockroachoids’ from Xiaheyan (Pennsylvanian; Ningxia, China). Alcheringa Australas. J. Palaeontol. 2013, 37, 547–557. [Google Scholar] [CrossRef]
- Trümper, S.; Schneider, J.W.; Nemyrovska, T.; Korn, D.; Linnemann, U.; Ren, D.; Béthoux, O. Age and depositional environment of the Xiaheyan insect fauna, embedded in marine black shales (Early Pennsylvanian, China). Palaeogeogr. Palaeoclimatol. Palaeoecol. 2020, 538, 109444. [Google Scholar] [CrossRef]
- Handlirsch, A. Die Fossilen Insekten und die Phylogenie der Rezenten Formen; Wilhelm Engelmann: Leipzig, Germany, 1906–1908; pp. 1–1430. [Google Scholar]
- Schneider, J. Die Blattodea (Insecta) des Paläozoikum. Teil 1: Systematik, Ökologie und Biostratigraphie. Freib. Forsch. 1983, 382, 106–145. [Google Scholar]
- Hmich, D.; Schneider, J.W.; Saber, H.; Voigt, S.; El Wartiti, M. New continental Carboniferous and Permian faunas of Morocco—Implications for biostratigraphy, palaeobiogeography and palaeoclimate. In Non-marine Permian Biostratigraphy and Biochronology; Lucas, S.G., Cassinis, G., Schneider, J.W., Eds.; Geological Society: London, UK, 2006; Volume 265, pp. 297–324. [Google Scholar]
- Belahmira, A.; Schneider, J.W.; Scholze, F.; Saber, H. Phyloblattidae and Compsoblattidae (Insecta, Blattodea) from the late Carboniferous Souss basin, Morocco. J. Paleontol. 2019, 93, 945–965. [Google Scholar] [CrossRef]
- Schneider, J.W.; Scholze, F.; Ross, A.J.; Blake, B.M., Jr.; Lucas, S.G. Improved blattoid insect and conchostracan zonation for the late Carboniferous, Pennsylvanian, of Euramerica. Geol. Soc. Lond. Spec. Publ. 2021, 512, 865–891. [Google Scholar] [CrossRef]
- Handlirsch, A. Revision of American Paleozoic insects. Proc. USA Nat. Mus. 1906, 29, 661–820. [Google Scholar] [CrossRef]
- Schneider, J. Revision der Poroblattinidae (Insecta, Blattodea) des euro päischen und nordamerikanischen Oberkarbon und Perm. Freib. Forsch. 1978, 342, 55–66. [Google Scholar]
- Schneider, J.W.; Lucas, S.G.; Rowland, J.M. The Blattida (Insecta) fauna of Carrizo Arroyo, New Mexico—Biostratigraphic link between marine and non-marine Pennsylvanian/Permian boundary profiles. New Mex. Mus. Nat. Hist. Sci. Bull. 2004, 25, 247–261. [Google Scholar]
- Vršansky, P.; Vishniakova, V.N.; Rasnitsyn, A.P. Order Blattida Latreille, 1810. In History of Insects; Rasnitsyn, A.P., Quicke, D.L.J., Eds.; Kluwer Academic Publishers: Dordrecht, The Netherlands, 2002; pp. 263–270. [Google Scholar]
- Vršansky, P. Phyloblatta grimaldii sp. nov.—A New Triassic Cockroach (Insecta, Blattaria) from Virginia. Entomol. Probl. 2003, 33, 51–53. [Google Scholar]
- Vršansky, P. A New Genus and Species of Cockroach (Blattida: Phyloblattidae) from the Permian/Triassic Boundary Beds of Tunguska Basin in Eastern Siberia, Russia. Zootaxa 2010, 2353, 55–61. [Google Scholar] [CrossRef]
- Aristov, D.S.; Bashkuev, A.S.; Golubev, V.K.; Gorochov, A.V.; Karasev, E.V.; Kopylov, D.S.; Ponomarenko, A.G.; Rasnitsyn, A.P.; Rasnitsyn, D.A.; Sinitshenkova, N.D.; et al. Fossil insects of the middle and upper Permian of European Russia. Paleontol. J. 2013, 47, 641–832. [Google Scholar] [CrossRef]
- Vršanský, P. New blattarians and a review of dictyopteran assemblages from the Lower Cretaceous of Mongolia. Acta Palaeontol. Pol. 2008, 53, 129–136. [Google Scholar] [CrossRef]
- Gurney, A.B. The largest cockroach (Orthoptera, Blattoidea). Proc. Entomol. Soc. Wash. 1959, 61, 133–134. [Google Scholar]
- Durden, C.J. Pennsylvanian correlation using blattoid insects. Can. J. Earth Sci. 1969, 6, 1159–1177. [Google Scholar] [CrossRef]
- Rasnitsyn, A.P.; Quicke, D.L.J. History of Insects; Kluwer Academic Publishers: Dordrecht, The Netherlands, 2002; p. XII, 517. [Google Scholar]
- Opluštil, S.; Cleal, C.J.; Wang, J.; Wan, M. Carboniferous macrofloral biostratigraphy: An overview. In The Carboniferous Timescale; Lucas, S.G., Schneider, J.W., Wang, X., Nikolaeva, S., Eds.; Geological Society: London, UK, 2021; Volume 512, p. 51. [Google Scholar]
- Lucas, S.G. Rethinking the Carboniferous Chronostratigraphic Scale. Newsl. Stratigr. 2021, 54, 257–274. [Google Scholar] [CrossRef]
- Wagner, R.H. Proposal for recognition of a new ‘Cantabrian’ Stage at the base of the Stephanian Series. 6th Congr. Int. Strat. Geol. Carbonif. 1969, 1, 139–150. [Google Scholar]
- Heckel, P.H. Pennsylvanian stratigraphy of northern Midcontinent shelf and biostratigraphic correlation of cyclothems. Stratigraphy 2013, 10, 3–39. [Google Scholar]
- Lucas, S.G.; Schneider, J.W.; Nikolaeva, S.; Wang, X. The Carboniferous chronostratigraphic scale: History, status and prospectus. In The Carboniferous Timescale; Lucas, S.G., Schneider, J.W., Wang, X., Nikolaeva, S., Eds.; Geological Society: London, UK, 2021; Volume 512, pp. 19–48. [Google Scholar] [CrossRef]
- Schneider, J.W.; Lucas, S.G.; Scholze, F.; Voigt, S.; Marchetti, L.; Klein, H.; Opluštil, S.; Werneburg, R.; Golubev, V.K.; Barrick, J.E.; et al. Late Paleozoic-early Mesozoic continental biostratigraphy—Links to the Standard Global Chronostratigraphic Scale. Palaeoworld 2020, 531, 186–238. [Google Scholar] [CrossRef]
- Lameere, A. Sur la nervation alaire des Insectes. Bull. Cl. Sci. Acad. Roy. Belg. 1922, 8, 138–149. [Google Scholar]
- Linnaeus, C. Systema Naturae per Regna Tria Naturae, Secundum Classes, Ordines, Genera, Species, Cum Characteribus, Differentiis, Synonymis, Locis; Editio Decima, Reformata; Holmiae, Impensis Direct, Laurentii Salvii; Linnean Society of London: London, UK, 1758; p. 824. [Google Scholar]
- Latreille, P.A. Le Règne Animal Distribué d’après son Organisation, pour Servir de Base à L’histoire Naturelle des Animaux et D’Introduction à L’anatomie Compare; Nouvelle édition, rev. et aug. Tome IV. Crustacés, arachnides et partie des insects; Chez Déterville: Paris, France, 1829; Volume 4, pp. 1–584. [Google Scholar]
- Brunner von Wattenwyll, C. Prodromus der Europäischen Orthopteren; Wilhelm Engelmann: Leipzig, Germany, 1882; p. 466. [Google Scholar]
- Scudder, S.H. Palaeozoic Cockroaches: A Complete Revision of the Species of Both Worlds, with an Essay Toward Their Classification; Boston Society of Natural History: Boston, MA, USA, 1879; Volume 3, pp. 23–134. [Google Scholar]
- Sellards, E.H. A study of the structure of Paleozoic Cockroaches, with descriptions of new forms from the coal measures. Amer. J. Sci. 1904, 18, 213–227. [Google Scholar] [CrossRef]
- Schneider, J.W.; Scholze, F.; Germann, S.; Lucas, S.G. The Late Pennsylvanian nearshore insect fauna of the Kinney Brick Quarry Fossil Lagerstätte, New Mexico. New Mexico Mus. Nat. Hist. Sci. Bull. 2021, 82, 255–286. [Google Scholar]
- Hörnig, M.K.; Haug, C.; Schneider, J.W.; Haug, J.T. Evolution of reproductive strategies in dictyopteran insects—Clues from ovipositor morphology of extinct roachoids. Acta Palaeontol. Polon. 2018, 63, 1–24. [Google Scholar] [CrossRef]
- Sellards, E.H. Some New Structural Characters of Paleozoic Cockroaches. Amer. J. Sci. 1903, 15, 307–315. [Google Scholar] [CrossRef]
- Sellards, E.H. Megablattina Sellards (non Brongniart): A correction. Amer. J. Sci. 1903, 15, 488. [Google Scholar]
- Laurentiaux, D. Les insectes des bassins houillers du Gard et de la Loire. Ann. Paléontol. 1950, 46, 65–84. [Google Scholar]
- Guthörl, P. Zur Arthropoden-Fauna des Karbons und Perms. 18. Weitere Insektenfunde (Blattodea) aus dem saar-lothringischen Karbon. N. Jb. Geol. Paläont. Abh. 1963, 118, 245–259. [Google Scholar]
- Heer, O. Ueber die fossilen Kakerlaken. Vierteljahrsschr. Naturforsch. Ges. Zürich 1864, 9, 273–302. [Google Scholar]
- Schneider, J. Entwurf einer Zonengliederung für das euramerische Permokarbon mittels der Spiloblattinidae (Blattodea, Insecta). Freib. Forsch. 1982, 375, 27–47. [Google Scholar]
- Hmich, D.; Schneider, J.W.; Saber, H.; El Wartiti, M. Spiloblattinidae (Insecta, Blattida) from the Carboniferous of Morocco, North Africa—Implications for biostratigraphy. New Mexico Mus. Nat. Hist. Sci. Bull. 2005, 30, 111–114. [Google Scholar]
- Schneider, J. Taxonomie, Biostratigraphie und Palökologie der Blattodea-Fauna aus dem Stefan von Commentry (Frankreich)—Versuch einer Revision. Freib. Forsch. 1983, 384, 77–100. [Google Scholar]
- Béthoux, O.; Schneider, J.W.; Klass, K. Redescription of the holotype of Phyloblatta gaudryi Agnus, 1903 (Pennsylvanian; Commentry, France), an exceptionally well-preserved stem-dictyopteran. Geobios 2011, 33, 623–634. [Google Scholar]
- Scudder, S.H. Insect Fauna of the Rhode Island Coal Field; US Government Printing Office: Washington, DC, USA, 1893; Volume 101, pp. 1–21. [Google Scholar]
- Lyons, P.C. Biostratigraphy of the Pennsylvanian of Massachusetts and Rhode Island. In The Mississippian and Pennsylvanian (Carboniferous) Systems in the United States-Massachusetts, Rhode Island, and Maine; Professional Paper 1110-A-L; Skehan, J.W., Murray, D.P., Hepburn, J.C., Billings, M.P., Lyons, P.C., Doyle, R.G., Eds.; US Geological Survey: Reston, VA, USA, 1979; pp. A20–A24. [Google Scholar]
- Agnus, A.N. Deuxième note sur les Blattidés palèozoiques (Orthopt. palaeoz.). Description d’une espèce nouvelle. Bull. Soc. Entomol. Fr. 1903, 17, 291–294. [Google Scholar] [CrossRef]
- Meunier, F. Nouvelles recherches sur les insectes du terrain houiller de Commenrry (Allier) (troisième partie). Orthoptères Blattidae. Ann. Paléontol. 1921, 10, 59–108. [Google Scholar]
- Handlirsch, A. Neue Untersuchungen über die fossilen Insekten mit Ergänzungen und Nachträgen sowie Ausblicken auf phylogenetische, palaeogeographische und allgemein biologische Probleme. I. Teil. Ann. Naturhist. Mus. Wien 1937, 48, 1–140. [Google Scholar]
- Schneider, J. Zur Variabilität der Flügel paläozoischer Blattodea (Insecta), Teil 1. Freib. Forsch. 1977, 326, 87–105. [Google Scholar]
- Schneider, J. Zur Variabilität der Flügel paläozoischer Blattodea (Insecta), Teil 2. Freib. Forsch. 1978, 334, 21–39. [Google Scholar]
- Agnus, A.N. 1903a. Première note sur les Blattidés paléozoïques. Description d’un Mylacridae de Commentry Orthopt. Paléoz. Bull. Soc. Entomol. Fr. 1903, 8, 272–275. [Google Scholar] [CrossRef]
- Sterzel, J.T. Über einige neue Fossilreste. Ber. Naturwiss. Ges. Chemnitz 1904, 15, 69–72. [Google Scholar]
- Meunier, F. Une nouvelle espèce de Etoblattina (Orth.) du houiller supérieur de Fontanes (Gard). Bull. Soc. Entomol. France 1906, 11, 82–85. [Google Scholar] [CrossRef]
- Haudour, J.; Laurentiaux, D.; Sarrot-Reynauld, J. Présence d’ailes d’insectes dans le bassin houiller de la Mure (Isère). Travaux Lab. Geol. Fac. Sci. Grenoble 1960, 36, 133–141. [Google Scholar]
- Laurentiaux, D. Présence du genre Dysmenes Handlirsch, insectes blattaires, dans le Stephanian de la Creuse. Ann. Soc. Geol. du Nord 1966, 86, 189–191. [Google Scholar]
- Brauckmann, C.; Hahn, G. Ein Schaben-Flügel aus dem Ober-Karbon des Schwarzwaldes. Oberrheinische Geol. Abh. 1983, 32, 69–74. [Google Scholar]
- Schneider, J.W.; Hoth, K.; Gaitzsch, B.G.; Berger, H.J.; Steinborn, H.; Walter, H.; Zeidler, M. Carboniferous stratigraphy and development of the Erzgebirge Basin, East Germany. Z. Dt. Ges. Geowiss. 2005, 156, 431–466. [Google Scholar] [CrossRef]
- Lyons, P.C.; Krogh, T.E.; Kwok, Y.Y.; Davis, D.W.; Outerbridge, W.F.; Evans, H.T., Jr. Radiometric ages of the Fire Clay tonstein [Pennsylvanian (Upper Carboniferous), Westphalian, Duckmantian]: A comparison of U–Pb zircon single-crystal ages and 40Ar/39Ar sanidine single-crystal plateau ages. Int. J. Coal Geol. 2006, 67, 259–266. [Google Scholar] [CrossRef]
- Pointon, M.A.; Chew, D.M.; Ovtcharova, M.; Sevastopulo, G.D.; Crowley, Q.G. New high-precision U–Pb dates from western European Carboniferous tuffs; implications for time scale calibration, the periodicity of late Carboniferous cycles and stratigraphical correlation. J. Geol. Soc. Lond. 2012, 169, 713–721. [Google Scholar] [CrossRef]
- Opluštil, S.; Schmitz, M.; Cleal, C.J.; Martinek, K. A review of the Middle-Late Pennsylvanian west European regional substages and floral biozones and their correlation to the geological time scale based on new U–Pb ages. Earth Sci. Rev. 2016, 154, 301–335. [Google Scholar] [CrossRef]
- Cui, Y.; Brauner, S.; Schneider, J.W.; Béthoux, O. Grylloblattidan insects from Sperbersbach and Cabarz (Germany), two new early Permian and insect-rich localities. J. Paleontol. 2022, 96, 355–374. [Google Scholar] [CrossRef]
- Selden, P.A.; Nudds, J.R. Evolution of Fossil Ecosystems, 2nd ed.; Manson Publishing Ltd.: London, UK, 2012; p. 304. [Google Scholar]
- Clements, T.; Purnell, M.; Gabbott, S. The Mazon Creek Lagerstätte: A diverse late Paleozoic ecosystem entombed within siderite concretions. J. Geol. Soc. 2019, 176, 1–11. [Google Scholar] [CrossRef]
- Laurentiaux, D. Contribution à l’ètude des Insects Carbonifères. Ph.D. Thesis, Université de Paris, Paris, France, 1958. [Google Scholar]
- Pruvost, P. Les insectes houillers du Nord de la France. Ann. Soc. Géol. Nord 1912, 41, 323–380. [Google Scholar]
- Pruvost, P. Introduction à L’étude du Terrain Houiller du Nord et du Pas-de-Calais: La Faune Continentale du Terrain Houiller du Nord de la France; Imprimerie Nationale: Paris, France, 1919; p. 616. [Google Scholar]
- Pruvost, P. La Faune continentale du terrain houiller de la Belgique. Mém. Mus. Roy. Hist. Nat. Belg. 1930, 44, 103–282. [Google Scholar]
- Sellards, E.H. Cockroaches of the Kansas Coal Measures, and of the Kansas Permian. Univ. Geol. Surv. Kansas 1908, 9, 501–541. [Google Scholar]
- Schneider, J.W.; Lucas, S.G.; Trümper, S.; Stanulla, C.; Krainer, K. Carrizo Arroyo, central New Mexico—A new late Paleozoic taphotype of arthropod fossillagerstätte. New Mex. Geol. Soc. Guideb. 2016, 67, 377–386. [Google Scholar] [CrossRef]
- Opluštil, S.; Pešek, J. Stratigraphy, palaeoclimatology and palaeogeography of the Late Palaeozoic continental deposits in the Czech Republic. Geodiversitas 1998, 20, 597–620. [Google Scholar]
- Opluštil, S. Evolution of the Middle Westphalian river valley drainage system in central Bohemia (Czech Republic) and its palaeogeographic implication. Palaeogeogr. Palaeoclimatol. Palaeoecol. 2005, 222, 223–258. [Google Scholar] [CrossRef]
- Gaitzsch, B.; Rössler, R.; Schneider, J.W.; Schretzenmayr, S. Neue Ergebnisse zur Verbreitung potentieller Muttergesteine im Karbon von Nord- und Mitteldeutschland. Geol. Jahrb. A 1998, 149, 25–58. [Google Scholar]
- Hoth, K.; Brause, H.; Döring, H.; Kahlert, E.; Schultka St Volkmann, N.; Berger, H.-J.; Adam Ch Felix, M.; Wünsche, M. Die Steinkohlenlagerstätte Zwickau; Bergbau in Sachsen 15; Sächsisches Landesamt für Umwelt, Landwirtschaft und Geologie: Freiberg, Germany, 2009; pp. 1–162. [Google Scholar]
- Hartlieb, J. Kohlenpetrographie des Lugau-Oelsnitzer und Zwickauer Reviers im Erzgebirgischen Becken. Fortschr. Geol. Rheinl. Westf. 1964, 12, 429–450. [Google Scholar]
- Gutbier, A. Abdrücke und Versteinerungen des Zwickauer Schwarzkohlengebirges und Seiner Umgebungen; G. Richtersche Buchhandlung: Zwickau, Germany, 1835; Volume 36, pp. 1–80. [Google Scholar]
- Sterzel, J.T. Palaeontologischer Charakter der oberen Steinkohlenformation und des Rothliegenden im Erzgebirgischen Becken. Ber. Naturwiss. Ges. Chemnitz 1881, 7, 155–270. [Google Scholar]
- Sterzel, J.T. Paläontologischer Charakter der Steinkohleformation von Zwickau und die Genesis der Dortigen Flötze. In Erläuterungen zur geologischen Specialkarte des Königreichs Sachsen, Section Zwickau-Werdau, Blatt 111, 2nd ed.; Siegert, T., Ed.; W. Engelmann: Leipzig, Germany, 1901; pp. 87–123. [Google Scholar]
- Döring, H. Sporenstratigraphische Untersuchungen in der Zwickau-Formation. In Die Steinkohlenlagerstätte Zwickau, 1st ed.; Hoth, K., Brause, H., Döring, H., Kahlert, E., Schultka, S., Volkmann, N., Berger, H.-J., Adam, C., Felix, M., Wünsche, M., Eds.; Bergbau in Sachsen 15; Sächsisches Landesamt für Umwelt, Landwirtschaft und Geologie: Freiberg, Germany, 2009; pp. 90–101. [Google Scholar]
- Kahlert, E.; Schultka, S. Makroflora der Zwickau-Formation. In Die Steinkohlenlagerstätte Zwickau, 1st ed.; Hoth, K., Brause, H., Döring, H., Kahlert, E., Schultka, S., Volkmann, N., Berger, H.-J., Adam, C., Felix, M., Wünsche, M., Eds.; Bergbau in Sachsen 15; Sächsisches Landesamt für Umwelt, Landwirtschaft und Geologie: Freiberg, Germany, 2009; pp. 72–90. [Google Scholar]
- Gutbier, A. Geognostische Beschreibung des Zwickauer Schwarzkohlengebirges und Seiner Umgebung; G. Richtersche Buchhandlung: Zwickau, Germany, 1834; pp. 1–160. [Google Scholar]
- Daber, R. Die Parallelisierung der Flöze des Zwickauer und Lugau-Oelsnitzer Steinkohlenreviers auf Grund paläobotanischer Untersuchungen. Geol. Beih. 1957, 19, 1–76. [Google Scholar]
- Döring, H.; Hoth, K.; Kahlert, E. Gegenwärtiger Stand der litho- und sporostratigraphischen Gliederung des Zwickauer Siles. Freib. Forsch. 1988, 419, 18–29. [Google Scholar]
- Döring, H.; Hoth, K.; Lobst, R. Korrelation des Oberkarbons von Lugau/Oelsnitz mit dem von Zwickau aus sporologischer und lithostratigraphischer Sicht. Geoprofil 2010, 13, 45–53. [Google Scholar]
- Barthel, M. Die Gattung Dicranophyllum Gr. Eury in den varistischen Innensenken der DDR. Hallesches Jahrb. Für Geowiss. 1977, 2, 73–86. [Google Scholar]
- Daber, R. Die Geologie und die Entstehung der Steinkohlen von Zwickau und Oelsnitz. Wiss. Z. Humboldt-Univ. Math.-Nat. R. 1992, 41, 71–80. [Google Scholar]
- Daber, R. Ein wertvoller Fund—Freigelegt durch das Augusthochwasser 2002 der Mulde in Zwickau. Veröff. Mus. Nat. Chemnitz 2002, 25, 127–128. [Google Scholar]
- Wagner, R.H. Dicranophyllum glabrum (Dawson) Stopes, an unusual element of Lower Westphalian floras in Atlantic Canada. Rev. Españ. Paleontol. 2005, 20, 7–13. [Google Scholar] [CrossRef]
- Lobin, M. Aufbau und Entwicklung des Permosiles im mittleren und östlichen Erzgebirge. Ph.D. Thesis, Bergakademie Freiberg, Freiberg, Germany, 1986; pp. 1–87. [Google Scholar]
- Rößler, R.; Barthel, M. Rotliegend taphocoenoses preservation favoured by rhyolitic explosive volcanism. Freib. Forsch. 1998, 474, 59–101. [Google Scholar]
- Donovan, M.P.; DiMichele, W.A.; Lucas, S.G.; Schneider, J.W. Atlas of selected Kinney Quarry plant fossils, Late Pennsylvanian, central New Mexico. New Mex. Mus. Nat. Hist. Sci. Bull. 2021, 84, 153–183. [Google Scholar]
- Grand’Eury, C. Flore carbonifère du Départment de la Loire et du centre de la France. Acad. Sci. Inst. Fr. Paris 1877, 24, 208–220. [Google Scholar]
- Grand’Eury, M.C. Géologie et Paléontologie du Basin Houiller du Gard; Fréderic Lantz: Saint-Étienne, France, 1890; pp. 1–324. [Google Scholar]
- Renault, B. Études sur le Terrain Houiller de Commentry. Livre 2, Flore Fossile, 2ème Partie; Theolier et Cie: Saint-Étienne, France, 1890; pp. 1–746. [Google Scholar]
- Broutin, J.; Ferrandini, J.; Saber, H. Implications stratigraphiques et paléogéographiques dela découverte d’une flore permienne euraméricaine dans le Haut-Atlas occidental (Maroc). In Comptes rendus de l’Académie des sciences. Série 2, Mécanique, Physique, Chimie, Sciences de l’univers, Sciences de la Terre; Gauthier-Villars: Paris, France, 1989; Volume 308, pp. 1509–1515. [Google Scholar]
- Daber, R. Pflanzengeographische Besonderheiten der Karbonflora des Zwickau-Lugauer Steinkohlenreviers. Geologie 1955, 13, 1–44. [Google Scholar]
- Rößler, R.; Dunlop, J.A. Redescription of the largest trigonotarbid arachnid—Kreischeria wiedei Geinitz 1882 from the Upper Carboniferous of Zwickau, Germany. Paläont. Z. 1997, 71, 237–245. [Google Scholar] [CrossRef]
- Barthel, M. Die Rotliegendflora des Thüringer Waldes Teil 4: Farnsamer und Farnlaub unbekannter taxonomischer Stellung. Veröff. Naturhist. Mus. Schleus. 2006, 21, 33–72. [Google Scholar]
- Laveine, J.-P.; Oudoire, T. Morphological analysis of the Grand’Eury 1890 large frond fragment attributed to Odontopteris reichiana, from the Late Pennsylvanian (Stephanian) of the Gard coalfield (SE of Massif Central, France). Morphological and taxonomical considerations on the odontopterids. Erection of Cyrillopteris gen. nov. Palaeontographica B 2016, 294, 1–84. [Google Scholar]
- Doubinger, J.; Vetter, P.; Langiaux, J.; Galtier, J.; Broutin, J. La flore fossile du bassin houiller de Saint-Etienne. Mém. Mus. Nat. Hist. Nat. Paris 1995, 164, 1–355. [Google Scholar]
- Looy, C.V.; Kerp, H.; Duijnstee, I.A.P.; DiMichele, W.A. The late Paleozoic ecological–evolutionary laboratory, a land–plant fossil record perspective. Sed. Record 2014, 12, 4–10. [Google Scholar] [CrossRef]
- Thomas, B.A.; Cleal, C.J. Distinguishing Pennsylvanian-age lowland, extra-basinal and upland vegetation. Palaeobiodivers. Palaeoenviron. 2017, 97, 273–293. [Google Scholar] [CrossRef]
- Sterzel, J.T. Über zwei neue Insektenarten aus dem Karbon von Lugau. Ber. Naturwiss. Ges. Chemnitz 1881, 7, 271–276. [Google Scholar]
- Handlirsch, A. Über die fossilen Insekten aus dem mittleren Oberkarbon des Königreiches Sachsen. Mitt. Geol. Ges. Wien 1909, 2, 373–381. [Google Scholar]
- Angelier, C.-H. Le Bassin houiller de La Mure. Rev. Géogr. Alpine 1940, 28, 249–346. [Google Scholar] [CrossRef]
- Haudour, J.; Sarrot-Reynauld, J. Le Bassin houiller de la Mure, ses Minéraux. Trav. Lab. Geol. Fac. Sci. Grenoble 1956, 32, 15–19. [Google Scholar]
- Bouroz, A.; Corsin, P. Sur l’âge du Houiller du Bassin de la Mure et de la zone externe des Alpes. C. R. Acad. Sci. Paris 1950, 220, 2035–2037. [Google Scholar]
- Guillot, S.; Ménot, R.-P. Paleozoic evolution of the External Crystalline Massifs of the Western Alps. C. R. Geosci. 2009, 341, 253–265. [Google Scholar] [CrossRef]
- Fréville, K.; Trap, P.; Faure, M.; Melleton, J.; Li, X.H.; Lin, W.; Blein, O.; Bruguier, O.; Poujol, M. Structural, metamorphic and geochronological insights on the Variscan evolution of the Alpine basement in the Belledonne massif (France). Tectonophysics 2018, 726, 14–42. [Google Scholar] [CrossRef]
- Ballèvre, M.; Manzotti, P.; Dal Piaz, G.V. Pre-Alpine (Variscan) inheritance: A key for the location of the future Valaisan Basin (Western Alps). Tectonics 2018, 37, 786–817. [Google Scholar] [CrossRef]
- Rastel, D.; Vallois, B.; Jouve, A.; Belleserre, G. Présentation et évaluation préliminaire de l’intérêt patrimonial du site fossilifère du Carbonifère supérieur alpin de Vaulnaveys-le-Bas (Isère, France). Bull. Mens. Soc. Linn. Lyon 2019, 88, 167–187. [Google Scholar]
- Von Raumer, J. The Palaeozoic evolution in the Alps: From Gondwana to Pangea. Geol. Rundschau 1998, 87, 407–435. [Google Scholar] [CrossRef]
- Pilloud, C. Structure de déformation alpines dans le synclinal de Permo-Carbonifere de Salvan-Dorénaz, (massif des Aiguilles Rouges, Valais). Mém. Géol. (Lausanne) 1991, 9, 101. [Google Scholar]
- Niklaus, P.-A.; Wetzel, A. Faziesanalyse und Ablagerungsmilieu der fluviatilen Sedimentfüllung des Karbontroges von Salvan-Dorénaz. Eclogae Geol. Helv. 1996, 89, 427–437. [Google Scholar]
- Capuzzo, N.; Wetzel, A. Facies and basin architecture of the Late Carboniferous Salvan-Dorénaz continental basin (Western Alps, Switzerland/France). Sedimentology 2004, 51, 675–697. [Google Scholar] [CrossRef]
- Jongmans, W.J. Die Karbonflora der Schweiz. Beiträge Zur Geol. Kt. Der Schweiz 1960, 108, 1–97. [Google Scholar]
- Capuzzo, N.; Bussy, F. High-precision dating and origin of synsedimentary volcanism in the Late Carboniferous Salvan-Dorénaz basin (Aiguilles-Rouges Massif, Western Alps). Schweiz. Min. Petrogr. Mitt. 2000, 80, 147–167. [Google Scholar]
- Sterzel, J.T. Die Flora des Rotliegenden von Oppenau im badischen Schwarzwalde (Blatt Petersthal-Reichenbach). Mitt. Grossherzogtum Baden Geol. Landesanst. 1895, 3, 259–352. [Google Scholar]
- Nitsch, E.; Zedler, H. Oberkarbon und Perm in Baden-Württemberg. LGRB-Inf. 2009, 22, 7–102. [Google Scholar]
- Sandberger, F. Über Steinkohlenformation und Rothliegendes im Schwarzwald und deren Floren. Jb. K.-K. Reichsanst. Wien 1890, 40, 77–102. [Google Scholar]
- Sterzel, J.T. Die Karbon- und Rotliegendflora im Großherzogtum Baden. Mitt. Grossherzogtum Baden Geol. Landesanst. 1907, 5, 345–892. [Google Scholar]
- Hartkopf-Fröder, C. 4.2 Palynologische Befunde. In Oberkarbon und Perm in Baden-Württemberg; Nitsch, E., Zedler, H., Eds.; LGRB-Informationen: Freiburg, Germany, 2009; Volume 22, pp. 30–31. [Google Scholar]
- Clayton, G.; Coquel, R.; Doubinger, J.; Gueinn, K.J.; Loboziak, S.; Owen, B.; Streel, M. Carboniferous miospores of Western Europe: Illustrations and zonations. Meded. -Rijks Geol. Dienst 1977, 29, 1–71. [Google Scholar]
- Hochuli, P.A. Palynostratigraphische Gliederung und Korrelation des Permokarbons der Nordschweiz. Eclogae Geol. Helv. 1985, 78, 719–831. [Google Scholar]
- Czygan, W.W. Das Jung-Paläozoikum an der Hohengeroldseck bei Lahr im mittleren Schwarzwald. Ber. Naturf. Ges. Freibg. I. Br. 1964, 54, 87–106. [Google Scholar]
- Saber, H. The Stephano-Permian of the western High Atlas: Geological and geodynamic evolution (Morocco). Ph.D. Thesis, Chouaïb Doukkali University, El Jadida, Morocco, 1998; p. 212. [Google Scholar]
- Saber, H.; El Wartiti, M.; Broutin, J. Dynamique sédimentaire comparative dans les bassins Stéphano-Permiens des Ida Ou Zal et Ida Ou Ziki, Haut Atlas Occidental. Maroc. J. African Earth Sci. 2001, 32, 573–594. [Google Scholar] [CrossRef]
- Clariond, L. Sur le Stéphanien des Ida ou Zal (Maroc occidental). C. R. Acad. Sci. Paris Ser. II 1932, 195, 62–64. [Google Scholar]
- Jongmans, W.J. Notes sur la flore du Carbonifère du versant Sud du Haut-Atlas. Notes Mém. Serv. Géol. Maroc 1950, 76, 155–172. [Google Scholar]
- Feys, R.; Greber, C. Le Stéphanian et l’Autunien du Souss dans les Ida ou Zal (Haut-Atlas occidental—Maroc). Notes Et Mém. Serv. Géol. Maroc 1963, 22, 19–43. [Google Scholar]
- Barthel, M. Alethopteris subelegans (H. Potonié) Franke ist ein Farn. Z. geol. Wiss. 1981, 9, 921–927. [Google Scholar]
- Schneider, J.W.; Werneburg, R. Biostratigraphie des Rotliegend mit Insekten und Amphibien. In Stratigraphie von Deutschland X. Rotliegend. Teil I: Innervariscische Becken; Lützner, H., Kowalczyk, G., Eds.; Schriftenreihe der Deutschen Gesellschaft für Geowissenschaften: Stuttgart, Germany, 2012; Volume 61, pp. 110–142. [Google Scholar]
- Doubinger, J. Contribution à l’étude des flores Autuno-Stéphaniennes. Mém. Soc. Géol. Fr. 1956, 75, 1–180. [Google Scholar]
- Broutin, J.; Doubinger, J.; Farjanel, G.; Freytet, P.; Kerp, H.; Langiaux, J.; Lebreton, M.L.; Sebban, S.; Satta, S. Le renouvellement des flores au passage Carbonifère Permian: Approche stratigraphique, biologique, sédimentologique. C.R. Acad. Sci. Paris II 1990, 311, 1563–1569. [Google Scholar]
- Belahmira, A.; Hmich, D.; Schneider, J.W.; Saber, H.; Lagnaoui, A.; Hminna, A. Evidence of insect-plant interaction in the Late Carboniferous Oued Issene Formation, Souss basin (Western High Atlas Mountains, Morocco). Arab. J. Earth Sci. 2015, 2, 6–7. [Google Scholar]
- McCann, T.; Skompski, S.; Poty, E.; Dusar, M.; Vozárová, A.; Schneider, J.; Wetzel, A.; Krainer, K.; Kornpihl, K.; Schäfer, A.; et al. Carboniferous. In The geology of Central Europe; McCann, T., Ed.; Precambrian and Palaeozoic; The Geological Society: London, UK, 2008; Volume 1, pp. 411–529. [Google Scholar]
- Courel, L. Intramontane Stephanian and Permian coal basins of the French Massif Central new sedimentological data. In Sedimentation in a Synorogenic Basin Complex, the Upper Carboniferous of Northwest Europe; Besly, B., Kelling, G., Eds.; Blackie, Chapman and Hall: New York, NY, USA, 1988; pp. 255–263. [Google Scholar]
- Fayol, H. Etudes sur le Terrain Houiller de Commentry. Ire. partie. Lithologie et Stratigraphie; Imprimerie Théolier: Saint-Etienne, France, 1887. [Google Scholar]
- Stevenson, J.J. The Coal Basin of Commentry in Central France. Ann. N.Y. Acad. Sci. 1910, 19, 161–204. [Google Scholar] [CrossRef]
- Carpenter, F.M. Studies on Carboniferous insects from Commentry, France; Part, I. Introduction and families Protagriidae, Meganeuridae, and Campylopteridae. Bull. Geol. Soc. Amer. 1943, 54, 527–554. [Google Scholar] [CrossRef]
- Bolton, H. Insects from the Coal Measures of Commentry; Fossil Insects, no. 2; British Museum of Natural History: London, UK, 1925; pp. 1–56. [Google Scholar]
- Schneider, J. Zur Taxonomie und Biostratigraphie der Blattodea (Insecta) des Karbon und Perm der DDR. Freib. Forsch. 1978, 340, 1–152. [Google Scholar]
- Nel, A.; Fleck, G.; Garrouste, R.; Gand, G.; Lapeyrie, J.; Bybee, S.M.; Prokop, J. Revision of Permo-Carboniferous griffenflies (Insecta: Odonatoptera: Meganisoptera) based upon new species and redescription of selected poorly known taxa from Eurasia. Palaeontogr. A 2009, 289, 89–121. [Google Scholar] [CrossRef]
- Zessin, W. Zur Taxonomie der jungpaläozoischen Familie Meganeuridae (Odonata) unter Einbeziehung eines Neufundes aus dem Stefan C der Halleschen Mulde (DDR). Freib. Forsch. 1983, 384, 58–76. [Google Scholar]
- Schulz, M.; Schneider, J.W.; Beckemeyr, R. Taphonomy of large cockroach wings of the Hamilton Quarry fossil lagerstätte, Late Pennsylvanian, Kansas. In Palaeobiology and Geobiology of Fossil Lagerstätten through Earth History, Proceedings of the Joint Conference of the “Paläontologische Gesellschaft” and the “Palaeontological Society of China”, Göttingen, Germany, 23–27 September 2013; Universitätsverlag Göttingen: Göttingen, Germany, 2013; Volume 150, p. 150. [Google Scholar]
- Schneider, J. Zur Entomofauna des Jungpaläozoikums der Boskovicer Furche (CSSR), Teil 1: Mylacridae (Insecta, Blattodea). Freib. Forsch. 1980, 357, 43–55. [Google Scholar]
- Schneider, J. Zur Entomofauna des Jungpaläozoikums der Boskovicer Furche (CSSR), Teil 2: Phyloblattidae (Insecta, Blattodea). Freib. Forsch. 1984, 395, 19–37. [Google Scholar]
- Roscher, M.; Schneider, J.W. Permo-Carboniferous climate: Early Pennsylvanian to Late Permian climate development of central Europe in a regional and global context. In Non-Marine Permian Biostratigraphy and Biochronology; Lucas, S.G., Cassinis, G., Schneider, J.W., Eds.; Geological Society: London, UK, 2006; Volume 265, pp. 95–136. [Google Scholar]
- Sellards, E.H. Fossil plants of the Upper Paleozoic of Kansas. Univ. Geol. Surv. Kans. (Spec. Rep. Oil Gas) 1908, 9, 386–480. [Google Scholar]
- Bashforth, A.R.; DiMichele, W.A.; Eble, C.E.; Falcon-Lang, H.J.; Looy, C.V.; Lucas, S.G. The environmental implications of upper Paleozoic plant-fossil assemblages with mixtures of wetland and drought-tolerant taxa in tropical Pangea. Geobios 2021, 68, 1–45. [Google Scholar] [CrossRef]
- Cridland, A.A.; Morris, J.E. Spermopteris, a new genus of pteridosperms from the Upper Pennsylvanian Series of Kansas. Am. J. Bot. 1960, 47, 855–859. [Google Scholar] [CrossRef]
- Cridland, A.A.; Morris, J.E. Taeniopteris, Walchia and Dichophyllum in the Pennsylvanian System of Kansas. Univ. Kansas Sci. Bull. 1963, 44, 71–85. [Google Scholar]
- Ricetti, J.H.Z.; Schneider, J.W.; Iannuzzi, R.; Weinschütz, L.C. Anthracoblattina mendesi Pinto et Sedor (Blattodea, Phyloblattidae): The most completely preserved South American Palaeozoic cockroach. Rev. Brasil. Paleontol. 2016, 19, 181–194. [Google Scholar] [CrossRef]
- Schneider, J.W.; Legler, B.; Brosig, A.; Krainer, K.; Lucas, S.G. Sedimentology and depositional environment of the Kinney Brick Quarry Fossil Lagerstätte (Missourian, Late Pennsylvanian), Central New Mexico. New Mex. Mus. Nat. Hist. Sci. Bull. 2021, 84, 93–123. [Google Scholar]
- DiMichele, W.A.; Donovan, M.P.; Schneider, J.W.; Lucas, S.G. Plant-fossil taphonomy, Kinney Quarry, Late Pennsylvanian age, New Mexico, USA. New Mex. Mus. Nat. Hist. Sci. Bull. 2021, 84, 125–151. [Google Scholar]
- Teixeira, C. Nouveaux insects du Stéphanien Portugais. Bol. Soc. Geol. Port. 1941, 1, 13–30. [Google Scholar]
- Correia, P.A.P. Contribution to the knowledge of the fossil flora and fauna of the Douro Carboniferous Basin (NW of Portugal). Ph.D. Thesis, University of Porto, Porto, Portugal, 2016; p. 258. [Google Scholar]
- Correia, P.; Schubnel, T.; Nel, A. What is the roachoid genus Eneriblatta (Dictyoptera: Phyloblattidae) from the Carboniferous of Portugal. Hist. Biol. 2019, 33, 777–782. [Google Scholar] [CrossRef]
- Cecil, C.B. Paleoclimate controls on stratigraphic repetition of chemical and siliciclastic rocks. Geology 1990, 18, 533–536. [Google Scholar] [CrossRef]
- DiMichele, W.A.; Phillips, T.L. Paleobotanical and paleoecological constraints on models of peat formation in the Late Carboniferous of Euramerica. Palaeogeogr. Palaeoclimatol. Palaeoecol. 1994, 106, 39–90. [Google Scholar] [CrossRef]
- DiMichele, W.A.; Tabor, N.J.; Chaney, D.S.; Nelson, W.J. From wetlands to wet spots: Environmental tracking and the fate of Carboniferous elements in early Permian tropical floras. In Wetlands through Time; GSA Special Papers; Geological Society of America: Boulder, CO, USA, 2006; Volume 399, pp. 223–248. [Google Scholar] [CrossRef]
- Cleal, C.J. The Westphalian-Stephanian macrofloral record from the South Wales Coalfield. Geol. Mag. 2007, 144, 465–486. [Google Scholar] [CrossRef]
- Cleal, C.J.; Uhl, D.; Cascales-Miñana, B.; Thomas, B.A.; Bashforth, A.R.; King, S.C.; Zodrow, E.L. Plant biodiversity changes in Carboniferous tropical wetlands. Earth-Sci. Rev. 2012, 114, 124–155. [Google Scholar] [CrossRef]
- DiMichele, W.A. Wetland-dryland vegetational dynamics in the Pennsylvanian ice age tropics. Int. J. Plant Sci. 2014, 175, 123–164. [Google Scholar] [CrossRef]
- DiMichele, W.A.; Bashforth, A.R.; Falcon-Lang, H.J.; Lucas, S.G. Uplands, lowlands, and climate: Taphonomic megabiases and the apparent rise of a xeromorphic, drought-tolerant flora during the Pennsylvanian-Permian transition. Palaeogeogr. Palaeoclimatol. Palaeoecol. 2020, 559, 109965. [Google Scholar] [CrossRef]
- Kerp, H. Post-Variscan late Palaeozoic northern Hemisphere gymnosperms: The onset to the Mesozoic. Rev. Palaeobot. Palynol. 1996, 90, 263–285. [Google Scholar] [CrossRef]
- Kerp, H. The modernization of landscapes during the Late Paleozoic–Early Mesozoic. Paleontol. Soc. Pap. 2000, 6, 79–113. [Google Scholar] [CrossRef]
- Barthel, M.; Eichler, B.; Reichel, W. Die Rotliegendflora des Weißig-Beckens. Geol. Saxonica 2010, 56, 159–192. [Google Scholar]
- Rößler, R. The most entirely known Permian terrestrial ecosystem on Earth—Kept by explosive volcanism. Palaeontogr. B 2021, 303, 1–75. [Google Scholar] [CrossRef]
- Opluštil, S.; Jirásek, J.; Schmitz, M.; Matýsek, D. Biotic changes around the radioisotopically constrained Carboniferous–Permian boundary in the Boskovice Basin (Czech Republic). Bull. Geosci. 2017, 92, 95–122. [Google Scholar] [CrossRef]
- Werneburg, R.; Schneider, J.W. New branchiopod crustaceans from the late Carboniferous and early Permian of the Thuringian Forest Basin, Germany, with a review of Permian notostracans from the Lodève basin, France. Semana 2022, 37, 57–103. [Google Scholar]
- Walter, H. Zur Taxonomie, Ökologie und Biostratigraphie der Ichnia limnisch- terrestrischer Arthropoden des mitteleuropäischen Jungpaläozoikums. Freib. Forsch. 1983, 382, 146–193. [Google Scholar]
- Minter, N.J.; Braddy, S.J.; Voigt, S. Die Arthropodenfährten aus dem Permokarbon des Saar-Nahe-Beckens In Kohlesümpfe, Seen und Halbwüsten—Dokumente einer rund 300 Millionen Jahre alten Lebewelt zwischen Saarbrücken und Mainz; Schindler, T., Heidtke, U.H.C., Eds.; Pollichia: Bad Dürkheim, Germany, 2007; pp. 198–205. [Google Scholar]
- Buchwitz, M.; Marchetti, L.; Jansen, M.; Falk, D.; Trostheide, F.; Schneider, J.W. Ichnotaxonomy and trackmaker assignment of tetrapod tracks and swimming traces from the middle Permian Hornburg formation of Saxony-Anhalt (Germany). Ann. Soc. Geol. Poloniae 2020, 90, 291–320. [Google Scholar] [CrossRef]
- Handlirsch, A. Über einige Insektenreste aus der Permformation Russlands. Mem. Acad. Imp. Sci. St.-Petersbourg 1904, 16, 1–8. [Google Scholar]
- Lin, Q.B. On the fossil Blattoidea of China. Acta Entomol. Sinica 1978, 21, 335–342. [Google Scholar]
- Berner, R.A. GEOCARB II: A revised model of atmospheric CO2 over Phanerozoic time. Am. J. Sci. 1994, 294, 56–91. [Google Scholar] [CrossRef]
- Berner, R.A. Phanerozoic atmospheric oxygen: New results using the GEOCARBSULF model. Am. J. Sci. 2009, 309, 603–606. [Google Scholar] [CrossRef]
- Montañez, I.P.; McElwain, J.C.; Poulsen, C.J.; White, J.D.; DiMichele, W.A.; Wilson, J.P.; Griggs, G.; Hren, M.T. Climate, pCO2 and terrestrial carbon cycle linkages during late Palaeozoic glacial-interglacial cycles. Nat. Geosci. 2016, 9, 824–828. [Google Scholar] [CrossRef]
- Graham, J.B.; Dudley, R.; Aguilar, N.; Gans, C. Implications of the late Palaeozoic oxygen pulse for physiology and evolution. Nature 1995, 375, 117–120. [Google Scholar] [CrossRef]
- Dudley, R. Atmospheric oxygen, giant Paleozoic insects and the evolution of aerial locomotor performance. J. Experim. Biol. 1998, 201, 81043–81050. [Google Scholar] [CrossRef]
- Wootton, R.J.; Kukalova-Peck, J. Flight adaptations in Palaeozoic Palaeoptera (Insecta). Biol. Rev. 2000, 75, 129–167. [Google Scholar] [CrossRef]
- Harrison, J.F.; Kaiser, A.; Van den Brooks, J.M. Atmospheric oxygen level and the evolution of insect body size. Proc. R. Soc. B Biol. Sci. 2010, 277, 1937–1946. [Google Scholar] [CrossRef] [PubMed]
- Schneider, J.W.; Lucas, S.G.; Werneburg, R.; Röβler, R. Euramerican Late Pennsylvanian/Early Permian arthropleurid/tetrapod associations–implications for the habitat and paleobiology of the largest terrestrial arthropod. Carboniferous–Permian transition in Canon del Cobre, northern New Mexico. New Mex. Mus. Nat. Hist. Sci. Bull. 2010, 49, 49–70. [Google Scholar]
- Schneider, J.W.; Werneburg, R. Arthropleura, der größte landlebende Arthropode der Erdgeschichte—Neue Funde und neue Ideen. Semana 2010, 25, 75–100. [Google Scholar]
- Davis, N.S.; Russell, J.G.; McMahon, W.J.; Schneider, J.W.; Shillito, A.P. The largest arthropod in Earth history: Insights from newly discovered Arthropleura remains (Serpukhovian Stainmore Formation, Northumberland, England). J. Geol. Soc. 2021, 179, 18. [Google Scholar] [CrossRef]
- Schachat, S.R.; Labandeira, C.C.; Saltzman, M.R.; Cramer, B.D.; Payne, J.L.; Boyce, C.K. Phanerozoic pO2 and the early evolution of terrestrial animals. Proc. R. Soc. B Biol. Sci. 2018, 285, 20172631. [Google Scholar] [CrossRef] [PubMed]










Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Schneider, J.W.; Rößler, R. The Early History of Giant Cockroaches: Gyroblattids and Necymylacrids (Blattodea) of the Late Carboniferous. Diversity 2023, 15, 429. https://doi.org/10.3390/d15030429
Schneider JW, Rößler R. The Early History of Giant Cockroaches: Gyroblattids and Necymylacrids (Blattodea) of the Late Carboniferous. Diversity. 2023; 15(3):429. https://doi.org/10.3390/d15030429
Chicago/Turabian StyleSchneider, Joerg W., and Ronny Rößler. 2023. "The Early History of Giant Cockroaches: Gyroblattids and Necymylacrids (Blattodea) of the Late Carboniferous" Diversity 15, no. 3: 429. https://doi.org/10.3390/d15030429