

Reproductive Biology and Pollination Ecology of Fritillaria michailovskyi Fomin (Liliaceae), Endemic to East Anatolia (Turkey)



Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Material and Study Site
2.2. Detection of Pollination Strategies
2.3. Detection of Flowering Dynamics
2.4. Detection of Pollen Viability and Stigma Receptivity
2.5. Detection of the Self-Incompatibility Index (SII) and Pollen/Ovule Ratio (P/O)
2.6. Detection of Seed Viability and Germination
2.7. Detection of Floral Visitors
3. Results
3.1. Floral Features and Reproductive Phenology of F. michailovskyi
3.2. Flowering Phenology of F. michailovskyi
3.3. Self-Incompatibility Index (SII) and Pollen/Ovule (P/O) Ratio of F. michailovskyi
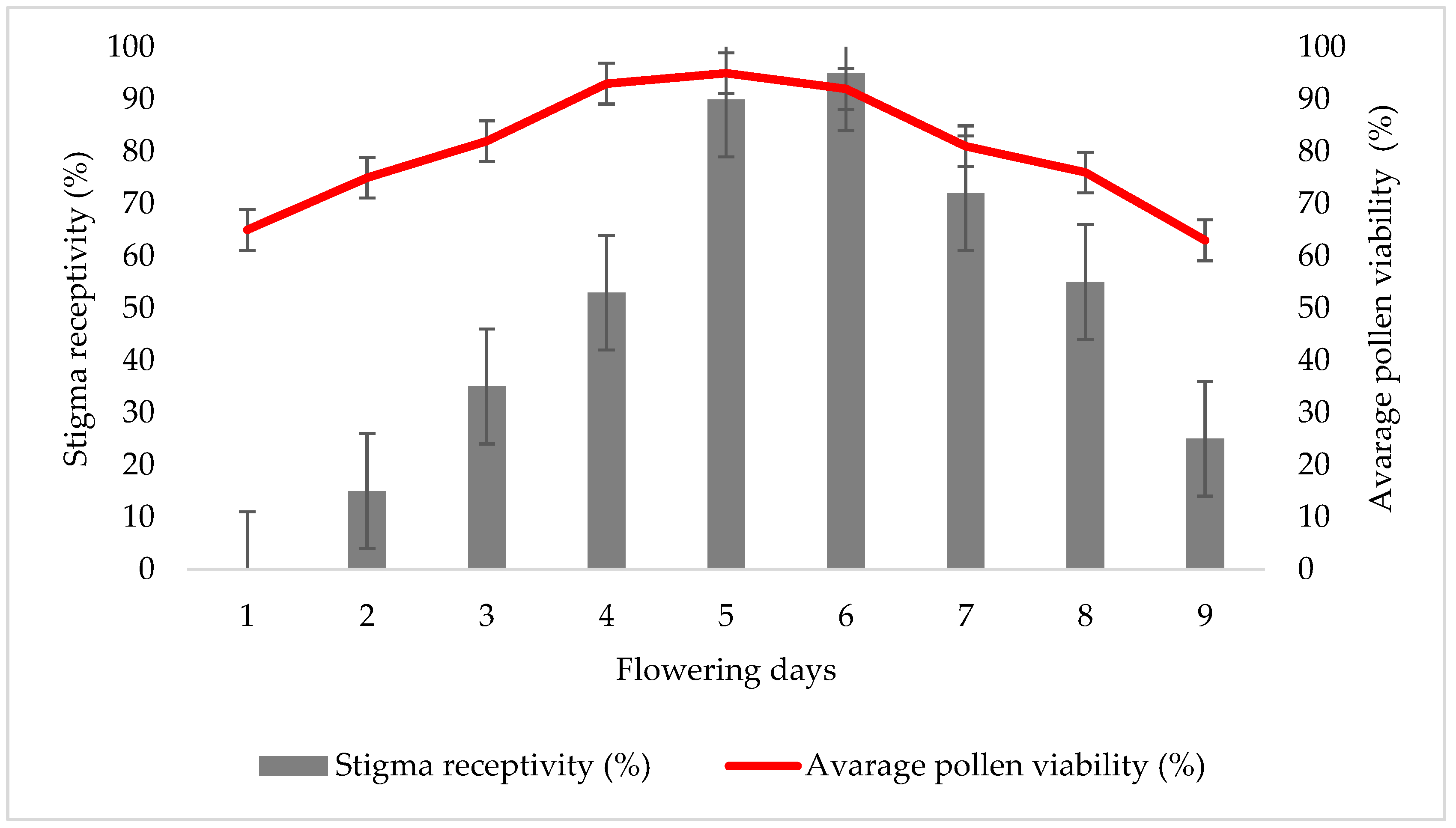
3.4. Pollen Viability and Stigma Receptivity of F. michailovskyi
3.5. Pollination Experiments of F. michailovskyi
3.6. Germination and Seed Viability of F. michailovskyi
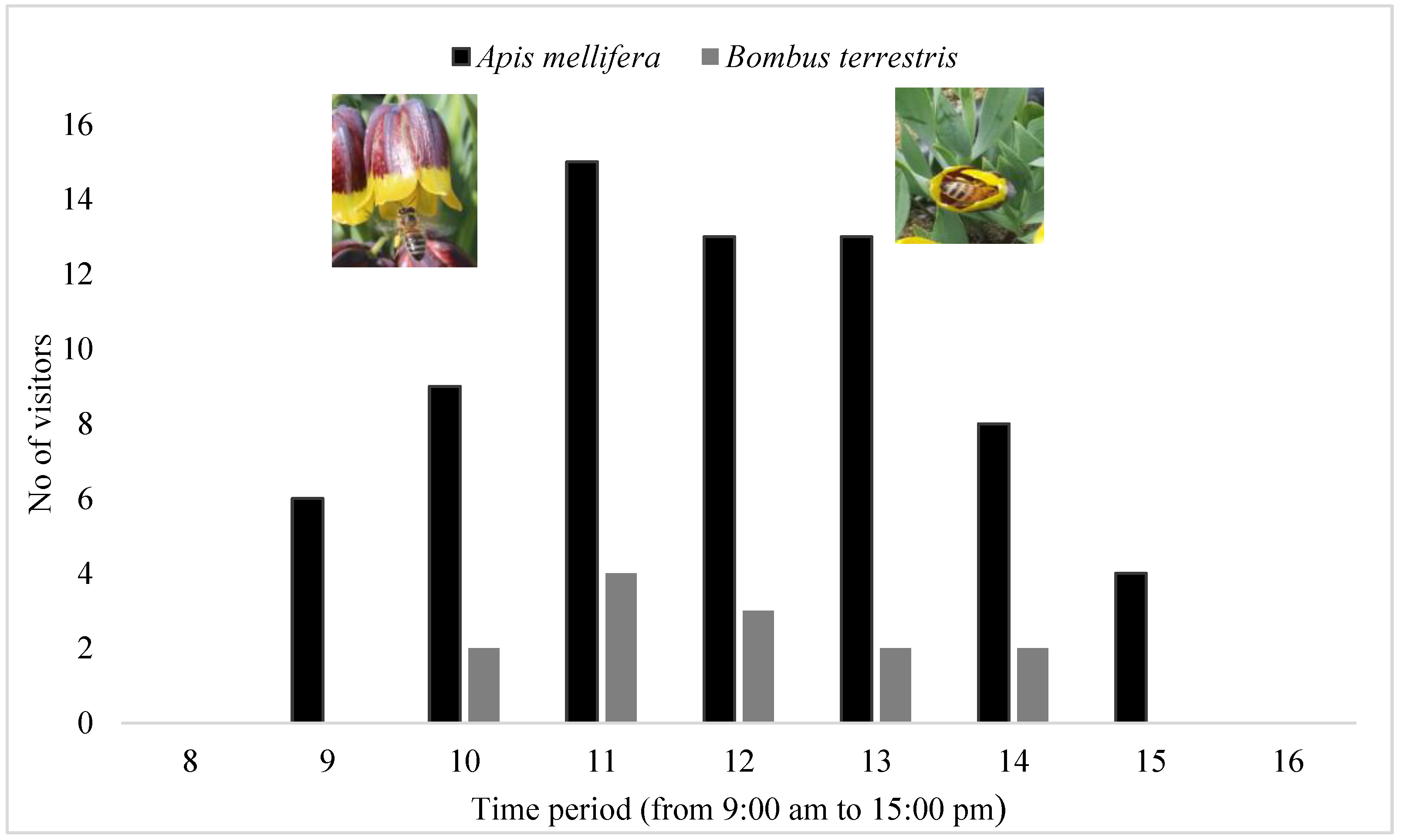
3.7. Pollinator Behavior Observation of F. michailovskyi
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Yıldız, F.; Aslay, M.; Kandemir, A.; Kaya, O. Reproductive Biology of Fritillaria aurea Schott (Liliaceae), a Rare Species Endemic to Turkey. Diversity 2022, 14, 1052. [Google Scholar]
- Gopalakrishnan, K.K.; Thomas, T.D. Reproductive biology of Pittosporum dasycaulon Miq., (Family Pittosporaceae) a rare medicinal tree endemic to Western Ghats. Bot. Stud. 2014, 55, 15. [Google Scholar] [PubMed] [Green Version]
- Li, T.; Liu, X.; Li, Z.; Ma, H.; Wan, Y.; Liu, X.; Fu, L. Study on Reproductive Biology of Rhododendron longipedicellatum: A Newly Discovered and Special Threatened Plant Surviving in Limestone Habitat in Southeast Yunnan, China. Front. Plant Sci. 2018, 9, 33. [Google Scholar]
- Moza, M.K.; Bhatnagar, A.K. Plant reproductive biology studies crucial for conservation. Curr. Sci. 2007, 92, 1207. [Google Scholar]
- Roguz, K.; Bajguz, A.; Gołębiewska, A.; Chmur, M.; Hill, L.; Kalinowski, P.; Zych, M. Functional diversity of nectary structure and nectar composition in the genus Fritillaria (Liliaceae). Front. Plant Sci. 2018, 9, 1246. [Google Scholar]
- Ono, A.; Dohzono, I.; Sugawara, T. Bumblebee pollination and reproductive biology of Rhododendron semibarbatum (Ericaceae). J. Plant Res. 2008, 121, 319–327. [Google Scholar] [PubMed]
- Stout, J.C. Pollination of invasive Rhododendron ponticum (Ericaceae) in Ireland. Apidologie 2007, 38, 198–206. [Google Scholar]
- Kuswantoro, F. Flower-insect visitor interaction: Case study on Rhododendron inundatum Sleumer in Bali Botanic Garden. J. Trop. Biodivers. Biotechnol. 2017, 2, 35–38. [Google Scholar]
- Sutherland, S. Floral sex ratios, fruit set, and resource allocation in plants. Ecology 1986, 67, 991–1001. [Google Scholar] [CrossRef]
- Bawa, K.S.; Webb, C.J. Flower, fruit and seed abortion in tropical forest trees: Implications for the evolution of paternal and maternal reproductive patterns. Am. J. Bot. 1984, 71, 736–751. [Google Scholar]
- Humphreys, A.M.; Govaerts, R.; Ficinski, S.Z.; Nic Lughadha, E.; Vorontsova, M.S. Global dataset shows geography and life form predict modern plant extinction and rediscovery. Nat. Ecol. Evol. 2019, 3, 1043–1047. [Google Scholar]
- Nic Lughadha, E.; Bachman, S.P.; Leão, T.C.; Forest, F.; Halley, J.M.; Moat, J.; Walker, B.E. Extinction risk and threats to plants and fungi. Plants People Planet 2020, 2, 389–408. [Google Scholar]
- Tekşen, M.; Aytaç, Z. The revision of the genus Fritillaria L. (Liliaceae) in the Mediterranean region (Turkey). Turk. J. Bot. 2011, 35, 447–478. [Google Scholar] [CrossRef]
- Hill, L. Fritillaria kiusiana: Liliaceae. Curtis’s Bot. Mag. 2016, 33, 275–285. [Google Scholar] [CrossRef]
- Kiani, M.; Mohammadi, S.; Babaei, A.; Sefidkon, F.; Naghavi, M.R.; Ranjbar, M.; Potter, D. Iran supports a great share of biodiversity and floristic endemism for Fritillaria spp. (Liliaceae): A review. Plant Divers. 2017, 39, 245–262. [Google Scholar] [PubMed]
- Rix, M.; Strange, K. Fritillaria sororum Liliaceae. Curtis’s Bot. Mag 2014, 31, 214–222. [Google Scholar]
- International Union for Conservation of Nature (IUCN). IUCN Red List Categories and Criteria: Version3.1; IUCN Species Survival Commission: Gland, Switzerland, 2001; p. 30. [Google Scholar]
- Demirkuş, N.; Koyuncu, M.; Gül, M. Endemic Plants of Lake Van Basin. Anatol. J. Bot. 2018, 2, 70–83. [Google Scholar]
- Tamura, M.N. Liliaceae. In The Families and Genera of Vascular Plants; Kubitzki, K., Ed.; Springer: Berlin/Heidelberg, Germany; New York, NY, USA, 1998; Volume 3, p. 352. [Google Scholar]
- Zych, M.; Stpiczyńska, M.; Roguz, K. Pollination biology and breeding system of European Fritillaria meleagris L. (Liliaceae). In Reproductive Biology of Plants; CRC Press: Boca Raton, FL, USA, 2014; pp. 147–163. [Google Scholar]
- Rix, E.M. Fritillaria L. In Flora of Turkey and the East Aegean Islands, 1st ed.; Davis, P.H., Ed.; Edinburgh University Press: Edinburgh, UK, 1975; Volume 8, pp. 284–302. [Google Scholar]
- Tekşen, M. Fritillaria L. In Resimli Türkiye Florası; Güner, A., Kandemir, A., Menemen, Y., Yıldırım, H., Aslan, S., Ekşi, G., Güner, I., Ve Çimen, A.Ö., Eds.; Ali Nihat Gökyiğit Vakfı Nezahat Gökyiğit Botanik Bahçesi Yayınları: İstanbul, Turkey, 2018; Volume 2, pp. 800–879. [Google Scholar]
- Yildirim, H.; Tekşen, M. Fritillaria arsusiana (Lilieae, Liliaceae), a new species from southern Anatolia. Phytotaxa 2021, 502, 149–159. [Google Scholar] [CrossRef]
- Eker, I.; Tekşen, M. Fritillaria umitkaplanii (Liliaceae), a new species from south Anatolia. Nord. J. Bot. 2023, 2023, e03803. [Google Scholar]
- Güner, A.; Aslan, S.; Ekim, T.; Vural, M.; Babaç, M.T. (Eds.) Türkiye Bitkileri Listesi: (Damarlı Bitkiler); Nezahat Gökyiǧit Botanik Bahçesi Yayınları: İstanbul, Turkey, 2012; p. s.63. (In Turkish) [Google Scholar]
- Aslay, M.; Çukadar, K.; Ünlü, H.M.; Kadıoglu, Z.; Teksen, M.; Kaya, E. Dogu Anadolu Fritillaria’larının Kültür Sartlarına Adaptasyonu. V. In Proceedings of the Süs Bitkileri Kongresi, Yalova, Turkey, 6–9 May 2013. [Google Scholar]
- Aslay, M.; Ünlü, M.H.; Kadıoğlu, Z.; Tuncer, S.; Kaya, E. Development of New Varıetıes from Turkeys Endemıc Specıes—Fritillaria michailovskyi Fomın. In Researches in Landscape and Ornamental Plants Kitap Bölümü; Gece Kitaplığı: Ankara, Turkey, 2019; pp. 27–49. [Google Scholar]
- Wang, Y.; Aamer, M.; Aslay, M.; Sener, B.; Khan, F.A.; Wahab, A.T.; Choudhary, M.I. A new steroidal alkaloid from Fritillaria michailovskyi Fomin. Nat. Prod. Res. 2021, 36, 361–366. [Google Scholar]
- Dafni, A. Pollination Ecology: A Practical Approach; University Press: New York, NY, USA, 1992. [Google Scholar]
- Dafni, A.; Maues, J. A rapid and simple procedure to determine stigma receptivity. Sex. Plant Reprod. 1998, 11, 177–180. [Google Scholar]
- Shivanna, K.R.; Tandon, R. Reproductive Ecology of Flowering Plants: A Manual; Springer: New Delhi, India, 2014; pp. 107–123. [Google Scholar]
- Cruden, R.W. Pollen-ovule ratios: A conservative indicator of breeding systems in flowering plants. Evolution 1977, 31, 32–46. [Google Scholar]
- Zapata, T.R.; Arroyo, M.T.K. Plant reproductive ecology of a secondary deciduous tropical forest in Venezuela. Biotropica 1978, 10, 221–230. [Google Scholar]
- Aslay, M.; Çukadar, K.; Ünlü, H.M.; Kadıoglu, Z.; Teksen, M. Ekim Öncesi Uygulamalar ve Farklı Çimlenme OrtamlarınınTchihatchewia isatidea Boiss. (Brassicaceae) Tohumlarının Çimlenmesi Üzerine Etkisi. IV. In Proceedings of the Süs Bitkileri Kongresi, Mersin, Turkey, 20–22 October 2010. [Google Scholar]
- Roguz, K.; Hill, L.; Roguz, A.; Zych, M. Evolution of bird and insect flower traits in Fritillaria L. (Liliaceae). Front. Plant Sci. 2021, 12, 656783. [Google Scholar]
- Tekşen, M.; Aytac, Z.; Pinar, N.M. Pollen morphology of the genus Fritillaria L.(Liliaceae) in Turkey. Turk. J. Bot. 2010, 34, 397–416. [Google Scholar]
- Zych, M.; Stpiczyńska, M. Neither protogynous nor obligatory out-crossed: Pollination biology and breeding system of the European Red List Fritillaria meleagris L. (Liliaceae). Plant Biol. 2012, 14, 285–294. [Google Scholar] [PubMed]
- Tong, L.; Lei, F.W.; Wu, Y.M.; Shen, X.L.; Xia, X.F.; Zhang, D.H.; Zhang, Z.X. Multiple reproductive strategies of a spring ephemeral plant, Fritillaria maximowiczii, enable its adaptation to harsh environments. Plant Species Biol. 2022, 37, 38–51. [Google Scholar]
- Zhang, X.; Cui, D.; Zong, X.; Ren, R.; Wei, J.; Situ, L.; Zhang, Y. Pollination biology and breeding system of Fritillaria ussuriensis Maxim. Acta Bot. Boreali-Occident. Sin. 2010, 30, 1404–1408. [Google Scholar]
- Gao, Y.Q.; Zhang, L.X.; Wang, M.R.; Song, B. Pollination biology of Fritillaria delavayi. China J. Chin. Mater. Med. 2014, 39, 1795–1798. [Google Scholar]
- Gao, Y.; Wang, C.; Song, B.; Du, F. Corolla retention after pollination facilitates the development of fertilized ovules in Fritillaria delavayi (Liliaceae). Sci. Rep. 2019, 9, 729. [Google Scholar]
- Chen, Y.; Xu, B.Q.; Guo, F.X.; Bai, G.; Zhang, J.F.; Zhang, Y. Studies of the floral organ characteristics and sexual breeding system of Fritillaria unibracteata. Acta Prataculturae Sin. 2017, 26, 90–98. [Google Scholar]
- Anderson, G.J. Systematics and reproductive biology. In Experimental and Molecular Approaches to Plant Systematics; Monogr Syst Bot 53; Hoch, P.C., Stephenson, A.G., Eds.; Missouri Botanical Garden: St. Louis, MI, USA, 1995; pp. 263–272. [Google Scholar]
- Bernardello, G.; Anderson, G.J.; Lopez, S.P.; Cleland, M.A.; Stuessy, T.F.; Crawford, D.J. Reproductive biology of Lactoris fernandeziana (Lactoridaceae). Am. J. Bot. 1999, 86, 829–840. [Google Scholar] [PubMed]
- Seifert, T.; Muller-Starck, G. Impacts of fructification on biomass production and correlated genetic effects in Norway spruce (Picea abies [L.] Karst.). Eur. J. For. Res. 2009, 128, 155–169. [Google Scholar]
- Yashima, T.; Kinoshita, E.; Shimizu, T. Flowering phenology and self-incompatibility in Fritillaria camtschatcensis (L.) Ker-Gawl. J. Phytogeogr. Taxon 1997, 45, 129–133. [Google Scholar]
- Nyman, Y. Pollination mechanisms in six Campanula species (Campanulaceae). Plant Syst. Evol. 1992, 181, 97–108. [Google Scholar] [CrossRef]
- Barrett, S.C.H.; Cruzan, M.B. Incompatibility in heterostylous plants. In Genetic Control of Self Incompatibility and Reproductive Development in Flowering Plants; Williams, E.G., Clarke, A.E., Knox, R.B., Eds.; Kluwer: Dordrecht, The Netherland, 1994; pp. 189–219. [Google Scholar]
- Brys, R.; Jacquemyn, H.; De Bruyn, L.; Hermy, M. Pollination success and reproductive output in experimental populations of the self-incompatible Primula vulgaris. Int. J. Plant Sci. 2007, 168, 571–578. [Google Scholar]
- Ma, Y.; Cui, Z.; Cheng, C.Y.; Li, R.; Wu, H.; Jin, L.; Wang, Z. Flowering characteristics and mating system of Fritillaria cirrhosa (Liliaceae), an endangered plant in China. Braz. J. Bot. 2022, 45, 1307–1318. [Google Scholar]
- Mancuso, E.; Peruzzi, L. Male individuals in cultivated Fritillaria persica L. (Liliaceae): Real androdioecy or gender disphasy? Turk. J. Bot. 2010, 34, 435–440. [Google Scholar] [CrossRef]
- Kawano, S.; Masuda, J.; Hayashi, K. Life-history monographs of Japanese plants. Fritillaria koidzumiana Ohwi (Liliaceae). Plant Species Biol. 2008, 23, 51–57. [Google Scholar]
- Hedstrom, I. Kungsangsliljans pollinationsekologi. Sven. Bot. Tidskr. 1983, 77, 313–325. [Google Scholar]
- Dafni, A.; Firmage, D. Pollen viability and longevity: Practical, ecological and evolutionary implications. In Pollen and Pollination; Springer: Vienna, Austria, 2000; pp. 113–132. [Google Scholar]
- He, G.; Hu, F.; Ming, J.; Liu, C.; Yuan, S. Pollen viability and stigma receptivity in Lilium during anthesis. Euphytica 2017, 213, 1–10. [Google Scholar]
- Stanley, R.G. Linskens HF Pollen: Biology Biochemistry and Management; Springer: Berlin, Germany, 1974. [Google Scholar]
- Colling, G.; Reckinger, C.; Matthies, D. Effects of pollen quantity and quality on reproduction and offspring vigour in the rare plant Scorzonera humilis (Asteraceae). Am. J. Bot. 2004, 91, 1774–1782. [Google Scholar] [PubMed]
- Rocha, O.J.; Aguilar, G. Reproductive biology of the dry forest tree Enterolobium cyclocarpum (Guanacaste) in Costa Rica: A comparison between trees left in pastures and in continuous forest. Am. J. Bot. 2001, 88, 1607–1614. [Google Scholar] [PubMed]
- Dudash, M.R.; Fenster, C.B. Inbreeding and out breeding depression in fragmented populations. In Genetics, Demography and Viability of Fragmented Populations; Young, A., Clarke, G., Eds.; Cambridge University Press: Cambridge, UK, 2000; pp. 35–53. [Google Scholar]
- Byers, D.L. Pollen quantity and quality as explanations for low seed set in small populations exemplified by Eupatorium (Asteraceae). Am. J. Bot. 2004, 82, 1000–1006. [Google Scholar] [CrossRef]
- Westerkamp, C. Honey bees are poor pollinators—Why? Plant Syst. Evol. 1991, 177, 71–75. [Google Scholar]
- Knuth, P. Handbuch der Blutenbiologie, Band II, Teil 2: Lobeliaceae bis Gnetaceae; Verlag von Wilhelm Engelmann: Leipzig, Germany, 1899. [Google Scholar]
- Rix, E.M.; Rast, D. Nectar sugars and subgeneric classification in Fritillaria. Biochem. Syst. Ecol. 1975, 2, 207–209. [Google Scholar]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Parameters Studied | Observations |
|---|---|
| Flowering period | March–April |
| Inflorescence | Broadly campanulate |
| Flower | Hermaphrodite and actinomorphic |
| Flower color | Purple-stricted yellow |
| Flower length | 16–33 mm |
| Odor | Nil |
| Nectar | Greenish, Oblanceolate |
| Anther dehiscence | Bursting inwards by slits |
| Number of anthers/flower | 6 |
| The average number of pollen grains/flower | 252,000 |
| The average number of ovules/ovary | 144 |
| Length of tepal | 18–36 mm |
| Length of filament | 6–10 mm |
| Length of anther | 5–11 mm |
| Length of stamen | 8–14 mm |
| Length of style | 6–10 mm |
| Pollen-ovule ratio | 1750 |
| Pollen surface | Granulate |
| Pollen shape | Subprolate |
| Pollen cell | 2 celled |
| Pollen size | 31–66 μm |
| Pollen viability | 95% |
| Stigma shape | Trifid or trilobate |
| Stigma surface | Wet |
| Diameter of ovary | 1.8–2.3 mm |
| Ovary placentation type | Axile |
| Ovary status | Hypogin |
| Ovary capsule | 6-lobed |
| Stamen type | Singenesis stamen |
| Periant type | Perigon (Tepal) |
| Corolla type | Gamosepal |
| Anther type | Basifix |
| Perianth estivation | Imbricate-alternate |
| Anther dehiscence direction | Extrorse |
| Treatment | Flowers | Fruits Developed (%) | Fruit Set (%) | Seed Viability (%) | Germination (%) |
|---|---|---|---|---|---|
| Open-pollination | 50 | 33 (66) | 68 | 63 | 43 |
| Spontaneous self-pollination | 50 | 0 | - | - | - |
| Spontaneous cross-pollination | 50 | 41 (82) | 60 | 64 | 45 |
| Induced self-pollination | 50 | 0 | - | - | - |
| Xenogamous pollination | 50 | 43 (86) | 59 | 74 | 51 |
| Apomixis | 30 | 0 | - | - | - |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Aslay, M.; Yıldız, F.; Kaya, O.; Bita-Nicolae, C. Reproductive Biology and Pollination Ecology of Fritillaria michailovskyi Fomin (Liliaceae), Endemic to East Anatolia (Turkey). Diversity 2023, 15, 414. https://doi.org/10.3390/d15030414
Aslay M, Yıldız F, Kaya O, Bita-Nicolae C. Reproductive Biology and Pollination Ecology of Fritillaria michailovskyi Fomin (Liliaceae), Endemic to East Anatolia (Turkey). Diversity. 2023; 15(3):414. https://doi.org/10.3390/d15030414
Chicago/Turabian StyleAslay, Meral, Faruk Yıldız, Ozkan Kaya, and Claudia Bita-Nicolae. 2023. "Reproductive Biology and Pollination Ecology of Fritillaria michailovskyi Fomin (Liliaceae), Endemic to East Anatolia (Turkey)" Diversity 15, no. 3: 414. https://doi.org/10.3390/d15030414