
Hybridization in the Fringed Orchids: An Analysis of Species Boundaries in the Face of Gene Flow

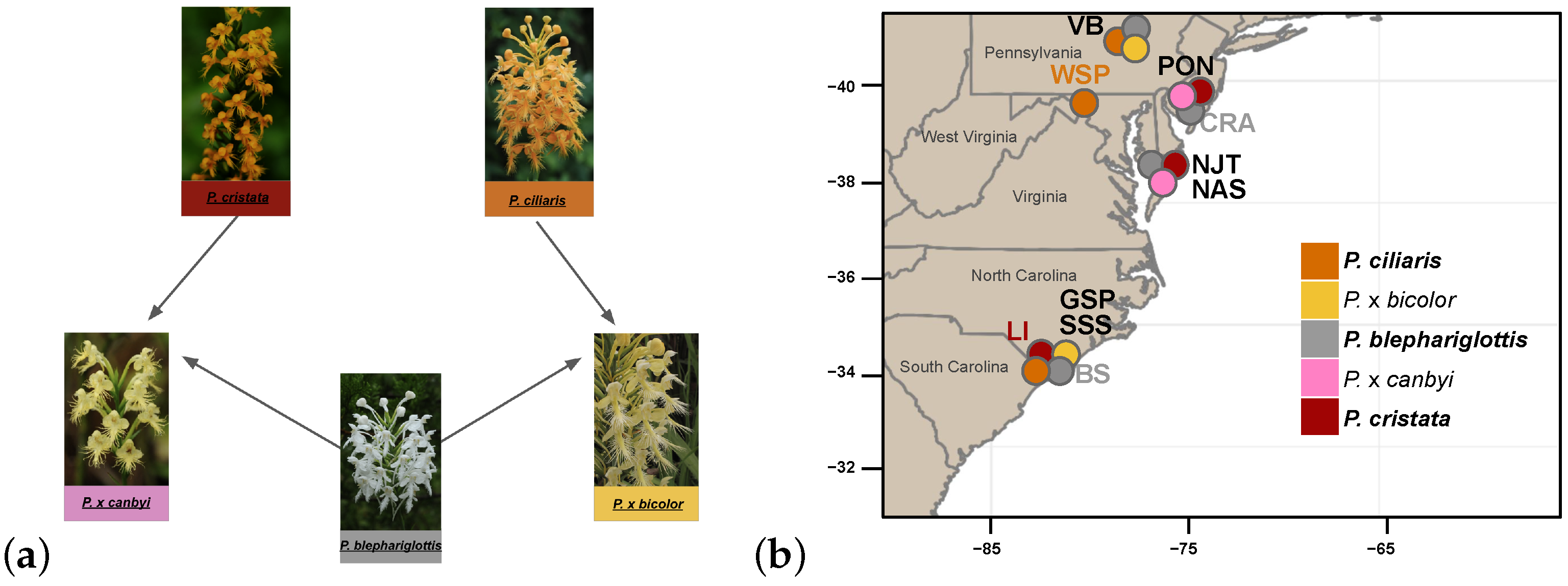{kind=link}
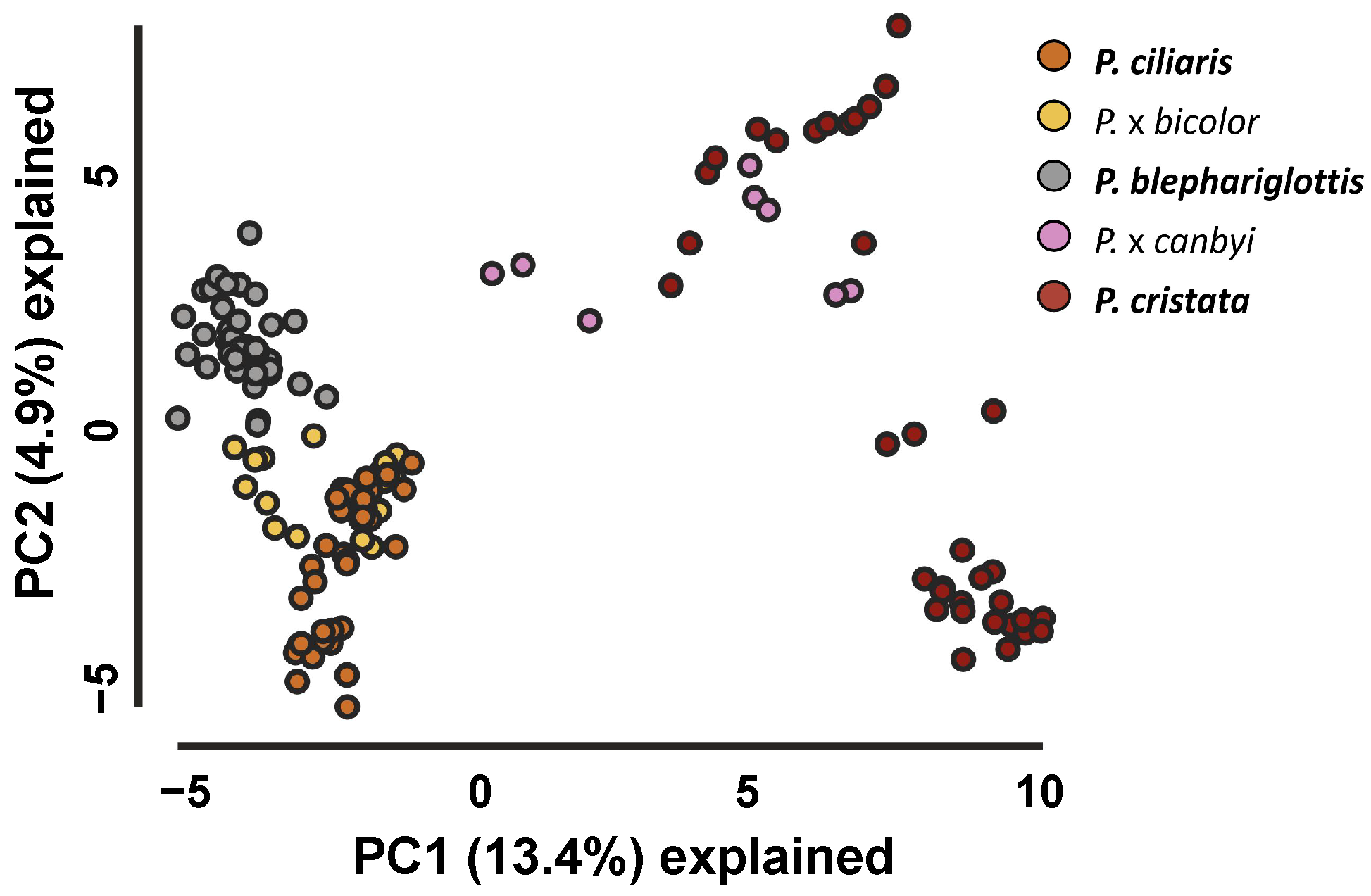{kind=link}
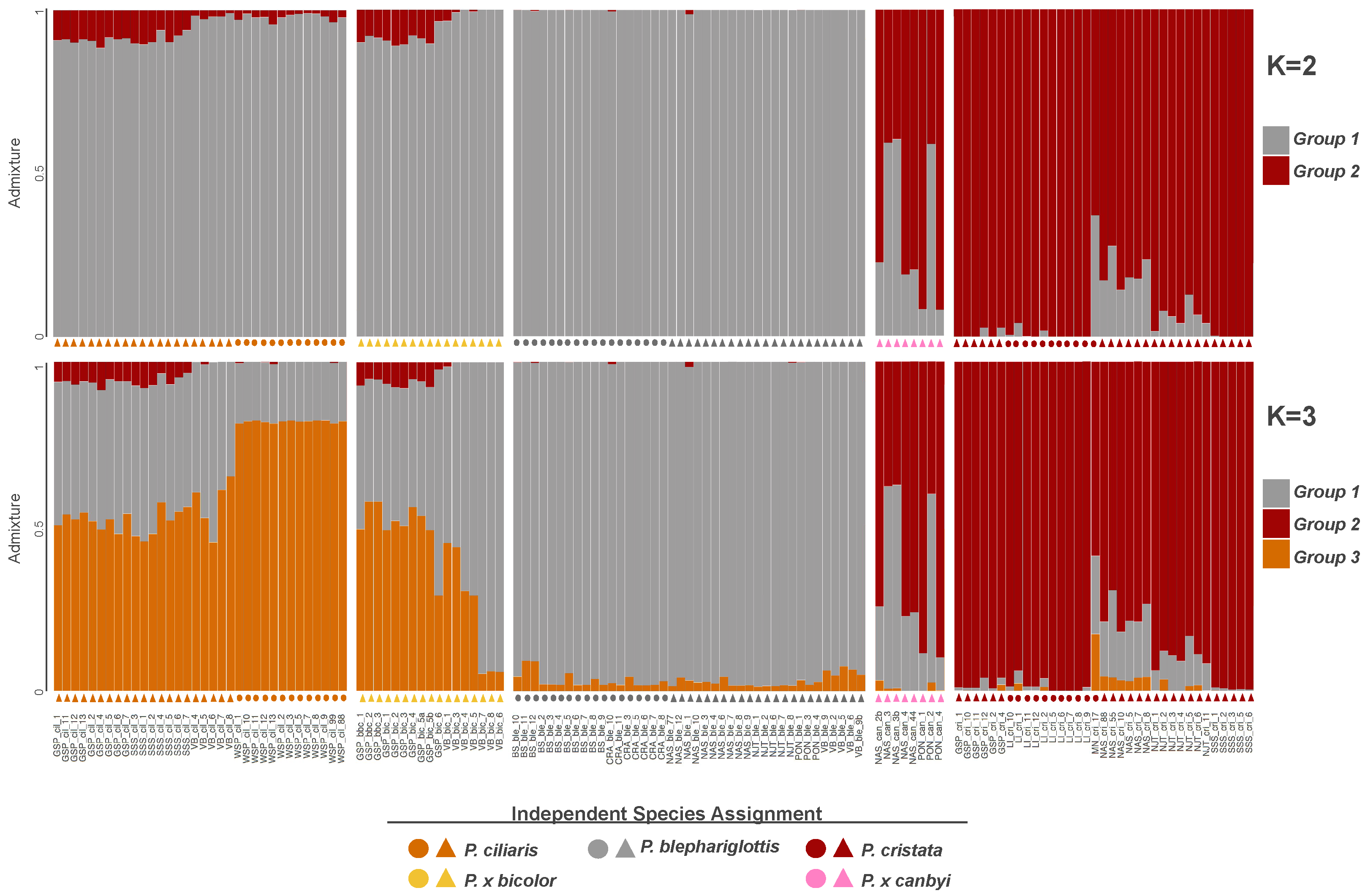{kind=link}
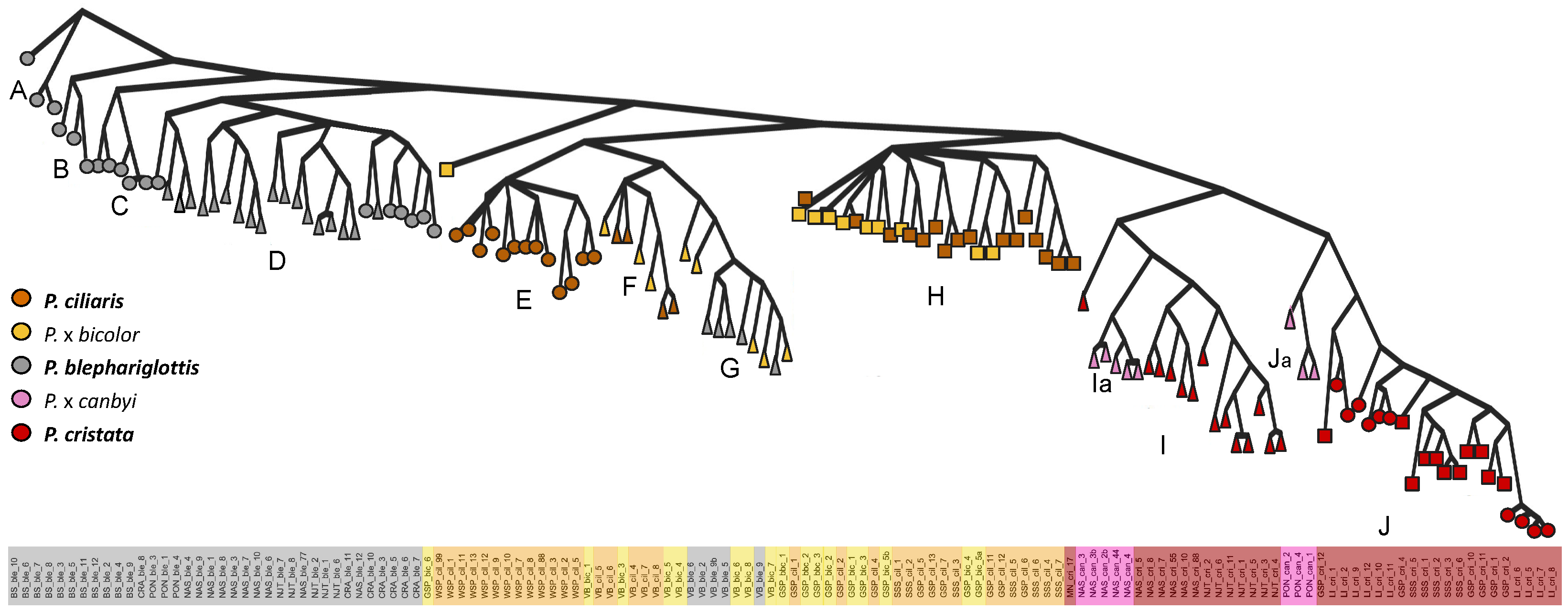{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Hybridization and Backcrossing in the Fringed Orchids: Morphology
2.2. Hybridization and Backcrossing in the Fringed Orchids: Genomics
2.3. Observed Hybridization Types
2.4. Morphological versus Genetic Composition
3. Results
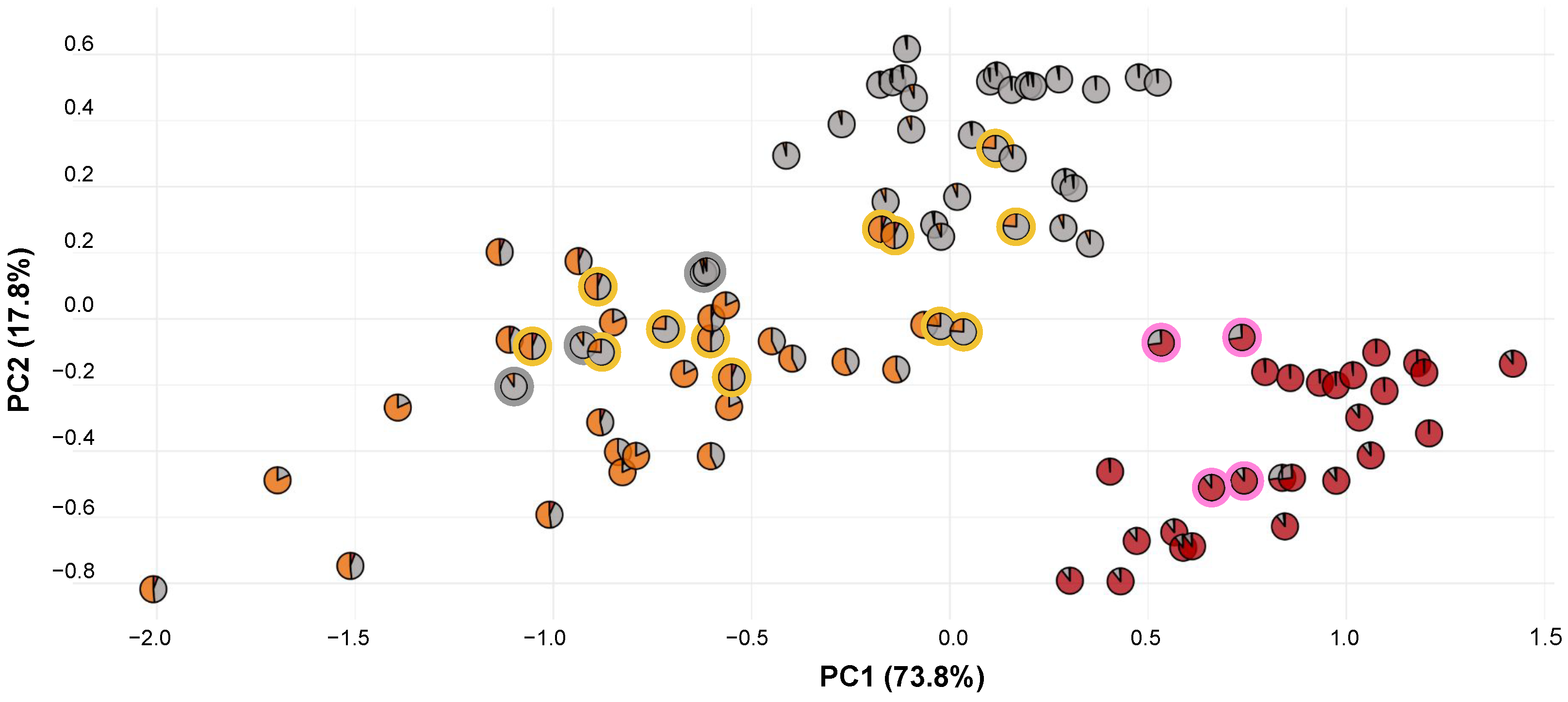
3.1. Morphology Analysis
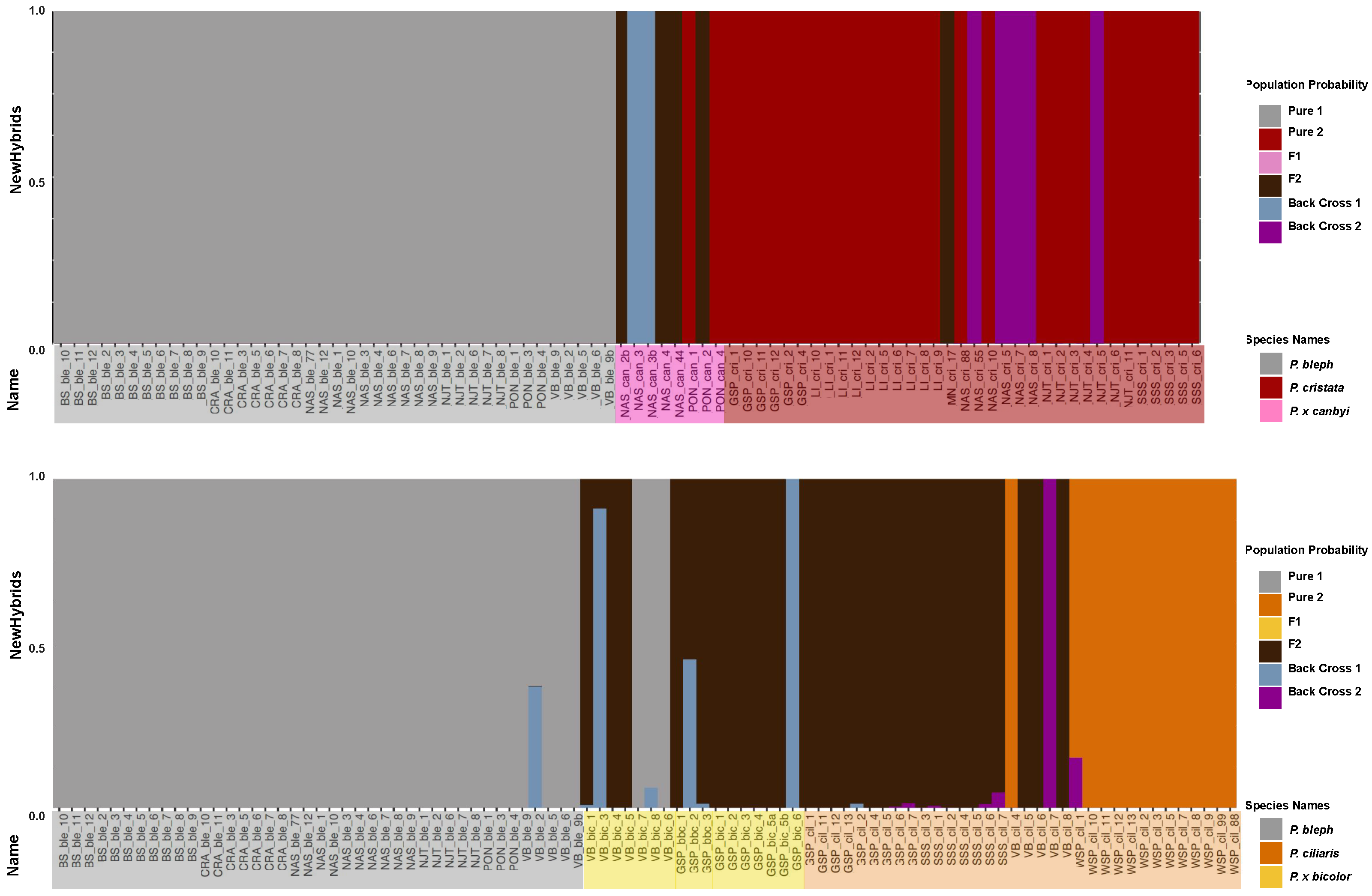
3.2. Genomic Analysis
4. Discussion
4.1. Hybridization in Allopatric and Sympatric Sites
4.2. Morphology versus Genetic Composition
4.3. Species Boundaries and Conservation Models
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
| RAD | Restriction site Associated DNA Sequencing |
| SNP | Single Nucleotide Polymorphism |
| F1 | First filial generation |
| F2 | Second and later filial generations |
| ble | Platanthera blephariglottis |
| cri | Platanthera cristata |
| cil | Platanthera ciliaris |
| bic | Platanthera x bicolor |
| bbc | Platanthera x bicolor |
| can | Platanthera x canbyi |
References
- Givnish, T.J.; Spalink, D.; Ames, M.; Lyon, S.P.; Hunter, S.J.; Zuluaga, A.; Iles, W.J.; Clements, M.A.; Arroyo, M.T.; Leebens-Mack, J.; et al. Orchid Phylogenomics and multiple drivers of their extraordinary diversification. Proc. R. Soc. B 2015, 282, 20151553. [Google Scholar] [CrossRef]
- Hassler, M. World Orchids. Synonymic Checklist and Distribution of the Orchids of the World. Version 14.5. Available online: www.worldplants.de/orchids/ (accessed on 4 December 2022).
- Harrison, R.G.; Larson, E.L. Hybridization, introgression, and the nature of Species Boundaries. J. Hered. 2014, 105, 795–809. [Google Scholar] [CrossRef] [PubMed]
- Mayr, E. Systematics and the Origin of Species; Columbia University Press: New York City, NY, USA, 1942. [Google Scholar]
- Queiroz, K. Ernst Mayr and the modern concept of species. Proc. Natl. Acad. Sci. USA 2005, 102, 6600–6607. [Google Scholar] [CrossRef] [PubMed]
- Arida, B.L.; Scopece, G.; Machado, R.M.; Moraes, A.P.; Forni-Martins, E.; Pinheiro, F. Reproductive barriers and fertility of two neotropical orchid species and their natural hybrid. Evol. Ecol. 2021, 35, 41–64. [Google Scholar] [CrossRef]
- Andriamihaja, C.F.; Ramarosandratana, A.V.; Grisoni, M.; Jeannoda, V.; Besse, P. The Leafless Vanilla Species-Complex from the South-West Indian Ocean Region: A Taxonomic Puzzle and a Model for Orchid Evolution and Conservation Research. Diversity 2020, 12, 443. [Google Scholar] [CrossRef]
- Cozzolino, S.; D’Emerico, S.; Widmer, A. Evidence for reproductive isolate selection in Mediterranean orchids: Karyotype differences compensate for the lack of pollinator specificity. Biol. Lett. 2004, 271, 259–262. [Google Scholar] [CrossRef]
- Dafni, A.; Ivri, Y. Pollination ecology of, and hybridization between, Orchis coriophora L. and Orchis collina Sol. ex Russ. (Orchidaceae) in Israel. New Phytol. 1979, 83, 181–187. [Google Scholar] [CrossRef]
- Esposito, F.; Vereecken, N.J.; Gammella, M.; Rinaldi, R.; Laurent, P.; Tyteca, D. Characterization of sympatric Platanthera bifolia and Platanthera chlorantha (Orchidaceae) populations with intermediate plants. PeerJ 2018, 6, 4256. [Google Scholar] [CrossRef]
- Vereecken, N.J.; Dafni, A.; Cozzolino, S. Pollination syndromes in Mediterranean orchids—implications for speciation, taxonomy and conservation. Bot. Rev. 2010, 76, 220–240. [Google Scholar] [CrossRef]
- Stull, G.W.; Pham, K.K.; Soltis, P.S.; Soltis, D.E. Deep reticulation: The long legacy of hybridization in vascular plant evolution. Plant J. 2023. Accepted Author Manuscript. [Google Scholar] [CrossRef]
- Fay, M.F. How can we meet the challenges in the twenty-first century? Bot Stud. 2018, 59, 16. [Google Scholar] [CrossRef] [PubMed]
- Krupnick, G.A.; McCormick, M.K.; Mirenda, T.; Whigham, D.F. The status and future of Orchid Conservation in North America. Ann. Mo. Bot. Gard. 2013, 99, 180–198. [Google Scholar] [CrossRef]
- Li, T.; Wu, S.; Yang, W.; Selosse, M.; Goa, J. How Mycorrhizal Associations Influence Orchid Distribution and Population Dynamics. Front. Plant Sci. 2021, 12, 647114. [Google Scholar] [CrossRef] [PubMed]
- Wallace, L.E. Biological Investigations In the Genus Platanthera (Orchidaceae): Conservation Issues in Platanthera Leucophaea Furthermore, Evolutionary Diversification in Section Limnorchis. Ph.D. Thesis, School of the Ohio State University, Columbus, OH, USA, 2002. Available online: https://etd.ohiolink.edu/apexprod/rws_olink/r/1501/10?clear=10&p10_accession_num=osu1038833679 (accessed on 20 February 2023).
- Gompert, Z.; Mandeville, E.G.; Buerkle, C.A. Analysis of population genomic data from hybrid zones. Annual Review of Ecology, Evolution, and Systematics. Annu. Rev. Ecol. Evol. Syst. 2017, 48, 207–209. [Google Scholar] [CrossRef]
- Knief, U.; Bossu, C.M.; Saino, N.; Hansson, B.; Poelstra, J.; Vijay, N.; Weissensteiner, M.; Wolf, J.B. Epistatic mutations under divergent selection govern phenotypic variation in the Crow Hybrid Zone. Nat. Ecol. Evol. 2019, 3, 570–576. [Google Scholar] [CrossRef]
- Runemark, A.; Vallejo-Marin, M.; Meier, J.I. Eukaryote hybrid genomes. PLoS Genet. 2019, 15, e1008404. [Google Scholar] [CrossRef]
- Taylor, S.A.; Larson, E.L. Insights from genomes into the evolutionary importance and prevalence of hybridization in nature. Nat. Ecol. Evol. 2019, 3, 170–177. [Google Scholar] [CrossRef]
- Vilà, M.; Weber, E.; D’Antonio, C.M. Conservation implications of invasion by plant hybridization. Biol. Invasions. 2000, 2, 207–217. [Google Scholar] [CrossRef]
- Swarts, N.D.; Dixon, K.W. Terrestrial orchid conservation in the age of extinction. Ann. Bot. 2009, 104, 543–556. [Google Scholar] [CrossRef]
- Stoutamire, W.P. Relationships of the purple-fringed orchids Platanthera psycodes and P. grandiflora. Brittonia 1974, 26, 42–58. [Google Scholar] [CrossRef]
- Brown, P.M. Understanding Platanthera chapmanii (Orchidaceae), its origins and hybrids. SIDA Contrib. Bot. 2004, 21, 853–859. [Google Scholar]
- Wallace, L.E. Spatial genetic structure and frequency of interspecific hybridization in Platanthera aquilonis and P. dilatata (Orchidaceae) occurring in sympatry. Am. J. Bot. 2006, 93, 1001–1009. [Google Scholar] [CrossRef]
- Wettewa, E.; Bailey, N.; Wallace, L.E. Comparative analysis of genetic and morphological variation within the Platanthera hyperborea complex (Orchidaceae). Syst. Bot. 2020, 45, 767–778. [Google Scholar] [CrossRef]
- Richard, L.M. De Orchideis Europaeis Annotationes; Ex typ. A. Belin: Paris, France, 1817. [Google Scholar]
- Bateman, R.M.; James, K.E.; Rudall, P.J. Contrast in levels of morphological versus molecular divergence between closely related Eurasian species of Platanthera (orchidaceae) suggests recent evolution with a strong allometric component. New J. Bot. 2012, 2, 110–148. [Google Scholar] [CrossRef]
- Hulten, E.; Fries, M. Atlas of North European Vascular Plants North of the Tropic of Cancer; Koeltz Scientific Books: Schmitten, Germany, 1986. [Google Scholar]
- Wettewa, E.; Wallace, L.E. Molecular phylogeny and ancestral biogeographic reconstruction of Platanthera subgenus Limnorchis (Orchidaceae) using target capture methods. Mol. Phylogenet. Evol. 2021, 157, 107070. [Google Scholar] [CrossRef] [PubMed]
- Efimov, P.G. A Revision of Platanthera (Orchi-daceae; Orchidoideae; Orchideae) in Asia. Phytotaxa 2016, 254, 1–233. [Google Scholar] [CrossRef]
- Hartvig, I.; Evans, S.; Ligon, J.; Eserman, L.; Coffey, E.; McCormick, M. Quantitative morphological variation in the orange and white fringed orchids (Platanthera) in the eastern US. American Orchid Society. Lindleyana 2022, 532–539. Available online: https://static-curis.ku.dk/portal/files/326038574/Hartvig_et_al_2022_Quantitative_morphology_Platanthera.pdf (accessed on 20 February 2023).
- Luer, C.A. The Native Orchids of the United States and Canada Excluding Florida; The New York Botanical Garden: Bronx, NY, USA, 1975. [Google Scholar]
- Argue, C.L. The Pollination Biology of North American Orchids; Springer: New York City, NY, USA, 2012. [Google Scholar]
- The PLANTS Database. United States Department of Agriculture, Natural Resources Conservation Service. Available online: http://plants.usda.gov (accessed on 20 February 2023).
- Pace, M.C. The Orchidaceae of northeastern North America: Systematics, evolution, diversity, and conservation. J. Torrey Bot. 2020, 29, 156–189. [Google Scholar]
- Cole, F.R.; Firmage, D.H. The Floral Ecology of Platanthera-Blephariglottis. Am. J. Bot. 1984, 71, 700–710. [Google Scholar] [CrossRef]
- Hapeman, J.R.; Inoue, K. Floral Diversification, Pollination Biology, and Molecular Evolution in Platanthera (Orchidaceae); Cambridge University Press: Cambridge, UK, 1997; pp. 433–454. [Google Scholar]
- Robertson, J.L.; Wyatt, R. Evidence for Pollination Ecotypes in the Yellow-Fringed Orchid, Platanthera ciliaris. Evolution 1990, 44, 121–133. [Google Scholar] [CrossRef]
- Smith, G.; Snow, G. Pollination Ecology of Platanthera (Habenaria) ciliaris and P. blephariglottis(Orchidaceae). Bot. Gaz. 1976, 137, 133–140. [Google Scholar] [CrossRef]
- Zale, P.J.; McCormick, M.K.; Whigham, D.F. Choosing a Favorable Substrate to Cultivate Native Orchids Symbiotically: Examples Using Goodyera tesselata and Platanthera blephariglottis. HortScience 1976, 57, 634–642. [Google Scholar] [CrossRef]
- Zettler, L.W.; Dvorak, C.J. Tulasnella calospora (UAMH 9824) retains its effectiveness at facilitating orchid symbiotic germination in vitro after two decades of subculturing. Bot Stud. 2021, 62, 34580790. [Google Scholar] [CrossRef] [PubMed]
- Zettler, L.W.; Hofer, C.J. Propagation of the little club-spur orchid (Platanthera clavellata) by symbiotic seed germination and its ecological implications. Environ. Exp. Bot. 1998, 39, 189–195. [Google Scholar] [CrossRef]
- Cozzolino, S.; Scopece, G.; Roma, L.; Schlüter, P.M. Different filtering strategies of genotyping by sequencing data provide complementary resolutions of species boundaries and relationships in a clade of sexually deceptive orchids. J. Syst. Evol. 2019, 52, 133–144. [Google Scholar] [CrossRef]
- Knowles, L.L.; Maddison, W.P. Statistical Phylogeography. Mol. Ecol. 2002, 11, 2623–2635. [Google Scholar] [CrossRef] [PubMed]
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2002; Available online: https://www.R-project.org/ (accessed on 20 February 2023).
- Koutecký, P. MorphoTools: A set of R functions for morphometric analysis. Plant Syst. Evol. 2015, 301, 1115–1121. [Google Scholar] [CrossRef]
- Oksanen, J.; Blanchet, F.; Friendly, M.; Kindt, R.; Legendre, P.; McGlinn, D.; Minchin, P.R.; O’Hara, R.B.; Simpson, G.L.; Solymos, P.; et al. Vegan: Community Ecology Package. R Package Version 2.2-1. 2. 1-2. 2015. Available online: https://cran.r-project.org/web/packages/vegan/index.html (accessed on 20 February 2023).
- Bayona-Vásquez, N.J.; Glenn, T.C.; Kieran, T.J.; Pierson, T.W.; Hoffberg, S.L.; Scott, P.A.; Bentley, K.E.; Finger, J.W.; Louha, S.; Troendle, N.; et al. Adapterama III: Quadruple-indexed, double/triple-enzyme RADseq libraries (2RAD/3RAD). PeerJ 2019, 7, e7724. [Google Scholar] [CrossRef] [PubMed]
- Eaton, D.A. Pyrad: Assembly of de novo RADseq loci for phylogenetic analyses. Bioinformatics 2014, 30, 1844–1849. [Google Scholar] [CrossRef]
- Eaton, D.A.; Overcast, I. Ipyrad: Interactive Assembly and analysis of RADseq datasets. Bioinformatics 2020, 36, 2592–2594. [Google Scholar] [CrossRef]
- Yu, G. SI/HPC: Smithsonian Institution High Performance Computing Cluster. Available online: https://researchcomputing.si.edu/high-performance-computing-cluster (accessed on 20 August 2022).
- McCartney-Melstad, E.; Gidiş, M.; Shaffer, H.B. An empirical pipeline for choosing the optimal clustering threshold in RADseq studies. Mol. Ecol. Resour. 2019, 19, 1195–1204. [Google Scholar] [CrossRef]
- Hodel, R.G.; Massatti, R.; Bishop, S.G.; Knowles, L.L. Testing which axes of species differentiation underlie covariance of phylogeographic similarity among montane sedge species. Evolution 2021, 75, 349–364. [Google Scholar] [CrossRef] [PubMed]
- Ronquist, F.; Huelsenbeck, J.P. MrBayes 3: Bayesian phylogenetic inference under Mixed Models. Bioinformatics 2003, 19, 1572–1574. [Google Scholar] [CrossRef] [PubMed]
- Eaton, D.A. Toytree: A minimalist tree visualization and manipulation library for Python. Methods Ecol. Evol. 2019, 11, 187–191. [Google Scholar] [CrossRef]
- Pritchard, J.K.; Stephens, M.; Donnelly, P. Inference of population structure using multilocus genotype data. Genetics 2000, 155, 945–959. [Google Scholar] [CrossRef]
- Evanno, G.; Regnaut, S.; Goudet, J. Detecting the number of clusters of individuals using the software structure: A simulation study. Mol. Ecol. 2005, 14, 2611–2620. [Google Scholar] [CrossRef]
- Gao, Y.; Harris, A.J.; Li, H.; Gao, X. Hybrid speciation and introgression both underlie the genetic structures and evolutionary relationships of three morphologically distinct species of lilium (Liliaceae) forming a hybrid zone along an elevational gradient. Front. Plant Sci. 2020, 11, 576407. [Google Scholar] [CrossRef]
- Jakobsson, M.; Rosenberg, N.A. CLUMPP: A cluster matching and permutation program for dealing with label switching and multimodality in analysis of population structure. Bioinformatics 2007, 23, 1801–1806. [Google Scholar] [CrossRef]
- Anderson, E.C.; Thompson, E.A. A model-based method for identifying species hybrids using multilocus genetic data. Genetics 2002, 160, 1217–1229. [Google Scholar] [CrossRef]
- Gramlich, S.; Wagner, N.D.; Hörandl, E. Rad-seq reveals genetic structure of the F2-generation of natural willow hybrids (Salix L.) and a great potential for interspecific introgression. BMC Plant Biol. 2018, 6, 7645–7655. [Google Scholar] [CrossRef]
- Hench, K.; Helmkampf, M.; McMillan, W.O.; Puebla, O. Rapid radiation in a highly diverse marine environment. Proc. Natl. Acad. Sci. USA 2022, 119, e2020457119. [Google Scholar] [CrossRef]
- Lischer, H.E.; Excoffier, L. PGDSpider: An automated data conversion tool for connecting population genetics and genomics programs. Bioinformatics 2012, 28, 298–299. [Google Scholar] [CrossRef]
- Anderson, E.C. User’s Guide to the Program NewHybrids, Version 1.1 beta. Technical Report. Available online: http://ib.berkeley.edu/labs/slatkin/eriq/software/new_hybs_doc1_1Beta3.pdf (accessed on 7 April 2022).
- Wringe, B.F.; Stanley, R.R.; Jeffery, N.W.; Anderson, E.C.; Bradbury, I.R. hybriddetective: A workflow and package to facilitate the detection of hybridization using genomic data in r. Mol. Ecol. Resour. 2017, 17, e275–e284. [Google Scholar] [CrossRef] [PubMed]
- Yu, G. scatterpie: Scatter Pie Plot. R Package Version 0.1.8; 2022. Available online: https://cran.r-project.org/web/packages/scatterpie/index.html (accessed on 3 April 2022).
- Janes, J.K.; Miller, J.M.; Dupuis, J.R.; Malefant, R.M.; Gorrell, J.C.; Cullingham, C.I.; Andrew, R.L. The K = 2 conundrum. Mol Ecol. 2017, 26, 3594–3602. [Google Scholar] [CrossRef] [PubMed]
- Cozzolino, S.; Nardella, A.M.; Impagliazzo, S.; Widmer, A.; Lexer, C. Hybridization and conservation of Mediterranean Orchids: Should we protect the orchid hybrids or the orchid hybrid zones? Biol. Conserv. 2006, 191, 14–23. [Google Scholar] [CrossRef]
- Bersweden, L.; Viruel, J.; Schatz, B.; Harland, J.; Gargiulo, R.; Cowan, R.S.; Calevo, J.; Juan, A.; Clarkson, J.J.; Leitch, A.R.; et al. Microsatellites and petal morphology reveal new patterns of admixture in Orchis hybrid zones. Am. J. Bot. 2021, 108, 1388–1404. [Google Scholar] [CrossRef]
- Lowry, D.B.; Modliszewski, J.L.; Wright, K.M.; Wu, C.A.; Willis, J.H. The strength and genetic basis of reproductive isolating barriers in flowering plants. Proc. R. Soc. B 2008, 363, 3009–3021. [Google Scholar] [CrossRef]
- Scopece, G.; Croce, A.; Lexer, C.; Cozzolino, S. Components of reproductive isolation between Orchis mascula and Orchis pauciflora. Evolution 2013, 67, 2083–2093. [Google Scholar] [CrossRef]
- Quilodrán, C.S.; Austerlitz, F.; Currat, M.; Montoya-Burgos, J.I. Cryptic biological invasions: A general model of hybridization. Sci. Rep. 2018, 8, 2414. [Google Scholar] [CrossRef]
- Hübner, S.; Bercovich, N.; Todesco, M.; Mandel, J.R.; Odenheimer, J.; Ziegler, E.; Lee, J.D.; Baute, G.J.; Owens, G.L.; Grass, C.J.; et al. Sunflower pan-genome analysis shows that hybridization altered gene content and disease resistance. Nat. Plants 2019, 5, 54–62. [Google Scholar] [CrossRef]
- Palaz, E.B.; Demirel, F.; Adali, S.; Demirel, S.; Yilmaz, A. Genetic relationships of salep orchid species and gene flow among Serapias vomeracea × Anacamptis morio hybrids. Plant Biotechnol. Rep. 2022, 16, 1863–5466. [Google Scholar] [CrossRef]
- Rice, A.; Glick, L.; Abadi, S.; Einhorn, M.; Kopelman, N.M.; Salman-Minkov, A.; Mayzel, J.; Chay, O.; Mayrose, I. The Chromosome Counts Database (CCDB)—A community resource of plant chromosome numbers. New Phytol. 2015, 206, 19–26. [Google Scholar] [CrossRef] [PubMed]
- Hollick, P.S.; Taylor, R.J.; McComb, J.A.; Dixon, K.W. If Orchid Mycorrhizal Fungi Are So Specific, How Do Natural Hybrids Cope? Selbyana 2005, 26, 159–170. [Google Scholar] [CrossRef]






Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Evans, S.A.; Whigham, D.F.; Hartvig, I.; McCormick, M.K. Hybridization in the Fringed Orchids: An Analysis of Species Boundaries in the Face of Gene Flow. Diversity 2023, 15, 384. https://doi.org/10.3390/d15030384
Evans SA, Whigham DF, Hartvig I, McCormick MK. Hybridization in the Fringed Orchids: An Analysis of Species Boundaries in the Face of Gene Flow. Diversity. 2023; 15(3):384. https://doi.org/10.3390/d15030384
Chicago/Turabian StyleEvans, Simone A., Dennis F. Whigham, Ida Hartvig, and Melissa K. McCormick. 2023. "Hybridization in the Fringed Orchids: An Analysis of Species Boundaries in the Face of Gene Flow" Diversity 15, no. 3: 384. https://doi.org/10.3390/d15030384