
Under-Ice Development of Silica-Scaled Chrysophytes with Different Trophic Mode in Two Ultraoligotrophic Lakes of Yakutia
 , , , and
, , , and
Abstract
:1. Introduction
2. Materials and Methods
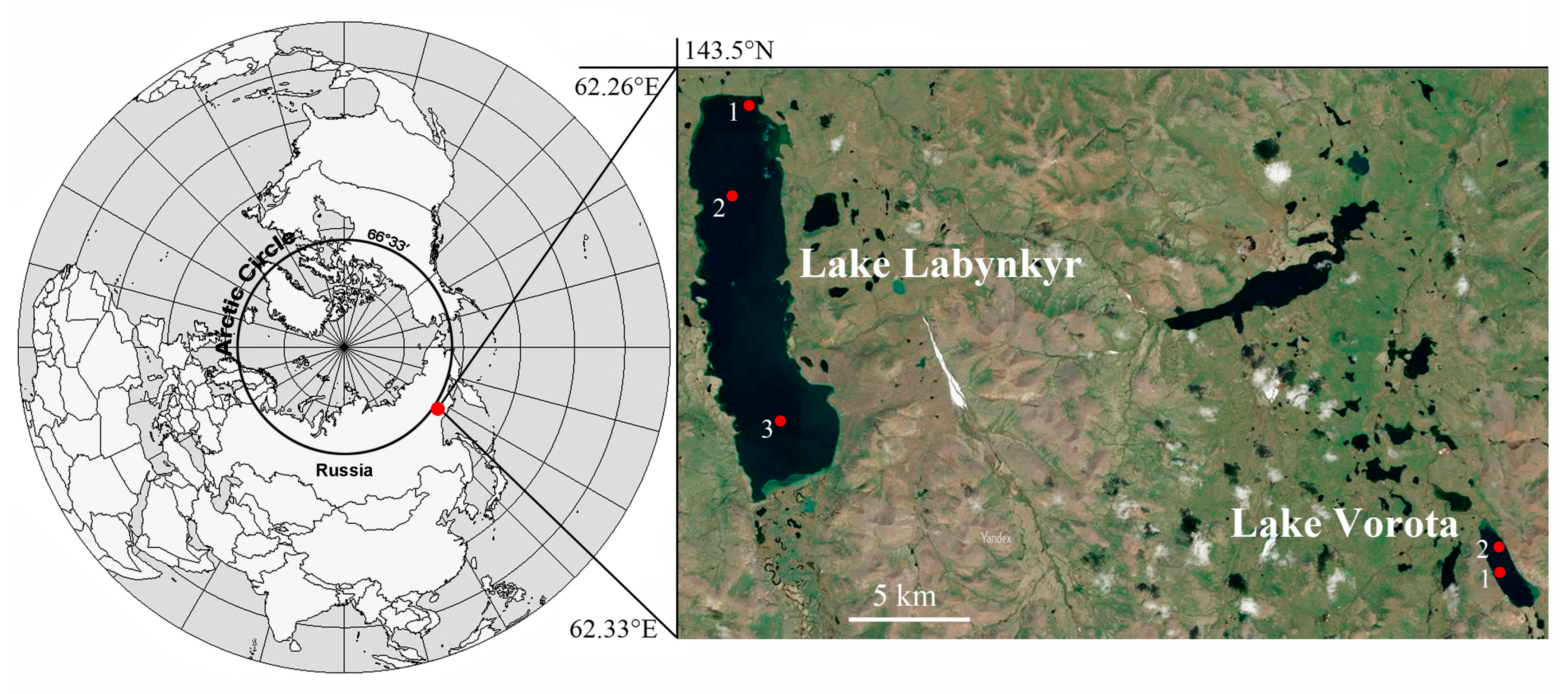
2.1. Study Site
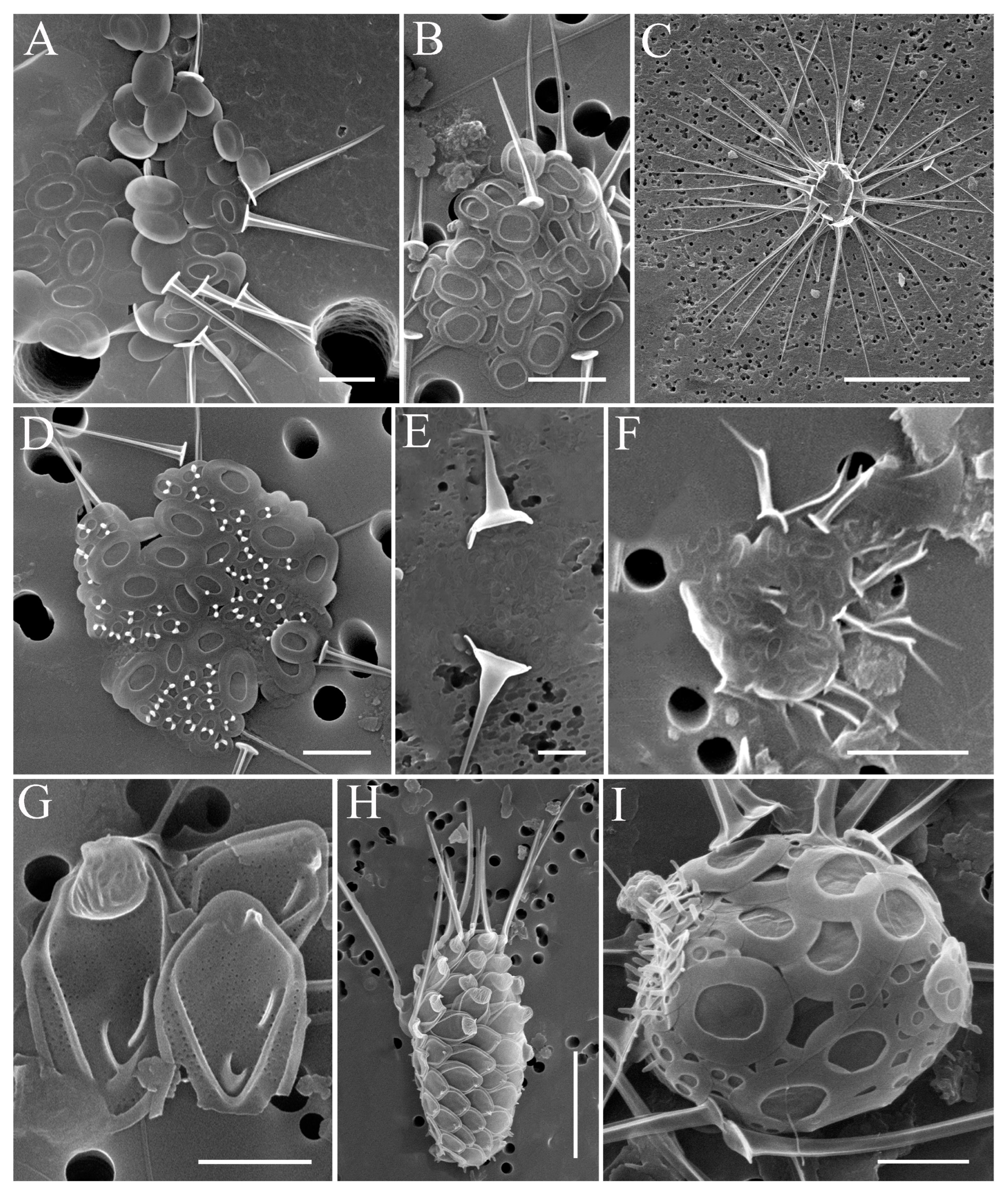
2.2. Study of Silica-Scaled Chrysophytes
2.3. Statistical Analysis and Visualization
3. Results
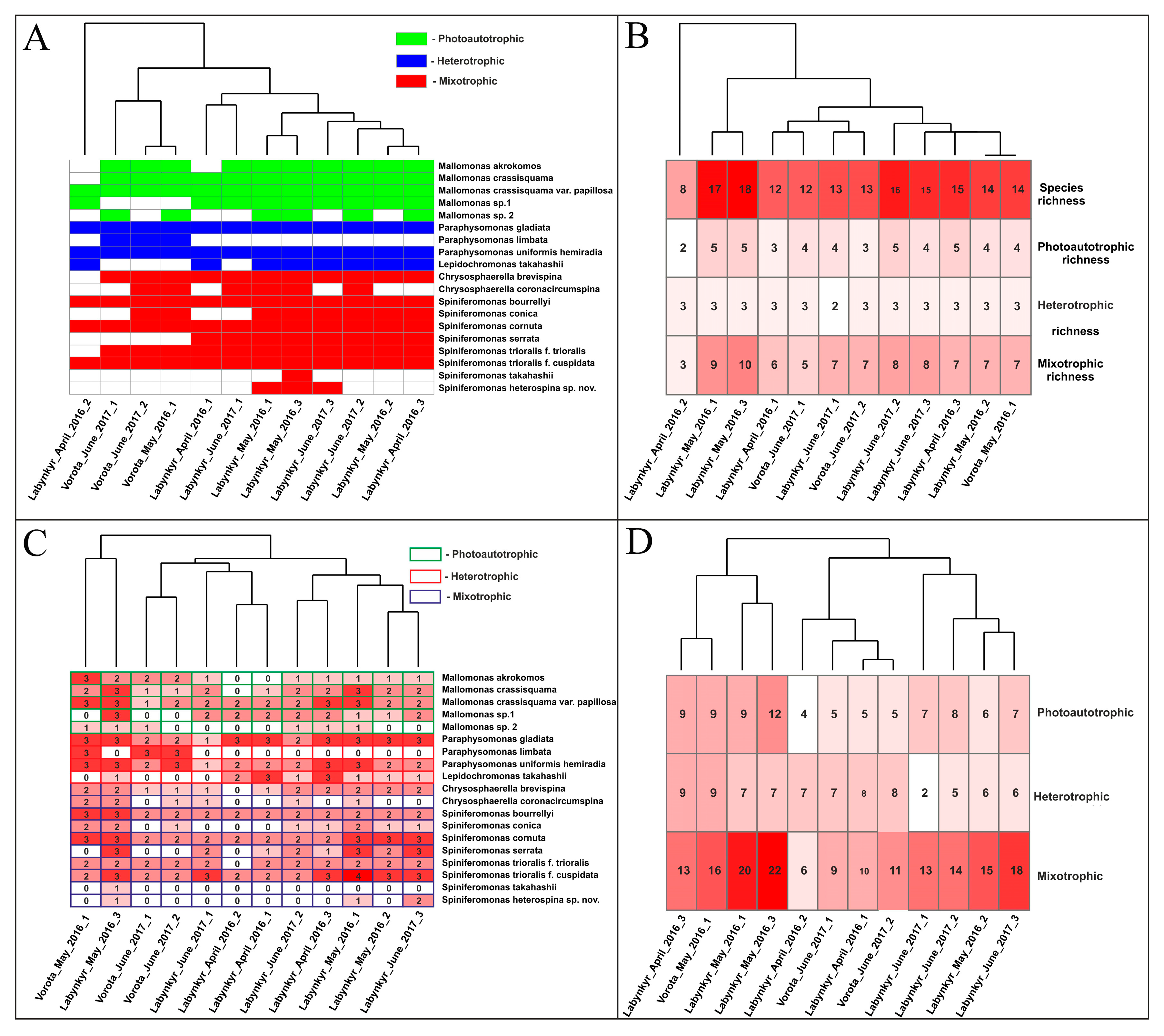
3.1. Features of the Species Composition of Silica-Scaled Chrysophytes in the Studied Lakes
3.2. Development of Silica-Scaled Chrysophytes Depending on Environmental Factors
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Kristiansen, J.; Škaloud, P. Chrysophyta. Handbook of the Protists, 2nd ed.; Springer International Publishing: Cham, Switzerland, 2016; pp. 1–38. [Google Scholar]
- Kapustin, D.; Kulikovskiy, M. Chrysosphaerella septentrionalis sp. nov. (Chrysophyceae, Chromulinales), a New Species from the Arctic Including the Description of Chrysosphaerellaceae, fam. nov. Plants 2022, 11, 3166. [Google Scholar] [CrossRef]
- Kapustin, D.A.; Guiry, M.D. Reinstatement of Lepidochromonas Kristiansen (Lepidochromonadaceae fam. nov., Chrysophyceae). Phytotaxa 2019, 413, 49–53. [Google Scholar] [CrossRef]
- Bhatti, S.; Colman, B. Inorganic carbon acquisition by the chrysophyte alga Mallomonas papillosa. Can. J. Bot. 2005, 83, 891–897. [Google Scholar] [CrossRef]
- Scoble, J.M.; Cavalier-Smith, T. Scale evolution in Paraphysomonadida (Chrysophyceae): Sequence phylogeny and revised taxonomy of Paraphysomonas, new genus Clathromonas, and 25 new species. Eur. J. Protistol. 2014, 50, 551–592. [Google Scholar] [CrossRef] [Green Version]
- Olsen, N.E.; Poulsen, L.K.; Reuss, N.; Steinarsdottir, S.S. A new subspecies of Paraphysomonas punctata (Paraphysomonadaceae, Chrysophyceae). Nord. J. Bot. 1999, 19, 635–640. [Google Scholar] [CrossRef]
- Zakharova, Y.; Bashenkhaeva, M.; Galachyants, Y.; Petrova, D.; Tomberg, I.; Marchenkov, A.; Kopyrina, L.; Likhoshway, Y. Variability of Microbial Communities in Two Long-Term Ice-Covered Freshwater Lakes in the Subarctic Region of Yakutia, Russia. Microb. Ecol. 2021, 84, 958–973. [Google Scholar] [CrossRef]
- Eloranta, P. Biogeography of chrysophytes in Finnish lakes. Chrysophyte Algae: Ecology, Phylogeny and Development; Cambridge University Press: Cambridge, UK, 1995; pp. 214–231. [Google Scholar] [CrossRef]
- Ikävalko, J. On the presence of some selected Heterokontophyta (Chrysophyceae, Dictyochophyceae, Bicocoecidae) and cysts (“archaeomonads”) from sea ice—A synopsis. Nova Hedw. Beih. 2001, 122, 41–54. [Google Scholar]
- Lim, E.L.; Dennett, M.R.; Caron, D.A. The ecology of Paraphysomonas imperforata based on studies employing oligonucleotide probe identification in coastal water samples and enrichment cultures. Limnol. Oceanogr. 1999, 44, 37–51. [Google Scholar] [CrossRef] [Green Version]
- Siver, P.A. The distribution of chrysophytes along environmental gradients: Their use as biological indicators. Chrysophyte Algae; Cambridge University Press: Cambridge, UK, 1995; pp. 232–268. [Google Scholar]
- Bessudova, A.Y.; Tomberg, I.; Firsova, A.D.; Kopyrina, L.I.; Likhoshway, Y.V. Silica-scaled chrysophytes in lakes Labynkyr and Vorota of the Sakha (Yakutia) Republic, Russia. Nova Hedwig. Beih. 2019, 148, 35–48. [Google Scholar] [CrossRef]
- Kopyrina, L.; Firsova, A.; Rodionova, E.; Zakharova, Y.; Bashenkhaeva, M.; Usoltseva, M.; Likhoshway, Y. The insight into diatom diversity, ecology, and biogeography of an extreme cold ultraoligotrophic Lake Labynkyr at the Pole of Cold in the northern hemisphere. Extremophiles 2020, 24, 603–623. [Google Scholar] [CrossRef]
- Warnes, G.R.; Bolker, B.; Bonebakker, L.; Gentleman, R.; Liaw, W.H.A.; Lumley, T.; Maechler, M.; Magnusson, A.; Moeller, S.; Schwartz, M.; et al. Package “gplots”: Various R Programming Tools for Plotting Data; R Packag, Version 2.17.0; ScienceOpen: Berlin, Germany, 2015.
- Oksanen, J. Vegan: Ecological Diversity. R. Project. 2018. Available online: https://cran.r-project.org/web/packages/vegan/vignettes/diversity-vegan.pdf (accessed on 30 August 2022).
- Voloshko, L.N. Species of the genus Spiniferomonas (Chrysophyceae, Paraphysomonadaceae) in water bodies of the Russian North. Bot. Zhurn. 2013, 98, 848–858. (In Russian) [Google Scholar]
- Bessudova, A.Y.; Likhoshway, Y.V. Silica chrysophytes (Chrysophyceae) of the Boguchany reservoir. Modern science: Actual problems of theory and practice. Series “Natural and technical sciences (General biology). Acta Biol. Sib. 2017, 11, 4–11. (In Russian) [Google Scholar]
- Siver, P.A. Inferring the specific conductivity of lake water with scaled chrysophytes. Limnol. Oceanogr. 1993, 38, 1480–1492. [Google Scholar] [CrossRef]
- Kapustin, D.A.; Gusev, E.S. Silica-scaled chrysophytes from West Java (Indonesia) including description of a new Chrysosphaerella species. Nova Hedwig. Beih. 2019, 148, 11–20. [Google Scholar] [CrossRef]
- Kristiansen, J. Silica-scaled chrysophytes from West Greenland: Disko Island and the Søndre Strømfjord region. Nord. J. Bot. 1992, 12, 525–536. [Google Scholar] [CrossRef]
- Nemcova, Y.; Rott, E. Diversity of Silica-scaled Chrysophytes in High-altitude Alpine Sites (North Tyrol, Austria) Including a Description of Mallomonas pechlaneri sp. nov. Cryptogam. Algologie 2018, 39, 63–83. [Google Scholar] [CrossRef]
- Bessudova, A.; Gabyshev, V.; Firsova, A.; Gabysheva, O.; Bukin, Y.; Likhoshway, Y. Diversity Variation of Silica-Scaled Chrysophytes Related to Differences in Physicochemical Variables in Estuaries of Rivers in an Arctic Watershed. Sustainability 2021, 13, 13768. [Google Scholar] [CrossRef]
- Goldman, J.C.; Caron, D.A. Experimental studies on an omnivorous microflagellate: Implications for grazing and nutrient regeneration in the marine microbial food chain. Deep. Sea Res. Part A. Oceanogr. Res. Pap. 1985, 32, 899–915. [Google Scholar] [CrossRef]
- Goldman, J. Dennett Dynamics of prey selection by an omnivorous flagellate. Mar. Ecol. Prog. Ser. 1990, 59, 183–194. [Google Scholar] [CrossRef]
- Charvet, S.; Vincent, W.F.; Lovejoy, C. Chrysophytes and other protists in High Arctic lakes: Molecular gene surveys, pigment signatures and microscopy. Polar Biol. 2011, 35, 733–748. [Google Scholar] [CrossRef]
- Terrado, R.; Pasulka, A.L.; Lie, A.A.-Y.; Orphan, V.; Heidelberg, K.; A Caron, D. Autotrophic and heterotrophic acquisition of carbon and nitrogen by a mixotrophic chrysophyte established through stable isotope analysis. ISME J. 2017, 11, 2022–2034. [Google Scholar] [CrossRef] [Green Version]
- Bessudova, A.Y.; Domysheva, V.M.; Firsova, A.D.; Likhoshway, Y.V. Silica-scaled chrysophytes of Lake Baikal. Acta Biol. Sib. 2017, 3, 47–56. [Google Scholar] [CrossRef] [Green Version]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Lake Labynkyr April 2016 | Lake Labynkyr May 2016 | Lake Vorota May 2016 | Lake Labynkyr June 2017 | Lake Vorota June 2017 | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Stations | 1 | 2 | 3 | 1 | 2 | 3 | 1 | 1 | 2 | 3 | 1 | 2 | |
| No. | Species | ||||||||||||
| Paraphysomonas gladiata Preisig et Hibberd | C | C | C | C | C | C | C | VR | R | C | R | R | |
| P. limbata Preisig et Hibberd | - | - | - | - | - | - | C | - | - | - | C | C | |
| P. uniformis hemiradia Scoble et Cavalier-Smith | R | R | C | C | R | C | C | VR | R | R | R | C | |
| Lepidochromonas takahashii (Cronberg et Kristiansen) Kapustin et Guiry | C | R | C | VR | VR | VR | - | - | VR | VR | - | - | |
| Chrysosphaerella brevispina Korshikov | VR | - | R | R | R | R | R | VR | R | R | V | VR | |
| C. coronacircumspina Wujek et Kristiansen | - | - | - | VR | - | R | R | VR | VR | - | - | VR | |
| Spiniferomonas bourrellyi Takahashi | R | R | R | R | R | C | C | R | R | R | R | R | |
| S. conica Takahashi | - | - | VR | R | VR | R | R | - | VR | VR | - | VR | |
| S. cornuta Balonov | R | R | R | C | C | C | C | R | R | C | R | R | |
| S. serrata Nicholls | VR | - | VR | C | R | C | - | R | R | C | - | - | |
| S. trioralis f. trioralis Takahashi | R | - | R | R | R | R | R | R | R | R | R | R | |
| S. trioralis f. cuspidata Balonov | R | R | C | A | C | A | R | C | R | C | R | R | |
| S. takahashii Nicholls | - | - | - | - | - | VR | - | - | - | - | - | - | |
| Spiniferomonas heterospina sp. nov. | - | - | - | VR | - | VR | - | - | - | R | - | - | |
| Mallomonas akrokomos Ruttner | - | - | VR | VR | VR | R | C | VR | VR | VR | R | R | |
| M. crassisquama (Asmund) Fott var. crassisquama | VR | - | R | C | R | C | R | R | R | R | VR | VR | |
| M. crassisquama var. papillosa Siver et Skogstad | R | R | C | C | R | C | C | R | R | R | VR | R | |
| Mallomonas sp. 1 | R | R | R | VR | VR | C | - | R | R | R | - | - | |
| Mallomonas sp. 2 | - | - | VR | VR | - | VR | VR | - | VR | - | VR | - | |
| Trophic Mode Type | F Value | p Value |
|---|---|---|
| Photoautotrophic species | 1.371 | 0.0424 |
| Heterotrophic species | 0.523 | 0.481 |
| Mixotrophic species | 4.412 | 0.0397 |
| Trophic Mode Type | Parameter | r Value | p Value |
|---|---|---|---|
| Photoautotrophic species | Ice thickness (cm) | −0.14 | 0.19 |
| Snow cover thickness (cm) | −0.15 | 0.15 | |
| T °C | −0.15 | 0.15 | |
| pH | 0.08 | 0.43 | |
| Conductivity (μS/cm) | 0.05 | 0.57 | |
| PO43− (mg/L) | −0.01 | 0.95 | |
| TOC (mgC/L) | −0.03 | 0.74 | |
| Heterotrophic species | Ice thickness (cm) | 0.04 | 0.72 |
| Snow cover thickness (cm) | 0.10 | 0.40 | |
| T °C | 0.10 | 0.40 | |
| pH | 0.16 | 0.13 | |
| Conductivity (μS/cm) | 0.15 | 0.17 | |
| PO43− (mg/L) | 0.01 | 0.98 | |
| TOC (mgC/L) | 0.06 | 0.57 | |
| Mixotrophic species | Ice thickness (cm) | −0.05 | 0.42 |
| Snow cover thickness (cm) | −0.21 | 0.01 | |
| T °C | −0.21 | 0.01 | |
| pH | 0.10 | 0.14 | |
| Conductivity (μS/cm) | 0.08 | 0.25 | |
| PO43− (mg/L) | 0.05 | 0.43 | |
| TOC (mgC/L) | −0.06 | 0.39 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bessudova, A.; Firsova, A.; Bukin, Y.; Kopyrina, L.; Zakharova, Y.; Likhoshway, Y. Under-Ice Development of Silica-Scaled Chrysophytes with Different Trophic Mode in Two Ultraoligotrophic Lakes of Yakutia. Diversity 2023, 15, 326. https://doi.org/10.3390/d15030326
Bessudova A, Firsova A, Bukin Y, Kopyrina L, Zakharova Y, Likhoshway Y. Under-Ice Development of Silica-Scaled Chrysophytes with Different Trophic Mode in Two Ultraoligotrophic Lakes of Yakutia. Diversity. 2023; 15(3):326. https://doi.org/10.3390/d15030326
Chicago/Turabian StyleBessudova, Anna, Alena Firsova, Yurij Bukin, Lubov Kopyrina, Yulia Zakharova, and Yelena Likhoshway. 2023. "Under-Ice Development of Silica-Scaled Chrysophytes with Different Trophic Mode in Two Ultraoligotrophic Lakes of Yakutia" Diversity 15, no. 3: 326. https://doi.org/10.3390/d15030326