
The Role of Uncultivated Habitats in Supporting Wild Bee Communities in Mediterranean Agricultural Landscapes

Abstract
:1. Introduction
2. Materials and Methods
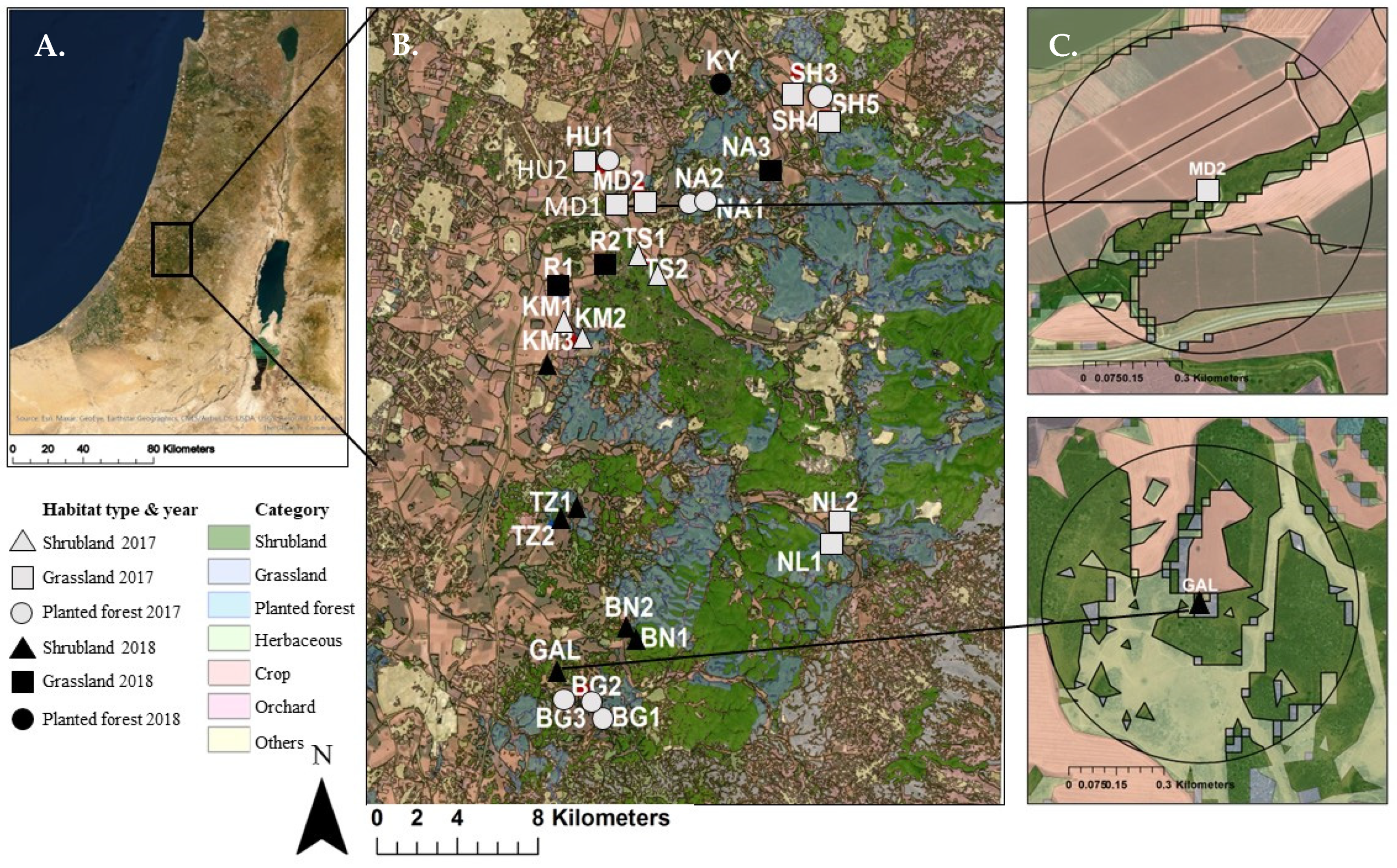
2.1. Study Area
2.2. Study Design
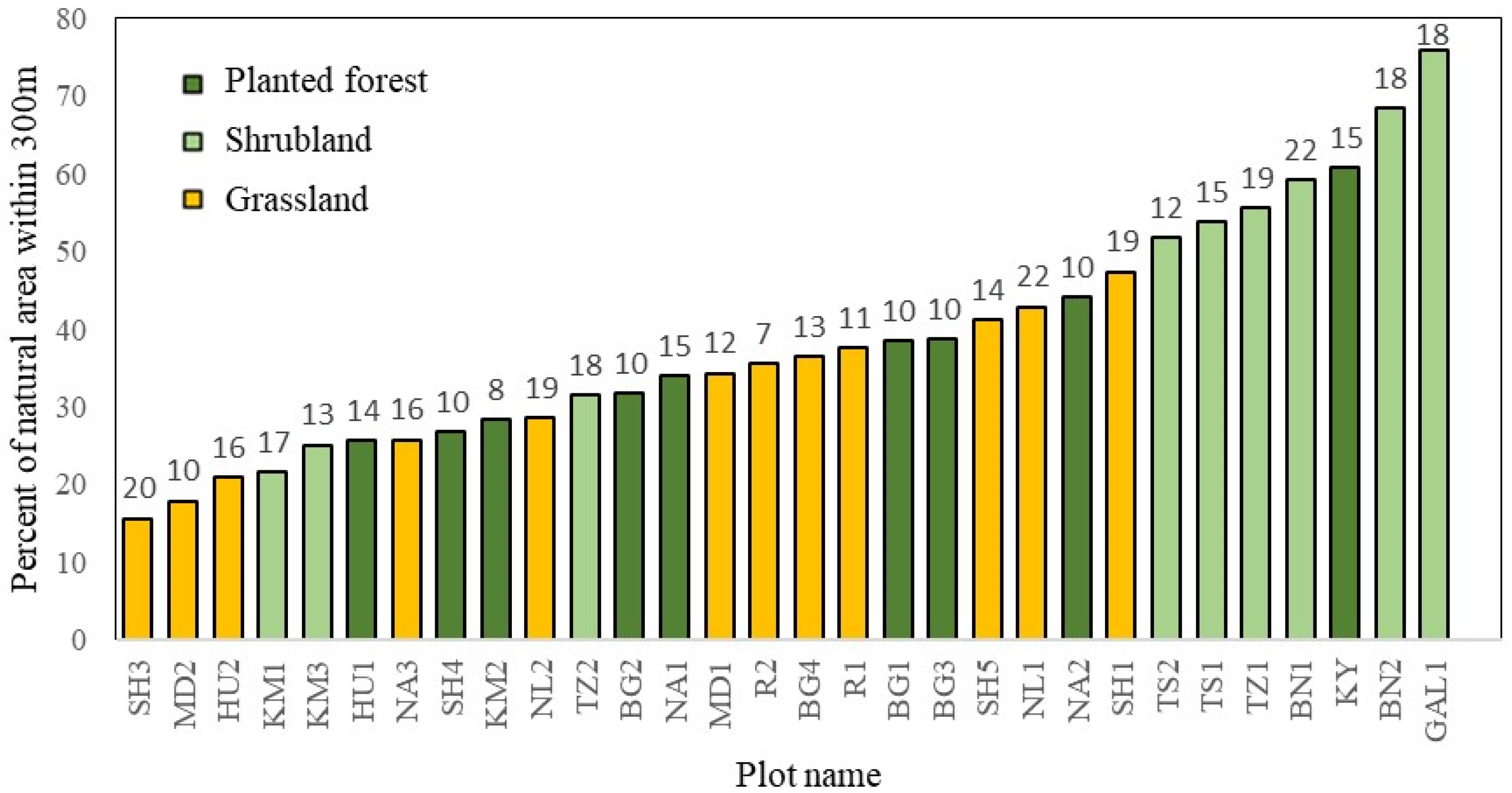
2.3. Landscape Parameters: Land-Use Classification and Analysis
2.4. Field Data Collection
2.4.1. Local Habitat Parameters
2.4.2. Bee Sampling
2.5. Bee Identification and Functional Traits
2.6. Data Analysis
2.6.1. Wild Bee and Flower Abundance and Species Richness
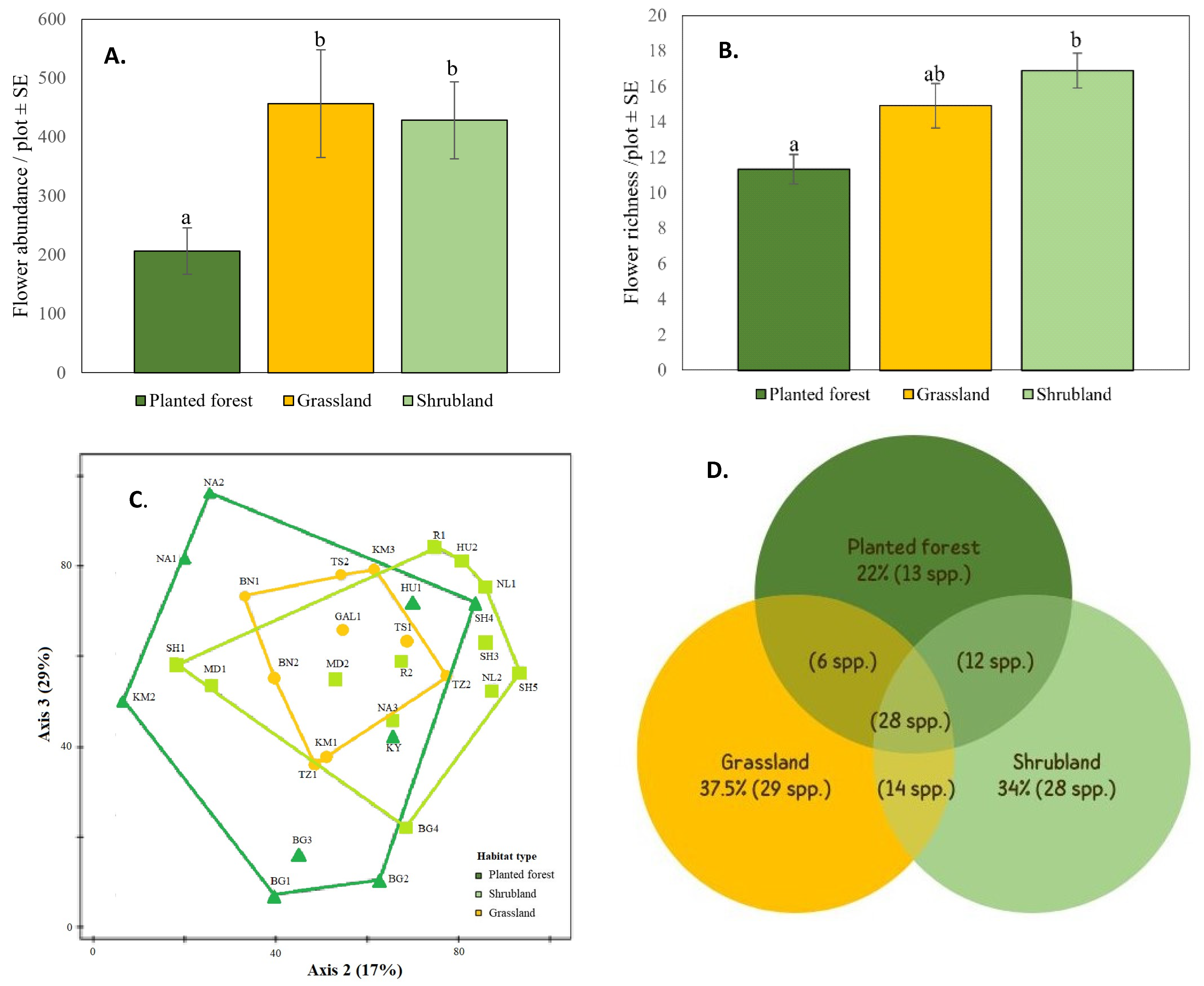
2.6.2. Wild Bee and Flower Species Composition
3. Results
3.1. Local Habitat Characteristics
3.1.1. Microclimate
3.1.2. Forage and Nesting Resources
3.2. Bee Community
3.2.1. Bee Abundance and Species Richness
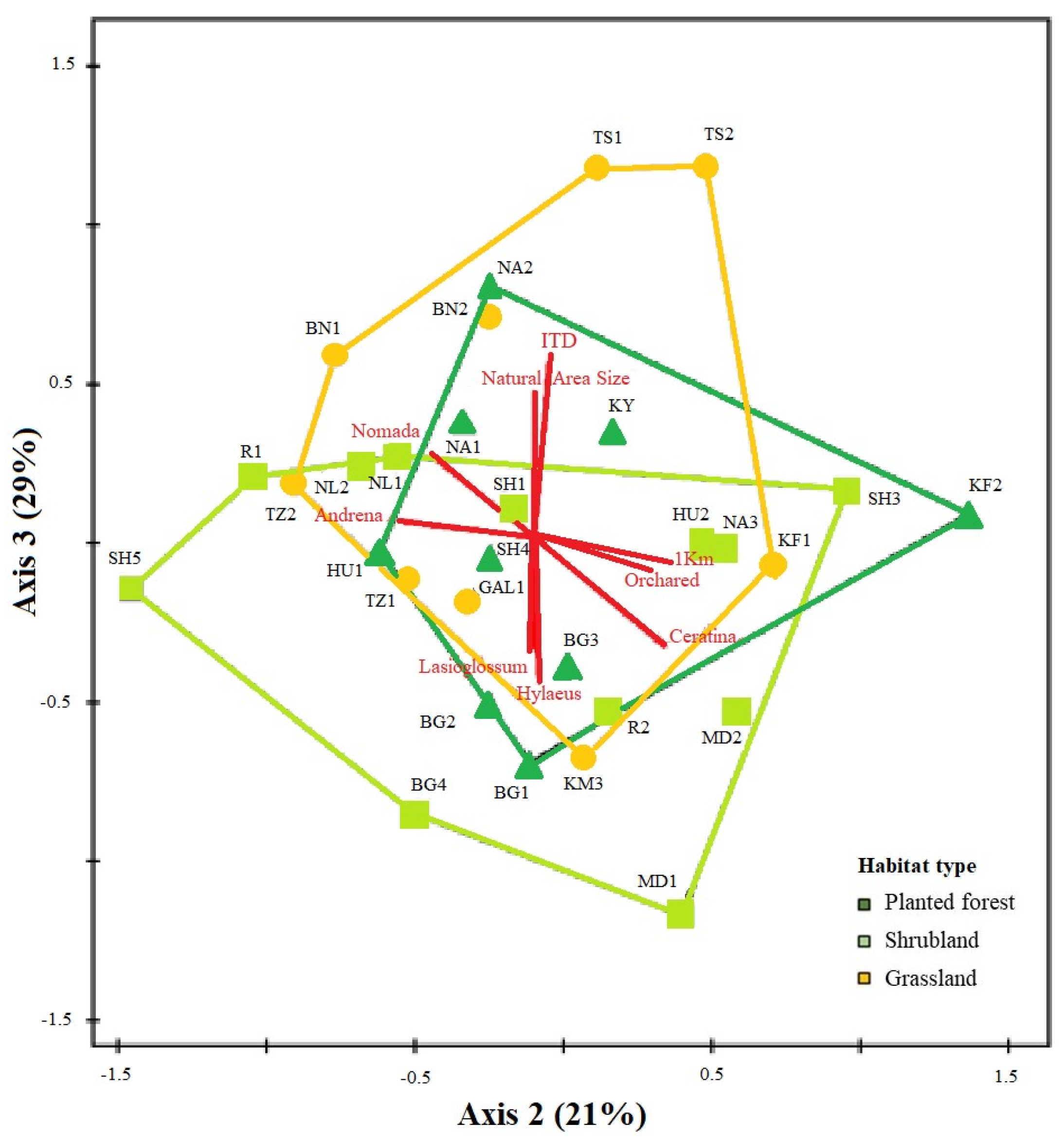
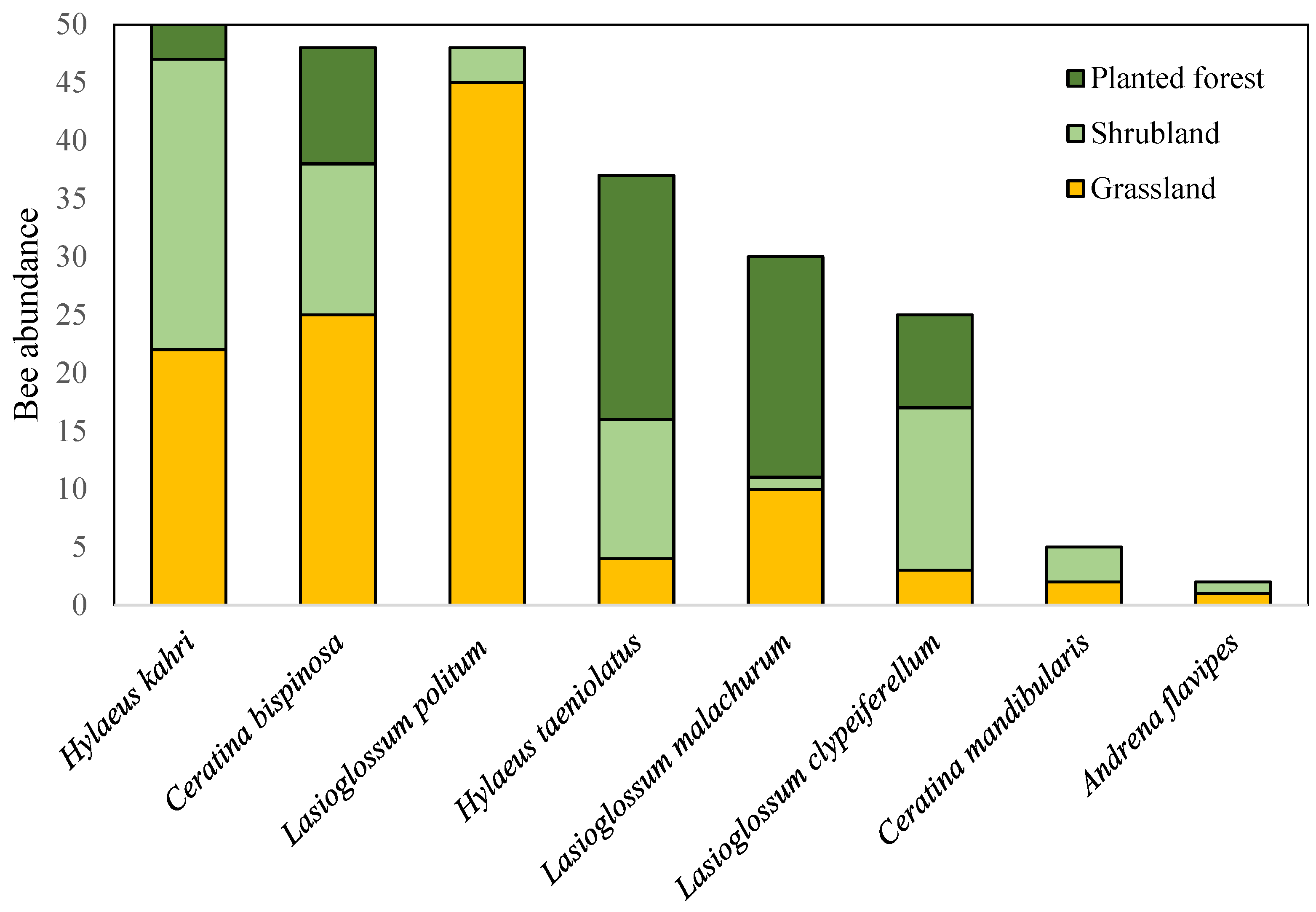
3.2.2. Bee Community Composition
3.2.3. Bee Functional Traits
- (a)
- Nesting guild
- (b) Sociality
- (c) Foraging guild (lecty)
- (d) Tongue length and body size (ITD)
3.2.4. Crop Pollinators Species
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Jackson, L.E.; Brussaard, L.; de Ruiter, P.C.; Pascual, U.; Perrings, C.; Bawa, K. Agrobiodiversity. In Encyclopedia of Biodiversity, 2nd ed.; Levin, S.A., Ed.; Academic Press: Waltham, MA, USA, 2013; pp. 126–135. [Google Scholar]
- Lichtenberg, E.M.; Kennedy, C.M.; Kremen, C.; Batáry, P.; Berendse, F.; Bommarco, R.; Bosque-Pérez, N.A.; Carvalheiro, L.G.; Snyder, W.E.; Williams, N.M.; et al. A global synthesis of the effects of diversified farming systems on arthropod diversity within fields and across agricultural landscapes. Glob. Chang. Biol. 2017, 23, 4946–4957. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Estrada-Carmona, N.; Sánchez, A.C.; Remans, R.; Jones, S.K. Complex agricultural landscapes host more biodiversity than simple ones: A global meta-analysis. Proc. Natl. Acad. Sci. USA 2022, 119, e2203385119. [Google Scholar] [CrossRef] [PubMed]
- Ollerton, J.; Winfree, R.; Tarrant, S. How many flowering plants are pollinated by animals? Oikos 2011, 120, 321–326. [Google Scholar] [CrossRef]
- Klein, A.-M.; Vaissière, B.E.; Cane, J.H.; Steffan-Dewenter, I.; Cunningham, S.A.; Kremen, C.; Tscharntke, T. Importance of pollinators in changing landscapes for world crops. Proc. R. Soc. B Biol. Sci. 2007, 274, 303–313. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Eilers, E.J.; Kremen, C.; Greenleaf, S.; Garber, A.K.; Klein, A.-M. Contribution of Pollinator-Mediated Crops to Nutrients in the Human Food Supply. PLoS ONE 2011, 6, e21363. [Google Scholar] [CrossRef] [PubMed]
- Brittain, C.; Kremen, C.; Garber, A.; Klein, A.-M. Pollination and Plant Resources Change the Nutritional Quality of Almonds for Human Health. PLoS ONE 2014, 9, e90082. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Klatt, B.K.; Holzschuh, A.; Westphal, C.; Clough, Y.; Smit, I.; Pawelzik, E.; Tscharntke, T. Bee pollination improves crop quality, shelf life and commercial value. Proc. R. Soc. B Biol. Sci. 2014, 281, 20132440. [Google Scholar] [CrossRef]
- Garibaldi, L.A.; Steffan-Dewenter, I.; Winfree, R.; Aizen, M.A.; Bommarco, R.; Cunningham, S.A.; Kremen, C.; Carvalheiro, L.G.; Harder, L.D.; Afik, O.; et al. Wild Pollinators Enhance Fruit Set of Crops Regardless of Honey Bee Abundance. Science 2013, 339, 1608–1611. [Google Scholar] [CrossRef]
- Kennedy, C.M.; Lonsdorf, E.; Neel, M.C.; Williams, N.M.; Ricketts, T.H.; Winfree, R.; Bommarco, R.; Brittain, C.; Burley, A.L.; Cariveau, D.; et al. A global quantitative synthesis of local and landscape effects on wild bee pollinators in agroecosystems. Ecol. Lett. 2013, 16, 584–599. [Google Scholar] [CrossRef]
- Senapathi, D.; Fründ, J.; Albrecht, M.; Garratt, M.P.D.; Kleijn, D.; Pickles, B.J.; Potts, S.G.; An, J.; Andersson, G.K.S.; Bänsch, S.; et al. Wild insect diversity increases inter-annual stability in global crop pollinator communities. Proc. R. Soc. B Biol. Sci. 2021, 288, 20210212. [Google Scholar] [CrossRef]
- Potts, S.G.; Biesmeijer, J.C.; Kremen, C.; Neumann, P.; Schweiger, O.; Kunin, W.E. Global pollinator declines: Trends, impacts and drivers. Trends Ecol. Evol. 2010, 25, 345–353. [Google Scholar] [CrossRef] [PubMed]
- Koh, I.; Lonsdorf, E.V.; Williams, N.M.; Brittain, C.; Isaacs, R.; Gibbs, J.; Ricketts, T.H. Modeling the status, trends, and impacts of wild bee abundance in the United States. Proc. Natl. Acad. Sci. USA 2016, 113, 140–145. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bartomeus, I.; Ascher, J.S.; Gibbs, J.; Danforth, B.N.; Wagner, D.L.; Hedtke, S.M.; Winfree, R. Historical changes in northeastern US bee pollinators related to shared ecological traits. Proc. Natl. Acad. Sci. USA 2013, 110, 4656–4660. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Carvalheiro, L.G.; Kunin, W.E.; Keil, P.; Aguirre-Gutiérrez, J.; Ellis, W.N.; Fox, R.; Groom, Q.; Hennekens, S.; Van Landuyt, W.; Maes, D.; et al. Species richness declines and biotic homogenisation have slowed down for NW-European pollinators and plants. Ecol. Lett. 2013, 16, 870–878. [Google Scholar] [CrossRef] [Green Version]
- Winfree, R.; Aguilar, R.; Vazquez, D.P.; LeBuhn, G.; Aizen, M.A. A meta-analysis of bees’ responses to anthropogenic disturbance. Ecology 2009, 90, 2068–2076. [Google Scholar] [CrossRef]
- Ollerton, J.; Erenler, H.; Edwards, M.; Crockett, R. Extinctions of aculeate pollinators in Britain and the role of large-scale agricultural changes. Science 2014, 346, 1360–1362. [Google Scholar] [CrossRef] [Green Version]
- Brown, M.J.F.; Dicks, L.V.; Paxton, R.J.; Baldock, K.C.R.; Barron, A.B.; Chauzat, M.-P.; Freitas, B.M.; Goulson, D.; Jepsen, S.; Kremen, C.; et al. A horizon scan of future threats and opportunities for pollinators and pollination. PeerJ 2016, 4, e2249. [Google Scholar] [CrossRef] [Green Version]
- Dicks, L.V.; Breeze, T.D.; Ngo, H.T.; Senapathi, D.; An, J.; Aizen, M.A.; Basu, P.; Buchori, D.; Galetto, L.; Garibaldi, L.A.; et al. A global-scale expert assessment of drivers and risks associated with pollinator decline. Nat. Ecol. Evol. 2021, 5, 1453–1461. [Google Scholar] [CrossRef]
- Kleijn, D.; van Langevelde, F. Interacting effects of landscape context and habitat quality on flower visiting insects in agricultural landscapes. Basic Appl. Ecol. 2006, 7, 201–214. [Google Scholar] [CrossRef]
- Mandelik, Y.; Winfree, R.; Neeson, T.; Kremen, C. Complementary habitat use by wild bees in agro-natural landscapes. Ecol. Appl. 2012, 22, 1535–1546. [Google Scholar] [CrossRef]
- Hannon, L.E.; Sisk, T.D. Hedgerows in an agri-natural landscape: Potential habitat value for native bees. Biol. Conserv. 2009, 142, 2140–2154. [Google Scholar] [CrossRef]
- Morandin, L.A.; Kremen, C. Hedgerow restoration promotes pollinator populations and exports native bees to adjacent fields. Ecol. Appl. 2013, 23, 829–839. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Campbell, A.J.; Wilby, A.; Sutton, P.; Wäckers, F.L. Do sown flower strips boost wild pollinator abundance and pollination services in a spring-flowering crop? A case study from UK cider apple orchards. Agric. Ecosyst. Environ. 2017, 239, 20–29. [Google Scholar] [CrossRef] [Green Version]
- Albrecht, M.; Kleijn, D.; Williams, N.M.; Tschumi, M.; Blaauw, B.R.; Bommarco, R.; Campbell, A.J.; Dainese, M.; Drummond, F.A.; Entling, M.H.; et al. The effectiveness of flower strips and hedgerows on pest control, pollination services and crop yield: A quantitative synthesis. Ecol. Lett. 2020, 23, 1488–1498. [Google Scholar] [CrossRef] [PubMed]
- Cole, L.J.; Kleijn, D.; Dicks, L.V.; Stout, J.C.; Potts, S.G.; Albrecht, M.; Balzan, M.V.; Bartomeus, I.; Bebeli, P.J.; Bevk, D.; et al. A critical analysis of the potential for EU Common Agricultural Policy measures to support wild pollinators on farmland. J. Appl. Ecol. 2020, 57, 681–694. [Google Scholar] [CrossRef] [PubMed]
- Sanchez, J.A.; Carrasco, A.; La Spina, M.; Pérez-Marcos, M.; Ortiz-Sánchez, F.J. How Bees Respond Differently to Field Margins of Shrubby and Herbaceous Plants in Intensive Agricultural Crops of the Mediterranean Area. Insects 2020, 11, 26. [Google Scholar] [CrossRef] [Green Version]
- Morrison, J.; Izquierdo, J.; Plaza, E.H.; González-Andújar, J.L. The role of field margins in supporting wild bees in Mediterranean cereal agroecosystems: Which biotic and abiotic factors are important? Agric. Ecosyst. Environ. 2017, 247, 216–224. [Google Scholar] [CrossRef]
- McKechnie, I.M.; Thomsen, C.J.M.; Sargent, R.D. Forested field edges support a greater diversity of wild pollinators in lowbush blueberry (Vaccinium angustifolium). Agric. Ecosyst. Environ. 2017, 237, 154–161. [Google Scholar] [CrossRef]
- Pywell, R.F.; Warman, E.A.; Carvell, C.; Sparks, T.H.; Dicks, L.V.; Bennett, D.; Wright, A.; Critchley, C.N.R.; Sherwood, A. Providing foraging resources for bumblebees in intensively farmed landscapes. Biol. Conserv. 2005, 121, 479–494. [Google Scholar] [CrossRef]
- Raderschall, C.A.; Bommarco, R.; Lindström, S.A.M.; Lundin, O. Landscape crop diversity and semi-natural habitat affect crop pollinators, pollination benefit and yield. Agric. Ecosyst. Environ. 2021, 306, 107189. [Google Scholar] [CrossRef]
- Neumüller, U.; Burger, H.; Krausch, S.; Blüthgen, N.; Ayasse, M. Interactions of local habitat type, landscape composition and flower availability moderate wild bee communities. Landsc. Ecol. 2020, 35, 2209–2224. [Google Scholar] [CrossRef]
- Mandelik, Y.; Dayan, T.; Chikatunov, V.; Kravchenko, V. Reliability of a Higher-Taxon Approach to Richness, Rarity, and Composition Assessments at the Local Scale. Conserv. Biol. 2007, 21, 1506–1515. [Google Scholar] [CrossRef] [PubMed]
- Pisanty, G.; Mandelik, Y. Profiling crop pollinators: Life history traits predict habitat use and crop visitation by Mediterranean wild bees. Ecol. Appl. 2015, 25, 742–752. [Google Scholar] [CrossRef] [PubMed]
- Weizel, Y.; Polak, G.; Cohen, Y. Ecology of the Vegetation in Israel; The Ecology Branch, Tel Aviv University: Tel Aviv, Israel, 1978. [Google Scholar]
- Westphal, C.; Bommarco, R.; Carre, G.; Lamborn, E.; Morison, N.; Petanidou, T.; Potts, S.G.; Roberts, S.P.M.; Szentgyorgyi, H.; Tscheulin, T.; et al. Measuring bee diversity in different european habitats and biogeographical regions. Ecol. Monogr. 2008, 78, 653–671. [Google Scholar] [CrossRef] [Green Version]
- Williams, N.M.; Crone, E.E.; Roulston, T.a.H.; Minckley, R.L.; Packer, L.; Potts, S.G. Ecological and life-history traits predict bee species responses to environmental disturbances. Biol. Conserv. 2010, 143, 2280–2291. [Google Scholar] [CrossRef]
- Greenleaf, S.S.; Williams, N.M.; Winfree, R.; Kremen, C. Bee foraging ranges and their relationship to body size. Oecologia 2007, 153, 589–596. [Google Scholar] [CrossRef]
- Cariveau, D.P.; Nayak, G.K.; Bartomeus, I.; Zientek, J.; Ascher, J.S.; Gibbs, J.; Winfree, R. The Allometry of Bee Proboscis Length and Its Uses in Ecology. PLoS ONE 2016, 11, e0151482. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pisanty, G.; Klein, A.-M.; Mandelik, Y. Do wild bees complement honeybee pollination of confection sunflowers in Israel? Apidologie 2014, 45, 235–247. [Google Scholar] [CrossRef]
- Pisanty, G.; Afik, O.; Wajnberg, E.; Mandelik, Y. Watermelon pollinators exhibit complementarity in both visitation rate and single-visit pollination efficiency. J. Appl. Ecol. 2016, 53, 360–370. [Google Scholar] [CrossRef] [Green Version]
- Bolker, B.M.; Brooks, M.E.; Clark, C.J.; Geange, S.W.; Poulsen, J.R.; Stevens, M.H.H.; White, J.-S.S. Generalized linear mixed models: A practical guide for ecology and evolution. Trends Ecol. Evol. 2009, 24, 127–135. [Google Scholar] [CrossRef]
- Harrison, X.A. Using observation-level random effects to model overdispersion in count data in ecology and evolution. PeerJ 2014, 2, e616. [Google Scholar] [CrossRef] [Green Version]
- Johnson, J.B.; Omland, K.S. Model selection in ecology and evolution. Trends Ecol. Evol. 2004, 19, 101–108. [Google Scholar] [CrossRef]
- Magurran, A.E. Measuring Biological Diversity; Wiley-Blackwell: Hoboken, NJ, USA, 2004. [Google Scholar]
- Peck, J.E. Multivariate Analysis for Ecologists: Step-by-Step, 2nd ed.; MjM Software Design: Gleneden Beach, OR, USA, 2016. [Google Scholar]
- Danin, A.O. Fragman-Sapir. Flora of Israel. Available online: http://flora.org.il/en/plants (accessed on 1 January 2023).
- Schmid, B.; Nottebrock, H.; Esler, K.J.; Pagel, J.; Pauw, A.; Böhning-Gaese, K.; Schurr, F.M.; Schleuning, M. Responses of nectar-feeding birds to floral resources at multiple spatial scales. Ecography 2016, 39, 619–629. [Google Scholar] [CrossRef]
- Biesmeijer, J.C.; Roberts, S.P.M.; Reemer, M.; Ohlemüller, R.; Edwards, M.; Peeters, T.; Schaffers, A.P.; Potts, S.G.; Kleukers, R.; Thomas, C.D.; et al. Parallel Declines in Pollinators and Insect-Pollinated Plants in Britain and the Netherlands. Science 2006, 313, 351. [Google Scholar] [CrossRef] [PubMed]
- Winfree, R. The conservation and restoration of wild bees. Ann. N. Y. Acad. Sci. 2010, 1195, 169–197. [Google Scholar] [CrossRef] [PubMed]
- De Palma, A.; Kuhlmann, M.; Roberts, S.P.M.; Potts, S.G.; Börger, L.; Hudson, L.N.; Lysenko, I.; Newbold, T.; Purvis, A. Ecological traits affect the sensitivity of bees to land-use pressures in European agricultural landscapes. J. Appl. Ecol. 2015, 52, 1567–1577. [Google Scholar] [CrossRef] [PubMed] [Green Version]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Model Parameters | 100 m | 200 m | 300 m | 400 m | 500 m |
|---|---|---|---|---|---|
| p (χ2) | p (χ2) | p (χ2) | p (χ2) | p (χ2) | |
| Bee abundance | |||||
| Total plant cover (%) | 0.01 (6.2) | 0.003 (8.7) | 0.003 (8.4) | ||
| Uncultivated area (%) | 0.02 (4.9) | 0.03 (4.5) | 0.04 (3.9) | ||
| Bee species richness | |||||
| Total plant cover (%) | 0.01 (5.8) | 0.004 (8.2) | 0.003 (8.3) | 0.003 (8.3) | 0.004 (8.1) |
| Uncultivated area (%) | 0.03 (4.7) | 0.04 (3.9) | 0.01 (5.7) | 0.02 (5.4) | 0.03 (4.5) |
| Modeled Variables | Model (ΔAIC, Weight) and Explanatory Variables (− with Negative Effect; + with Positive Effect) | χ2 | p |
|---|---|---|---|
| Relative abundance of above-ground nesting bees | Best-fitted model: (ΔAIC = 0, Weight = 0.32) | 19.5 | <0.001 |
| Flower abundance (−) | 31 | <0.001 | |
| Alternative model: (ΔAIC = 0.5, weight = 0.25) | |||
| Flower abundance (−) | 29.8 | <0.001 | |
| Flower richness (+) | 2.9 | 0.08 | |
| Woody cover (%) | 13.6 | <0.001 | |
| Alternative model: (ΔAIC = 0.5, Weight = 0.25) | |||
| Flower abundance (−) | 20.2 | <0.001 | |
| Woody cover (%) (+) | 26.4 | <0.001 | |
| Uncultivated area within a 300 m radius (−) | 2.9 | 0.08 | |
| Alternative model: (ΔAIC = 1.1, Weight = 0.18) | |||
| Flower abundance (−) | 20.6 | <0.001 | |
| Woody cover (%) (+) | 27.6 | <0.001 | |
| Uncultivated area within a 400 m radius (−) | 2.2 | 0.1 | |
| Proportion of above-ground nesting bee species | Best-fitted model: (ΔAIC = 0, Weight = 0.54) | ||
| Uncultivated area within a 200 m radius (−) | 5.5 | 0.02 | |
| Alternative model: (ΔAIC = 0.3, Weight = 0.46) | |||
| Flower abundance (−) | 2.8 | 0.09 | |
| Uncultivated area within a 200 m radius (−) | 4.8 | 0.02 |
| Explained Variables, Model Type | Model (ΔAIC, Weight)/Explanatory Variables (− Negative Effect; + Positive Effect) | χ2 | p |
|---|---|---|---|
| Relative abundance of oligolectic bees GLMM, Poisson | Best-fitted model: (ΔAIC = 0, Weight = 0.36) | ||
| Flower abundance (+) | 13.9 | <0.001 | |
| Habitat type | 6.7 | 0.07 | |
| Proportion of oligolectic bee species GLMM, Poisson | Best-fitted model: (ΔAIC = 0, Weight = 0.51) | ||
| Woody cover % (−) | 6.6 | 0.01 | |
| Uncultivated area within a 200 m radius (+) | 6.6 | 0.02 | |
| Alternative model: (ΔAIC = 0.1, Weight = 0.49) | |||
| Flower abundance (+) | 5.4 | 0.01 | |
| Proportion of polylectic bee species GLMM, Binomial | Best-fitted model: (ΔAIC = 0, Weight = 0.38) | ||
| Flower richness (−) | 6.9 | <0.001 | |
| Alternative model: (ΔAIC = 0.7, Weight = 0.28) | |||
| Flower abundance (−) | 6.2 | 0.01 | |
| Alternative model: (ΔAIC = 1.4 Weight = 0.19) | |||
| Flower abundance (−) | 5.7 | 0.01 | |
| Uncultivated area within a 200 m radius (−) | 2.4 | 0.11 |
| Explained Variables Model Type | Model (ΔAIC, Weight)/Explanatory Variables (− Negative Effect; + Positive Effect) | χ2 | p |
|---|---|---|---|
| Calculated tongue length LME, Normal | Best-fitted model: (ΔAIC = 0, Weight = 0.9) | ||
| Flower abundance (+) | 17.9 | <0.001 | |
| Proportion of long tongue bee species GLMM, Binomial | Best-fitted model: (ΔAIC = 0, Weight = 0.61) | ||
| Flower abundance (+) | 8.5 | 0.003 | |
| Uncultivated area within a 100 m radius (−) | 5.8 | 0.01 | |
| Alternative model: (ΔAIC = 0.8, Weight = 0.31) | |||
| Flower abundance (+) | 11 | <0.001 | |
| Total plant cover (%) (−) | 2.6 | 0.1 | |
| Uncultivated area within a 100 m radius (−) | 6.4 | 0.01 | |
| Body size (ITD) LME, Normal | Best-fitted model: (ΔAIC = 0, Weight = 0.66) | ||
| Flower abundance (+) | 5.4 | 0.02 | |
| Alternative model: (ΔAIC = 0.2, Weight = 0.34) | |||
| Flower abundance (+) | 5.5 | 0.01 | |
| Uncultivated patch area (+) | 6.1 | 0.01 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Roth, T.; Coll, M.; Mandelik, Y. The Role of Uncultivated Habitats in Supporting Wild Bee Communities in Mediterranean Agricultural Landscapes. Diversity 2023, 15, 294. https://doi.org/10.3390/d15020294
Roth T, Coll M, Mandelik Y. The Role of Uncultivated Habitats in Supporting Wild Bee Communities in Mediterranean Agricultural Landscapes. Diversity. 2023; 15(2):294. https://doi.org/10.3390/d15020294
Chicago/Turabian StyleRoth, Tohar, Moshe Coll, and Yael Mandelik. 2023. "The Role of Uncultivated Habitats in Supporting Wild Bee Communities in Mediterranean Agricultural Landscapes" Diversity 15, no. 2: 294. https://doi.org/10.3390/d15020294