
Effects of Stream Connectivity on Phytoplankton Diversity and Community Structure in Sunken Lakes: A Case Study from an August Survey

Abstract
:1. Introduction
2. Materials and Methods
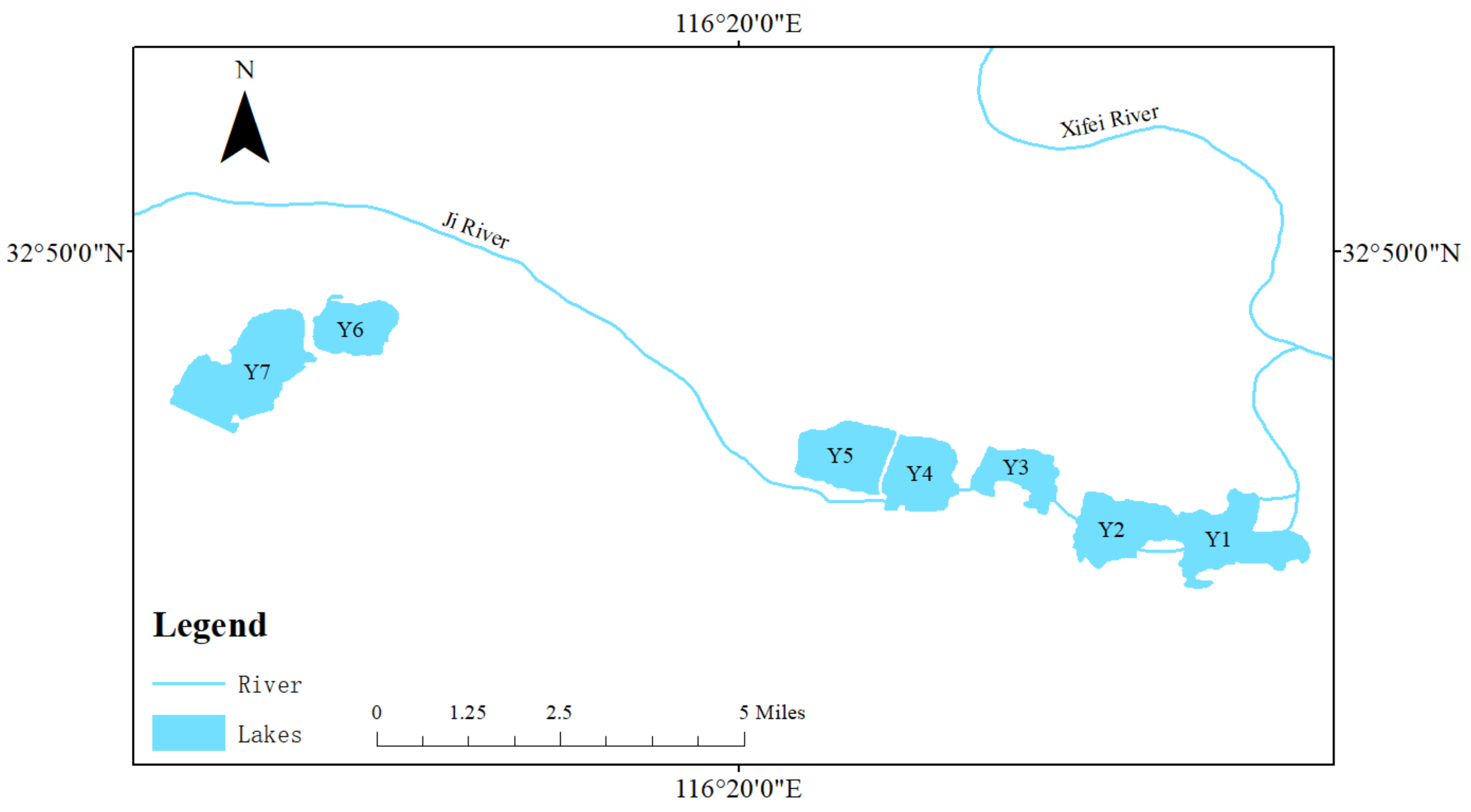
2.1. Sampling Area and Sampling Time
2.2. Collection and Identification of Phytoplankton Samples
2.3. Determination of Physical and Chemical Indicators
2.4. Data Analyses
3. Results
3.1. Ecological Factors Parameters
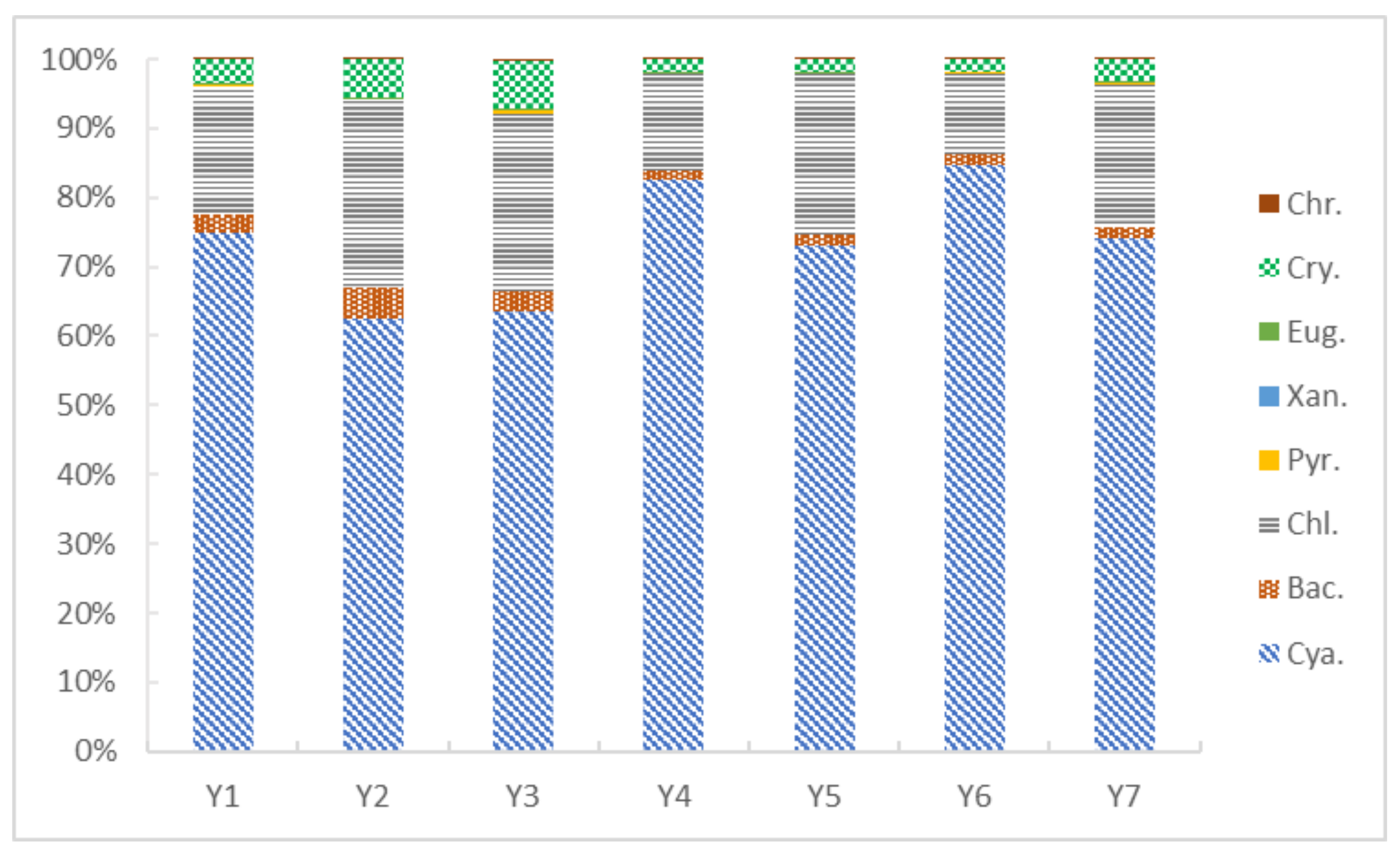
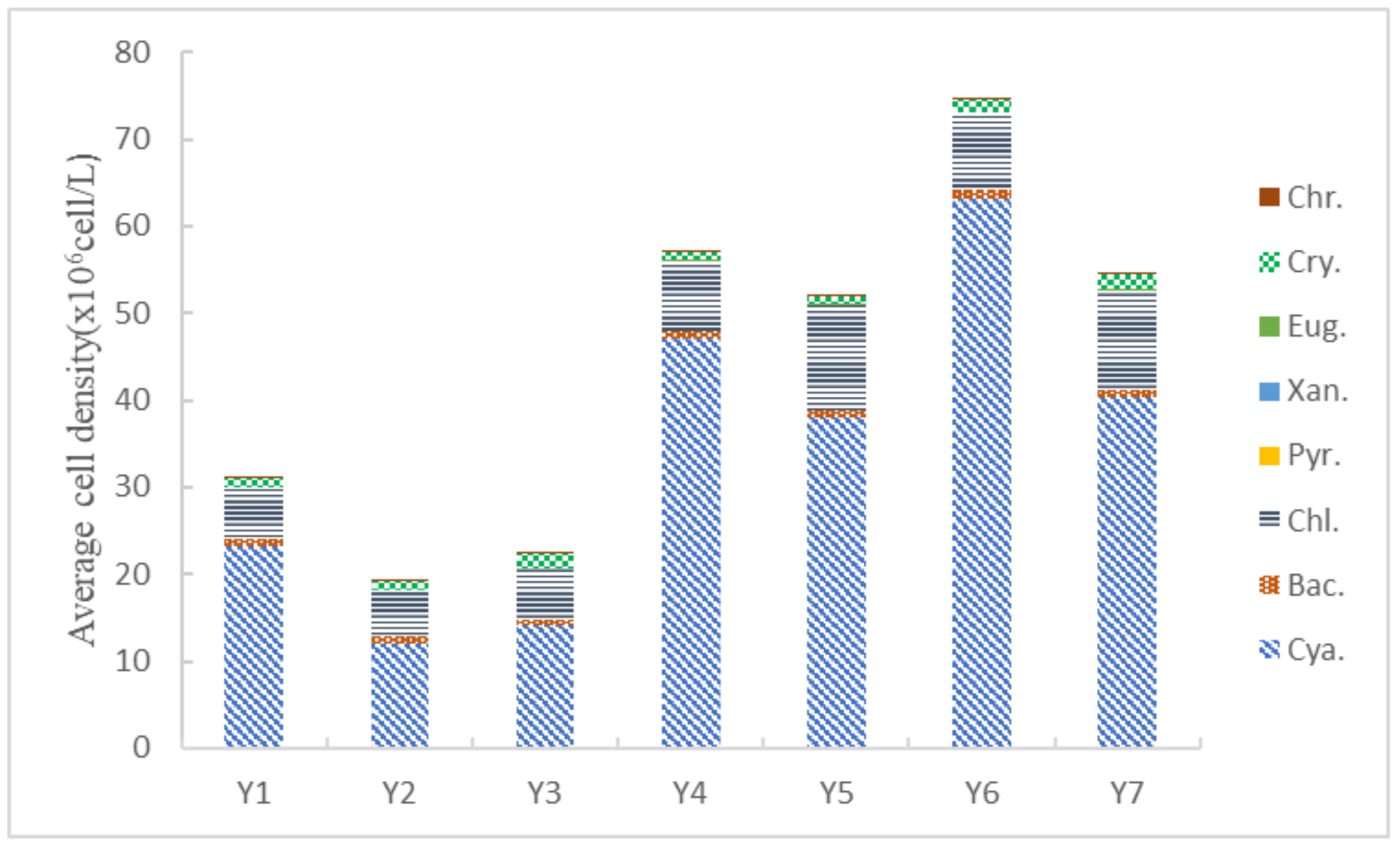
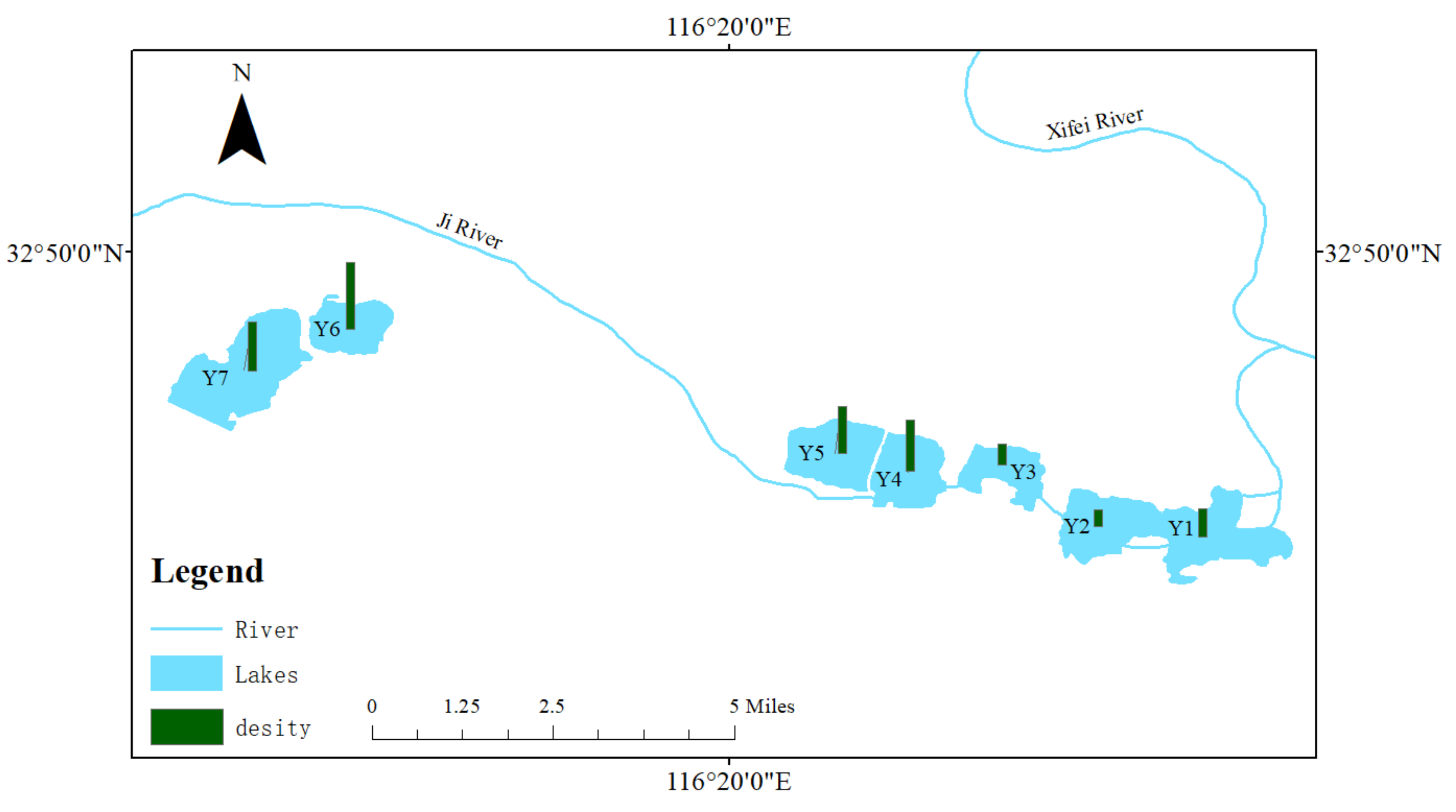
3.2. Phytoplankton Composition and Cell Density
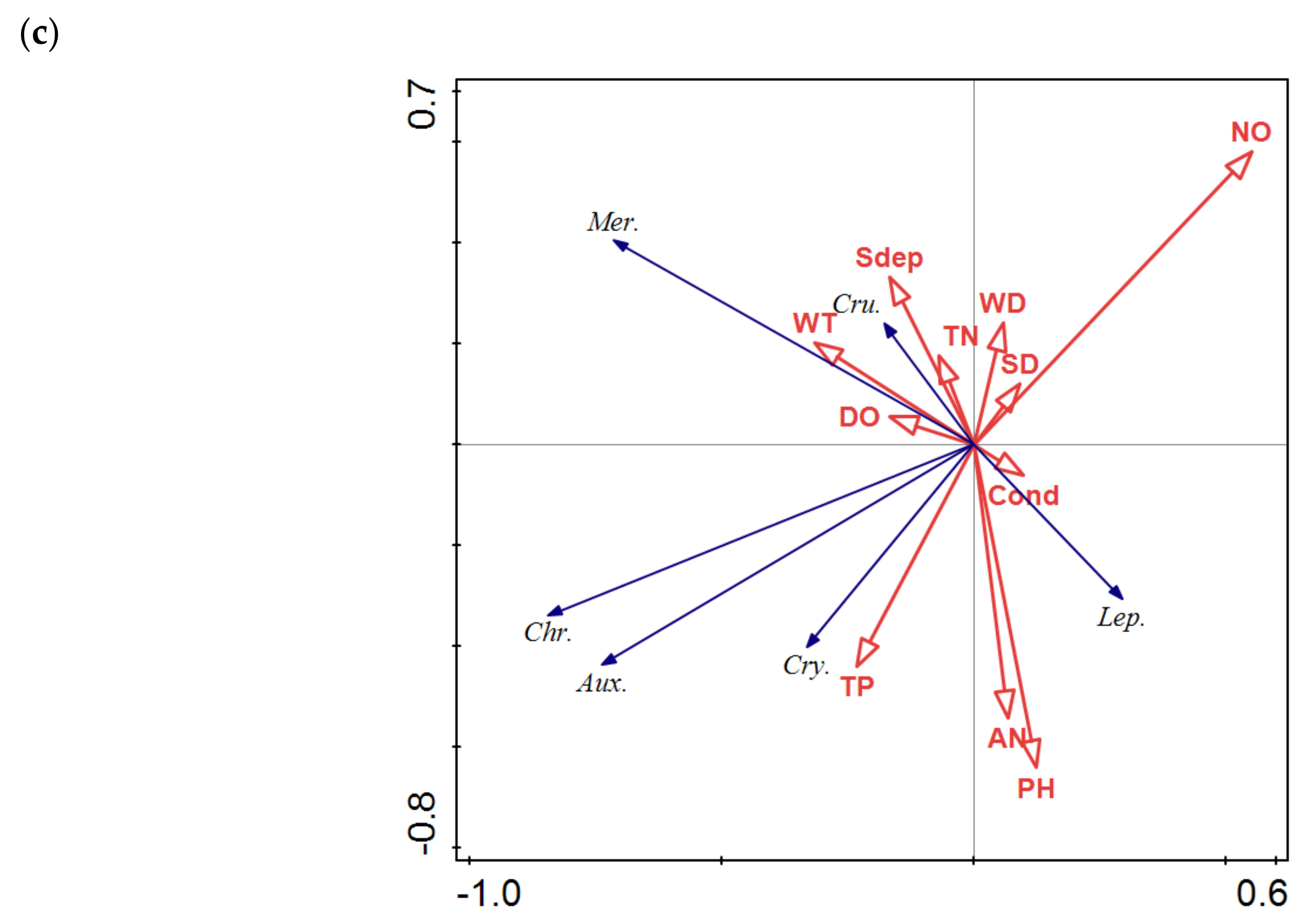
3.3. Relationship between Phytoplankton and Environmental Factors
3.3.1. Pearson Correlation Analysis
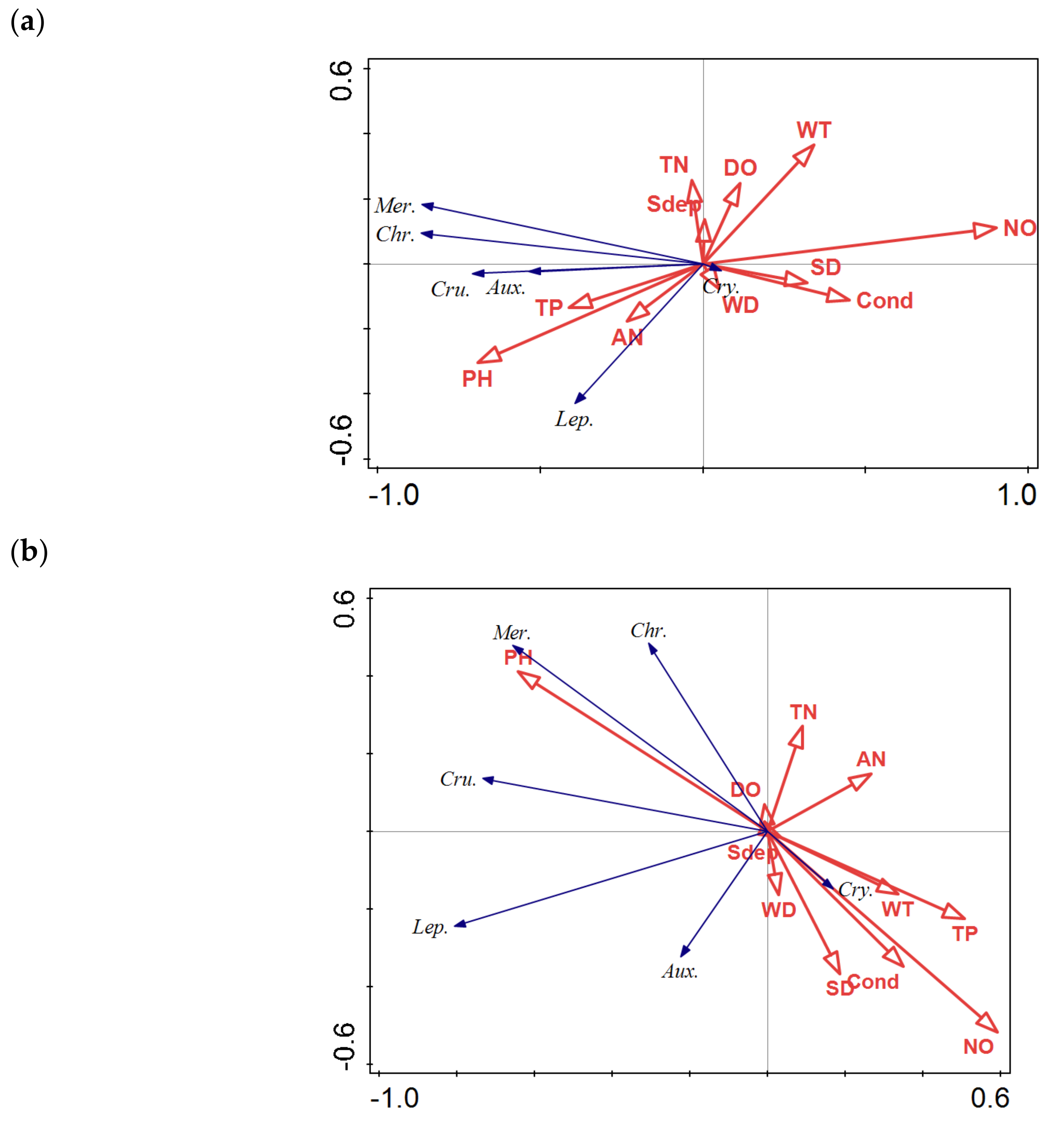
3.3.2. RDA Analysis
3.3.3. Relationships between Phytoplankton and the Environmental Factors in Lakes with Different Stream Connectivity
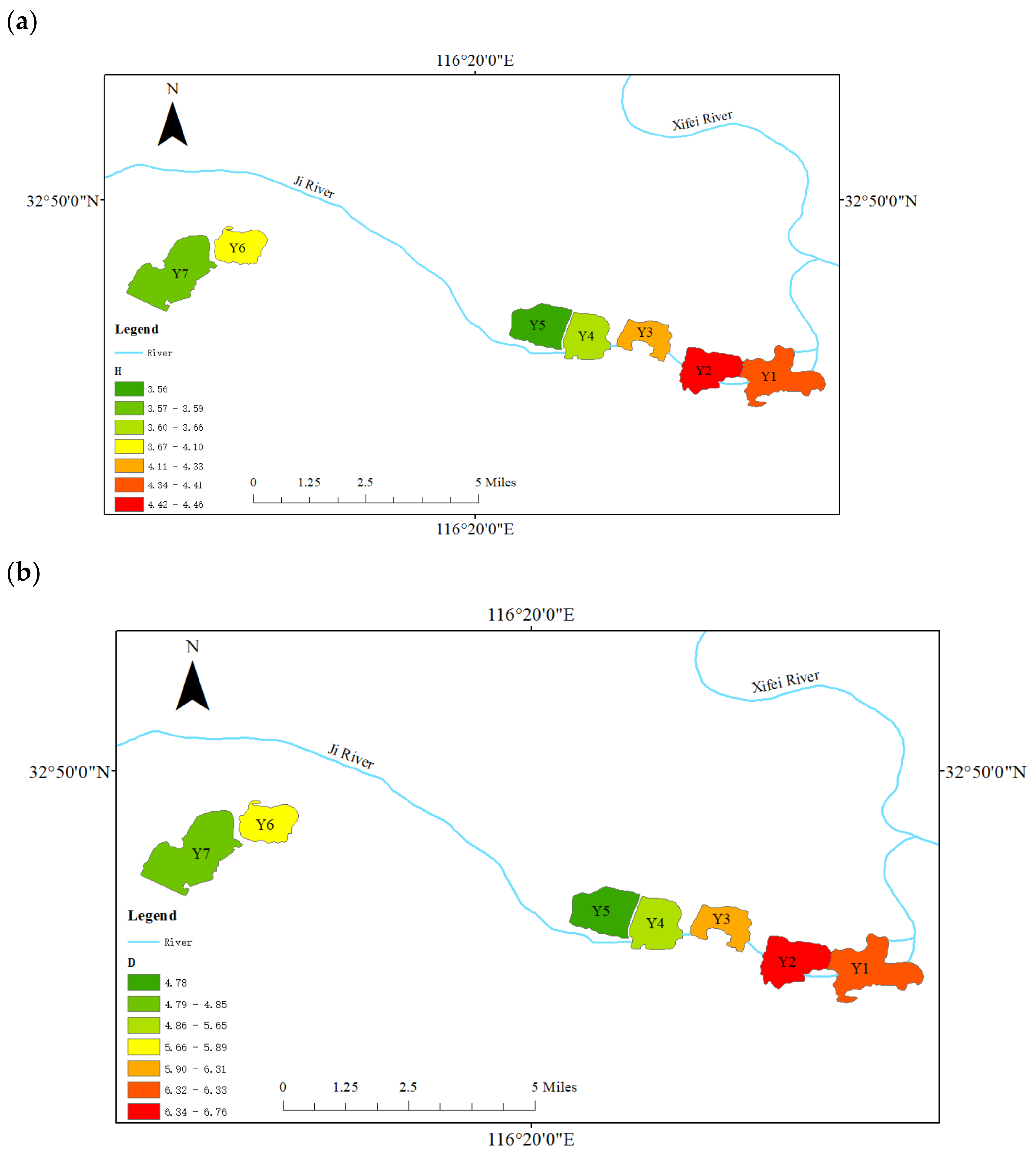
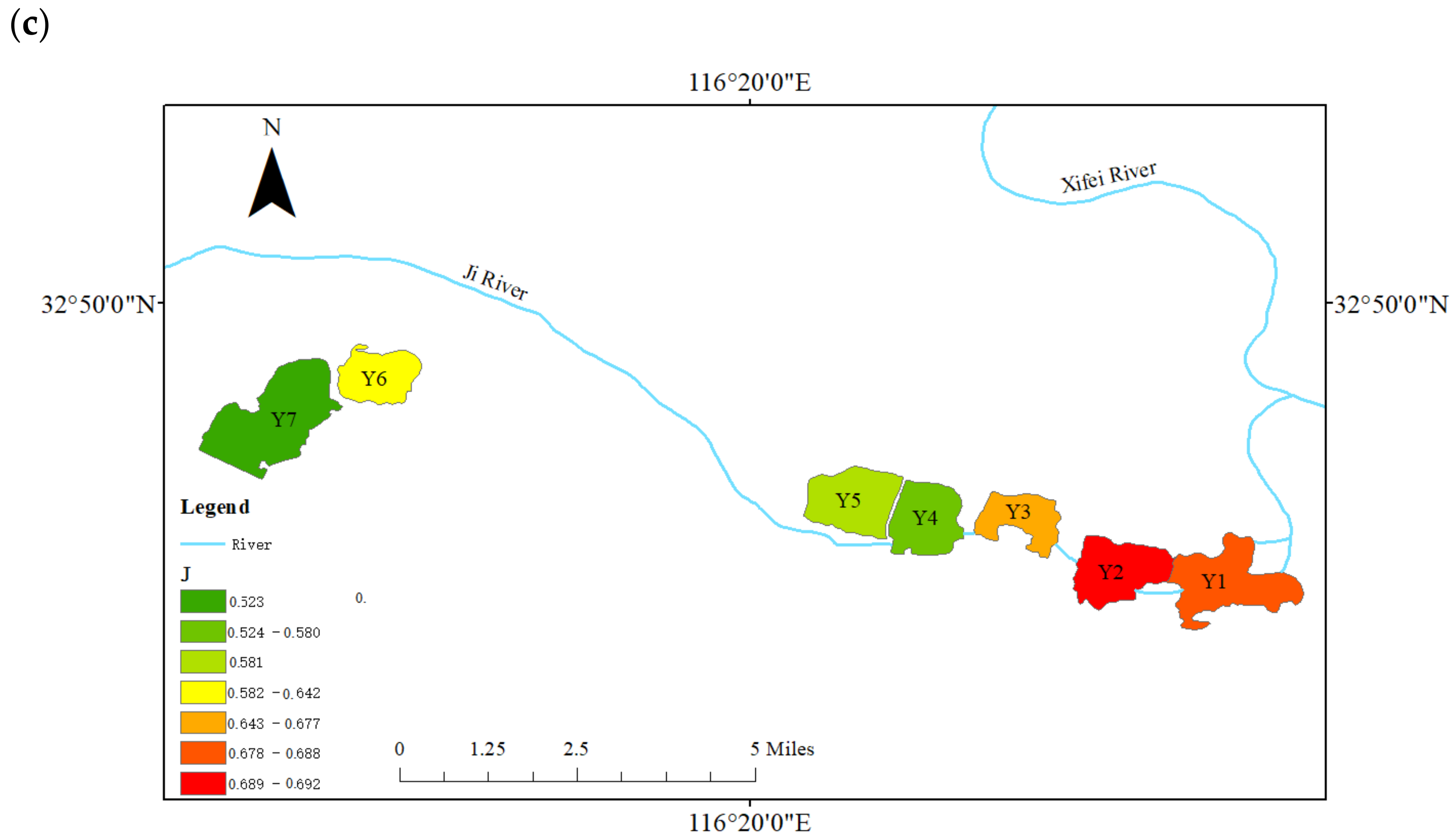
3.4. Spatial Distribution of Phytoplankton
4. Discussion
4.1. General Evaluation of Water Quality
4.2. Effects of Environmental Factors on Phytoplankton Diversity and Cell Density
4.3. Effects of Stream Connectivity on Phytoplankton Communities
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Fu, Y.; He, Z.; Xi, W. Mining subsidence wetland and its ecological management in coal mine area with high diving level. Wetl. Sci. 2016, 5, 6. [Google Scholar]
- Custodio, E. Aquifer overexploitation: What does it mean? Hydrogeol. J. 2002, 10, 254–277. [Google Scholar] [CrossRef]
- Zhou, J.; Zhao, S.; Chai, B. Ecological Environment Management Model and Key Technologies of Coal Mining Subsidence Area in Huang-Huai-Hai Plain; China University of Geosciences Press: Wuhan, China, 2015. [Google Scholar]
- Zhang, B.; Yan, J.; Fan, T. Evaluation and analysis of water eutrophication in coalmine subsidence area. Coal Technol. 2012, 31, 159–161. [Google Scholar]
- Chen, X.; Gao, L.; Zhuo, L. Temporal and spatial distribution of nitrogen and phosphorus of Xieqiao coal mining subsided water area. Environ. Chem. 2013, 32, 446–450. [Google Scholar]
- Chen, J. Analysis and Evaluation of Heavy Metal Pollution in Water Bodies in Coal Mining Subsidence Area of Pan Coal Mine, Huainan, Anhui Province; Nanjing University: Nanjing, China, 2017. [Google Scholar]
- Lin, Z.; Wan, Y.; Xu, M.; Wang, C.; Wu, Q.; Zhou, Z. Metazoan zooplankton community structure and environmental drivers of Digou coal mining subsided lakes in Huainan. J. Lake Sci. 2018, 301, 171–182. [Google Scholar]
- Wang, L.; Wang, C.; Deng, D.; Zhao, X.; Zhou, Z. Temporal and spatial variations in phytoplankton: Correlations with environmental factors in Shengjin Lake, China. Environ. Sci. Pollut. Res. 2015, 22, 14144–14156. [Google Scholar] [CrossRef]
- Irigoien, X.; Harris, R.P.; Verheye, H.M. Copepod hatching success in marine ecosystems with high diatom concentrations. Nature 2002, 419, 387–389. [Google Scholar] [CrossRef]
- Cardinale, B.J. Biodiversity improves water quality through niche partitioning. Nature 2011, 472, 86–89. [Google Scholar] [CrossRef]
- Wang, C.; Wu, Q.; Wan, Y.; Lin, Z.; Xu, J.; Zhou, Z. Phytoplankton community structure of coal mining subsided area with environmental drivers in Linhuan Lake, Huaibei. J. Biol. 2019, 36, 37–43. [Google Scholar]
- Wan, Y.; Zhou, Z.; Wang, C.; Lin, Z.; Wu, Q. Community structure characteristics of phytoplankton in Digou coal mining subsidence lake. J. Biol. 2018, 35, 75–82. [Google Scholar]
- Arrigo, K.R. Erratum: Marine microorganisms and global nutrient cycles. Nature 2005, 437, 349–355. [Google Scholar] [CrossRef]
- Gilstad, M.; Sakshaug, E. Growth rates of ten diatom species from the Barents Sea at different irradiances and day lengths. Mar. Ecol. Prog. Ser. 1990, 64, 169–173. [Google Scholar] [CrossRef]
- Maguer, J.F.; l’Helguen, S.; Caradec, J.; Klein, C. Size-dependent uptake of nitrate and ammonium as a function of light in well-mixed temperate coastal waters. Cont. Shelf Res. 2011, 31, 1620–1631. [Google Scholar] [CrossRef] [Green Version]
- Zhang, O.; Bu, H.; Wang, C. The influence of water system connectivity of Yangtze River for river health. Yangtze River 2010, 41, 1–5. [Google Scholar]
- Junk, W. The Flood Pulse Concept in river-floodplain Systems. Can. J. Spec. Publ. Fish. Aquat. Sci. 1989, 106, 110–127. [Google Scholar]
- Gumiero, B.; Mant, J.; Hein, T. Linking the restoration of rivers and riparian zones/wetlands in Europe: Sharing knowledge through case studies. Ecol. Eng. 2013, 56, 36–50. [Google Scholar] [CrossRef]
- Hu, H.J.; Wei, Y.X. The Freshwater Algae of China: Systematics, Taxonomy and Ecology; Science Press: Beijing, China, 2006. [Google Scholar]
- State Environmental Protection Bureau (SEPB). Methods of Monitoring and Analysis for Water and Wastewater, 4th ed.; China Environmental Science Press: Beijing, China, 2002. [Google Scholar]
- Jin, X.C.; Tu, Q.Y. The Standard Methods for Observation and Analysis of Lake Eutrophication; China Environmental Science Press: Beijing, China, 1990. [Google Scholar]
- Shannon, C.E.; Wiener, W. The Mathematical Theory of Communication; University of Illinois Press: Urbana, IL, USA, 1949. [Google Scholar]
- Pielou, E.C. An Introduction to Mathematical Ecology; Wiley-Interscience, Inc.: New York, NY, USA, 1969. [Google Scholar]
- Margalef, R. Information theory in ecology. Gen. Syst. 1958, 3, 36–71. [Google Scholar]
- Wu, C.; Zhang, Q.G. Progress of freshwater plankton diversity and quantitative analysis methods. Anhui Agric. Bull. 2009, 15, 41–42. [Google Scholar]
- Liu, Y. Research on Coordinated Utilization of Water and Soil Resources in Coal Mine Area with High Diving Position; China University of Mining and Technology: Beijing, China, 2014. [Google Scholar]
- Jin, X.C.; Tu, Q.Y. Lake Eutrophication Investigation Specification; China Environmental Science Press: Beijing, China, 1990. [Google Scholar]
- Li, Y.; Geng, M.; Yu, J.; Du, Y.; Xu, M.; Zhang, W.; Chen, F. Eutrophication decrease compositional dissimilarity in freshwater plankton communities. Sci. Total Environ. 2022, 821, 153434. [Google Scholar] [CrossRef]
- Chao, C.; Lv, T.; Wang, L.; Li, Y.; Han, C.; Yu, W.; Liu, C. The spatiotemporal characteristics of water quality and phytoplankton community in a shallow eutrophic lake: Implications for submerged vegetation restoration. Sci. Total Environ. 2022, 821, 153460. [Google Scholar] [CrossRef]
- Wang, Y. Characteristics of Phytoplankton Community Structure in Dongting Lake under Different Landscape Connectivity; Hubei University of Technology: Wuhan, China, 2021. [Google Scholar]
- Dieter, L.; Andrew, F.; Brigitte, N. Experimental eutrophication of a shallow acidic mining lake and effects on the phytoplankton. Hydrobiologia 2003, 506, 753–758. [Google Scholar]
- Li, Y.; Shen, R.; Liu, X.; Su, H.; Wu, F.; Chen, F. Impacts of nutrient reduction on temporal β-diversity of rotifers: A 19-year limnology case study on Lake Wuli, China. Water Res. J. Int. Water Assoc. 2022, 216, 1–14. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Lu, Y.; He, G.; Han, J.; Wang, T. Multivariate analysis of interactions between phytoplankton biomass and environmental variables in Taihu Lake, China. Environ. Monit. Assess. 2007, 133, 243–253. [Google Scholar] [CrossRef] [PubMed]
- Hinder, S.L.; Hays, G.C.; Edwards, M.; Roberts, E.C.; Walne, A.W.; Gravenor, M.B. Changes in marine dinoflagellate and diatom abundance under climate change. Nat. Clim. Change 2012, 2, 271–275. [Google Scholar] [CrossRef]
- Ma, J.; Qin, B.; Wu, P.; Zhou, J.; Niu, C.; Deng, J.; Niu, H. Controlling cyanobacterial blooms by managing nutrient ratio and limitation in a large hyper-eutrophic lake: Lake Taihu, China. J. Environ. Sci. 2015, 27, 80–86. [Google Scholar] [CrossRef]
- Smith, W.O.; Ainley, D.G.; Cattaneo-Vietti, R. Trophic interactions within the Ross Sea continental shelf ecosystem. Philos. Trans. R. Soc. B Biol. Sci. 2007, 362, 95–111. [Google Scholar] [CrossRef]
- Jiang, Y.; He, W.; Liu, W.; Qin, N.; Ouyang, H.; Wang, Q.; Kong, X.; He, Q.; Yang, C.; Yang, B.; et al. The seasonal and spatial variations of phytoplankton community and their correlation with environmental factors in a large eutrophic Chinese lake (Lake Chaohu). Ecol. Indic. 2014, 40, 58–67. [Google Scholar] [CrossRef]
- Howarth, R.W.; Marino, R. Nitrogen as the limiting nutrient for eutrophication in coastal marine ecosystems: Evolving views over three decades. Limnol. Oceanogr. 2006, 51, 364–376. [Google Scholar] [CrossRef] [Green Version]
- Chen, K.; Bao, X.; Shi, L. Reconstruction demonstration project of Wuli Lake in Taihu Lake (large-scale enclosure experiment). J. Lake Sci. 2006, 18, 139–149. [Google Scholar]
- Paul, W.J.; Hamilton, D.P.; Ostrovsky, I.; Miller, S.D.; Zhang, A.; Muraoka, K. Catchment land use and trophic state impacts on phytoplankton composition: A case study from the Rotorua lakes’ district, New Zealand. Hydrobiologia 2012, 698, 133–146. [Google Scholar] [CrossRef]
- Mitrovic, S.M.; Chessman, B.C.; Davie, A.; Avery, E.L.; Ryan, N. Development of blooms of Cyclotella meneghiniana and Nitzschia spp. (Bacillariophyceae) in a shallow river and estimation of effective suppression flows. Hydrobiologia 2008, 596, 173–185. [Google Scholar] [CrossRef]
- Zhang, S.; Li, C.; Fu, Y. Nutrient status and nutrient output of tributary bay of Three Gorges Reservoir. Environ. Sci. 2008, 29, 6. [Google Scholar]
- Wang, Y. Effect mechanism of hydrodynamic forces on algae. J. Anhui Agric. Sci. 2013, 41, 2. [Google Scholar]
- Xu, W.W.; Hu, W.P.; Deng, J.C.; Zhu, J.G.; Li, Q.Q. Effects of harvest management of Trapa bispinosa on an aquatic macrophyte community and water quality in a eutrophic lake. Ecol. Eng. 2014, 64, 120–129. [Google Scholar] [CrossRef]
- Mulderij, G.; Nes, E.H.V.; Donk, E.V. Macrophyte-phytoplankton interactions: The relative importance of allelopathy versus other factors. Ecol. Model. 2007, 204, 85–92. [Google Scholar] [CrossRef]
- Felpeto, A.B.; Roy, S.; Vasconcelos, V.M. Allelopathy prevents competitive exclusion and promotes phytoplankton biodiversity. Oikos J. Ecol. 2017, 127, 85–98. [Google Scholar] [CrossRef] [Green Version]
- Chia, A.M.; Iortsuun, D.N.; Stephen, B.J.; Ayobamire, A.E.; Ladan, Z. Phytoplankton responses to changes in macrophyte density in a tropical artificial pond in Zaria, Nigeria. Afr. J. Aquat. Sci. 2011, 36, 35–46. [Google Scholar] [CrossRef]
- Wang, Y.; Wang, W.; Zhou, Z.; Xia, W.; Zhang, Y. Effect of fast restoration of aquatic vegetation on phytoplankton community after removal of purse seine culture in huayanghe lakes. Sci. Total Environ. 2021, 768, 144024. [Google Scholar] [CrossRef]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Y1 | Y2 | Y3 | Y4 | Y5 | Y6 | Y7 | |
|---|---|---|---|---|---|---|---|
| WT/℃ | 27.58 ± 0.20 | 26.75 ± 0.22 | 26.75 ± 0.46 | 26.60 ± 0.11 | 26.85 ± 0.23 | 26.15 ± 0.63 | 26.60 ± 0.16 |
| pH | 7.57 ± 0.10 | 7.49 ± 0.09 | 7.57 ± 0.12 | 7.84 ± 0.04 | 7.94 ± 0.23 | 8.39 ± 0.21 | 8.17 ± 0.12 |
| DO/(mg/L) | 6.43 ± 1.03 | 5.17 ± 0.91 | 6.41 ± 0.86 | 5.68 ± 0.62 | 5.19 ± 0.57 | 4.67 ± 1.05 | 6.04 ± 0.77 |
| Cond/(Μs/cm) | 745.75 ± 2.59 | 768.75 ± 11.16 | 761.50 ± 11.59 | 742.14 ± 0.99 | 710.25 ± 3.19 | 717.50 ± 12.06 | 767.63 ± 1.80 |
| SD/cm | 53.13 ± 14.99 | 73.88 ± 11.90 | 58.75 ± 10.46 | 51.29 ± 7.21 | 52.63 ± 9.42 | 62.13 ± 7.93 | 55.25 ± 7.45 |
| WD/m | 5.23 ± 1.58 | 4.23 ± 1.46 | 5.58 ± 1.06 | 4.03 ± 2.37 | 5.18 ± 2.58 | 5.13 ± 2.36 | 4.65 ± 2.58 |
| Sdep/m | 2.73 ± 2.50 | 2.24 ± 2.01 | 2.83 ± 2.44 | 1.56 ± 1.60 | 2.64 ± 2.85 | 2.65 ± 2.68 | 2.38 ± 2.58 |
| NO/(mg/L) | 0.93 ± 0.10 | 1.50 ± 0.17 | 1.52 ± 0.16 | 0.62 ± 0.11 | 0.14 ± 0.02 | 0.07 ± 0.05 | 0.18 ± 0.02 |
| TP/(mg/L) | 0.24 ± 0.02 | 0.32 ± 0.10 | 0.21 ± 0.07 | 0.20 ± 0.04 | 0.27 ± 0.04 | 0.45 ± 0.21 | 0.67 ± 0.07 |
| TN/(mg/L) | 0.41 ± 0.47 | 1.62 ± 0.20 | 1.89 ± 0.48 | 1.47 ± 0.19 | 1.22 ± 0.27 | 1.07 ± 0.76 | 1.42 ± 0.43 |
| AN/(mg/L) | 0.68 ± 0.07 | 0.20 ± 0.02 | 0.21 ± 0.11 | 0.20 ± 0.02 | 0.30 ± 0.05 | 1.15 ± 0.81 | 0.57 ± 0.04 |
| Phylum | Dominant Species | Dominance of Phytoplankton Species (Y) | Proportion of Densities of Different Species (%) |
|---|---|---|---|
| Cyanobacteria | Leptolyngbya tenuis | 0.881 | 40.90 |
| Chroococcus minor | 0.023 | 1.49 | |
| Merismopedia sinica | 0.243 | 2.94 | |
| Chlorophyta | Auxenochlorella pyrenoidosa | 0.426 | 3.21 |
| Crucigenia quadrata | 0.051 | 4.45 | |
| Cryptophyta | Cryptomonas ovata | 0.133 | 1.80 |
| Parameter | Cell Density |
|---|---|
| r | |
| WT | −0.412 ** |
| pH | 0.554 ** |
| DO | −0.168 |
| Cond | −0.351 ** |
| SD | −0.255 |
| WD | −0.011 |
| Sdep | 0.036 |
| NO | −0.713 ** |
| TP | 0.335 * |
| TN | −0.087 |
| AN | 0.489 ** |
| Axes | 1 | 2 | 3 | 4 | Total Variance |
|---|---|---|---|---|---|
| Eigenvalues | 0.4603 | 0.0611 | 0.0243 | 0.0164 | 1 |
| Species–environment correlations | 46.03 | 52.14 | 54.57 | 56.22 | |
| Cumulative percentage variance of species data | 0.8926 | 0.5399 | 0.6520 | 0.5346 | |
| Cumulative percentage variance of species–environment relationship | 80.11 | 90.75 | 94.98 | 97.84 | |
| Sum of all eigenvalues | 1 |
| Axes | 1 | 2 | 3 | 4 | Total Variance |
|---|---|---|---|---|---|
| Eigenvalues | 0.3910 | 0.1086 | 0.0579 | 0.0280 | 1 |
| Species–environment correlations | 39.10 | 49.96 | 55.75 | 58.55 | |
| Cumulative percentage variance of species data | 0.8691 | 0.8250 | 0.6604 | 0.5387 | |
| Cumulative percentage variance of species–environment relationship | 63.94 | 81.69 | 91.16 | 95.73 | |
| Sum of all eigenvalues | 1 |
| Axes | 1 | 2 | 3 | 4 | Total Variance |
|---|---|---|---|---|---|
| Eigenvalues | 0.3170 | 0.1216 | 0.0757 | 0.0563 | 1 |
| Species–environment correlations | 31.70 | 43.85 | 51.43 | 57.06 | |
| Cumulative percentage variance of species data | 0.9328 | 0.7733 | 0.5222 | 0.8531 | |
| Cumulative percentage variance of species–environment relationship | 52.52 | 72.66 | 85.21 | 94.54 | |
| Sum of all eigenvalues | 1 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Jiang, L.; Yao, Y.; Zhang, S.; Wan, L.; Zhou, Z. Effects of Stream Connectivity on Phytoplankton Diversity and Community Structure in Sunken Lakes: A Case Study from an August Survey. Diversity 2023, 15, 291. https://doi.org/10.3390/d15020291
Jiang L, Yao Y, Zhang S, Wan L, Zhou Z. Effects of Stream Connectivity on Phytoplankton Diversity and Community Structure in Sunken Lakes: A Case Study from an August Survey. Diversity. 2023; 15(2):291. https://doi.org/10.3390/d15020291
Chicago/Turabian StyleJiang, Lingli, Yuping Yao, Siyong Zhang, Linqiang Wan, and Zhongze Zhou. 2023. "Effects of Stream Connectivity on Phytoplankton Diversity and Community Structure in Sunken Lakes: A Case Study from an August Survey" Diversity 15, no. 2: 291. https://doi.org/10.3390/d15020291