
Long-Term Succession on Offshore Wind Farms and the Role of Species Interactions

 ,
,  ,
,
Abstract
:1. Introduction
2. Materials and Methods
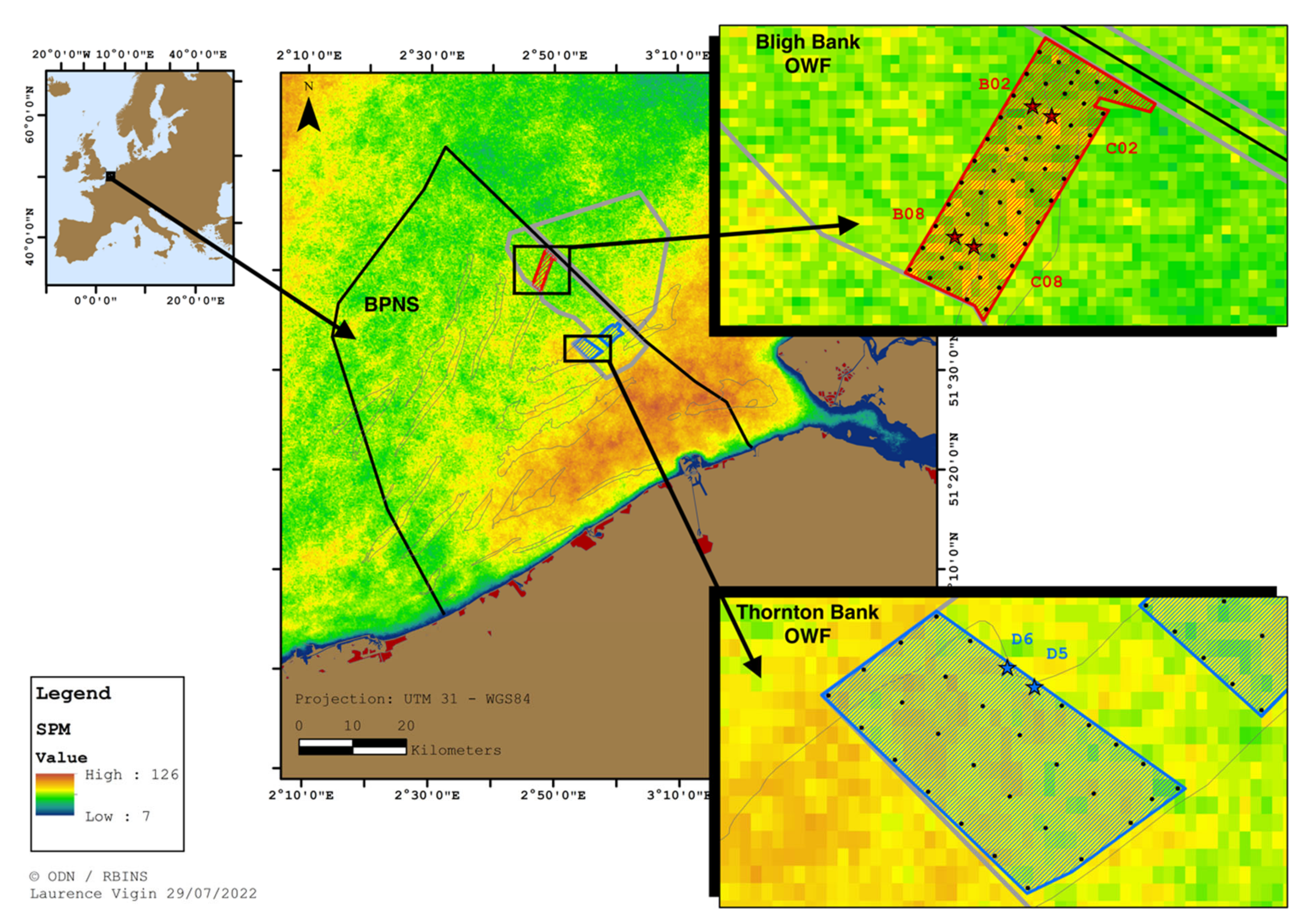
2.1. Study Site
2.2. The Data Set
2.3. Data Analysis
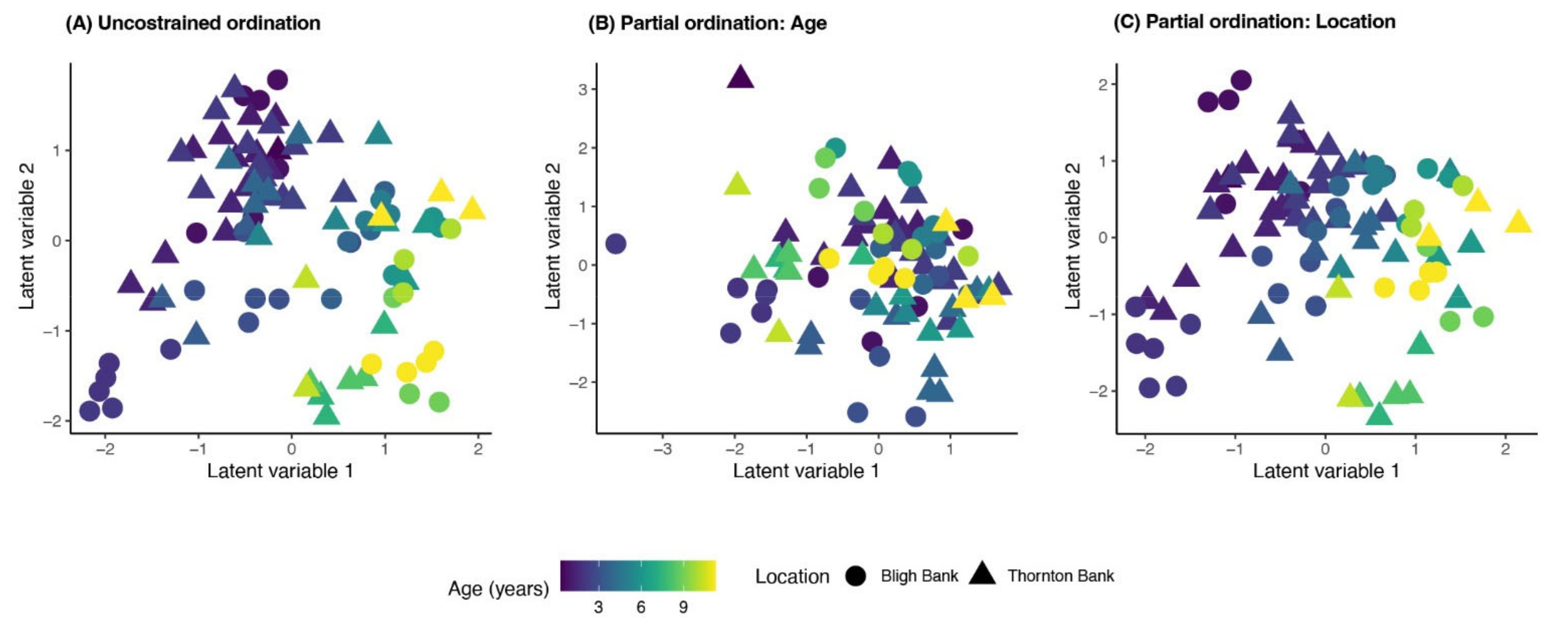
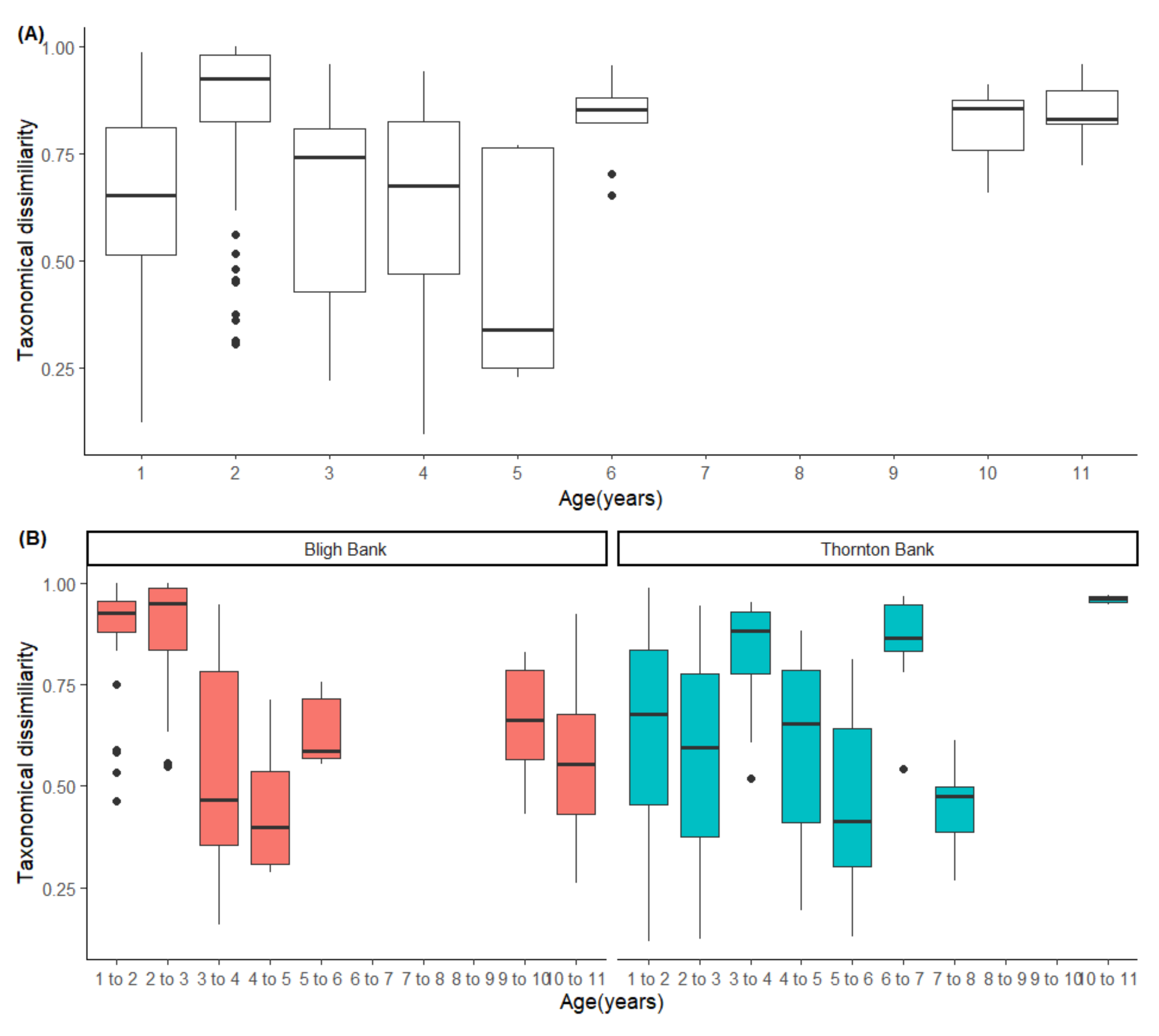
2.3.1. Multivariate Response Changes in Community Composition
Temporal Trends in Community Dynamics
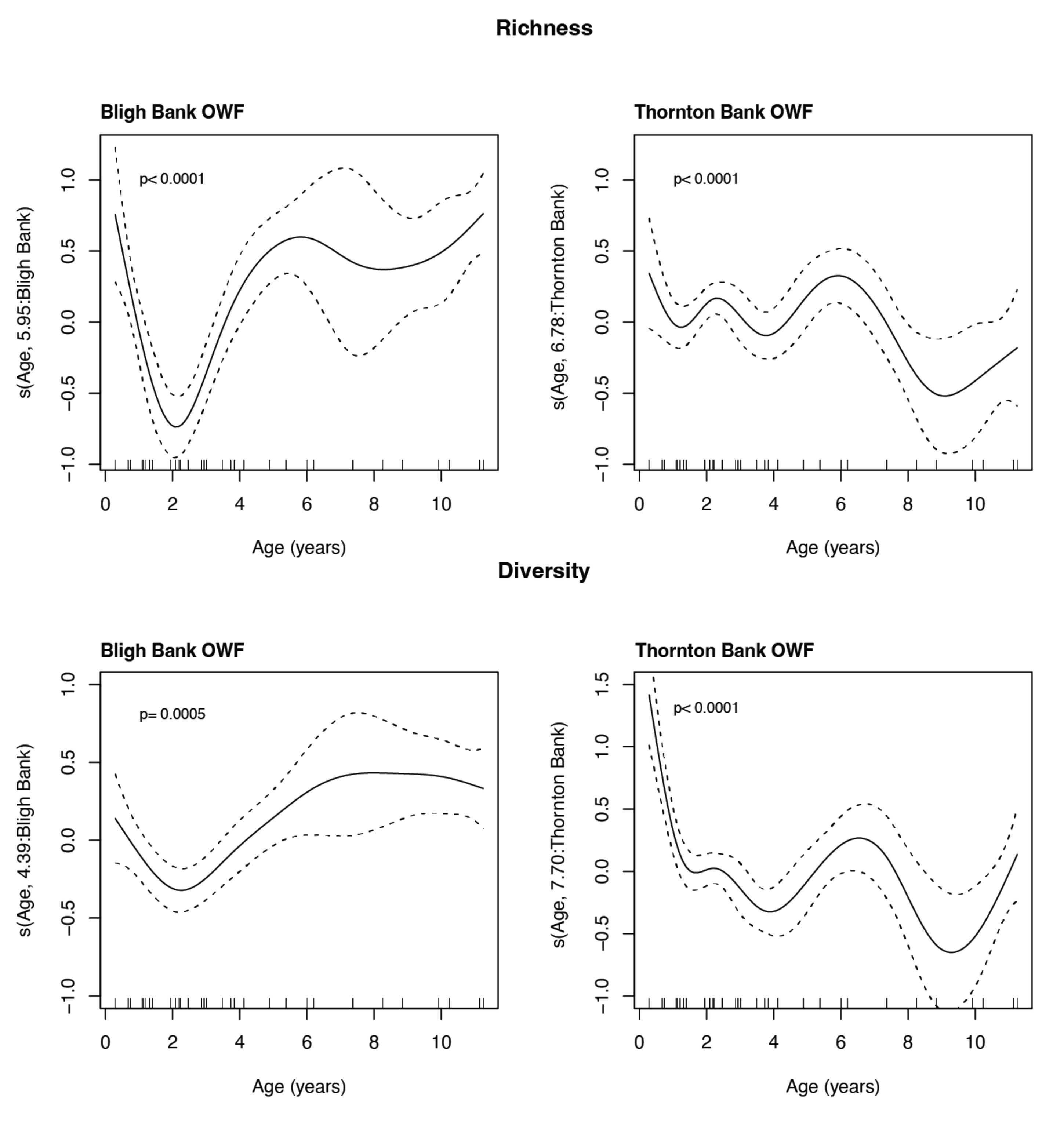
2.3.2. Univariate Response: Diversity Indices
- A full model with the sequential removal of each non-significant variable, until only significant variables were retained.
- A model with only environmental explanatory variables.
- A model with only biological explanatory variables.
- A model with only the abundance of the predator species (A. rubens and P. miliaris) as the explanatory variable.
- A model with only the abundance of the foundation species M. edulis as the explanatory variable.
- A model with only the abundance of the space occupiers (M. senile and amphipods).
3. Results
3.1. Biofouling Community Composition
3.2. Temporal Trends in Community Dynamics
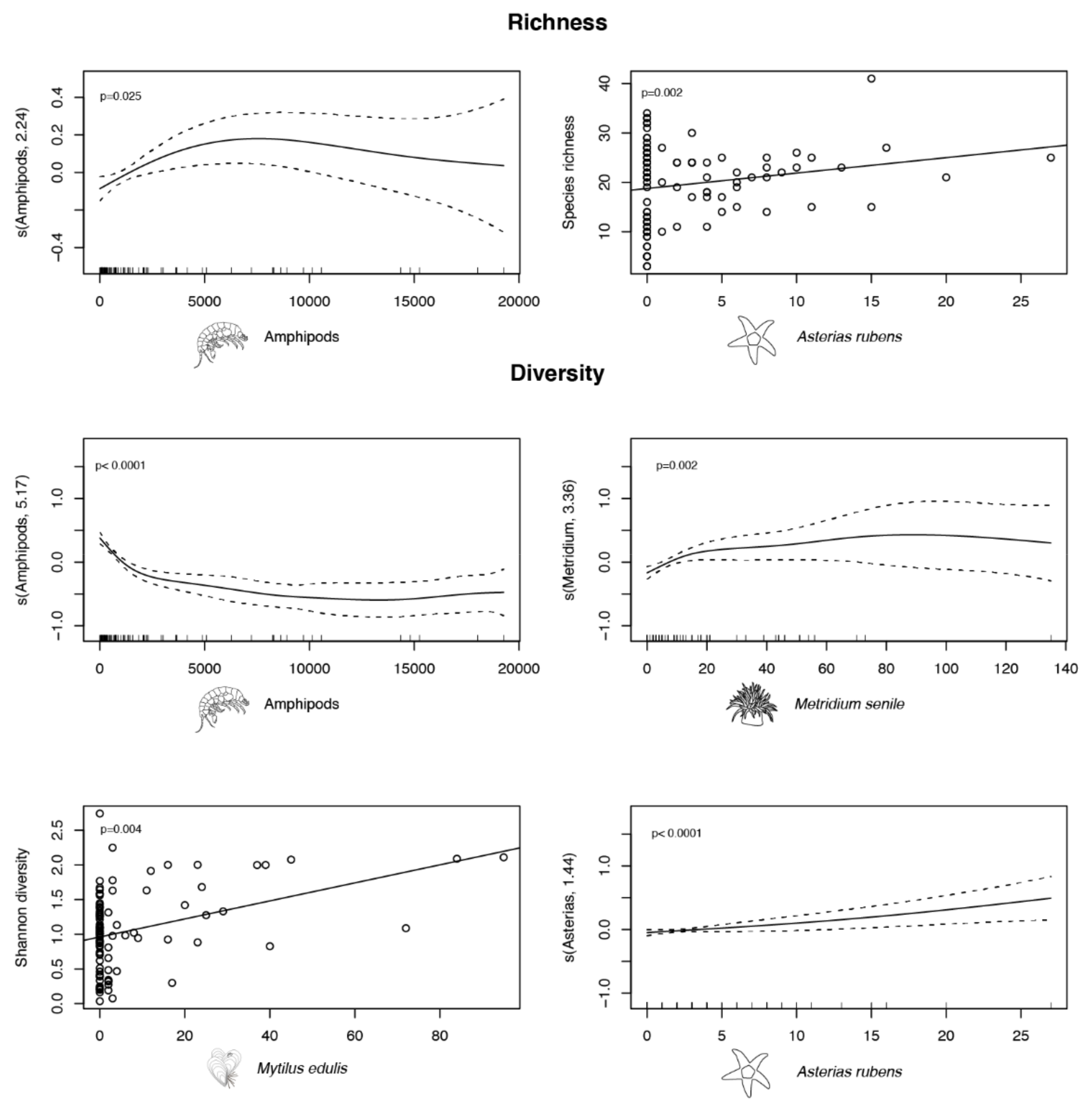
3.3. Univariate Response
4. Discussion
4.1. Local Differences in Biofouling Communities
4.2. Temporal Trends of Influential Species and Their Effects on Diversity
4.3. Divergence in Community Composition and No Apparent Stable State
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- North Seas Energy Cooperation. In Proceedings of the Joint Statement of North Seas Countries and the European Commission, Online, 6 July 2020; Available online: https://ec.europa.eu/info/news/progress-north-seas-energy-cooperation-2020-jul-06_en (accessed on 20 July 2022).
- GWEC Global Wind Energy Council. Global Offshore Wind Report; GWEC: Brussels, Belgium, 2021. [Google Scholar]
- Lindeboom, H.J.; Kouwenhoven, H.J.; Bergman, M.J.N.; Bouma, S.; Brasseur, S.; Daan, R.; Fijn, R.C.; De Haan, D.; Dirksen, S.; Van Hal, R.; et al. Short-term ecological effects of an offshore wind farm in the Dutch coastal zone; A compilation. Environ. Res. Lett. 2011, 6, 035101. [Google Scholar] [CrossRef] [Green Version]
- Vanagt, T.; Van de Moortel, L.; Faasse, M.A. Development of hard substrate fauna in the Princess Amalia Wind Farm. Monitoring 3.5 years after construction. eCoast Rep. 2013. Available online: https://core.ac.uk/download/pdf/80852871.pdf (accessed on 20 July 2022).
- Schröder, A.; Orejas, C.; Joschko, T. Benthos in the Vicinity of Piles: FINO 1 (North Sea). In Offshore Wind Energy: Research on Environmental Impacts; Köller, J., Köppel, J., Peters, W., Eds.; Springer: Berlin/Heidelberg, Germany, 2006; pp. 185–200. ISBN 978-3-540-34677-7. [Google Scholar]
- Coates, D.A.; Deschutter, Y.; Vincx, M.; Vanaverbeke, J. Enrichment and shifts in macrobenthic assemblages in an offshore wind farm area in the Belgian part of the North Sea. Mar. Environ. Res. 2014, 95, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Reubens, J.T.; Degraer, S.; Vincx, M. The ecology of benthopelagic fishes at offshore wind farms: A synthesis of 4 years of research. Hydrobiologia 2014, 727, 121–136. [Google Scholar] [CrossRef]
- Mavraki, N.; Degraer, S.; Vanaverbeke, J. Offshore wind farms and the attraction–production hypothesis: Insights from a combination of stomach content and stable isotope analyses. Hydrobiologia 2021, 848, 1639–1657. [Google Scholar] [CrossRef]
- Petersen, J.K.; Malm, T. Offshore Windmill Farms: Threats to or Possibilities for the Marine Environment. Ambio 2006, 35, 75–80. [Google Scholar] [CrossRef]
- Degraer, S.; Carey, D.A.; Coolen, J.W.P.; Hutchison, Z.L.; Kerckhof, F.; Rumes, B.; Vanaverbeke, J. Offshore wind farm artificial reefs affect ecosystem structure and functioning: A Synthesis. Oceanography 2020, 33, 48–57. [Google Scholar] [CrossRef]
- Mavraki, N.; Degraer, S.; Vanaverbeke, J.; Braeckman, U. Organic matter assimilation by hard substrate fauna in an offshore wind farm area: A pulse-chase study. ICES J. Mar. Sci. 2020, 77, 2681–2693. [Google Scholar] [CrossRef]
- Voet, H.E.E.; Vlaminck, E.; Van Colen, C.; Bodé, S.; Boeckx, P.; Degraer, S.; Moens, T.; Vanaverbeke, J.; Braeckman, U. Organic Matter Processing in a [Simulated] Offshore Wind Farm Ecosystem in Current and Future Climate and Aquaculture Scenarios. Sci. Total Environ. 2023, 857, 159285. [Google Scholar] [CrossRef]
- Slavik, K.; Lemmen, C.; Zhang, W.; Kerimoglu, O.; Klingbeil, K.; Wirtz, K.W. The large-scale impact of offshore wind farm structures on pelagic primary productivity in the southern North Sea. Hydrobiologia 2019, 845, 35–53. [Google Scholar] [CrossRef] [Green Version]
- Mavraki, N.; De Mesel, I.; Degraer, S.; Moens, T.; Vanaverbeke, J. Resource Niches of Co-occurring Invertebrate Species at an Offshore Wind Turbine Indicate a Substantial Degree of Trophic Plasticity. Front. Mar. Sci. 2020, 7, 379. [Google Scholar] [CrossRef]
- Ivanov, E.; Capet, A.; Borger, E.D.; Degraer, S.; Delhez, E.; Soetaert, K.; Vanaverbeke, J.; Grégoire, M. Modelling of the offshore wind farm footprint on organic and mineral particle deposition flux. In Proceedings of the Copernicus Meetings, Online, 19–30 April 2021. [Google Scholar]
- De Borger, E.; Ivanov, E.; Capet, A.; Braeckman, U.; Vanaverbeke, J.; Grégoire, M.; Soetaert, K. Offshore Windfarm Footprint of Sediment Organic Matter Mineralization Processes. Front. Mar. Sci. 2021, 8, 632243. [Google Scholar] [CrossRef]
- Coolen, J.W.P.; Vanaverbeke, J.; Dannheim, J.; Garcia, C.; Birchenough, S.N.R.; Krone, R.; Beermann, J. Generalized changes of benthic communities after construction of wind farms in the southern North Sea. J. Environ. Manag. 2022, 315, 115173. [Google Scholar] [CrossRef]
- Clements, F.E. Plant Succession: An Analysis of the Development of Vegetation; Carnegie Institution of Washington: Washington, DC, USA, 1916; ISBN 978-0-598-48534-2. [Google Scholar]
- Hill, M.F.; Witman, J.D.; Caswell, H. Spatio-temporal variation in Markov chain models of subtidal community succession. Ecol. Lett. 2002, 5, 665–675. [Google Scholar] [CrossRef]
- Scheer, B.T. The development of marine fouling communities. Biol. Bull. 1945, 89, 103–121. [Google Scholar] [CrossRef]
- Sutherland, J.P. Multiple Stable Points in Natural Communities. Am. Nat. 1974, 108, 859–873. [Google Scholar] [CrossRef]
- Boyd, S.E.; Limpenny, D.S.; Rees, H.L.; Cooper, K.M.; Campbell, S. Preliminary observations of the effects of dredging intensity on the re-colonisation of dredged sediments off the southeast coast of England (Area 222). Estuar. Coast. Shelf Sci. 2003, 57, 209–223. [Google Scholar] [CrossRef]
- Whomersley, P.; Picken, G.B. Long-term dynamics of fouling communities found on offshore installations in the North Sea. J. Mar. Biol. Assoc. UK 2003, 83, 897–901. [Google Scholar] [CrossRef]
- Taormina, B.; Percheron, A.; Marzloff, M.P.; Caisey, X.; Quillien, N.; Lejart, M.; Desroy, N.; Dugornay, O.; Tancray, A.; Carlier, A. Succession in epibenthic communities on artificial reefs associated with marine renewable energy facilities within a tide-swept environment. ICES J. Mar. Sci. 2020, 77, 2656–2668. [Google Scholar] [CrossRef]
- Oshurkov, V.V. Succession and climax in some fouling communities. Biofouling 1992, 6, 1–12. [Google Scholar] [CrossRef]
- Sutherland, J.P.; Karlson, R.H. Development and Stability of the Fouling Community at Beaufort, North Carolina. Ecol. Monogr. 1977, 47, 425–446. [Google Scholar] [CrossRef]
- Roberts, D.; Rittschof, D.; Holm, E.; Schmidt, A.R. Factors influencing initial larval settlement: Temporal, spatial and surface molecular components. J. Exp. Mar. Biol. Ecol. 1991, 150, 203–221. [Google Scholar] [CrossRef]
- Fortič, A.; Mavrič, B.; Pitacco, V.; Lipej, L. Temporal changes of a fouling community: Colonization patterns of the benthic epifauna in the shallow northern Adriatic Sea. Reg. Stud. Mar. Sci. 2021, 45, 101818. [Google Scholar] [CrossRef]
- Nydam, M.; Stachowicz, J. Predator effects on fouling community development. Mar. Ecol. Prog. Ser. 2007, 337, 93–101. [Google Scholar] [CrossRef] [Green Version]
- Cifuentes, M.; Krueger, I.; Dumont, C.P.; Lenz, M.; Thiel, M. Does primary colonization or community structure determine the succession of fouling communities? J. Exp. Mar. Biol. Ecol. 2010, 395, 10–20. [Google Scholar] [CrossRef]
- Sandin, S.A.; McNamara, D.E. Spatial dynamics of benthic competition on coral reefs. Oecologia 2012, 168, 1079–1090. [Google Scholar] [CrossRef]
- Loureiro, T.G.; Peters, K.; Robinson, T.B. Light, shade and predation: Who wins and who loses in sessile fouling communities? Mar. Biodivers. 2021, 51, 94. [Google Scholar] [CrossRef]
- Dayton, P.K. Toward an understanding of community resilience and the potential effects of enrichments to the benthos at McMurdo Sound, Antarctica. In Proceedings of the Colloquium on Conservation Problems Allen Press, Lawrence, Kansas, 1 January 1972; pp. 81–96. [Google Scholar]
- Stachowicz, J.J. Mutualism, Facilitation, and the Structure of Ecological Communities. BioScience 2001, 51, 235. [Google Scholar] [CrossRef]
- Bruno, J.F.; Stachowicz, J.J.; Bertness, M.D. Inclusion of facilitation into ecological theory. Trends Ecol. Evol. 2003, 18, 119–125. [Google Scholar] [CrossRef]
- Yakovis, E.; Artemieva, A. Epibenthic predators control mobile macrofauna associated with a foundation species in a subarctic subtidal community. Ecol. Evol. 2019, 9, 10499–10512. [Google Scholar] [CrossRef] [Green Version]
- Gutiérrez, J.L.; Jones, C.G.; Strayer, D.L.; Iribarne, O.O. Mollusks as ecosystem engineers: The role of shell production in aquatic habitats. Oikos 2003, 101, 79–90. [Google Scholar] [CrossRef]
- Yakovis, E.; Artemieva, A. Cockles, barnacles and ascidians compose a subtidal facilitation cascade with multiple hierarchical levels of foundation species. Sci. Rep. 2017, 7, 237. [Google Scholar] [CrossRef] [Green Version]
- Searles, A.R.; Gipson, E.E.; Walters, L.J.; Geoffrey, S. Cook Oyster reef restoration facilitates the recovery of macroinvertebrate abundance, diversity, and composition in estuarine communities. Sci. Rep. 2022, 12, 8163. [Google Scholar] [CrossRef]
- Coolen, J.W.P.; van der Weide, B.; Cuperus, J.; Blomberg, M.; Van Moorsel, G.W.N.M.; Faasse, M.A.; Bos, O.G.; Degraer, S.; Lindeboom, H.J. Benthic biodiversity on old platforms, young wind farms, and rocky reefs. ICES J. Mar. Sci. 2020, 77, 1250–1265. [Google Scholar] [CrossRef]
- Mills, L.S.; Soulé, M.E.; Doak, D.F. The Keystone-Species Concept in Ecology and Conservation. BioScience 1993, 43, 219–224. [Google Scholar] [CrossRef] [Green Version]
- Paine, R.T. Food Web Complexity and Species Diversity. Am. Nat. 1966, 100, 65–75. [Google Scholar] [CrossRef]
- Menge, B.A.; Sutherland, J.P. Species Diversity Gradients: Synthesis of the Roles of Predation, Competition, and Temporal Heterogeneity. Am. Nat. 1976, 110, 351–369. [Google Scholar] [CrossRef]
- Sarnelle, O. Daphnia as keystone predators: Effects on phytoplankton diversity and grazing resistance. J. Plankton Res. 2005, 27, 1229–1238. [Google Scholar] [CrossRef]
- Navarrete, S.A.; Menge, B.A. Keystone Predation and Interaction Strength: Interactive Effects of Predators on Their Main Prey. Ecol. Monogr. 1996, 66, 409–429. [Google Scholar] [CrossRef]
- Mouquet, N.; Gravel, D.; Massol, F.; Calcagno, V. Extending the concept of keystone species to communities and ecosystems. Ecol. Lett. 2013, 16, 1–8. [Google Scholar] [CrossRef]
- Lewbel, G.S.; Howard, R.L.; Gallaway, B.J. Zonation of dominant fouling organisms on northern gulf of Mexico petroleum platforms. Mar. Environ. Res. 1987, 21, 199–224. [Google Scholar] [CrossRef]
- Venugopalan, V.P.; Wagh, A.B. Biofouling of an offshore oil platform: Faunal composition and biomass. Indian J. Mar. Sci. 1990, 19, 4. [Google Scholar]
- Richardson, C.A.; Seed, R. Predictions of mussel (Mytilus edulis) biomass on an offshore platform from single population samples. Biofouling 1990, 2, 289–297. [Google Scholar] [CrossRef]
- Sell, D. Marine Fouling. Proc. Royal Soc. Edinb. Sect. B Biol. Sci. 1992, 100, 169–184. [Google Scholar] [CrossRef]
- Relini, G.; Tixi, F.; Relini, M.; Torchia, G. The macrofouling on offshore platforms at Ravenna. Int. Biodeterior. Biodegrad. 1998, 41, 41–55. [Google Scholar] [CrossRef]
- Yan, T.; Yan, W.X. Fouling of Offshore Structures in China-a Review. Biofouling 2003, 19, 133–138. [Google Scholar] [CrossRef]
- Bram, J.B.; Page, H.M.; Dugan, J.E. Spatial and temporal variability in early successional patterns of an invertebrate assemblage at an offshore oil platform. J. Exp. Mar. Biol. Ecol. 2005, 317, 223–237. [Google Scholar] [CrossRef]
- Oricchio, F.; Dias, G. Predation and competition interact to determine space monopolization by non-indigenous species in a sessile community from the southwestern Atlantic Ocean. Aquat. Invasions 2020, 15, 127–139. [Google Scholar] [CrossRef]
- Fettweis, M.; Schartau, M.; Desmit, X.; Lee, B.J.; Terseleer, N.; Zande, D.V.d.; Parmentier, K.; Riethmüller, R. Organic Matter Composition of Biomineral Flocs and Its Influence on Suspended Particulate Matter Dynamics Along a Nearshore to Offshore Transect. J. Geophys. Res. Biogeosci. 2022, 172, e2021JG006332. [Google Scholar] [CrossRef]
- Baeye, M.; Fettweis, M. In situ observations of suspended particulate matter plumes at an offshore wind farm, southern North Sea. Geo. Mar. Lett. 2015, 35, 247–255. [Google Scholar] [CrossRef]
- De Mesel, I.; Kerckhof, F.; Norro, A.; Rumes, B.; Degraer, S. Succession and seasonal dynamics of the epifauna community on offshore wind farm foundations and their role as stepping stones for non-indigenous species. Hydrobiologia 2015, 756, 37–50. [Google Scholar] [CrossRef]
- Horton, T.; Kroh, A.; Ahyong, S.; Bailly, N.; Boyko, C.B.; Brandão, S.N.; Gofas, S.; Hooper, J.N.A.; Hernandez, F.; Holovachov, O.; et al. World Register of Marine Species (WoRMS); WoRMS Editorial Board: Oostende, Belgium, 2021. [Google Scholar]
- Wang, Y.; Naumann, U.; Wright, S.T.; Warton, D.I. Mvabund—An R package for model-based analysis of multivariate abundance data. Methods Ecol. Evol. 2012, 3, 471–474. [Google Scholar] [CrossRef]
- Niku, J.; Hui, F.K.C.; Taskinen, S.; Warton, D.I. Gllvm: Fast analysis of multivariate abundance data with generalized linear latent variable models in r. Methods Ecol. Evol. 2019, 10, 2173–2182. [Google Scholar] [CrossRef] [Green Version]
- Hui, F.K.C.; Taskinen, S.; Pledger, S.; Foster, S.D.; Warton, D.I. Model-based approaches to unconstrained ordination. Methods Ecol. Evol. 2015, 6, 399–411. [Google Scholar] [CrossRef]
- Bray, J.R.; Curtis, J.T. An Ordination of the Upland Forest Communities of Southern Wisconsin. Ecol. Monogr. 1957, 27, 326–349. [Google Scholar] [CrossRef]
- Oksanen, J.; Blanchet, F.G.; Friendly, M.; Kindt, R.; Legendre, P.; McGlinn, D.; Minchin, P.R.; O’Hara, R.B.; Simpson, G.; Solymos, P.; et al. Vegan: Community Ecology Package, R Package Version 2.5–7; University of Helsinki: Helsinki, Finland, 2020; Available online: https://CRAN.R-project.org/package=vegan (accessed on 15 June 2022).
- Buckley, H.L.; Day, N.J.; Case, B.S.; Lear, G.; Ellison, A.M. Multivariate methods for testing hypotheses of temporal community dynamics. bioRxiv 2019, 362822. [Google Scholar] [CrossRef] [Green Version]
- Zuur, A.F.; Ieno, E.N.; Elphick, C.S. A protocol for data exploration to avoid common statistical problems: Data exploration. Methods Ecol. Evol. 2010, 1, 3–14. [Google Scholar] [CrossRef]
- Fox, J.; Weisberg, S. An R Companion to Applied Regression, 3rd ed.; Sage: Thousand Oaks, CA, USA, 2019. [Google Scholar]
- Zuur, A.F.; Ieno, E.N.; Walker, N.; Saveliev, A.A.; Smith, G.M. Mixed Effects Models and Extensions in Ecology with R; Statistics for Biology and Health; Springer: New York, NY, USA, 2009; ISBN 978-0-387-87457-9. [Google Scholar]
- Wood, S.; Scheipl, F. Package ‘Gamm4′: Generalized Additive Mixed Models Using “mgcv” and “lme4”; CRAN: Online, 2020. [Google Scholar]
- Akaike, H. Information Theory and the Maximum Likelihood Principle in 2nd International Symposium on Information Theory; Petrov, B.N., Cs ä ki, F., Eds.; Akademiai Ki à do: Budapest, Hungary, 1973. [Google Scholar]
- Anderson, D.R. Model Based Inference in the Life Sciences: A Primer on Evidence; Springer: New York, NY, USA, 2008; ISBN 978-0-387-74073-7. [Google Scholar]
- Parsons, D.; Suthers, I.; Cruz, D.; Smith, J. Effects of habitat on fish abundance and species composition on temperate rocky reefs. Mar. Ecol. Prog. Ser. 2016, 561, 155–171. [Google Scholar] [CrossRef]
- R Core Team. R: A Language and Environment for Statistical Computing; R Core Team: Vienna, Austria. Available online: https://www.R-project.org/ (accessed on 15 June 2022).
- Zintzen, V.; Norro, A.; Massin, C.; Mallefet, J. Spatial variability of epifaunal communities from artificial habitat: Shipwrecks in the Southern Bight of the North Sea. Estuar. Coast. Shelf Sci. 2008, 76, 327–344. [Google Scholar] [CrossRef]
- Schutter, M.; Dorenbosch, M.; Driessen, F.M.F.; Lengkeek, W.; Bos, O.G.; Coolen, J.W.P. Oil and gas platforms as artificial substrates for epibenthic North Sea fauna: Effects of location and depth. J. Sea Res. 2019, 153, 101782. [Google Scholar] [CrossRef]
- Krone, R.; Gutow, L.; Joschko, T.J.; Schröder, A. Epifauna dynamics at an offshore foundation–Implications of future wind power farming in the North Sea. Mar. Environ. Res. 2013, 85, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Todd, V.L.G.; Susini, I.; Williamson, L.D.; Todd, I.B.; McLean, D.L.; Macreadie, P.I. Characterizing the second wave of fish and invertebrate colonization of an offshore petroleum platform. ICES J. Mar. Sci. 2021, 78, 1131–1145. [Google Scholar] [CrossRef]
- Agüera, A.; Saurel, C.; Møller, L.F.; Fitridge, I.; Petersen, J.K. Bioenergetics of the common seastar Asterias rubens: A keystone predator and pest for European bivalve culture. Mar. Biol. 2021, 168, 48. [Google Scholar] [CrossRef]
- Hardy, F.G. Fouling on North Sea Platforms. Bot. Mar. 1981, 24, 173–176. [Google Scholar] [CrossRef]
- Forteath, G.; Picken, G.; Ralph, R.; Williams, J. Marine Growth Studies on the North Sea Oil Platform Montrose Alpha. Mar. Ecol. Prog. Ser. 1982, 8, 61–68. [Google Scholar] [CrossRef]
- Wolfson, A.; Van Blaricom, G.; Davis, N.; Lewbel, G. The Marine Life of an Offshore Oil Platform. Mar. Ecol. Prog. Ser. 1979, 1, 81–89. [Google Scholar] [CrossRef]
- Southgate, T.; Myers, A.A. Mussel fouling on the Celtic Sea Kinsale Field gas platforms. Estuar. Coast. Shelf Sci. 1985, 20, 651–659. [Google Scholar] [CrossRef]
- Suchanek, T. Mussels and their role in structuring rocky shore communities. In The Ecology of Rocky Coasts; Hodder & Stoughton: London, UK, 1985; pp. 70–96. [Google Scholar]
- Lintas, C.; Seed, R. Spatial variation in the fauna associated with Mytilus edulis on a wave-exposed rocky shore. J. Molluscan Stud. 1994, 60, 165–174. [Google Scholar] [CrossRef]
- Ragnarsson, S.Á.; Raffaelli, D. Effects of the mussel Mytilus edulis L. on the invertebrate fauna of sediments. J. Exp. Mar. Biol. Ecol. 1999, 241, 31–43. [Google Scholar] [CrossRef]
- Drent, J.; Dekker, R. Macrofauna Associated with Mussels, Mytilus edulis L., in the subtidal of the western Dutch Wadden Sea; NIOZ, Royal Netherlands Institute for Sea Research: Texel, The Netherlands, 2013; p. 56. [Google Scholar]
- Tsuchiya, M.; Nishihira, M. Islands of Mytilus as a habitat for small intertidal animals: Effect of island size on community structure. Mar. Ecol. Prog. Ser. 1985, 25, 71–81. [Google Scholar] [CrossRef]
- Norling, P.; Kautsky, N. Structural and functional effects of Mytilus edulis on diversity of associated species and ecosystem functioning. Mar. Ecol. Prog. Ser. 2007, 351, 163–175. [Google Scholar] [CrossRef] [Green Version]
- Miner, C.M.; Burnaford, J.L.; Ammann, K.; Becker, B.H.; Fradkin, S.C.; Ostermann-Kelm, S.; Smith, J.R.; Whitaker, S.G.; Raimondi, P.T. Latitudinal variation in long-term stability of North American rocky intertidal communities. J. Anim. Ecol. 2021, 90, 2077–2093. [Google Scholar] [CrossRef]
- Nelson, M.; Craig, S. Role of the sea anemone Metridium senile in structuring a developing subtidal fouling community. Mar. Ecol. Prog. Ser. 2011, 421, 139–149. [Google Scholar] [CrossRef] [Green Version]
- Zintzen, V.; Massin, C.; Norro, A.; Mallefet, J. Epifaunal inventory of two shipwrecks from the Belgian Continental Shelf. In Marine Biodiversity: Patterns and Processes, Assessment, Threats, Management and Conservation; Developments in, Hydrobiology; Martens, K., Queiroga, H., Cunha, M.R., Cunha, A., Moreira, M.H., Quintino, V., Rodrigues, A.M., Seroôdio, J., Warwick, R.M., Eds.; Springer: Dordrecht, The Netherlands, 2006; pp. 207–219. ISBN 978-1-4020-4697-1. [Google Scholar]
- Coolen, J.W.P.; Bos, O.G.; Glorius, S.; Lengkeek, W.; Cuperus, J.; van der Weide, B.; Agüera, A. Reefs, sand and reef-like sand: A comparison of the benthic biodiversity of habitats in the Dutch Borkum Reef Grounds. J. Sea Res. 2015, 103, 84–92. [Google Scholar] [CrossRef]
- Van der Stap, T.; Coolen, J.W.P.; Lindeboom, H.J. Marine Fouling Assemblages on Offshore Gas Platforms in the Southern North Sea: Effects of Depth and Distance from Shore on Biodiversity. PLoS ONE 2016, 11, e0146324. [Google Scholar] [CrossRef] [Green Version]
- Ardizzone, G.D.; Gravina, M.F.; Belluscio, A. Temporal Development of Epibenthic Communities on Artificial Reefs in the Central Mediterranean Sea. Bull. Mar. Sci. 1989, 44, 592–608. [Google Scholar]
- Butler, A.; Connolly, R. Assemblages of sessile marine invertebrates:still changing after all these years? Mar. Ecol. Prog. Ser. 1999, 182, 109–118. [Google Scholar] [CrossRef] [Green Version]
- Hooper, T.; Beaumont, N.; Hattam, C. The implications of energy systems for ecosystem services: A detailed case study of offshore wind. Renew. Sustain. Energy Rev. 2017, 70, 230–241. [Google Scholar] [CrossRef] [Green Version]
- Fowler, A.M.; Jorgensen, A.-M.; Svendsen, J.C.; Macreadie, P.I.; Jones, D.O.B.; Boon, A.R.; Booth, D.J.; Brabant, R.; Callahan, E.; Claisse, J.T.; et al. Environmental benefits of leaving offshore infrastructure in the ocean. Front. Ecol. Environ. 2018, 16, 571–578. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Years Since Installation | 1 | 2 | 3 | 4 | 5 | 6 | 7 | 8 | 9 | 10 | 11 | Total | |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Bligh Bank | BB B2 | 3 | 3 | ||||||||||
| BB B8 | 3 | 3 | 2 | 3 | 3 | 4 | 18 | ||||||
| BB C2 | 3 | 3 | 4 | 4 | 3 | 17 | |||||||
| Thornton Bank | D5 | 16 | 11 | 3 | 2 | 3 | 3 | 3 | 3 | 44 | |||
| D6 | 3 | 3 | 3 | 2 | 11 | ||||||||
| Total | 22 | 20 | 9 | 9 | 6 | 6 | 3 | 3 | 3 | 5 | 7 | 93 |
| Parameter | Residual DF | DF | Deviance | p-Value | Species with Significant Effect |
|---|---|---|---|---|---|
| Countable Species (Abundance) | |||||
| Location | 91 | 1 | 352.3 | 0.001 | Heteranomia squamula (9.5%), Monocorophium acherusicum (7.8%), Spirobranchus triqueter (6.8%), Verruca stroemia (5.2%), Phyllodoce mucosa (5.1%), Alcyonium digitatum (4.5 %), Eulalia (4.3 %), and Amphipholis squamata (3.5%) |
| Age | 90 | 1 | 766.3 | 0.001 | Asterias rubens (6.7%), Psammechinus miliaris (6.1%), Monocorophium acherusicum (6%), Corophium sextonae (4.9%), Phyllodoce mucosa (4.1%), Verruca stroemia (3.8%), Nudibranchia (3.8%), Metridium senile (3.6%), Mytilus edulis (3.2%), Gitana sarsi (3.12%), Lanice conchilega (2.8%), Heteranomia squamula (2.7 %), Sabellaria spinulosa (2.6%), Syllis gracilis (2.4%), Pisidia longicornis (2.2%), Nototropis swammerdamei (1.8%), Pilumnus hirtellus (1.7%), Eunereis longissima (1.7%), and Pagurus bernhardus (1.6%) |
| Location:Age | 89 | 1 | 171.8 | 0.023 | Balanus perforatus (19.4%) |
| Model | Park | p | Age, BW | Age, CP | Asterias rubens | p | Psammechinus miliaris | p | Mytilus edulis | Metridium senile | p | Amphipods | AIC | ΔAIC | % Deviance |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Full | 0.11 | ns | *** | ns | 0.013 | * | −0.009 | ns | ns | 0.001 | ns | *** | 590 | 4 | 85.8 |
| Full simplified 1 | 0.21 | ns | *** | *** | 0.016 | * | −0.008 | ns | * | - | - | * | 590 | 4 | 85.3 |
| Full simplified 2 | 0.18 | ns | *** | *** | 0.019 | ** | - | - | ns | - | - | * | 589 | 3 | 85.3 |
| Full simplified 3 | - | - | *** | ** | 0.021 | ** | - | - | ns | - | - | * | 588 | 2 | 82.7 |
| Full simplified 4 | - | - | *** | ** | 0.021 | *** | - | - | - | - | - | ** | 586 | 0 | 82.5 |
| Environment | 0.20 | ns | *** | *** | - | - | - | - | - | - | - | 603 | 17 | 76.9 | |
| Biological | - | - | - | - | 0.026 | *** | −0.001 | ns | *** | 0.0024 | Ns | *** | 654 | 68 | 52.6 |
| Predators | - | - | - | - | 0.018 | *** | −0.016 | *** | - | - | - | - | 746 | 160 | 7 |
| Foundation | - | - | - | - | - | - | - | *** | - | - | - | 695 | 109 | 24.7 | |
| Space occupier | - | - | - | - | - | - | - | - | - | 0.004 | *** | *** | 714 | 125 | 29.9 |
| Model | Park | p | Age, BW | Age, CP | Asterias rubens | Psammechinus miliaris | Mytilus edulis | p | Metridium senile | Amphipods | AIC | ΔAIC | % Deviance |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Full | 0.013 | ns | *** | *** | ** | ns | 0.005 | * | * | *** | 102 | 12 | 92.2 |
| Full simplified 1 | - | - | *** | *** | ** | ns | 0.005 | * | * | *** | 97 | 7 | 93.8 |
| Full simplified 2 | - | - | *** | *** | * | - | 0.005 | * | * | *** | 90 | 0 | 91.7% |
| Environment | 0.005 | ns | *** | *** | - | - | - | - | - | 117 | 27 | 73 | |
| Biological | - | - | - | * | ns | 0.008 | *** | *** | *** | 117 | 27 | 73 | |
| Predators | - | - | - | * | ns | - | - | - | 177 | 88 | 16.8 | ||
| Foundation | - | - | - | - | *** | - | - | 166 | 76 | 15.1 | |||
| Space occupier | - | - | - | - | - | - | *** | *** | 148 | 58 | 66.5 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zupan, M.; Rumes, B.; Vanaverbeke, J.; Degraer, S.; Kerckhof, F. Long-Term Succession on Offshore Wind Farms and the Role of Species Interactions. Diversity 2023, 15, 288. https://doi.org/10.3390/d15020288
Zupan M, Rumes B, Vanaverbeke J, Degraer S, Kerckhof F. Long-Term Succession on Offshore Wind Farms and the Role of Species Interactions. Diversity. 2023; 15(2):288. https://doi.org/10.3390/d15020288
Chicago/Turabian StyleZupan, Mirta, Bob Rumes, Jan Vanaverbeke, Steven Degraer, and Francis Kerckhof. 2023. "Long-Term Succession on Offshore Wind Farms and the Role of Species Interactions" Diversity 15, no. 2: 288. https://doi.org/10.3390/d15020288