
Lichen and Lichenicolous Fungal Communities Tested as Suitable Systems for the Application of Cross-Taxon Analysis



Abstract
:1. Introduction
2. Materials and Methods
2.1. Data Collection
2.2. Analysis of Species Diversity Patterns at Different Spatial Scales
2.3. Congruence in Species Composition
3. Results
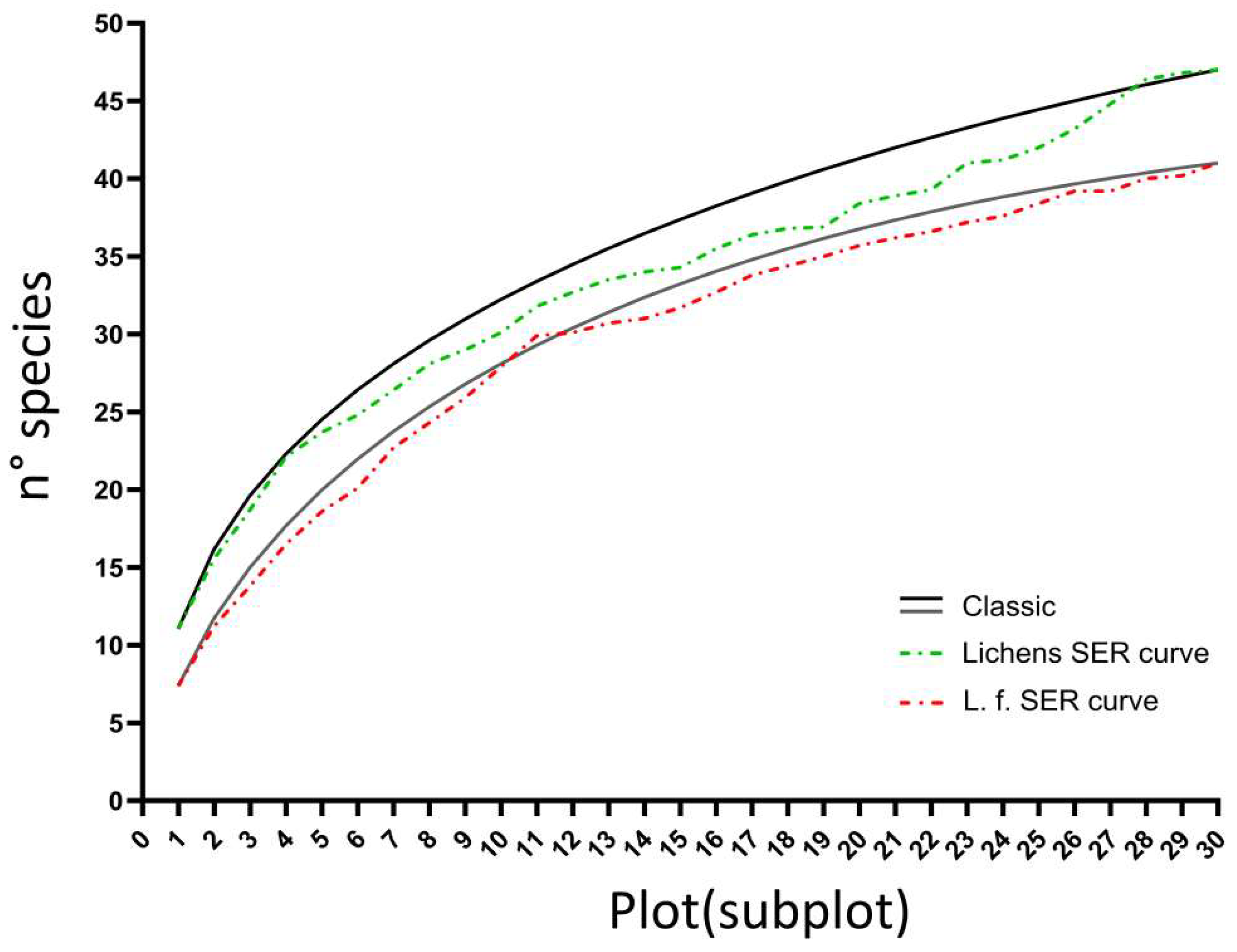
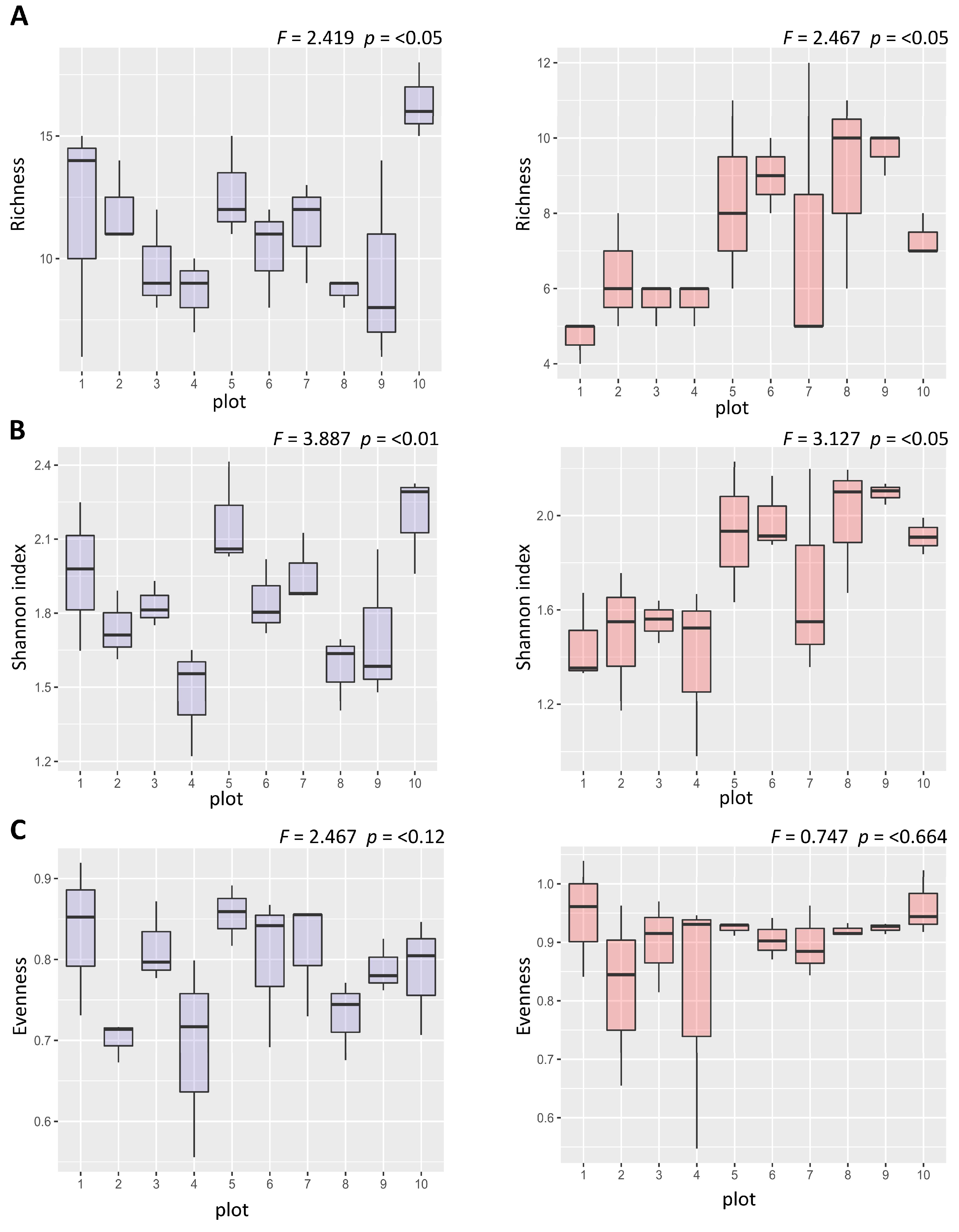
3.1. Diversity Patterns at Different Spatial Scales
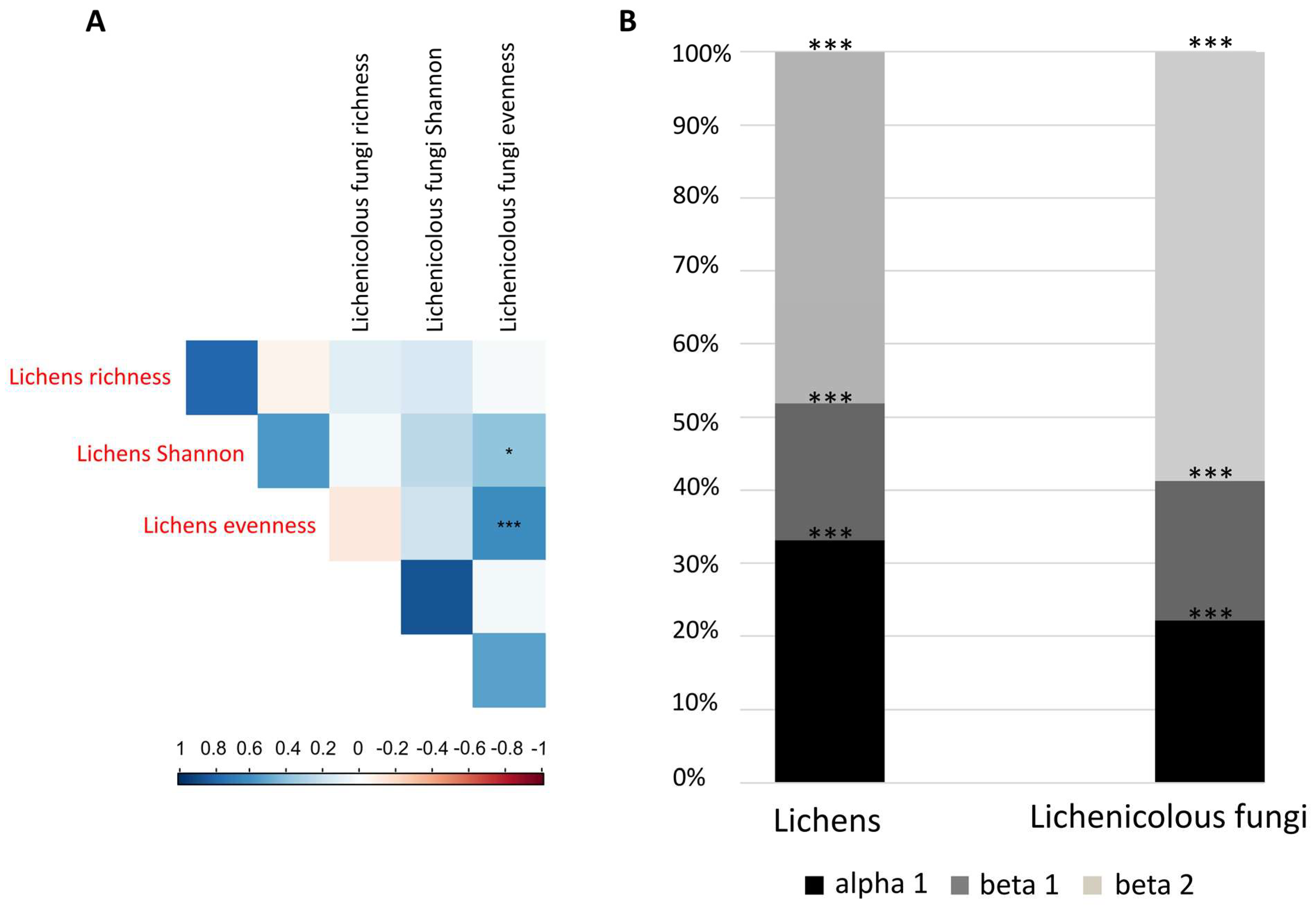
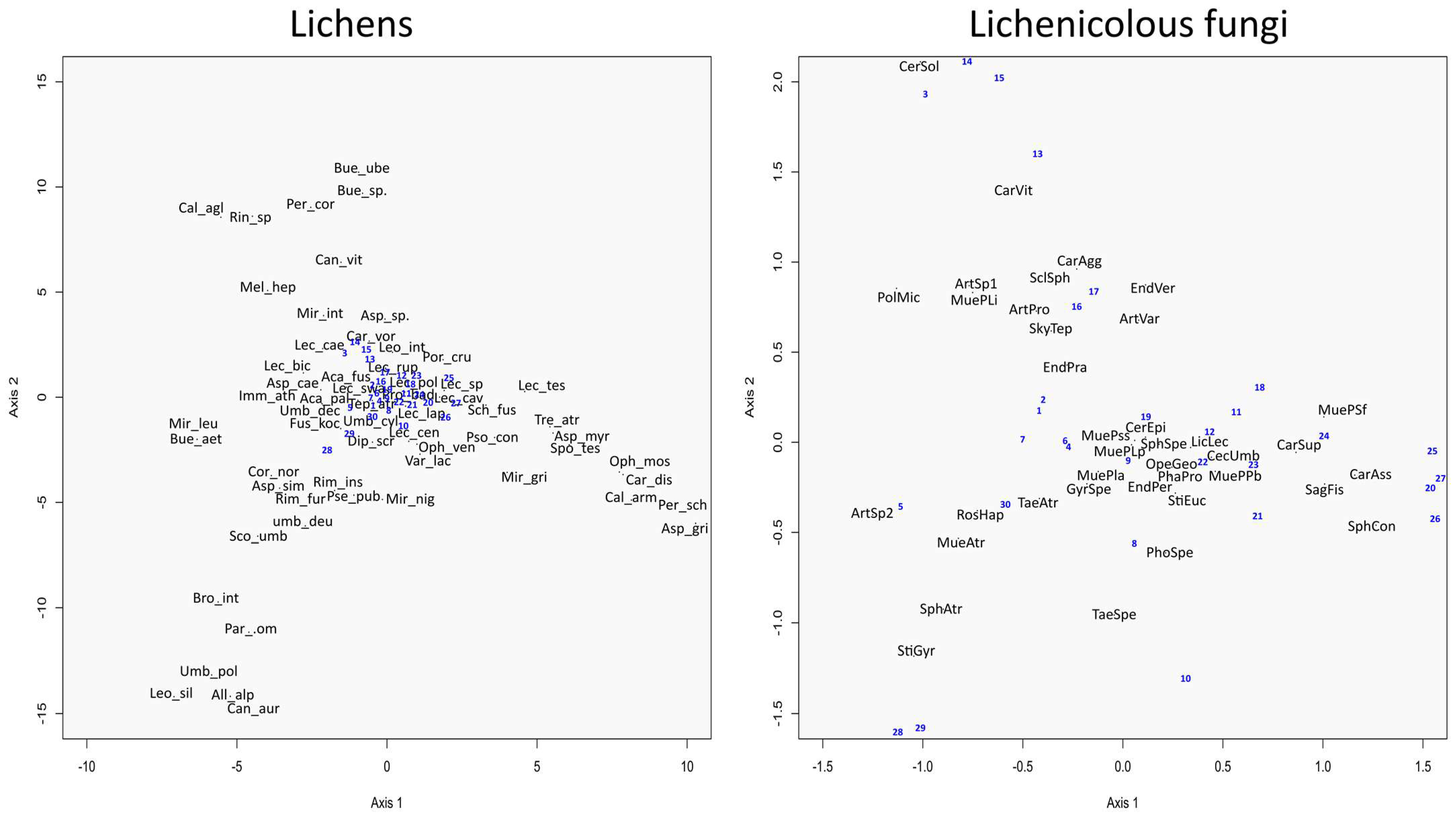
3.2. Congruence in Species Composition
4. Discussion
Cross-Taxon Analysis
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Margulis, L. Origin of Eukaryotic Cells; Yale University Press: New Haven, CT, USA, 1970. [Google Scholar]
- Gilbert, S.F.; Sapp, J.; Tauber, A.I. A symbiotic view of life: We have never been individuals. Q. Rev. Biol. 2012, 7, 325–341. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Grube, M.; Cardinale, M.; de Castro, J.V.; Müller, H.; Berg, G. Species-specific structural and functional diversity of bacterial communities in lichen symbioses. ISME J. 2009, 3, 1105–1115. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hawksworth, D.L.; Hill, D.J.; Phil, D. The Lichen-Forming Fungi, 1st ed.; Springer: New York, NY, USA, 1984. [Google Scholar]
- Honegger, R. The lichen symbiosis—What is so spectacular about it? Lichenologist 1998, 30, 193–212. [Google Scholar] [CrossRef] [Green Version]
- Hawksworth, D.L.; Grube, M. Lichens redefined as complex ecosystems. New Phytol. 2020, 227, 1281. [Google Scholar] [CrossRef]
- Denton, G.H.; Karlén, W. Lichenometry: Its application to Holocene moraine studies in southern Alaska and Swedish Lapland. Arctic Alpine Res. 1973, 5, 347–372. [Google Scholar] [CrossRef]
- Lawrey, J.D.; Diederich, P. Lichenicolous fungi: Interactions, evolution, and biodiversity. Bryologist 2003, 106, 80–120. [Google Scholar] [CrossRef]
- Muggia, L.; Grube, M. Fungal diversity in lichens: From extremotolerance to interactions with algae. Life 2018, 8, 15. [Google Scholar] [CrossRef] [Green Version]
- Spribille, T.; Resl, P.; Stanton, D.E. and Tagirdzhanova, G. Evolutionary biology of lichen symbioses. New Phytol. 2022, 234, 1566–1582. [Google Scholar] [CrossRef]
- Fernández-Mendoza, F.; Fleischhacker, A.; Kopun, T.; Grube, M.; Muggia, L. ITS 1 metabarcoding highlights low specificity of lichen mycobiomes at a local scale. Mol. Ecol. 2017, 26, 4811–4830. [Google Scholar] [CrossRef]
- Banchi, E.; Stankovic, D.; Fernández-Mendoza, F.; Gionechetti, F.; Pallavicini, A.; Muggia, L. ITS2 metabarcoding analysis complements lichen mycobiome diversity data. Mycol. Prog. 2018, 17, 1049–1066. [Google Scholar] [CrossRef] [Green Version]
- Diederich, P.; Lawrey, J.D.; Ertz, D. The 2018 classification and checklist of lichenicolous fungi, with 2000 non-lichenized, obligately lichenicolous taxa. Bryologist 2018, 121, 340–425. [Google Scholar] [CrossRef]
- Harutyunyan, S.; Muggia, L.; Grube, M. Black fungi in lichens from seasonally arid habitats. Stud. Mycol. 2008, 61, 83–90. [Google Scholar] [CrossRef]
- Arnold, A.E.; Miadlikowska, J.; Higgins, K.L.; Sarvate, S.D.; Gugger, P.; Way, A.; Hofstetter, V.; Kauff, F.; Lutzoni, F. A phylogenetic estimation of trophic transition networks for ascomycetous fungi: Are lichens cradles of symbiotrophic fungal diversification? Syst. Biol. 2009, 58, 283–297. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Muggia, L.; Fleischhacker, A.; Kopun, T.; Grube, M. Extremotolerant fungi from alpine rock lichens and their phylogenetic relationships. Fun. Div. 2016, 76, 119–142. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hafellner, J. Focus on lichenicolous fungi: Diversity and taxonomy under the principle “one fungus–one name”. Biosyst. Ecol. Ser 2018, 34, 227–243. [Google Scholar]
- Kumar, V.; Joseph, S.; Sharma, Y.P.; Nayaka, S. An annotated catalogue of the lichenicolous fungi of Jammu and Kashmir and Ladakh, India with new records and identification key. J. Asia-Pac. Biodiv. 2022, 15, 527–540. [Google Scholar] [CrossRef]
- Varga, N.; Lőkös, L.; Farkas, E. Annotated checklist of the lichenicolous fungi of Hungary. Diversity 2021, 13, 557. [Google Scholar] [CrossRef]
- Zhurbenko, M.P. Lichenicolous fungi from the Holarctic. Part IV: New reports and a key to species on Dermatocarpon. Opusc. Philolichenum 2021, 20, 44–53. [Google Scholar]
- Roux, C. Catalogue des lichens et champignons lichénicoles de France métropolitaine, 2nd ed.; Association française de lichénologie (A. F. L.): Fontainebleau, France, 2017; 1581p. [Google Scholar]
- Spribille, T.; Pérez-Ortega, S.; Tønsberg, T.; Schirokauer, D. Lichens and lichenicolous fungi of the Klondike Gold Rush National Historic Park, Alaska, in a global biodiversity context. Bryologist 2010, 113, 439–515. [Google Scholar] [CrossRef]
- Hafellner, J.; Herzog, G.; Mayrhofer, H. Zur Diversität von lichenisierten und lichenicolen Pilzen in den Ennstaler Alpen (Österreigh: Steiermark, Oberösterreich). Mitt. Naturwiss. Ver. Steiermark 2008, 137, 131–204. [Google Scholar]
- Fleischhacker, A.; Grube, M.; Kopun, T.; Hafellner, J.; Muggia, L. Community analyses uncover high diversity of lichenicolous fungi in alpine habitats. Microb. Ecol. 2015, 70, 348–360. [Google Scholar] [CrossRef]
- Toranza, C.; Arim, M. Cross-taxon congruence and environmental conditions. BMC Ecol. 2010, 10, 18. [Google Scholar] [CrossRef] [Green Version]
- Fattorini, S.; Dennis, R.L.; Cook, L.M. Use of cross-taxon congruence for hotspot identification at a regional scale. PLoS ONE 2012, 7, e40018. [Google Scholar] [CrossRef] [Green Version]
- Zara, L.; Tordoni, E.; Castro-Delgado, S.; Colla, A.; Maccherini, S.; Marignani, M.; Panepinto, F.; Trittoni, M.; Bacaro, G. Cross-taxon relationships in Mediterranean urban ecosystem: A case study from the city of Trieste. Ecol. Indic. 2021, 125, 107538. [Google Scholar] [CrossRef]
- Uboni, C.; Tordoni, E.; Brandmayr, P.; Battistella, S.; Bragato, G.; Castello, M.; Colombetta, G.; Poldini, L.; Bacaro, G. Exploring cross-taxon congruence between carabid beetles (Coleoptera: Carabidae) and vascular plants in sites invaded by Ailanthus altissima versus non-invaded sites: The explicative power of biotic and abiotic factors. Ecol. Indic. 2019, 103, 145–155. [Google Scholar] [CrossRef]
- Corcos, D.; Lami, F.; Nardi, D.; Boscutti, F.; Sigura, M.; Giannone, F.; Pantini, P.; Tagliapietra, A.; Busato, F.; Sibella, R.; et al. Cross-taxon congruence between predatory arthropods and plants across Mediterranean agricultural landscapes. Ecol. Indic. 2021, 123, 107366. [Google Scholar] [CrossRef]
- Prendergast, J.R.; Wood, S.N.; Lawton, J.H.; Eversham, B.C. Correcting for variation in recording effort in analyses of diversity hotspots. Biodiv. Lett. 1993, 1, 39–53. [Google Scholar] [CrossRef]
- Howard, P.C.; Viskanic, P.; Davenport, T.R.; Kigenyi, F.W.; Baltzer, M.; Dickinson, C.J.; Lwanga, J.S.; Matthews, R.A.; Balmford, A. Complementarity and the use of indicator groups for reserve selection in Uganda. Nature 1998, 394, 472–475. [Google Scholar] [CrossRef]
- Rodrigues, A.S.; Brooks, T.M. Shortcuts for biodiversity conservation planning: The effectiveness of surrogates. Ann. Rev. Ecol. Evol. Syst. 2007, 38, 713–737. [Google Scholar] [CrossRef]
- Heino, J. Are indicator groups and cross-taxon congruence useful for predicting biodiversity in aquatic ecosystems? Ecol. Indic. 2010, 10, 112–117. [Google Scholar] [CrossRef]
- Bacaro, G.; Tordoni, E.; Martellos, S.; Maccherini, S.; Marignani, M.; Muggia, L.; Petruzzellis, F.; Napolitano, R.; Da Re, D.; Guidi, T.; et al. Cross taxon congruence between lichens and vascular plants in a riparian ecosystem. Diversity 2019, 11, 133. [Google Scholar] [CrossRef] [Green Version]
- Santi, E.; Bacaro, G.; Rocchini, D.; Chiarucci, A.; Bonini, I.; Brunialti, G.; Muggia, L.; Maccherini, S. Methodological issues in exploring cross-taxon congruence across vascular plants, bryophytes and lichens. Folia Geobot. 2016, 51, 297–304. [Google Scholar] [CrossRef]
- Chiarucci, A.; Bacaro, G.; Rocchini, D.; Ricotta, C.; Palmer, M.W.; Scheiner, S.M. Spatially constrained rarefaction: Incorporating the autocorrelated structure of biological communities into sample-based rarefaction. Comm. Ecol. 2009, 10, 209–214. [Google Scholar] [CrossRef]
- Bacaro, G.; Altobelli, A.; Cameletti, M.; Ciccarelli, D.; Martellos, S.; Palmer, M.W.; Ricotta, C.; Rocchini, D.; Scheiner, S.M.; Tordoni, E.; et al. Incorporating spatial autocorrelation in rarefaction methods: Implications for ecologists and conservation biologists. Ecol. Indic. 2016, 69, 233–238. [Google Scholar] [CrossRef] [Green Version]
- Thouverai, E.; Pavoine, S.; Tordoni, E.; Rocchini, D.; Ricotta, C.; Chiarucci, A.; Bacaro, G. Rarefy: Rarefaction Methods, R Package Version 1.1. 2021. Available online: https://CRAN.R-project.org/package=Rarefy (accessed on 1 April 2022).
- Wei, T.; Simko, V. R Package ‘Corrplot’: Visualization of a Correlation Matrix, Version 0.92. 2021. Available online: https://github.com/taiyun/corrplot (accessed on 1 April 2022).
- Veech, J.A.; Summerville, K.S.; Crist, T.O.; Gering, J.C. The additive partitioning of species diversity: Recent revival of an old idea. Oikos 2002, 99, 3–9. [Google Scholar] [CrossRef] [Green Version]
- Whittaker, R.H. New Concepts of Kingdoms of Organisms: Evolutionary relations are better represented by new classifications than by the traditional two kingdoms. Science 1969, 163, 150–160. [Google Scholar] [CrossRef] [Green Version]
- Lande, R. Statistics and partitioning of species diversity, and similarity among multiple communities. Oikos 1996, 1, 5–13. [Google Scholar] [CrossRef]
- Oksanen, J.; Blanchet, F.G.; Kindt, R.; Legendre, P.; Minchin, P.R.; O’hara, R.B.; Simpson, G.L.; Solymos, P.; Stevens, M.H.; Wagner, H.; et al. Package ‘Vegan’. Community Ecology Package, Version. 2013. Available online: https://www.researchgate.net/publication/313502495_Vegan_Community_Ecology_Package/link/5c285a96458515a4c700bac2/download (accessed on 1 April 2022).
- Legendre, P.; Legendre, L. Numerical Ecology, 3rd ed.; Elsevier: Amsterdam, The Netherlands, 2012. [Google Scholar]
- Lários, M.C.; da Cunha, C.N.; Penha, J.; Landeiro, V.L.; Pinho, J.B.; Aragona, M.; Valerio, L.M.; Strüssmann, C.; Marques, M.I.; Lourenço, L.S.; et al. Evidence of cross-taxon congruence in Neotropical wetlands: Importance of environmental and spatial factors. Global Ecol. Cons. 2017, 12, 108–118. [Google Scholar] [CrossRef]
- Legendre, P.; Legendre, L. Numerical Ecology, 2nd ed.; Elsevier Science: Amsterdam, The Netherlands, 1998; 852p. [Google Scholar]
- ter Braak, C.J.F.; Schaffers, A.P. Co-correspondence analysis: A new ordination method to relate two community compositions. Ecology 2004, 85, 834–846. [Google Scholar] [CrossRef] [Green Version]
- Schaffers, A.P.; Raemakers, I.P.; Sýkora, K.V.; ter Braak, C.J.F. Arthropod assemblages are best predicted by plant species composition. Ecology 2008, 89, 782–794. [Google Scholar] [CrossRef]
- Martens, H.; Naes, T. Multivariate Calibration; Wiley: Chicester, UK, 1992. [Google Scholar]
- Bacaro, G.; Gioria, M.; Ricotta, C. Beta diversity reconsidered. Ecol. Res. 2012, 28, 537–540. [Google Scholar] [CrossRef]
- Anderson, M.J. Distance-Based Tests for Homogeneity of Multivariate Dispersions. Biometrics 2006, 62, 245–253. [Google Scholar] [CrossRef]
- Simpson, G.L. Co-Correspondence Analysis Ordination Methods, R PackageVersion 0.4-3. 2009. Available online: https://cran.r-project.org/package=cocorresp (accessed on 1 April 2022).
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2021. [Google Scholar]
- Harry, I.; Höfer, H.; Schielzeth, H.; Assmann, T. Protected habitats of Natura 2000 do not coincide with important diversity hotspots of arthropods in mountain grasslands. Insect Conserv. Div. 2019, 12, 329–338. [Google Scholar] [CrossRef]
- U’Ren, J.M.; Lutzoni, F.; Miadlikowska, J.; Laetsch, A.D.; Arnold, A.E. Host and geographic structure of endophytic and endolichenic fungi at a continental scale. Am. J. Bot. 2012, 99, 898–914. [Google Scholar] [CrossRef] [Green Version]
- Oh, S.-Y.; Yang, J.H.; Woo, J.-J.; Oh, S.-O.; Hur, J.-S. Diversity and distribution patterns of endolichenic fungi in Jeju Island, South Korea. Sustainability 2020, 12, 3769. [Google Scholar] [CrossRef]
- Jackson, D.A.; Harvey, H.H. Fish and benthic invertebrates: Community concordance and community–environment relationships. Can. J. Fish. Aquat. Scie. 1993, 50, 2641–2651. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Plot | State | Region | Mountain | Coordinates | Altitude |
|---|---|---|---|---|---|
| 1 | Austria | Styria | Koralpe | 46°50′20″ N/15°02′35″ E | ca. 1760 m |
| 2 | Austria | Styria | Koralpe | 46°50′38″ N/15°01′10″ E | ca. 1800 m |
| 3 | Austria | Styria | Koralpe | 46°49′22″ N/14°59′29″ E | ca. 1910 m |
| 4 | Austria | Styria | Koralpe | 46°48′23″ N/14°58′57″ E | ca. 1980 m |
| 5 | Austria | Styria | Koralpe | 46°48′23″ N/15°00′33″ E | ca. 1790 m |
| 6 | Austria | Styria | Koralpe | 46°46′50″ N/15°01′35″ E | ca. 1800 m |
| 7 | Austria | Styria | Koralpe | 46°46′47″ N/14°58′13″ E | ca. 2070 m |
| 8 | Austria | Styria | Koralpe | 46°46′29″ N/15°00′52″ E | ca. 1760 m |
| 9 | Austria | Carinthia | Koralpe | 46°47′39″ N/14°57′42″ E | ca. 2000 m |
| 10 | Austria | Carinthia | Koralpe | 46°48′54″ N/14°58′14″ E | ca. 1860 m |
| Groups | Data | r | p-Value |
|---|---|---|---|
| lichens vs. lichenicolous fungi | Species richness (abundance) | 0.0068 | 0.486 |
| Species richness (p/a) | 0.0130 | 0.376 | |
| lichens vs. lichenicolous fungi | Species composition (abundance) | 0.2543 | 0.006 ** |
| Species composition (p/a) | 0.1906 | 0.012 * |
| Data | Taxa | Axes | Cross-Validation Fit (%) | Cumulative Expl. Variance (%) | p-Value |
|---|---|---|---|---|---|
| Abundance | Lichenicolous fungi | 1 | 8.26 | 7.07 | 0.009 |
| 2 | 7.91 | 13.06 | 0.002 | ||
| P/A | Lichenicolous fungi | 1 2 | 7.91 6.63 | 7.27 14.50 | 0.002 0.085 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
De Carolis, R.; Muggia, L.; Bacaro, G. Lichen and Lichenicolous Fungal Communities Tested as Suitable Systems for the Application of Cross-Taxon Analysis. Diversity 2023, 15, 285. https://doi.org/10.3390/d15020285
De Carolis R, Muggia L, Bacaro G. Lichen and Lichenicolous Fungal Communities Tested as Suitable Systems for the Application of Cross-Taxon Analysis. Diversity. 2023; 15(2):285. https://doi.org/10.3390/d15020285
Chicago/Turabian StyleDe Carolis, Roberto, Lucia Muggia, and Giovanni Bacaro. 2023. "Lichen and Lichenicolous Fungal Communities Tested as Suitable Systems for the Application of Cross-Taxon Analysis" Diversity 15, no. 2: 285. https://doi.org/10.3390/d15020285