
Genus Apterocuris Jacobson, 1901 (Coleoptera: Chrysomelidae) in the Subfamily Chrysomelinae with Description of a New Species and Remarks on Significance of Preimaginal Characters †

Abstract
:1. Introduction
2. Materials and Methods
3. Results
3.1. Genus Apterocuris Jacobson, 1901
3.2. Apterocuris sibirica (Gebler, 1830)
3.2.1. Material Examined
3.2.2. Redescription
3.2.3. Differential Diagnosis
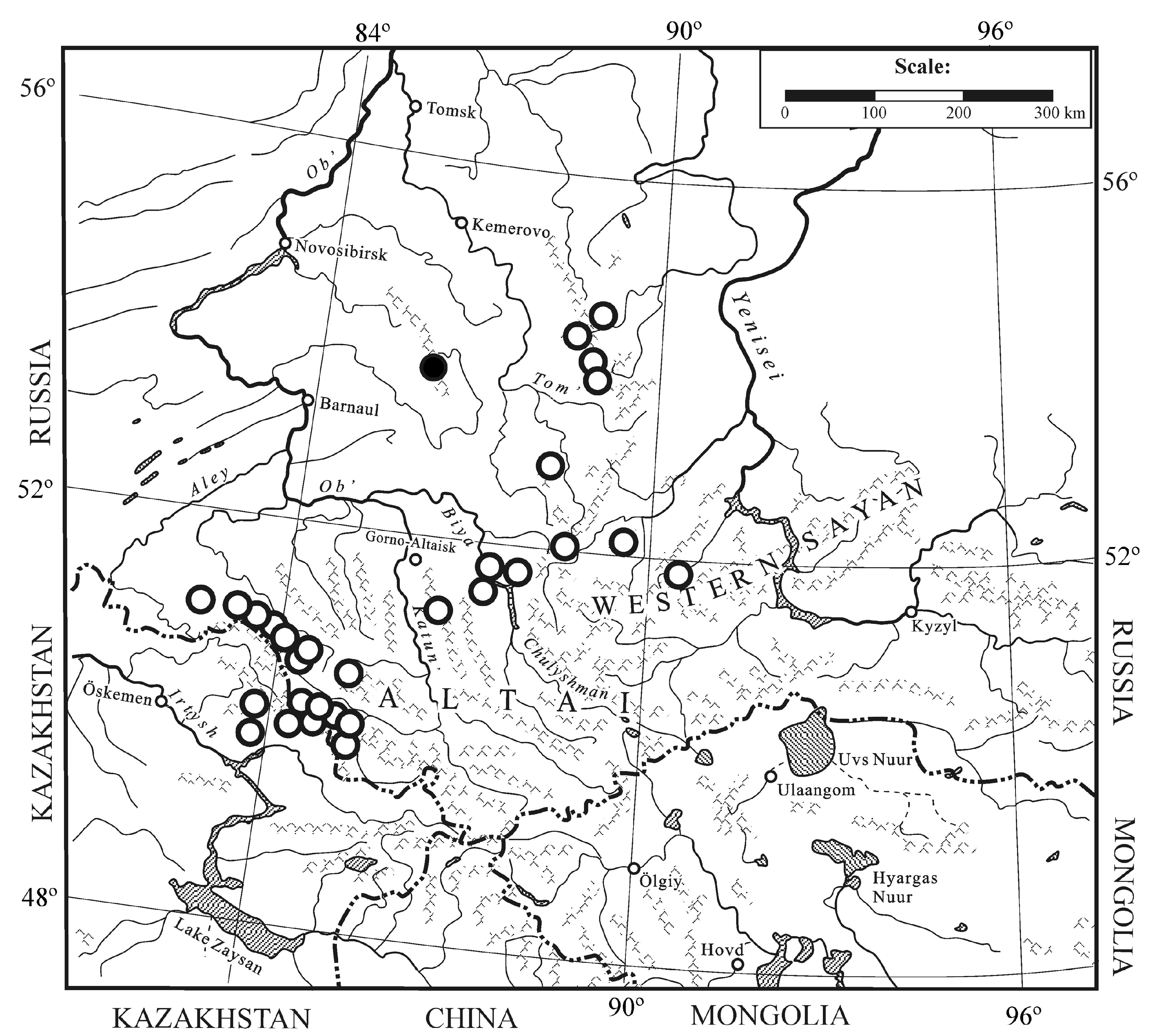
3.2.4. Distribution
3.2.5. Biotopes
3.2.6. Host Plants
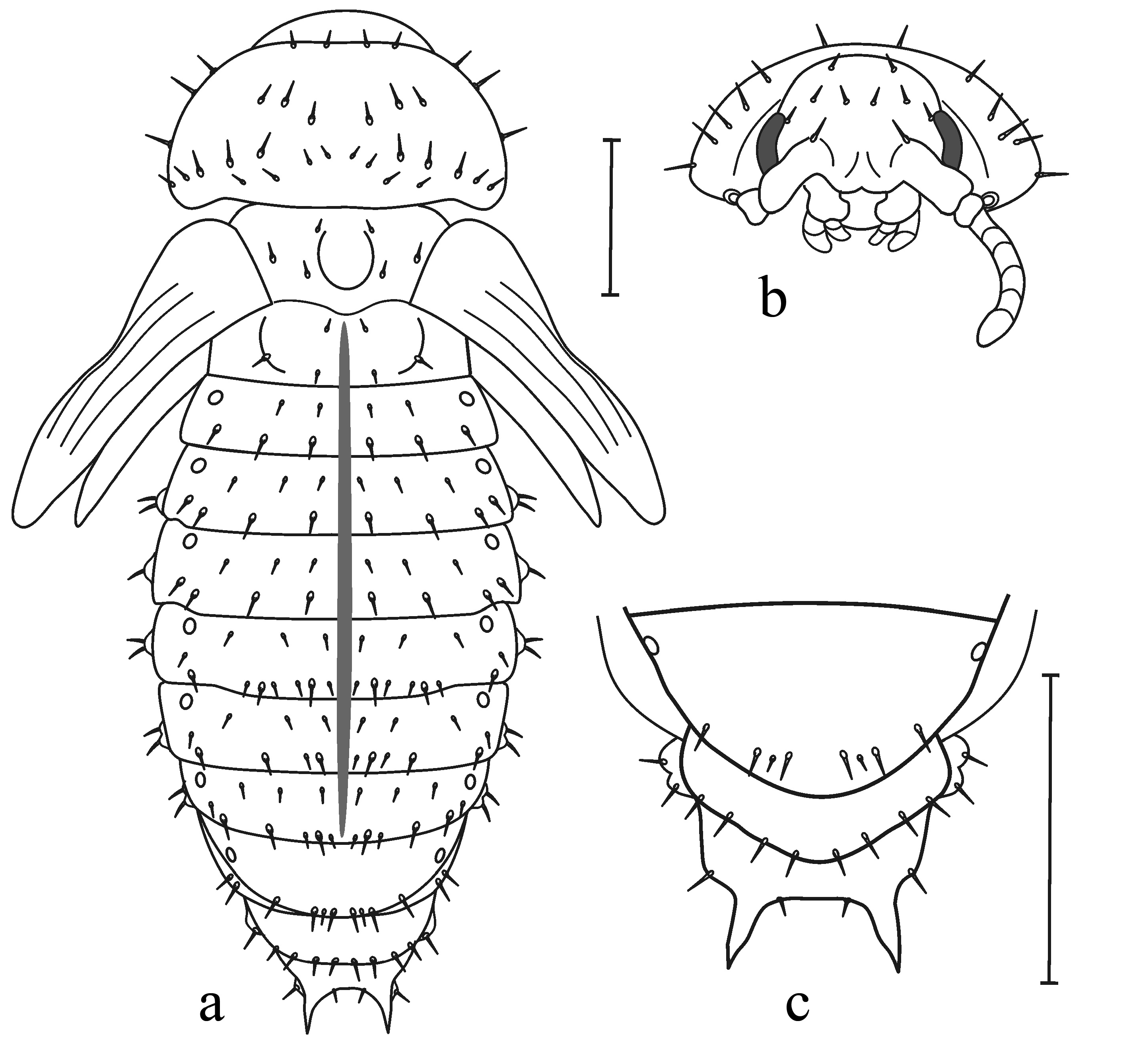
3.2.7. Larva
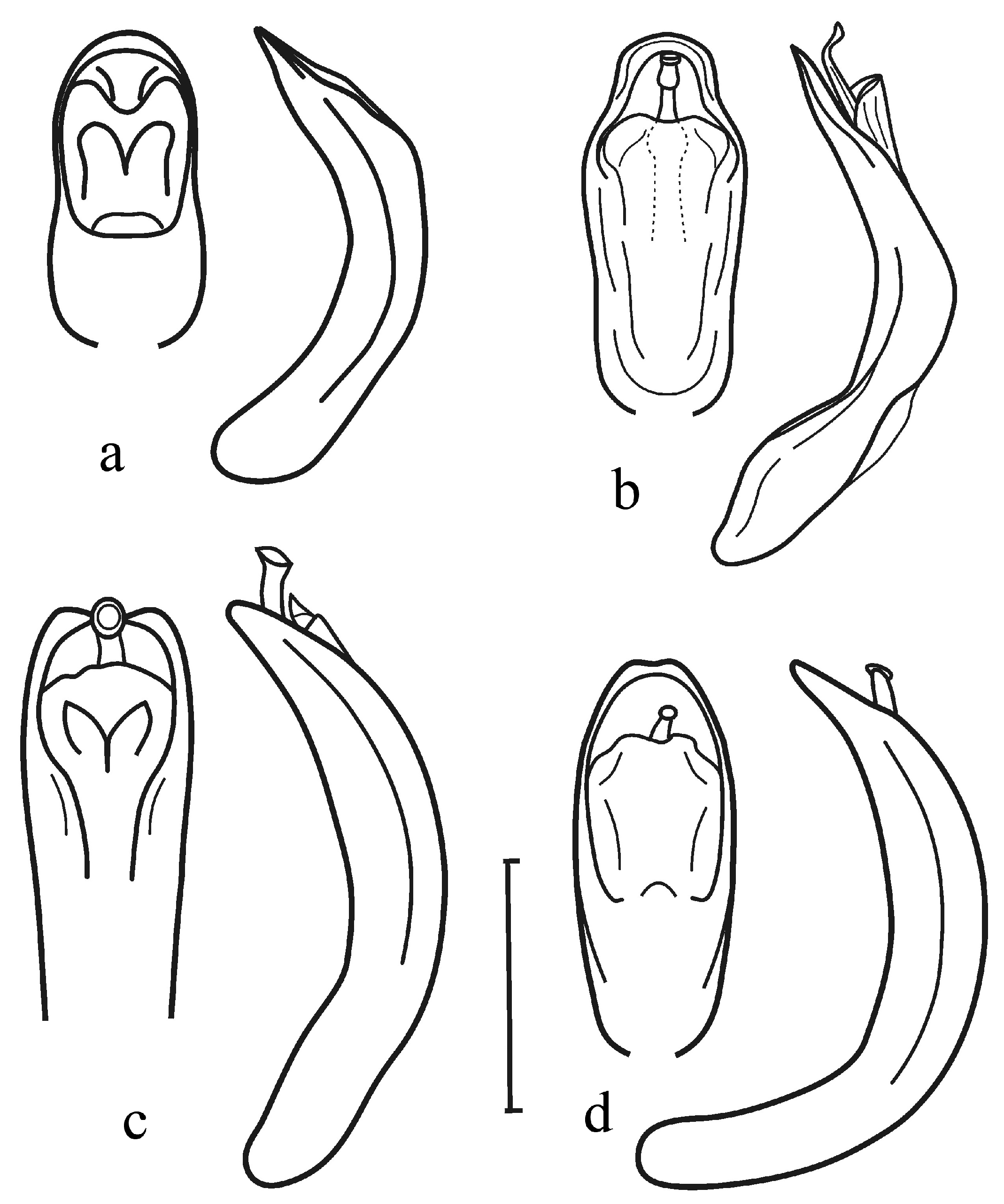
3.2.8. Pupa
3.3. Apterocuris brinevi sp. n.
3.3.1. Type Material
3.3.2. Description
3.3.3. Etymology
3.3.4. Differential Diagnosis
3.3.5. Distribution
4. Discussion


5. Conclusions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Jacobson, G. Chrysomelidae Sibiriae occidentalis. Tr. Rus. Entomol. Obs. Horae Soc. Entomol. Ross. 1901, 35, 73–102. [Google Scholar]
- Dolgin, M.M.; Medvedev, L.N. Previously unknown larvae of leaf beetles from Altai, Part 1. Zool. Zhurnal 1974, 53, 642–645. [Google Scholar]
- Daccordi, M. Notes for phylogenetic study of Chrysomelinae, with descriptions of new taxa and a list of all the known genera (Coleoptera: Chrysomelidae: Chrysomelinae). In Proceedings of the Third International symposium on the Chrysomelidae, Beijing, China, 4 July 1992; Backhuys Publ.: Leiden, The Netherlands, 1994; pp. 60–81. [Google Scholar]
- Medvedev, L.N. On taxonomy and nomenclature of leaf beetles species (Coleoptera, Chrysomelidae) of Russia. Euroasian Entomol. J. 2011, 10, 475–478. [Google Scholar]
- Takizawa, H. Note on the Pupae of the Genus Chrysolina Motschulsky in Japan (Coleoptera: Chrysomelidae). Entomol. Rev. Jpn. 1971, 23 (Suppl. 11–14), 110–111. [Google Scholar]
- Steinhausen, W.R. Die Puppen mitteleuropäischer Blattkäfer-Eine vorläufige Bestimmungstabelle. 1 Teil (Coleoptera, Chrysomelidae). Mitt. Münchener Entomol. Ges. 2001, 91, 35–63. [Google Scholar]
- Steinhausen, W.R. Comparative morphology of pupae of the leaf beetle subfamilies Chrysomelinae and Galerucinae. In Proceedings of the Fourth International Symposium on the Chrysomelidae: Proceedings of XX ICE, Florence, Italy, 30 August 1996; Biondi, M., Daccordi, M., Furth, D.G., Eds.; Museo Regionale di Scienze Naturale: Torino, Italy, 1998; pp. 311–318. [Google Scholar]
- Reid, C.A.M. Chrysomelinae Latreille, 1802. In Handbook of Zoology. Arthropoda: Insecta. Coleoptera, Beetles, Vol. 3: Morphology and Systematics (Phytophaga); Walter de Gruyter GmbH: Berlin, Germany; Walter de Gruyter GmbH: Boston, MA, USA, 2014; pp. 243–250. [Google Scholar]
- Lopatin, I.K. Zhuki-Listoedy (Chrysomelidae) Srednei Azii i Kazakhstana [The Leaf Beetles (Chrysomelidae) of Middle Asia and Kazakhstan]; Nauka Publ.: Leningrad USSR, Russia, 1977; p. 270. [Google Scholar]
- Lopatin, I.K.; Kulenova, K.Z. Zhuki-Listoedy (Coleoptera, Chrysomelidae) Kazakhstana: Opredelitel’ [The Leaf Beetles (Coleoptera, Chrysomelidae) of Kazakhstan: Identification keys]; Nauka Publ.: Alma-Ata, Kazakhstan, 1986; p. 200. [Google Scholar]
- Medvedev, L.N.; Dubeshko, L.N. Opredelitel’ Listoedov Sibiri [Identification Keys to Siberian Leaf Beetles]; Irkutsk University Publ.: Irkutsk, Russia, 1992; p. 224. [Google Scholar]
- Lopatin, I.K. Zhuki-Listoedy (Insecta, Coleoptera, Chrysomelidae) Central’noi Azii [The Leaf Beetles (Insecta, Coleoptera, Chrysomelidae) of Central Asia]; Belarusian University Publ.: Minsk, Belarus, 2010; p. 511. [Google Scholar]
- Warchalowski, A. The Palaearctic Chrysomelidae: Identification Keys. Vol. 1; Natura Optima dux Foundation: Warszawa, Poland, 2010; p. 629. [Google Scholar]
- Kippenberg, H. Subfamily Chrysomelinae Latreille, 1802. In Catalogue of Palaearctic Coleoptera. Vol. 6. Chrysomeloidea.; Löbl, I., Smetana, A., Eds.; Apollo Books: Stenstrup, Denmark, 2010; pp. 390–443. [Google Scholar]
- Gebler, F. Bemerkungen über die Insecten Sibiriens, vorzüglich des Altai. In Reise Durch das Altai-Gebirge und die Soongorische Kirgisen-Steppe. Zweiter Theil; Ledebour, C.F., Ed.; G. Reimer: Berlin, Germany, 1830; pp. 1–228. [Google Scholar]
- Gebler, F. Notae et additamenta ad Catalogum Coleopterorum Sibiriae occidentalis et confinis Tatariae operis. Bull. Soc. Nat. Moscou 1841, 14, 577–625. [Google Scholar]
- Daccordi, M.; Lavarini, N. Le specie Italiane del genere Phaedon (Coleoptera: Chrysomelidae). Boll. Mus. Civ. St. Nat. Verona 1993, 17, 481–512. [Google Scholar]
- Ge, S.-Q.; Daccordi, M.; Cui, J.-Z.; Ren, J.; Yang, X.-K. Revision of genus Sclerophaedon Weise from China (Coleoptera, Chrysomelidae, Chrysomelinae). Ann. Del Mus. Civ. Di Stor. Nat. G. Doria 2013, 105, 81–102. [Google Scholar]
- Hennig, W. Übersicht über die Larven der wichtigsten deutschen Chrysomelinen. Arb. Phys. Angew. Ent. 1938, 5, 85–136. [Google Scholar]
- Steinhausen, W.R. Versuch einer Übereinstimmung imaginaler und larvaler Systeme (Coleoptera, Chrysomelidae, Chrysomelinae). In Verhandlungen des 14, Proceedings of the Internationalen Symposiums für Entomofaunistik in Mitteleuropa, SIEEC, München, Germany, 4–9 September 1994; Gerstmeier, R., Scherer, G., Eds.; SIEEC XIV: München, Germany, 1996; pp. 261–263. [Google Scholar]
- Boland, W. Sequestration of plant-derived glycosides by leaf beetles: A model system for evolution and adaptation. Perspect. Sci. 2015, 6, 38–48. [Google Scholar] [CrossRef]
- Zaitsev, Y.M.; Medvedev, L.N. Lichinki Zhukov-Listoedov Rossii [The Larvae of Leaf Beetles of Russia]; T-vo Nauchnyh Izdaniy KMK: Moscow, Russia, 2009; p. 246. [Google Scholar]
- Weise, J. Chrysomelidae. Lieferung 2. In Naturgeschichte der Insecten Deutschlands. Erste Abteilung Coleoptera. Sechster Band; Nicolaische Verlags-Buchhandlung: Berlin, Germany, 1882; pp. 193–368. [Google Scholar]
- Daccordi, M.; Medvedev, L. A study of the species of Sclerophaedon from the Himalaya (Coleoptera: Chrysomelidae). Boll. Soc. entom. Ital. Genova 2000, 132, 219–231. [Google Scholar]
- Lopatin, I.K. New species of the leaf beetles (Coleoptera, Chrysomelidae) from China, VI. Entom. Rev. 2006, 86, 662–669. [Google Scholar] [CrossRef]
- Warchalowski, A. Chrysomelidae-Stonkowate (Insecta, Coleoptera). Część IV (podplemiona: Chrysomelina, Gonioctenina, Phratorina i Entomoscelina oraz podrodzina Galerucinae). In Fauna Polski-Fauna Poloniae; Dzial Wydawnictw MiIZ PAN: Warszawa, Poland, 1994; Volume 16, p. 302. [Google Scholar]
- Petitpierre, E. Coleoptera Chrysomelidae II. In Fauna Iberica. Vol. 46; Ramos, M.A., Fernandes, J., Daccordi, M., Vela, J.M., Eds.; Museo Nacional de Ciencias Naturales: Madrid, Spain, 2019; Volume 46, p. 596. [Google Scholar]
- Kippenberg, H.; Mikhailov, Y.E. Subfamily Chrysomelinae Latreille, 1802. In Catalogue of Palaearctic Coleoptera. Chrysomeloidea., Volume 6; Revised and updated, ed.; Bezdek, J., Sekerka, L., Eds.; Koninklijke Brill NV: Leiden, The Netherlands, 2023; in press. [Google Scholar]
- Steinhausen, W.R. Integrierte Systematik bei Blattkäfern (Coleoptera, Chrysomelidae, Chrysomelinae). Entomol. Blätter 2001, 97, 225–236. [Google Scholar]
- Chen, S.H. Recherches sur les Chrysomelinae de la Chine et du Tonkin. Ann. Soc. Entomol. Fr. 1935, 104, 127–158. [Google Scholar]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Larvae | Pupae | Imagines | |
|---|---|---|---|
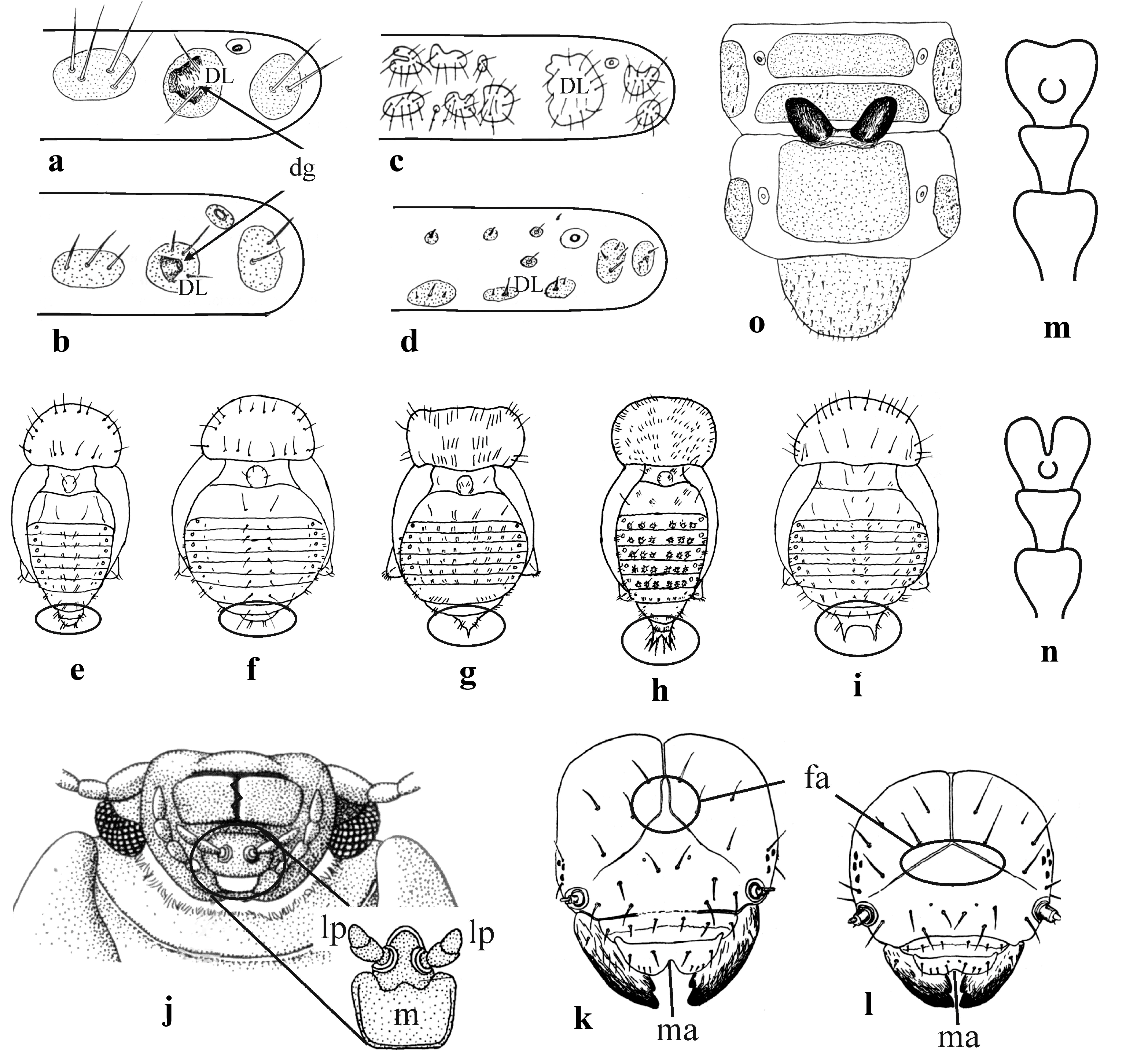
| 1 | Dorsolateral sclerites of thoracic and abdominal terga with large openings of dorsal defensive glands (Figure 8 and Figure 9a,b) …………… | Last abdominal segment without any appendages (Figure 9e,f) ……… | Labial palps are attached to mentum close to each other (Figure 9j) …… 2 |
| -- | Dorsolateral sclerites without openings of dorsal defensive glands (Figure 9c,d) ………………… | Last abdominal segment with one or a pair of urogomphi (Figure 9g–i)… | Labial palps are attached to mentum at larger distance from each other …7 |
| 2 | Anterior margin of labrum with well-pronounced median incisure (Figure 9k). Claws with large tooth (dendrophagous genera of subtribe Chrysomelina) ……………… | Dorsum completely coloured or with brown markings, especially thoracic and abdominal terga. Leaf-pupating species of Chrysomelina ……… | Tarsomere 3 entire apically or shallowly emarginated (Figure 9m), rarely deeply incised (Figure 9n). Elytra without sutural stria on apical slope, tibiae without triangular process at apex ………………3 |
| -- | Anterior margin of labrum with or without median incisure (Figure 9l). Claws with or without tooth at base … | Dorsum almost white, conspicuously setose. Soil-pupating species of subtribe Chrysomelina ………… | Tarsomere 3 deeply incised, bilobed (Figure 9n). Elytra with sutural stria on apical slope, tibiae with triangular process at apex ……………… 4 |
| 3 | Antennae short, antennomere 2 transverse, antennomere 3 short, not longer than preceding, maximum twice longer than wide ………… | -- | Hind wings developed. Elytral punctures confused …. Plagiodera Chevrolat, 1836, Plagiosterna Motschulsky, 1860, Chrysomela Linnaeus, 1758, Gastrolina Baly, 1859 |
| -- | Antennae long, antennomere 2 of equal length and width, antennomere 3 long, 3–4 times longer than wide … | Pupae unknown | Hind wings reduced. Elytral punctures arranged in regular rows …… Sternoplatys Motschulsky, 1860 |
| 4 | Anterior margin of labrum without median incisure. Claws with or without tooth at base ………… | Abdominal segments without post-stigmal setae …………… | Claws with tooth at base ………… Phratora Chevrolat, 1836 |
| -- | Anterior margin of labrum with well-pronounced median incisure (Figure 9k). Claws without tooth at base ……………………… | Abdominal segments with post-stigmal setae …………… | Claws without tooth at base …… 5 |
| 5 | Frontal arms first parallel at base, then diverge (Figure 9k). Abdominal terga without pre-tergal sclerites or with their rudiments ………… | Body elongate. Length to width ratio more than 2.0 ……………… | Body elongate. Pronotum base and base of elytra with distinct convex margin (Figure 2). Prosternal appendix wide ……… Prasocuris Latreille, 1802, Oreothassa Jacobson, 1901 |
| -- | Frontal arms diverge from the very base, not parallel at any distance (Figure 9l). Post-tergal sclerites of abdominal terga with two setae … | Body short, oval. Length to width ratio less than 2.0 …………… | Body outline oval. Pronotum base narrowly margined, base of elytra not margined. Prosternal appendix narrow …………………… 6 |
| 6 | Microsculpture of chitinous body surface (tubercles and spikes) covers not only interspaces but also sclerites. Mesothorax with two pretergal sclerites bearing two setae each … | Outer setae of dorsal pairs at segments 7 and 9 very short, inner setae large and long; rarely outer setae completely absent ……… | Punctation of elytra arranged in regular rows ………………… Phaedon Megerle von Muhlfeld, 1823, Neophaedon Jacobson, 1901 |
| -- | Microsculpture of body surface visible only between sclerites and does not cover the latter. Mesothorax with four pretergal sclerites bearing one seta each or tergal sclerites unclear and their situation is indicated by setae …………… | Abdominal terga with a pair of setae at posterior side, from which inner setae slightly longer and at apex of last segment bent inwards ……… | Punctation of elytra confused …………Gastrophysa Chevrolat, 1836 |
| 7 | Body convex. Sclerites hardly visible or absent …………………… | Last abdominal segment with one sharply pointed urogomphus (Figure 9g) … subtribe Chrysolinina | Elytral epipleura with setae at interior border at least in apical part … Chrysolina Motschulsky, 1860, Oreina Chevrolat, 1836, Ambrostoma Motschulsky, 1845 |
| -- | Body weakly convex. Sclerites clearly visible and well-defined ……… | Last abdominal segment with a pair of urogomphi (Figure 4 and Figure 9h,i) … | Elytral epipleura without setae … 8 |
| 8 | A pair of eversible glands is between abdominal segments 7 and 8 (Figure 9o) ………………… | A pair of urogomphi at last abdominal segment only slightly separated at base from each other, divergent (Figure 9h) ………… | Last maxillary palpomere transversely shortened …… Gonioctena Chevrolat, 1836 (subtribe Gonioctenina) |
| -- | No eversible glands between abdominal segments 7 and 8. Mesoterga and metaterga with two rows of sclerites ……………… | A pair of urogomphi at last abdominal segment distinctly separated at base from each other by concave line and placed in parallel (Figure 4 and Figure 9i) …………… | Last maxillary palpomere sharpened ………………………… 9 |
| 9 | Head capsule with numerous secondary setae similar in length with primary setae. Thoracic and abdominal terga with three transverse rows of sclerites ……………… | A pair of urogomphi separated from each other at base by distance twice of their length……………… | Hind wings developed. Elytral punctures confused ………… 10 |
| -- | Head capsule with primary setae only or vertex with few secondary setae. Thoracic and abdominal terga with two transverse rows of sclerites (sometimes reduced). Claws with tooth at base ……………… | A pair of urogomphi separated from each other at base by distance equal to their length ……………… | Hind wings reduced. Elytral punctures arranged in regular rows (at least partly) ……………… 11 |
| 10 | Second transverse row of sclerites on meso-, metaterga and abdominal segments consists of 6 sclerites of equal size. All setae light-coloured, 5 to 10 on each sclerite ………… | Frons with 1 pair of setae, vertex with 10 pairs of short setae ………… | Anterior coxae with closed cavities ……… Entomoscelis Chevrolat, 1836 (subtribe Entomoscelina) |
| -- | Second transverse row of sclerites on meso-, metaterga and abdominal segments consists of 2 sclerites much larger than others. All setae dark, 10 to 20 on each sclerite ………… | Frons and vertex both with 10 pairs of setae ……………………… | Anterior coxae with open cavities … Colaphellus Weise, 1916 |
| 11 | Meso- and metathorax bear two post-tergal sclerites each. All sclerites of abdominal terga bear one long seta ………………………… | Body short, oval. Length to width ratio less than 2.0. Urogomphi with two pairs of setae outside (Figure 9i)………………………… | Body outline oval. Pronotum as broad as base of elytra, its hind angles acute. Elytral punctures large and sparse, elytral striae (usually striae 6–7) may be irregular at the sides (Figure 10)… Sclerophaedon Weise, 1882 |
| -- | Dorsal setae short, always shorter than length of sclerites. On abdominal terga each pretergal sclerite bears one seta and each post-tergal sclerite bears two or three setae ……………………… | Body elongate. Length to width ratio more than two. Urogomphi with one pair of setae outside (Figure 4)…… | Body outline oblong. Pronotum narrower than base of elytra, its hind angles right. Elytral punctures mostly confused and arranged in rows only along suture and lateral margins (Figure 1 and Figure 5) … Apterocuris Jacobson, 1901 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the author. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mikhailov, Y.E. Genus Apterocuris Jacobson, 1901 (Coleoptera: Chrysomelidae) in the Subfamily Chrysomelinae with Description of a New Species and Remarks on Significance of Preimaginal Characters. Diversity 2023, 15, 228. https://doi.org/10.3390/d15020228
Mikhailov YE. Genus Apterocuris Jacobson, 1901 (Coleoptera: Chrysomelidae) in the Subfamily Chrysomelinae with Description of a New Species and Remarks on Significance of Preimaginal Characters. Diversity. 2023; 15(2):228. https://doi.org/10.3390/d15020228
Chicago/Turabian StyleMikhailov, Yuri E. 2023. "Genus Apterocuris Jacobson, 1901 (Coleoptera: Chrysomelidae) in the Subfamily Chrysomelinae with Description of a New Species and Remarks on Significance of Preimaginal Characters" Diversity 15, no. 2: 228. https://doi.org/10.3390/d15020228