
Floral Specialization and Bat Pollination in Subtribe Cereinae (Cactaceae): A Morphological Approach

 , and
, and
Abstract
:
1. Introduction
2. Material and Methods
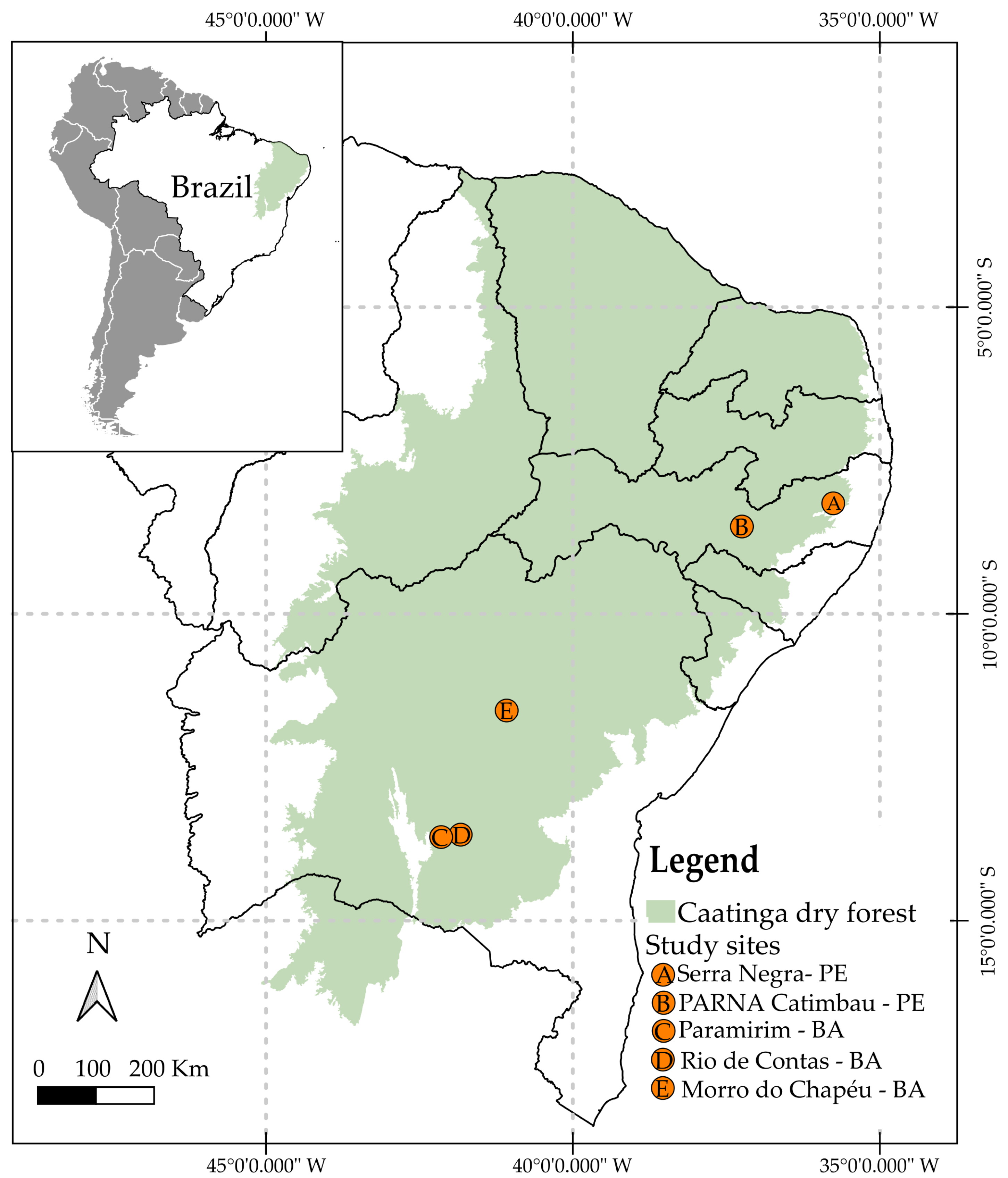
2.1. Study Sites
2.2. Studied Taxa
2.3. Floral Biology
2.4. Floral Traits and Morphometry
2.5. Floral Visitors
3. Results
3.1. Floral Biology
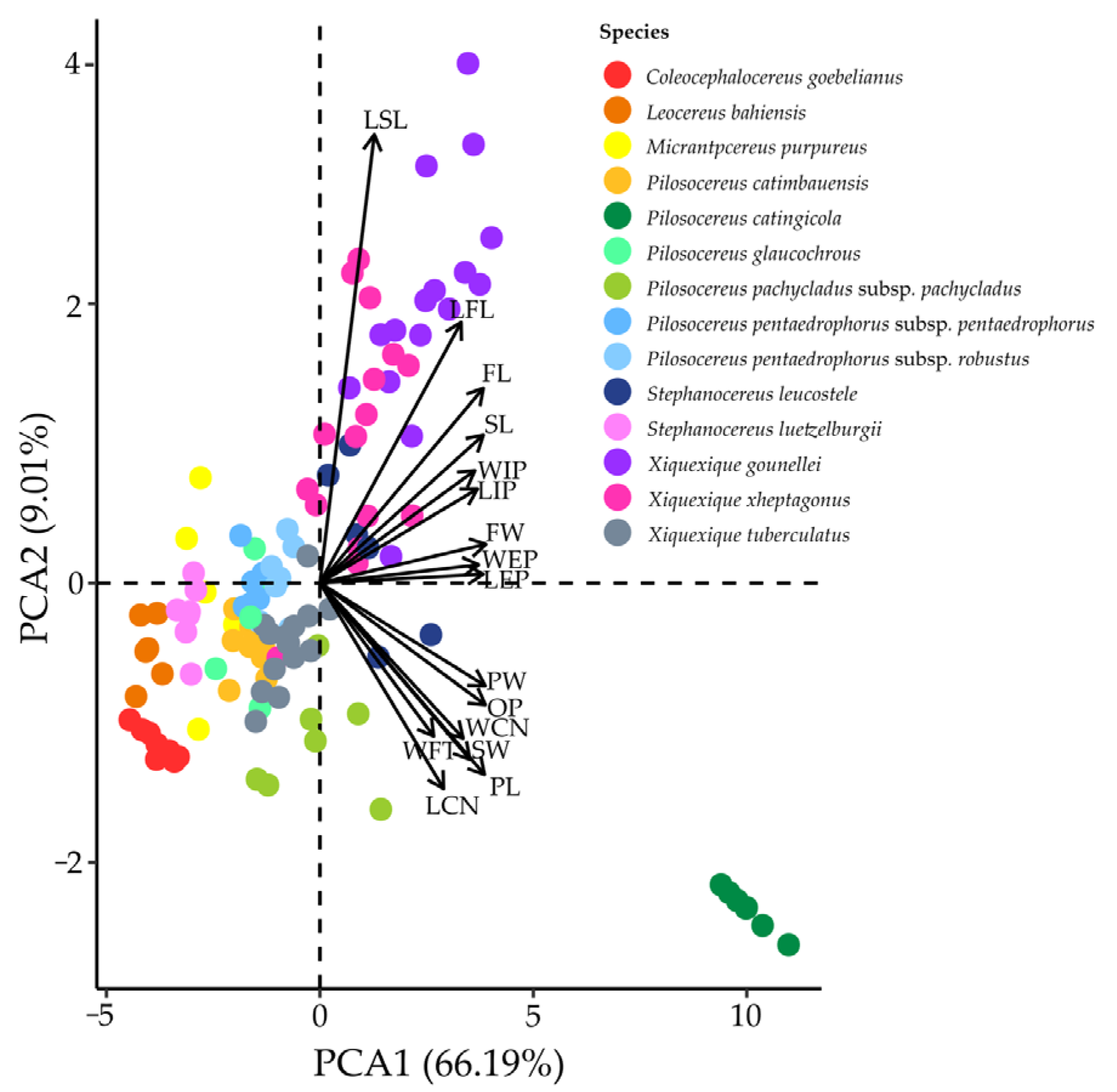
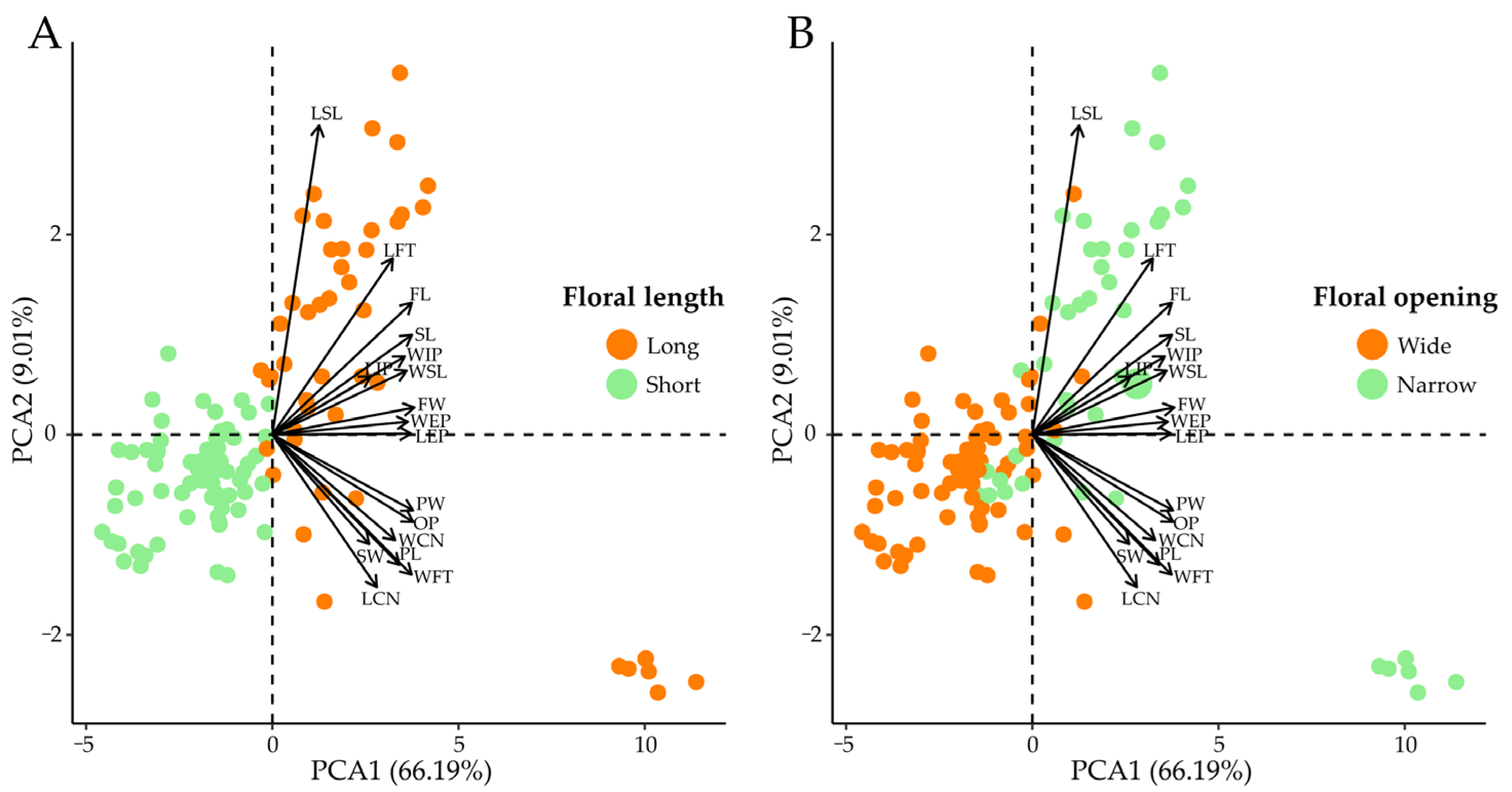
3.2. Quantification of Floral Traits
3.3. Nectar Standing Crop and Floral Visitors
4. Discussion
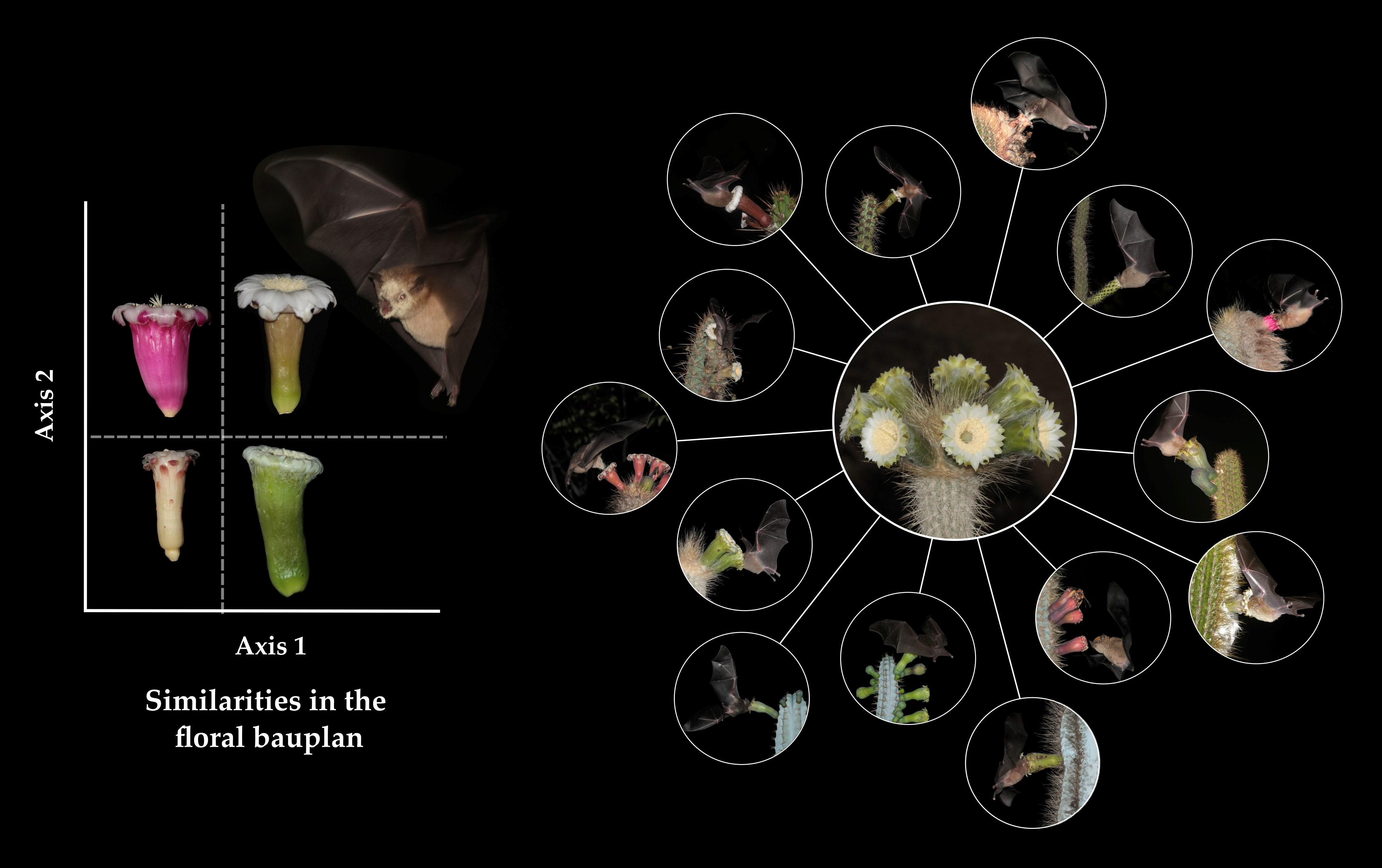
4.1. Floral Biology and Variation in Floral Morphology among Chiropterophilous Taxa of the Subtribe Cereinae
4.2. Participation of Bats in the Pollination Systems of Cereinae
5. Final Considerations
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Appendix A

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Traits * | GOE | BAH | PUR | CAT | CAG | GLAU | PAC | PPE | PPR | LEU | LUE | GOU | HEP | TUB |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| FL | 39.5 | 47.9 | 42.57 | 53.03 | 90.38 | 51.92 | 55.85 | 55.12 | 56.22 | 70.62 | 43.05 | 78.23 | 70.74 | 56.37 |
| FW | 22.61 | 16.01 | 30.6 | 29.35 | 72.73 | 24.58 | 30.77 | 24.17 | 25.15 | 36.59 | 26.39 | 49.29 | 40.43 | 32.86 |
| OP | 11.16 | 9.12 | 13.2 | 15.5 | 38.29 | 13.2 | 19.65 | 12.82 | 13.59 | 17.51 | 12.94 | 19.72 | 16.42 | 15.42 |
| PL | 4.3 | 8.86 | 6.83 | 7.92 | 21.75 | 10.81 | 8.88 | 10.64 | 10.78 | 11.47 | 6.92 | 8.96 | 9.54 | 8.9 |
| PW | 6.44 | 7.7 | 10.08 | 15.18 | 28.44 | 13.59 | 13.32 | 13.07 | 13.47 | 15.61 | 8.81 | 14.95 | 14.52 | 14.35 |
| LFT | 30.47 | 33.4 | 26.9 | 35.82 | 54.56 | 35.38 | 40.85 | 38.18 | 39.75 | 54.48 | 29.55 | 54.05 | 48.6 | 38.58 |
| WFT | 14.54 | 13.33 | 17.33 | 20.38 | 46.83 | 18.71 | 24.52 | 18.51 | 19.12 | 25.33 | 16.1 | 21.88 | 20.95 | 18.06 |
| LEP | 9.58 | 7 | 8.52 | 9.38 | 24.29 | 9.6 | 10.86 | 9.41 | 9.68 | 11.81 | 8.95 | 15.69 | 12.84 | 9.31 |
| WEP | 4.46 | 2.2 | 7.67 | 5.57 | 17.54 | 6.92 | 8.3 | 6.77 | 7.22 | 6.88 | 7.24 | 11.16 | 8.59 | 7.67 |
| LIP | 9.96 | 6.66 | 12.13 | 7.59 | 25.65 | 7.8 | 11.08 | 7.64 | 8.02 | 16.05 | 10.79 | 18.7 | 15.46 | 12.54 |
| WIP | 4.95 | 3.08 | 7.76 | 4.52 | 14.95 | 6.18 | 7.27 | 6.16 | 6.5 | 7.4 | 6.66 | 12.04 | 7.76 | 6.54 |
| LCN | 8.65 | 11.6 | 4.66 | 9.06 | 20.29 | 12.66 | 12.32 | 12.12 | 12.7 | 13.75 | 7.6 | 9.35 | 10.89 | 9.16 |
| WCN | 7.54 | 6.36 | 9.06 | 8.45 | 14.65 | 7.98 | 8.18 | 7.62 | 8.4 | 8.49 | 7.69 | 9.41 | 8.84 | 9.48 |
| SL | 31.62 | 38.46 | 32.82 | 39.27 | 68.56 | 35.02 | 41.06 | 38.89 | 40.52 | 47.98 | 30.61 | 56.19 | 47.83 | 42.76 |
| SW | 1.46 | 0.98 | 0.95 | 1.67 | 2.38 | 1.27 | 2.18 | 1.26 | 1.7 | 1.52 | 0.91 | 1.63 | 1.59 | 1.64 |
| LSL | 2.57 | 4.85 | 5.86 | 6.8 | 5.51 | 7.98 | 5.14 | 7.59 | 8.58 | 6.54 | 5.41 | 9.37 | 8.26 | 5.41 |
| WSL | 0.49 | 0.63 | 0.53 | 0.46 | 0.98 | 0.64 | 0.68 | 0.64 | 0.63 | 0.51 | 0.61 | 0.81 | 0.73 | 0.75 |
References
- Kaiser, R.; Tollsten, L. An introduction to the scent of cacti. Flavour Fragr. J. 1995, 10, 153–164. [Google Scholar] [CrossRef]
- Borges, R.M.; Somanathan, H.; Kelber, A. Patterns and processes in nocturnal and crepuscular pollination services. Q. Rev. Biol. 2016, 91, 389–418. [Google Scholar] [CrossRef] [PubMed]
- Macgregor, C.J.; Scott-Brown, A.S. Nocturnal pollination: An overlooked ecosystem service vulnerable to environmental change. Emer. Top. Lif. Scien. 2020, 4, 19–32. [Google Scholar] [CrossRef]
- Fleming, T.H.; Geiselman, C.; Kress, W.J. The evolution of bat pollination: A phylogenetic perspective. Ann. Bot. 2009, 104, 1017–1043. [Google Scholar] [CrossRef]
- Willmer, P. Pollination and Floral Ecology; Princeton University Press: Princeton, NJ, USA, 2011; ISBN 9781400838943. [Google Scholar]
- Rech, A.R.; Agostini, K.; Oliveira, P.E.; Machado, I.C. Biologia da Polinização, 1st ed.; Projeto Cultural: Rio de Janeiro, Brazil, 2014; ISBN 9788568126011. [Google Scholar]
- Faegri, K.; Van Der Pijl, L. Principles of Pollination Ecology, 3rd ed.; Elsevier Science: Amsterdam, The Netherlands, 1979; ISBN 9781483293035. [Google Scholar]
- Domingos-Melo, A.; Milet-Pinheiro, P.; Navarro, D.M.d.A.F.; Lopes, A.V.; Machado, I.C. It’s raining fragrant nectar in the Caatinga: Evidence of nectar olfactory signaling in bat-pollinated flowers. Ecology 2019, 101, e01640. [Google Scholar] [CrossRef]
- Rosas-Guerrero, V.; Aguilar, R.; Martén-Rodríguez, S.; Ashworth, L.; Lopezaraiza-Mikel, M.; Bastida, J.M.; Quesada, M. A Quantitative review of pollination syndromes: Do floral traits predict effective pollinators? Ecol. Lett. 2014, 17, 388–400. [Google Scholar] [CrossRef]
- Rocha, E.A.; Domingos-Melo, A.; Zappi, D.C.; Machado, I.C. Reproductive biology of columnar cacti: Are bats the only protagonists in the pollination of Pilosocereus, a typical chiropterophilous genus? Folia Geobot. 2020, 54, 239–256. [Google Scholar] [CrossRef]
- Hunt, D.; Taylor, N.; Charles, G. The New Cactus Lexicon; Dh Books: Milborne Port, UK, 2006; ISBN 9780953813445. [Google Scholar]
- Hunt, D.R.; Taylor, N.P.; Charles, G. The New Cactus Lexicon—Illustrations; Dh Books: Milborne Port, UK, 2013; ISBN 9780953813490. [Google Scholar]
- Vogel, S. Chiropterophilie in der Neotropischen Flora. Flora 1968, 157, 562–602. [Google Scholar] [CrossRef]
- Dobat, K.; Peikert-Holle, T. Blüten und Fledermäuse; Waldemar Kramer: Frankfurt, Germany, 1985; ISBN 3782910958. [Google Scholar]
- Valiente-Banuet, A.; Arizmendi, M.D.C.; Rojas-Martínez, A.; Domínguez-Canseco, L. Ecological relationships between columnar cacti and nectar-feeding bats in Mexico. J. Trop. Ecol. 1996, 12, 103–119. [Google Scholar] [CrossRef]
- Locatelli, E.; Machado, I.C.; Medeiros, P. Floral biology and bat pollination in Pilosocereus catingicola (Cactaceae) in Northeastern Brazil. Bradleya 1997, 15, 28–34. [Google Scholar] [CrossRef]
- Valiente-Banuet, A.; Rojas-Martínez, A.; Arizmendi, M.d.C.; Dávila, P. Pollination biology of two columnar cacti (Neobuxbaumia mezcalaensis and Neobuxbaumia macrocephala) in the Tehuacan Valley, Central Mexico. Am. J. Bot. 1997, 84, 452–455. [Google Scholar] [CrossRef]
- Valiente-Banuet, A.; Rojas-Martínez, A.; Casas, A.; del Coro Arizmendi, M.; Dávila, P. Pollination biology of two winter-blooming giant columnar cacti in the Tehuacán Valley, Central Mexico. J. Arid Environ. 1997, 37, 331–341. [Google Scholar] [CrossRef]
- Tschapka, M.; Helversen, O.; Barthlott, W. Bat pollination of Weberocereus tunilla, an epiphytic rain forest cactus with functional flagelliflory. Plant Biol. 1999, 1, 554–559. [Google Scholar] [CrossRef]
- Rocha, E.A.; Machado, I.C.; Zappi, D.C. Floral biology of Pilosocereus tuberculatus (Werderm.) Byles & Rowley: A bat pollinated cactus endemic from the “Caatinga” in Northeastern Brazil. Bradleya 2007, 25, 129–144. [Google Scholar] [CrossRef]
- Valiente-Banuet, A.; Santos Gally, R.; Arizmendi, M.C.; Casas, A. Pollination biology of the hemiepiphytic cactus Hylocereus undatus in the Tehuacán Valley, Mexico. J. Arid Environ. 2007, 68, 1–8. [Google Scholar] [CrossRef]
- Munguía-Rosas, M.A.; Sosa, V.J.; Jácome-Flores, M.E. Pollination system of the Pilosocereus leucocephalus columnar cactus (Tribe Cereeae) in Eastern Mexico. Plant Biol. 2009, 12, 578–586. [Google Scholar] [CrossRef]
- Mutke, J.; Burstedde, K.; Geffert, J.L.; Miebach, A.; Rafiqpoor, M.D.; Stein, A.; Barthlott, W. Patterns of diversity and endemism. Biogeography and biodiversity of cacti. Schumannia 2015, 7, 37–67. [Google Scholar]
- Martins, C.; Oliveira, R.; Filho, C.V.M.; Lopes, L.T.; Silveira, R.A.; de Silva, J.A.P.; Aguiar, L.M.S.; Antonini, Y. Reproductive biology of Cipocereus minensis (Cactaceae)—A columnar cactus endemic to rupestrian fields of a neotropical savannah. Flora 2016, 218, 62–67. [Google Scholar] [CrossRef]
- Martins, C.; Oliveira, R.; Aguiar, L.M.S.; Antonini, Y. Pollination biology of the endangered columnar cactus Cipocereus crassisepalus: A case of close relationship between plant and pollinator. Acta Bot. Bras. 2020, 34, 177–184. [Google Scholar] [CrossRef]
- Rowley, G.D. Pollination syndromes and cactus taxonomy. Bradleya 1980, 42, 95–98. [Google Scholar]
- Schlumpberger, B.O. A survey on pollination modes in cacti and a potential key innovation. In Evolution of Plant–Pollinator Interactions; Patiny, S., Ed.; Cambridge University Press: Cambridge, UK, 2012; pp. 301–319. ISBN 9780521198929. [Google Scholar]
- Tripp, E.A.; Manos, P.S. Is floral specialization an evolutionary dead-end? pollination system transitions in Ruellia (Acanthaceae). Evolution 2008, 62, 1712–1737. [Google Scholar] [CrossRef] [PubMed]
- Porsch, O. Cactaceae, – Das Bestäubungsleben der kakteenbülte II; Jahrb. D. Deutsch Kakteengesellschaft: Neudam, Germany, 1939; pp. 81–142. [Google Scholar]
- Hernandez-Hernandez, T.; Hernandez, H.M.; De-Nova, J.A.; Puente, R.; Eguiarte, L.E.; Magallon, S. Phylogenetic relationships and evolution of growth form in Cactaceae (Caryophyllales, Eudicotyledoneae). Am. J. Bot. 2011, 98, 44–61. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Taylor, N.; Zappi, D.C. Cacti of Eastern Brazil; Royal Botanic Gardens: Kew, VIC, Australia, 2004; ISBN 1842460560. [Google Scholar]
- Nyffeler, R.; Eggli, U. A farewell to dated ideas and concepts: Molecular phylogenetics and a revised suprageneric classification of the family Cactaceae. Schumannia 2010, 6, 109–149. [Google Scholar]
- Zappi, D.; Taylor, N.; Cactaceae in Flora e Funga do Brasil. Jardim Botânico do Rio de Janeiro. Available online: https://floradobrasil.jbrj.gov.br/FB70 (accessed on 15 December 2022).
- Taylor, N.P.; Zappi, D.C. An alternative view of generic delimitation and relationships in Tribe Cereeae (Cactaceae). Bradleya 1989, 7, 13–40. [Google Scholar] [CrossRef]
- Lendel, A. South American Cacti in Time and Space: Studies on the Diversification of the Tribe Cereeae, with Particular Focus on Subtribe Trichocereinae (Cactaceae). Ph.D. Dissertation, University of Zurich, Zürich, Switzerland, 2013; pp. 284p. [Google Scholar]
- Fantinati, M.R.; Soffiatti, P.; Calvente, A. A new phylogenetic hypothesis for Cereinae (Cactaceae) points to a monophyletic subtribe. Syst. Bot. 2021, 46, 689–699. [Google Scholar] [CrossRef]
- Romeiro-Brito, M.; Telhe, M.C.; Amaral, D.T.; Franco, F.F.; Moraes, E.M. A target capture probe set useful for deep- and shallow-level phylogenetic studies in Cactaceae. Genes 2022, 13, 707. [Google Scholar] [CrossRef]
- Rivera-Marchand, B.; Ackerman, J.D. Bat pollination breakdown in the caribbean columnar cactus Pilosocereus royenii. Biotropica 2006, 38, 635–642. [Google Scholar] [CrossRef]
- Rego, J.O.; Franceschinelli, E.V.; Zappi, D.C. Reproductive biology of a highly endemic species: Cipocereus laniflorus N.P. Taylor & Zappi (Cactaceae). Acta Bot. Bras. 2012, 26, 243–250. [Google Scholar] [CrossRef]
- Sampaio, E.V.S.B. Overview of the Brazilian caatinga. In Seasonally Dry Tropical Forests; Bullock, S.H., Mooney, H.A., Medina, E., Eds.; Cambridge University Press: Cambrigde, UK, 1995; pp. 35–63. ISBN 9780521112840. [Google Scholar]
- Silva, J.M.C.; Leal, I.R.; Tabarelli, M. (Eds.) Caatinga; Springer International Publishing: Cham, Switzerland, 2017; ISBN 9783319683386. [Google Scholar]
- Diaz, C.C.F.; Soares, D.B.; Ribeiro, A.S.; Santos, P.F.C. Diagnóstico ambiental do Parque Ecológico da Serra Negra, Bezerros-PE. In Os Desafios da Geografia Física na Fronteira do Conhecimento; Instituto de Geociências—UNICAMP: Campinas, Brazil, 2017; pp. 1181–1192. ISBN 9788585369163. [Google Scholar]
- Vieira, L.A.F.; Tabarelli, M.; Souza, G.; Queiroz, R.T.; Santos, B.A. Divergent herb communities in drier and chronically disturbed areas of the Brazilian Caatinga. Persp. Ecol. Conserv. 2022, 20, 132–140. [Google Scholar] [CrossRef]
- Barreto, J.M.C. Potencial Geoturístico da Região de Rio de Contas-Bahia–Brasil. Master’s Thesis, Universidade Federal da Bahia, Salvador, Brazil, 2017; pp. 164p. [Google Scholar]
- CPRM—Companhia de Pesquisa de Recursos Minerais. Projeto Mapas Municipais do Município de Morro do Chapéu; CPRM: Salvador, Bahia, Brazil, 1995. [Google Scholar]
- Dafni, A.; Kevan, P.G.; Husband, B.C. Practical Pollination Biology; Enviroquest Ltd.: Cambrigde, ON, Canada, 2005; ISBN 0968012302. [Google Scholar]
- Galetto, L.; Bernadello, G. Rewards in flowers: Nectar. In Practical Pollination Biology; Dafni, A., Kevan, P.G., Husbands, B.C., Eds.; Enviroquest: Cambridge, UK, 2005; pp. 264–313. [Google Scholar]
- Scogin, R. Nectar constituents of the Cactaceae. Southwest. Nat. 1985, 30, 77. [Google Scholar] [CrossRef]
- Zappi, D.C. Pilosocereus (Cactaceae): The Genus in Brazil; Royal Botanical Garden: Kew, VIC, Australia, 1994; ISBN 9780951723449. [Google Scholar]
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2020; Available online: https://www.R-project.org/ (accessed on 31 December 2022).
- Simmons, N.B.; Wetterer, A.L. Phylogeny and convergence in cactophilic bats. In Columnar Cacti and Their Mutualists: Evolution, Ecology, and Conservation; Fleming, T.H., Valiente-Banuet, A., Eds.; University of Arizona Press: Tucson, AZ, USA, 2022; ISBN 0816522049. [Google Scholar]
- Alcorn, S.M.; McGregor, S.E.; Butler, G.D., Jr.; Kurtz, E.B., Jr. Pollination requirements of the Saguaro (Carnegiea gigantea). Cactus Succul. J. Am. 1959, 31, 39–41. [Google Scholar]
- Sosa, M.; Soriano, P.J. Resource availability, diet and reproduction in Glossophaga longirostris (Mammalia: Chiroptera) in an arid zone of the Venezuelan Andes. J. Trop. Ecol. 1996, 12, 805–818. [Google Scholar] [CrossRef]
- McGregor, S.E.; Alcorn, S.M.; Olin, G. Pollination and pollinating agents of the Saguaro. Ecology 1962, 43, 259. [Google Scholar] [CrossRef]
- Viana, M.L.; Ortega Baes, P.; Saravia, M.; Badano, E.I.; Schlumpberger, B. Biología floral y polinizadores de Trichocereus pasacana (Cactaceae) en el Parque Nacional los Cardones, Argentina. Ver. Biol. Trop. 2001, 49, 279–285. [Google Scholar]
- Mandujano, M.d.C.; Carrillo-Angeles, I.; Martínez-Peralta, C.; Golubov, J. Reproductive biology of Cactaceae. In Desert Plants; Ramawat, K.G., Ed.; Springer: Berlin/Heidelberg, Germany, 2010; pp. 197–230. ISBN 9783642025495. [Google Scholar]
- Martén-Rodríguez, S.; Almarales-Castro, A.; Fenster, C.B. Evaluation of pollination syndromes in Antillean Gesneriaceae: Evidence for bat, hummingbird and generalized flowers. J. Ecol. 2009, 97, 348–359. [Google Scholar] [CrossRef]
- Lagomarsino, L.P.; Forrestel, E.J.; Muchhala, N.; Davis, C.C. Repeated evolution of vertebrate pollination syndromes in a recently diverged Andean plant clade. Evolution 2017, 71, 1970–1985. [Google Scholar] [CrossRef]
- Levin, D.A. The origin of reproductive isolating mechanisms in flowering plants. Taxon 1971, 20, 91–113. [Google Scholar] [CrossRef]
- Baack, E.; Melo, M.C.; Rieseberg, L.H.; Ortiz-Barrientos, D. The origins of reproductive isolation in plants. New Phytol. 2015, 207, 968–984. [Google Scholar] [CrossRef]
- Grant, V. Pollination systems as isolating mechanisms in Angiosperms. Evolution 1949, 3, 82. [Google Scholar] [CrossRef]
- Kay, K.M. Reproductive isolation between two closely related hummingbird pollinated neotropical gingers. Evolution 2006, 60, 538–552. [Google Scholar] [CrossRef]
- Celep, F.; Atalay, Z.; Dikmen, F.; Doǧan, M.; Sytsma, K.J.; Claßen-Bockhoff, R. Pollination ecology, specialization, and genetic isolation in sympatric bee-pollinated Salvia (Lamiaceae). Int. J. Plant Sci. 2020, 181, 800–811. [Google Scholar] [CrossRef]
- Muchhala, N.; Thomson, J.D. Interspecific competition in pollination systems: Costs to male fitness via pollen misplacement. Func. Ecol. 2012, 26, 476–482. [Google Scholar] [CrossRef]
- Albuquerque-Lima, S. Isolamento reprodutivo entre duas espécies quiropterófilas simpátricas de Bauhinia L. (Fabaceae). Master’s Thesis, Universidade Federal de Pernambuco, Recife, Brazil, 2019; pp. 93p. [Google Scholar]
- Queiroz, J.A.; Diniz, U.M.; Vázquez, D.P.; Quirino, Z.M.; Santos, F.A.R.; Mello, M.A.R.; Machado, I.C. Bats and hawkmoths form mixed modules with flowering plants in a nocturnal interaction network. Biotropica 2021, 53, 596–607. [Google Scholar] [CrossRef]
- Zhang, N.-N.; Yu, J.-J.; Wang, Y.-H.; Gong, X. Molecular evidence for asymmetric hybridization in three closely related sympatric species. AoB Plants 2018, 10, ply011. [Google Scholar] [CrossRef]
- Albuquerque-Lima, S.; Milet-Pinheiro, P.; Navarro, D.M.A.F.; Taylor, N.; Zappi, D.; Machado, I.C. Intermediary floral cues and morphology between natural hybrid and its parents in the Xiquexique (Cactaceae). manuscript to be submitted. Universidade Federal de Pernambuco: Recife, Brazil, 2023. [Google Scholar]
- Rowley, G.D. Spontaneous bigeneric hybrids in Cactaceae. Bradleya 1994, 12, 2–7. [Google Scholar] [CrossRef]
- Vite, F. A Natural hybrid population between Neobuxbaumia tetetzo and Cephalocereus columna-trajani (Cactaceae). J. Arid Environ. 1996, 32, 395–405. [Google Scholar] [CrossRef]
- Machado, M.C. What Is the role of hybridization in the evolution of the Cactaceae? Bradleya 2008, 26, 1–18. [Google Scholar] [CrossRef]
- Fleming, T.H.; Muchhala, N. Nectar-feeding bird and bat niches in two worlds: Pantropical comparisons of vertebrate pollination systems. J. Biogeogr. 2008, 35, 764–780. [Google Scholar] [CrossRef]
- Sazima, I.; Vogel, S.; Sazima, M. Bat pollination of Encholirium glaziovii, a terrestrial bromeliad. Plant Syst. Evol. 1989, 168, 167–179. [Google Scholar] [CrossRef]
- Prado, D.E.; Gibbs, P.E. Patterns of species distributions in the dry seasonal forests of South America. Ann. Mo. Bot. Gard. 1993, 80, 902. [Google Scholar] [CrossRef]
- Ribeiro-Silva, S.; Zappi, D.C.; Taylor, N.P.; Machado, M. Plano de Ação Nacional para Conservação das Cactáceas; Série Espécies Ameaçadas 24; Instituto Chico Mendes de Conservação da Biodiversidade, ICMBIO: Brasília, Brazil, 2011. [Google Scholar]
- Aona, L.Y.S.; Machado, M.; Pansarin, E.R.; Castro, C.C.d.; Zappi, D.; Amaral, M.d.C.E. Pollination biology of three Brazilian species of Micranthocereus Backeb. (Cereeae, Cactoideae) Endemic to the “Campos Rupestres”. Bradleya 2006, 24, 39–52. [Google Scholar] [CrossRef]
- Queiroz, J.A.; Quirino, Z.G.M.; Machado, I.C. Floral traits driving reproductive isolation of two co-flowering taxa that share vertebrate pollinators. AoB Plants 2015, 7, plv127. [Google Scholar] [CrossRef] [PubMed]
- Queiroz, J.A.; Quirino, Z.G.M.; Lopes, A.V.; Machado, I.C. Vertebrate mixed pollination system in Encholirium spectabile: A bromeliad pollinated by bats, opossum and hummingbirds in a tropical dry forest. J. Arid Environ. 2016, 125, 21–30. [Google Scholar] [CrossRef]
- Albuquerque-Lima, S.; Domingos-Melo, A.; Milet-Pinheiro, P.; Navarro, D.M.A.F.; Taylor, N.P.; Zappi, D.C.; Machado, I.C. The iconic cactus of the Caatinga dry forest, Cereus jamacaru (Cactaceae) has high sphingophily specialization and pollinator dependence. An. Acad. Bras. Cienc. Manuscript accepted. 2023. [Google Scholar]
- Primo, L.M.; Domingos-Melo, A.; Galetto, L.; Machado, I.C. Nectar secretion patterns are associated to nectar accessibility in a guild of crepuscular-nocturnal flowering plants. Plant. Ecol. 2022, 223, 951–964. [Google Scholar] [CrossRef]
- Locatelli, E.; Machado, I.C.S. Floral biology of Cereus fernambucensis: A sphingophilous cactus of restinga. Bradleya 1999, 17, 86–94. [Google Scholar] [CrossRef]
- Goettsch, B.; Hilton-Taylor, C.; Cruz-Piñón, G.; Duffy, J.P.; Frances, A.; Hernández, H.M.; Inger, R.; Pollock, C.; Schipper, J.; Superina, M.; et al. High Proportion of cactus species threatened with extinction. Nat. Plants 2015, 1, 1–7. [Google Scholar] [CrossRef]
- Silva, U.B.T.; Delgado-Jaramillo, M.; de Souza Aguiar, L.M.; Bernard, E. Species richness, geographic distribution, pressures, and threats to bats in the Caatinga drylands of Brazil. Biol. Conserv. 2018, 221, 312–322. [Google Scholar] [CrossRef]





| Cereeae Taxa | Study Sites * | Coordinates | Number of Plants | Number of Flowers | Hours of Observation |
|---|---|---|---|---|---|
| Coleocephalocereus goebelianus (Vaupel) Buining | C | 13°40′23″ S 42°04′03″ W | 7 | 7 | 8 |
| Leocereus bahiensis Britton & Rose | D | 13°35′54″ S 41°49′43″ W | 3 | 8 | 8 |
| Micranthocereus purpureus (Gürke) F.Ritter | D | 13°35′54″ S 41°49′43″ W | 4 | 4 | 5 |
| Pilosocereus catimbauensis (N.P.Taylor & Albuq.-Lima) N.P.Taylor & Albuq.-Lima | B | 8°34′34″ S 37°14′49″ W | 6 | 8 | 4 |
| P. glaucochrous (Werderm.) Byles & G.D.Rowley | E | 11°32′47″ S 41°10′34″ W | 3 | 5 | 4 |
| P. pachycladus F.Ritter subsp. pachycladus | D | 13°35′54″ S 41°49′43″ W | 3 | 6 | 4 |
| P. catingicola subsp. salvadorensis (Werderm.) Zappi | B | 8°34′34″ S 37°14′49″ W | 6 | 7 | 5 |
| P. pentaedrophorus (Cels) Byles & G.D.Rowley subsp. pentaedrophorus | A | 8°11′53″ S 35°45′34″ W | 3 | 5 | 4 |
| P. pentaedrophorus subsp. robustus Zappi | C | 13°40′23″ S 42°04′03″ W | 3 | 6 | 4 |
| Stephanocereus leucostele (Gürke) A.Berger | C | 13°40′23″ S 42°04′03″ W | 4 | 5 | 5 |
| S. luetzelburgii (Vaupel) N.P.Taylor & Eggli | D | 13°35′54″ S 41°49′43″ W | 3 | 7 | 6 |
| Xiquexique gounellei (F.A.C.Weber) Lavor & Calvente | C | 13°40′23″ S 42°04′03″ W | 10 | 15 | 8 |
| X. ×heptagonus N.P.Taylor & Albuq.-Lima. | B | 8°34′34″ S 37°14′49″ W | 6 | 15 | 12 |
| X. tuberculatus (Werderm.) Lavor & Calvente | B | 8°34′34″ S 37°14′49″ W | 10 | 15 | 16 |
| Characteristics |
|---|
| 1. Flower length (FL) |
| 2. Flower width (FW) |
| 3. Flower tube opening diameter (OP) |
| 4. Pericarpel length (PL) |
| 5. Pericarpel width (PW) |
| 6. Length of the floral tube (LFT) |
| 7. Width of the widest portion of the floral tube (WFT) |
| 8. Length of the external elements of the perianth (LEP) |
| 9. Width of the external elements of the perianth (WEP) |
| 10. Length of the internal elements of the perianth (LIP) |
| 11. Width of the internal elements of the perianth (WIP) |
| 12. Length of the nectar chamber (LCN) |
| 13. Width of the nectar chamber (WCN) |
| 14. Style length (SL) |
| 15. Style width (SW) |
| 16. Length of the stigma lobes (LSL) |
| 17. Width of the stigma lobes (WSL) |
| Cereinae Taxa | Habit * | Cephalium | Floral Form | Symmetry | External Colour | Internal Colour | No. of Flowers ** |
|---|---|---|---|---|---|---|---|
| Coleocephalocereusgoebelianus | sg | pre | tub | act | Pale beige | White | 2–4 |
| Leocereusbahiensis | ere/ss | abs | tub | zyg | Green | White | 1–3 |
| Micranthocereuspurpureus | cbb | pre | cam | act | Magenta | White | 1–2 |
| Pilosocereuscatimbauensis | sg/cbb | pre | cam | zyg | Olive green | White | 1–2 |
| P. glaucochrous | tr | abs | tuc | zyg | Magenta | White | 1–3 |
| P. pachycladus subsp. pachycladus | tr | abs | cam | zyg | Olive green | White | 1–3 |
| P. catingicola subsp. salvadorensis | tr | abs | cam | zyg | Blue | White | 1–5 |
| P. pentaedrophorus subsp. pentaedrophorus | tr | abs | cam | zyg | Green | White | 1–3 |
| P. pentaedrophorus subsp. robustus | tr | abs | cam | zyg | Green | White | 1–3 |
| Stephanocereusleucostele | cab | pre | tuc | act | Green | White | 1–7 |
| S. luetzelburgii | bot | pre | cam | act | Reddish | White | 1–4 |
| Xiquexiquegounellei | can | abs | tub | zyg | Beige | White | 1–5 |
| X. ×heptagonus | can | abs | tub | zyg | Dark magenta | White | 1–7 |
| X. tuberculatus | can | abs | tub | zyg | Olive green | White | 1–5 |
| Traits * | Axis 1 | Axis 2 |
|---|---|---|
| FL | 0.81 | - |
| FW | 0.84 | 0 |
| OP | 0.83 | 0.04 |
| PL | 0.68 | 0.1 |
| PW | 0.83 | 0.03 |
| LFT | 0.61 | 0.18 |
| WFT | 0.81 | 0.11 |
| LEP | 0.8 | 0 |
| WEP | 0.77 | 0 |
| LIP | 0.75 | 0.02 |
| WIP | 0.73 | 0.04 |
| LCN | 0.46 | 0.13 |
| WCN | 0.63 | 0.06 |
| SL | 0.81 | 0.06 |
| SW | 0.39 | 0.07 |
| LSL | 0.09 | 0.55 |
| WSL | 0.4 | 0.02 |
| Traits * | Kruskal–Wallis | Dunn Test ** | ||||||
|---|---|---|---|---|---|---|---|---|
| FL | H(13) = 96.81 | p < 0.001 | goe-cag | goe-gou | goe-hep | cag-lue | lue-gou | |
| FW | H(13) = 96.76 | p < 0.001 | goe-cag | goe-gou | bah-cag | bah-gou | ||
| OP | H(13) = 76.84 | p < 0.001 | goe-cag | bah-cag | ||||
| PL | H(13) = 76.16 | p < 0.001 | goe-cag | cag-lue | ||||
| PW | H(13) = 80.23 | p < 0.001 | cag-lue | |||||
| LFT | H(13) = 89.91 | p < 0.001 | goe-gou | cag-lue | lue-gou | |||
| WFT | H(13) = 91.42 | p < 0.001 | goe-cag | bah-cag | cag-lue | cag-tub | ||
| LEP | H(13) = 73.47 | p < 0.001 | bah-cag | |||||
| WEP | H(13) = 81.1 | p < 0.001 | goe-cag | goe-gou | bah-cag | bah-gou | cat-cag | |
| LIP | H(13) = 99.93 | p < 0.001 | bah-cag | bah-gou | cat-cag | cat-gou | ||
| WIP | H(13) = 87.13 | p < 0.001 | goe-cag | goe-gou | bah-cag | bah-gou | cat-cag | cat-gou |
| LCN | H(13) = 72.18 | p < 0.001 | pur-cag | |||||
| WCN | H(13) = 49.54 | p < 0.001 | bah-cag | |||||
| SL | H(13) = 85.46 | p < 0.001 | goe-cag | goe-gou | cag-lue | lue-gou | ||
| SW | H(13) = 64.02 | p < 0.001 | cag-lue | |||||
| LSL | H(13) = 76.42 | p < 0.001 | goe-gou | goe-hep | ||||
| WSL | H(13) = 55.99 | p < 0.001 | cat-cag | |||||
| Cereinae Taxa | Nectar Volume (µL) | Nectar Concentration (%) | Energy (Calories/Grams) | Number of Bat Visits | Ratio Number of Visits/Hour |
|---|---|---|---|---|---|
| Coleocephalocereusgoebelianus | 132 ± 50 | 17.5 ± 0.25 | 90.09 | 24 | 3 h |
| Leocereusbahiensis | 175 ± 33 | 20 ± 1.7 | 136.5 | 29 | 3.6 h |
| Micranthocereuspurpureus | 212.5 ± 53 | 19 ± 1.4 | 157.09 | 3 | 0.6 h |
| Pilosocereuscatimbauensis | 339.5 ± 50 | 20 ± 0.5 | 264.42 | 20 | 5 h |
| Pilosocereusglaucochrous | 250 ± 60 | 21 ± 0.9 | 204.75 | 3 | 0.75 h |
| P. pachycladus subsp. pachycladus | 688.30 ± 207 | 20 ± 1.3 | 536.64 | 4 | 1 h |
| P. catingicola subsp. salvadorensis | 1331.25 ± 486.25 | 20 ± 4.5 | 908.40 | 8 | 1.6 h |
| P. pentaedrophorus subsp. pentaedrophorus | 240 ± 26.7 | 21 ± 0.6 | 196.56 | 7 | 1.75 h |
| P. pentaedrophorus subsp. robustus | 228 ± 55.5 | 20 ± 0.9 | 177.84 | 19 | 4.75 h |
| Stephanocereusleucostele | 388.125 ± 146 | 19 ± 0.9 | 265.3 | 13 | 2.6 h |
| S. luetzelburgii | 202.85 ± 44.70 | 19.20 ± 1.02 | 151.25 | 1 | 0.16 h |
| Xiquexiquegounellei | 365 ± 94 * | 23 ± 11 * | 327.40 | 7 | 0.87 h |
| X. ×heptagonus | 256 ± 28.24 | 21.9 ± 4.8 | 218.6 | 22 | 1.46 h |
| X. tuberculatus | 213.3 * | 12.3 * | 102.17 | 25 | 1.66 h |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Albuquerque-Lima, S.; Taylor, N.P.; Zappi, D.C.; Machado, I.C. Floral Specialization and Bat Pollination in Subtribe Cereinae (Cactaceae): A Morphological Approach. Diversity 2023, 15, 207. https://doi.org/10.3390/d15020207
Albuquerque-Lima S, Taylor NP, Zappi DC, Machado IC. Floral Specialization and Bat Pollination in Subtribe Cereinae (Cactaceae): A Morphological Approach. Diversity. 2023; 15(2):207. https://doi.org/10.3390/d15020207
Chicago/Turabian StyleAlbuquerque-Lima, Sinzinando, Nigel P. Taylor, Daniela C. Zappi, and Isabel C. Machado. 2023. "Floral Specialization and Bat Pollination in Subtribe Cereinae (Cactaceae): A Morphological Approach" Diversity 15, no. 2: 207. https://doi.org/10.3390/d15020207