
Isotopic Space of the House Mouse in the Gradient of Anthropogenic Habitats

 , ,
, , 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Study Sites and Sampling
2.2. Stable Isotope Analysis of Hair
2.3. Data Analyses
3. Results
3.1. House Mouse in Small Mammal Communities
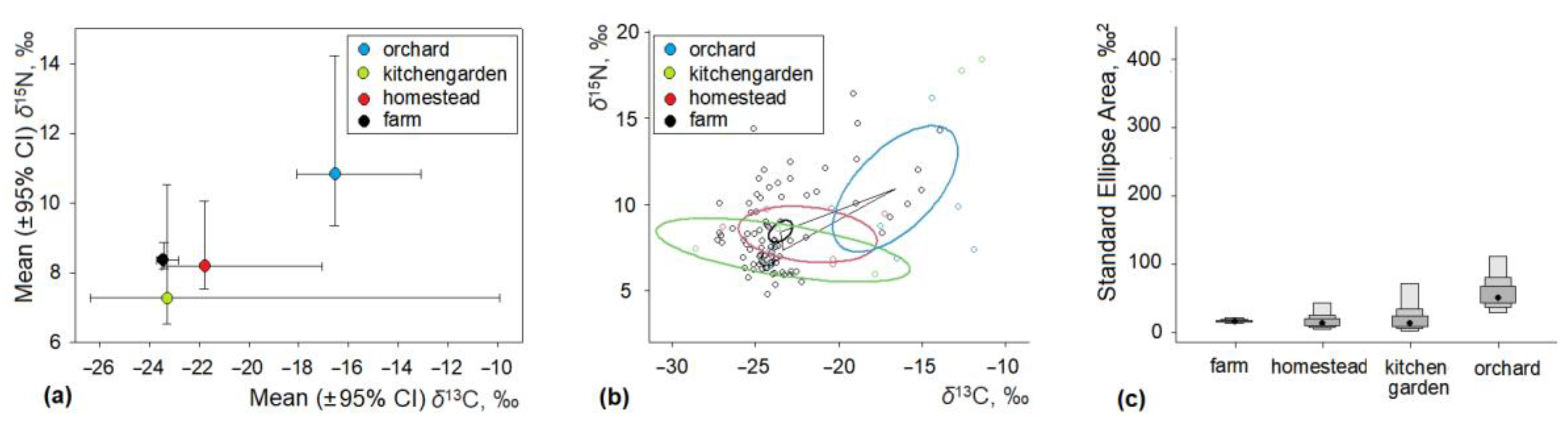
3.2. Stable Isotope Ratios of House Mouse in Anthropogenic Habitats
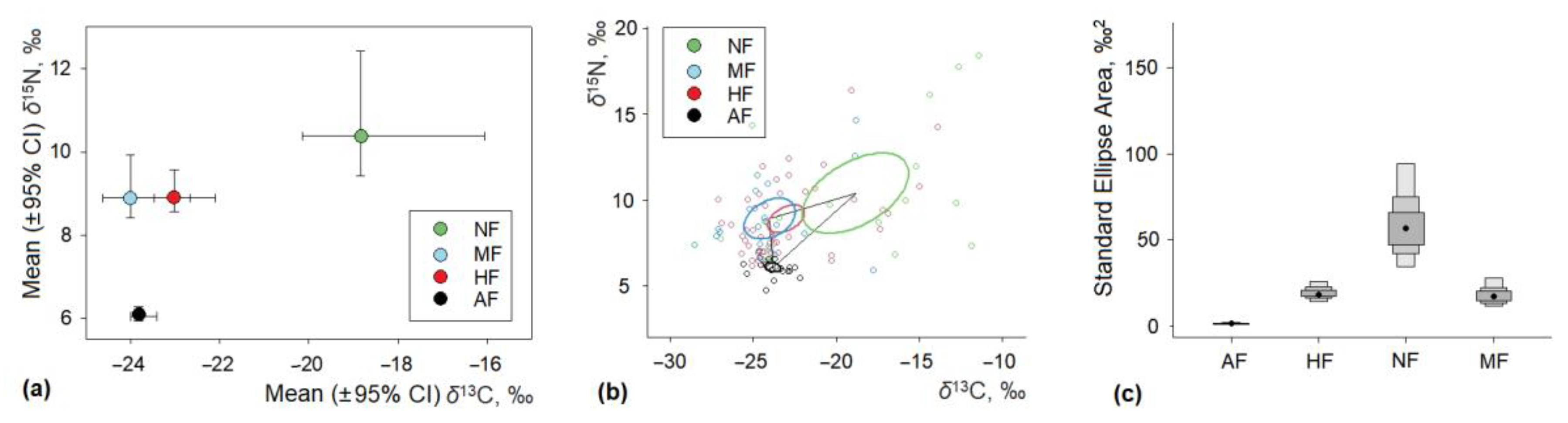
3.3. Intraspecific Differences of Isotopic Ratios in House Mice
3.4. Seasonal Differences of Isotopic Ratios in House Mice
4. Discussion
5. Conclusions
- The main factors, responsible for the diet variation are the season and the source of available food.
- The dietary niches of mice eating natural food in orchards were completely separated from those of mice eating livestock feed on farms, the latter having the lowest dietary variation.
- Based on isotopic data, all mice (males, females, adults, subadults and juveniles) were likely to consume similar diets.
- M. musculus has much wider dietary niche and consumes more foods of animal origin than the other synoptically living omnivore, the bank vole.
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Musser, G.; Hutterer, R.; Kryštufek, B.; Yigit, N.; Mitsainas, G. Mus musculus (amended version of 2016 assessment). IUCN Red List. Threat. Species 2021, e.T13972A197519724. [Google Scholar] [CrossRef]
- Wilson, D.E.; Lacher, T.E.; Mittermeier, R.A. Handbook of the Mammals of the World: Rodents II; Wilson, D.E., Lacher, T.E., Mittermeier, R.A., Eds.; Lynx Edicions: Barcelona, Spain, 2017; p. 789. [Google Scholar]
- Mazza, V.; Dammhahn, M.; Lösche, E.; Eccard, J.A. Small mammals in the big city: Behavioural adjustments of non-commensal rodents to urban environments. Glob. Chang. Biol. 2020, 26, 6326–6337. [Google Scholar] [CrossRef] [PubMed]
- Khlyap, L.A.; Warshavsky, A.A. Synanthropic and agrophilic rodents as invasive alien mammals. Russ. J. Biol. Invasions 2010, 1, 301–312. [Google Scholar] [CrossRef]
- Polito, M.J.; Robinson, B.; Warzybok, P.; Bradley, R.W. Population Dynamics and Resource Availability Drive Seasonal Shifts in the Consumptive and Competitive Impacts of Introduced House Mice (Mus musculus) on an Island Ecosystem. bioRxiv. 2022, 10, e13904. [Google Scholar] [CrossRef] [PubMed]
- Macholán, M. Mus Musculus; Academic Press: London, UK, 1999. [Google Scholar]
- Berry, R.J.; Jakobson, M.E. Adaptation and adaptability in wild-living house mice (Mus musculus). J. Zool. 1975, 176, 391–402. [Google Scholar] [CrossRef]
- Pocock, M.J.; Searle, J.B.; White, P.C. Adaptations of animals to commensal habitats: Population dynamics of house mice Mus musculus domesticus on farms. J. Anim. Ecol. 2004, 73, 878–888. [Google Scholar] [CrossRef]
- Gomez, D.; Provensal, C.; Polop, J. Microhabitat use by the house mouse Mus musculus in an urban area. Acta Theriol. 2009, 54, 183–192. [Google Scholar] [CrossRef]
- Häberle, S.; Schäfer, M.; Soteras, R.; Martínez-Grau, H.; Hajdas, I.; Jacomet, S.; Röder, B.; Schibler, J.; van Willigen, S.; Antolín, F. Small Animals, Big Impact? Early Farmers and Pre- and Post-Harvest Pests from the Middle Neolithic Site of Les Bagnoles in the South-East of France (L’Isle-sur-la-Sorgue, Vaucluse, Provence-Alpes-Côte-d’Azur). Animals 2022, 12, 1511. [Google Scholar] [CrossRef] [PubMed]
- Prūsaitė, J. (Comp.). Fauna of Lithuania. In Mammals; Mokslas: Vilnius, Lithuania, 1988; p. 295. [Google Scholar]
- Balčiauskas, L.; Trakimas, G.; Juškaitis, R.; Ulevičius, A.; Balčiauskienė, L. Atlas of Lithuanian Mammals, Amphibians and Reptiles, 2nd ed.; Akstis: Vilnius, Lithuania, 1999; p. 47. [Google Scholar]
- Balčiauskas, L.; Juškaitis, R. Diversity of small mammal communities in Lithuania (1. A review). Acta Zool. Litu. 1997, 7, 29–45. [Google Scholar] [CrossRef]
- Balčiauskas, L.; Balčiauskienė, L. Small Mammal Diversity Changes in a Baltic Country, 1975–2021: A Review. Life 2022, 12, 1887. [Google Scholar] [CrossRef]
- Balčiauskas, L.; Balčiauskienė, L.; Garbaras, A.; Stirkė, V. Diversity and Diet Differences of Small Mammals in Commensal Habitats. Diversity 2021, 13, 346. [Google Scholar] [CrossRef]
- Atkočaitis, O. Sodybos pastatuose sugauti smulkieji žinduoliai. Theriol. Litu. 2003, 3, 57–61. [Google Scholar]
- Balčiauskienė, L.; Balčiauskas, L.; Vitkauskas, V.; Podėnas, S. Indoor small mammals in Lithuania: Some morphometrical, body condition, and reproductive characteristics. Zool. Ecol. 2015, 25, 305–313. [Google Scholar] [CrossRef]
- Bronson, F.H. The reproductive ecology of the house mouse. Q. Rev. Biol. 1979, 54, 265–299. [Google Scholar] [CrossRef]
- Latham, N.; Mason, G. From house mouse to mouse house: The behavioural biology of free-living Mus musculus and its implications in the laboratory. Appl. Anim. Behav. Sci. 2004, 86, 261–289. [Google Scholar] [CrossRef] [Green Version]
- Russell, J.C.; Meyer, J.-Y.; Holmes, N.D.; Pagad, S. Invasive alien species on islands: Impacts, distribution, interactions and management. Environ. Conserv. 2017, 44, 359–370. [Google Scholar] [CrossRef] [Green Version]
- Watts, C.; Innes, J.; Wilson, D.J.; Thornburrow, D.; Bartlam, S.; Fitzgerald, N.; Cave, V.; Smale, M.; Barker, G.; Padamsee, M. Do mice matter? Impacts of house mice alone on invertebrates, seedlings and fungi at Sanctuary Mountain Maungatautari. N. Z. J. Ecol. 2022, 46, 3472. [Google Scholar] [CrossRef]
- Shiels, A.B.; Flores, C.A.; Khamsing, A.; Krushelnycky, P.D.; Mosher, S.M.; Drake, D.R. Dietary niche differentiation among three species of invasive rodents (Rattus rattus, R. exulans, Mus musculus). Biol. Invasions 2013, 15, 1037–1048. [Google Scholar] [CrossRef]
- Norbury, G.; Wilson, D.J.; Clarke, D.; Hayman, E.; Smith, J.; Howard, S. Density-impact functions for invasive house mouse (Mus musculus) effects on indigenous lizards and invertebrates. Biol. Invasions 2022. [Google Scholar] [CrossRef]
- Simberloff, D. Rats are not the only introduced rodents producing ecosystem impacts on islands. Biol. Invasions 2009, 11, 1735–1742. [Google Scholar] [CrossRef]
- Russell, J.C.; Peace, J.E.; Houghton, M.J.; Bury, S.J.; Bodey, T.W. Systematic prey preference by introduced mice exhausts the ecosystem on Antipodes Island. Biol. Invasions 2020, 2, 1265–1278. [Google Scholar] [CrossRef] [Green Version]
- Balčiauskas, L.; Skipitytė, R.; Balčiauskienė, L.; Jasiulionis, M. Resource partitioning confirmed by isotopic signatures allows small mammals to share seasonally flooded meadows. Ecol. Evol. 2019, 9, 5479–5489. [Google Scholar] [CrossRef]
- Ruffino, L.; Russell, J.C.; Pisanu, B.; Caut, S.; Vidal, E. Low individual-level dietary plasticity in an island-invasive generalist forager. Pop. Ecol. 2011, 53, 535–548. [Google Scholar] [CrossRef]
- Ribeiro, J.F.; Guaraldo, A.; Nardoto, G.B.; Santoro, G.; Vieira, E.M. Habitat type and seasonality influence the isotopic trophic niche of small mammals in a neotropical savanna. Hystrix 2019, 30, 3038. [Google Scholar] [CrossRef]
- Abt, K.F.; Bock, W.F. Seasonal variations of diet composition in farmland field mice Apodemus spp. and bank voles Clethrionomys glareolus. Acta Theriol. 1998, 43, 379–389. [Google Scholar] [CrossRef] [Green Version]
- Pisanu, B.; Caut, S.; Gutjar, S.; Vernon, P.; Chapuis, J.-L. Introduced black rats (Rattus rattus) on Ile de la Possession (Iles Crozet, Subantarctic): Diet and trophic position in food webs. Polar Biol. 2011, 34, 169–180. [Google Scholar] [CrossRef]
- Pinho, C.J.; Lopes, E.P.; Paupério, J.; Gomes, I.; Romeiras, M.M.; Vasconcelos, R. Trust your guts? The effect of gut section on diet composition and impact of Mus musculus on islands using metabarcoding. Ecol. Evol. 2022, 12, e8638. [Google Scholar] [CrossRef] [PubMed]
- Symes, C.T.; Wilson, J.W.; Woodborne, S.M.; Shaikh, Z.S.; Scantlebury, M. Resource partitioning of sympatric small mammals in an A frican forest-grassland vegetation mosaic. Austral Ecol. 2013, 38, 721–729. [Google Scholar] [CrossRef]
- Baltensperger, A.P.; Huettmann, F.; Hagelin, C.; Welker, J.M. Quantifying trophic niche spaces of small mammals using stable isotopes (δ15N and δ13C) at two scales across Alaska. Can. J. Zool. 2015, 93, 579–588. [Google Scholar] [CrossRef] [Green Version]
- Phillips, D.L. Converting isotope values to diet composition: The use of mixing models. J. Mammal. 2012, 93, 342–352. [Google Scholar] [CrossRef] [Green Version]
- Caut, S.; Angulo, E.; Courchamp, F. Variation in discrimination factors (Δ15N and Δ13C): The effect of diet isotopic values and applications for diet reconstruction. J. Appl. Ecol. 2009, 46, 443–453. [Google Scholar] [CrossRef]
- Jackson, A.L.; Inger, R.; Parnell, A.C.; Bearhop, S. Comparing isotopic niche widths among and within communities: SIBER—Stable Isotope Bayesian Ellipses in R. J. Anim. Ecol. 2011, 80, 595–602. [Google Scholar] [CrossRef]
- Hwang, Y.T.; Millar, J.S.; Longstaffe, F.J. Do δ15N and δ13C values of feces reflect the isotopic composition of diets in small mammals? Can. J. Zool. 2007, 85, 388–396. [Google Scholar] [CrossRef]
- Kurle, C.M.; Koch, P.L.; Tershy, B.R.; Croll, D.A. The effects of sex, tissue type, and dietary components on stable isotope discrimination factors (Δ13C and Δ15N) in mammalian omnivores. Isot. Environ. Health Stud. 2014, 50, 307–321. [Google Scholar] [CrossRef] [Green Version]
- Chibowski, P.; Brzeziński, M.; Suska-Malawska, M.; Zub, K. Diet/Hair and Diet/Faeces Trophic Discrimination Factors for Stable Carbon and Nitrogen Isotopes, and Hair Regrowth in the Yellow-Necked Mouse and Bank Vole. Ann. Zool. Fenn. 2022, 59, 171–185. [Google Scholar] [CrossRef]
- Calandra, I.; Labonne, G.; Mathieu, O.; Henttonen, H.; Lévêque, J.; Milloux, M.; Renvoisé, É.; Montuire, S.; Navarro, N. Isotopic partitioning by small mammals in the subnivium. Ecol. Evol. 2015, 5, 4132–4140. [Google Scholar] [CrossRef] [PubMed]
- Tieszen, L.L.; Boutton, T.W.; Tesdahl, K.G.; Slade, N.A. Fractionation and turnover of stable carbon isotopes in animal tissues: Implications for δ13C analysis of diet. Oecologia 1983, 57, 32–37. [Google Scholar] [CrossRef]
- Sponheimer, M.; Robinson, T.; Ayliffe, L.; Passey, B.; Roeder, B.; Shipley, L.; Lopez, E.; Cerling, T.; Dearing, D.; Ehleringer, J. An experimental study of carbon-isotope fractionation between diet, hair, and feces of mammalian herbivores. Can. J. Zool. 2003, 81, 871–876. [Google Scholar] [CrossRef]
- Schwertl, M.; Auerswald, K.; Schnyder, H. Reconstruction of the isotopic history of animal diets by hair segmental analysis. Rapid Commun. Mass Sp. 2003, 17, 1312–1318. [Google Scholar] [CrossRef]
- Plikus, M.V.; Chuong, C.M. Complex hair cycle domain patterns and regenerative hair waves in living rodents. J. Investig. Dermatol. 2008, 128, 1071–1080. [Google Scholar] [CrossRef] [Green Version]
- Tejada-Lara, J.V.; MacFadden, B.J.; Bermudez, L.; Rojas, G.; Salas-Gismondi, R.; Flynn, J.J. Body mass predicts isotope enrichment in herbivorous mammals. Proc. R. Soc. B 2018, 285, 20181020. [Google Scholar] [CrossRef] [Green Version]
- Rioux, È.; Pelletier, F.; St-Laurent, M.H. Influence of lipids on stable isotope ratios in mammal hair: Highlighting the importance of validation. Ecosphere 2019, 10, e02723. [Google Scholar] [CrossRef] [Green Version]
- Taylor, D.; DeJesus, E.J.; Novak, M.; Terry, R.C. The effects of formalin fixation and fluid storage on stable isotopes in rodent hair. J. Mammal. 2021, 102, 1635–1647. [Google Scholar] [CrossRef]
- Smiley, T.M.; Cotton, J.M.; Badgley, C.; Cerling, T.E. Small-mammal isotope ecology tracks climate and vegetation gradients across western North America. Oikos 2015, 125, 1100–1109. [Google Scholar] [CrossRef]
- Balčiauskas, L.; Skipitytė, R.; Jasiulionis, M.; Trakimas, G.; Balčiauskienė, L.; Remeikis, V. The impact of Great Cormorants on biogenic pollution of land ecosystems: Stable isotope signatures in small mammals. Sci. Total Environ. 2016, 565, 376–383. [Google Scholar] [CrossRef] [PubMed]
- Balčiauskas, L.; Skipitytė, R.; Garbaras, A.; Stirkė, V.; Balčiauskienė, L.; Remeikis, V. Isotopic Niche of Syntopic Granivores in Commercial Orchards and Meadows. Animals 2021, 11, 2375. [Google Scholar] [CrossRef] [PubMed]
- Balčiauskas, L.; Skipitytė, R.; Garbaras, A.; Stirkė, V.; Balčiauskienė, L.; Remeikis, V. Stable Isotopes Reveal the Dominant Species to Have the Widest Trophic Niche of Three Syntopic Microtus Voles. Animals 2021, 11, 1814. [Google Scholar] [CrossRef]
- Balčiauskas, L.; Stirkė, V.; Garbaras, A.; Skipitytė, R.; Balčiauskienė, L. Stable Isotope Analysis Supports Omnivory in Bank Voles in Apple Orchards. Agriculture 2022, 12, 1308. [Google Scholar] [CrossRef]
- Moors, P.J. Norway rats (Rattus norvegicus) on the Noises and Motukawao islands, Hauraki Gulf, New Zealand. N. Z. J. Ecol. 1985, 8, 37–54. Available online: https://www.jstor.org/stable/24052744 (accessed on 9 December 2022).
- The Kolmogorov-Smirnov Test of Normality. Available online: https://www.socscistatistics.com/tests/kolmogorov/default.aspx (accessed on 9 December 2022).
- Hulme-Beaman, A.; Dobney, K.; Cucchi, T.; Searle, J.B. An ecological and evolutionary framework for commensalism in anthropogenic environments. Trends Ecol. Evol. 2016, 31, 633–645. [Google Scholar] [CrossRef] [Green Version]
- Gomez, M.D.; Provensal, M.C.; Polop, J.J. Effect of interspecific competition on Mus musculus in an urban area. J. Pest Sci. 2008, 81, 235–240. [Google Scholar] [CrossRef]
- Balčiauskas, L.; Balčiauskienė, L. On the doorstep, rodents in homesteads and kitchen gardens. Animals 2020, 10, 856. [Google Scholar] [CrossRef] [PubMed]
- Evans, J.C.; Lindholm, A.K.; König, B. Long-term overlap of social and genetic structure in free-ranging house mice reveals dynamic seasonal and group size effects. Curr. Zool. 2021, 67, 59–69. [Google Scholar] [CrossRef]
- Souquet, L.; Chevret, P.; Ganem, G.; Auffray, J.C.; Ledevin, R.; Agret, S.; Hautier, L.; Renaud, S. Back to the wild: Does feralization affect the mandible of non-commensal house mice (Mus musculus domesticus)? Biol. J. Linn. Soc. 2019, 126, 471–486. [Google Scholar] [CrossRef]
- Fairley, J.S.; Smal, C.M. Feral house mice in Ireland. Ir. Nat. J. 1987, 22, 284–290. Available online: https://www.jstor.org/stable/25539173 (accessed on 9 December 2022).
- Delong, K.T. Population Ecology of Feral House Mice: Interference by Microtus. Ecology 1966, 47, 481–484. [Google Scholar] [CrossRef]
- Canova, L.; Fasola, M. Communities of small mammals in six biotopes of northern Italy. Acta Theriol. 1991, 36, 73–86. Available online: https://rcin.org.pl/ibs/dlibra/publication/edition/11774 (accessed on 9 December 2022). [CrossRef] [Green Version]
- Chalmers, R.; Sturdee, A.; Bull, S.; Miller, A.; Wright, S.E. The prevalence of Cryptosporidium parvum and C. muris in Mus domesticus, Apodemus sylvaticus and Clethrionomys glareolus in an agricultural system. Parasitol. Res. 1997, 83, 478–482. [Google Scholar] [CrossRef]
- Michalski, A.; Niemcewicz, M.; Bielawska-Drózd, A.; Nowakowska, A.; Gaweł, J.; Pitucha, G.; Joniec, J.; Zielonka, K.; Marciniak-Niemcewicz, A.; Kocik, J. Surveillance of hantaviruses in Poland: A study of animal reservoirs and human hantavirus disease in Subcarpathia. Vector-Borne Zoonot. 2014, 14, 514–522. [Google Scholar] [CrossRef] [Green Version]
- Le Roux, V.; Chapuis, J.L.; Frenot, Y.; Vernon, P. Diet of the house mouse (Mus musculus) on Guillou Island, Kerguelen archipelago, Subantarctic. Polar Biol. 2002, 25, 49–57. [Google Scholar] [CrossRef]
- Rowe, F.P.; Swinney, T.; Quy, R.J. Reproduction of the House mouse (Mus musculus) in farm buildings. J. Zool. 2009, 199, 259–269. [Google Scholar] [CrossRef]
- Vadell, M.V.; Villafañe, I.E.G.; Cavia, R. Are life-history strategies of Norway rats (Rattus norvegicus) and house mice (Mus musculus) dependent on environmental characteristics? Wildlife Res. 2014, 41, 172–185. [Google Scholar] [CrossRef]
- Gomez, M.D.; Priotto, J.; Provensal, M.C.; Steinmann, A.; Castillo, E.; Polop, J.J. A population study of house mice (Mus musculus) inhabiting different habitats in an Argentine urban area. Int. Biodeter. Biodegr. 2008, 62, 270–273. [Google Scholar] [CrossRef]
- León, V.A.; Fraschina, J.; Guidobono, J.S.; Busch, M. Habitat use and demography of Mus musculus in a rural landscape of Argentina. Integr. Zool. 2012, 8, 18–29. [Google Scholar] [CrossRef]
- Sked, S.; Liu, C.; Abbar, S.; Corrigan, R.; Cooper, R.; Wang, C. The Spatial Distribution of the House Mouse, Mus musculus domesticus, in Multi-Family Dwellings. Animals 2022, 12, 197. [Google Scholar] [CrossRef]
- Pocock, M.J.O.; Hauffe, H.C.; Searle, J.B. Dispersal in house mice. Biol. J. Linn. Soc. 2005, 84, 565–583. [Google Scholar] [CrossRef] [Green Version]
- Vrbanec, L.; Matijević, V.; Guenther, A. Enhanced problem-solving ability as an adaptation to urban environments in house mice. Proc. R. Soc. B 2021, 288, 20202504. [Google Scholar] [CrossRef]
- Boitani, L.; Loy, A.; Molinari, P. Temporal and spatial displacement of two sympatric rodents (Apodemus sylvaticus and Mus musculus) in a Mediterranean coastal habitat. Oikos 1985, 45, 246–252. [Google Scholar] [CrossRef]
- de la Peña, N.M.; Butet, A.; Delettre, Y.; Paillat, G.; Morant, P.; Le Du, L.; Burel, F. Response of the small mammal community to changes in western French agricultural landscapes. Landscape Ecol. 2003, 18, 265–278. [Google Scholar] [CrossRef]
- Csanády, A.; Kisková, J.; Galušková, S.; Duranková, S.; Pristaš, P.; Sedláková-Kaduková, J. The house mouse (Mus musculus) in small farmstead buildings in Slovakia. Biologia 2021, 76, 1205–1213. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Site No | Year | Habitat | N | Males | Females | Adults | Subadults | Juveniles |
|---|---|---|---|---|---|---|---|---|
| 1 | 2022 | Apple orchard | 1 | 1 | 0 | 0 | 0 | 1 |
| 2 | 2022 | Apple orchard | 2 | 2 | 0 | 0 | 2 | 0 |
| 2 | 2018, 2021 | Meadow | 4 | 2 | 2 | 2 | 0 | 2 |
| 3 | 2021–2022 | Farm | 65 | 43 | 22 | 38 | 12 | 15 |
| 4 | 2018 | Currant plantation | 1 | 1 | 0 | 0 | 0 | 1 |
| 5 | 2022 | Homestead | 1 | 1 | 0 | 0 | 1 | 0 |
| 6 | 2018–2019 | Raspberry plantation | 2 | 1 | 1 | 1 | 1 | 0 |
| 7 | 2022 | Homestead | 4 | 2 | 2 | 1 | 0 | 3 |
| 8 | 2022 | Kitchen garden | 3 | 1 | 2 | 2 | 1 | 0 |
| 9 | 2018 | Farm | 21 | 11 | 10 | 10 | 3 | 8 |
| 10 | 2019 | Currant plantation | 1 | 1 | 0 | 1 | 0 | 0 |
| In total | 105 | 66 | 39 | 55 | 20 | 30 |
| Habitat | Year | N | S | H | P% | RA |
|---|---|---|---|---|---|---|
| Meadow | 2018–2022 | 152 | 10 | 1.48 a | 3.3 a (CI = 1.1–7.5) | 0.05 |
| Apple orchard | 2018–2022 | 134 | 8 | 1.35 a | 2.2 a (CI = 0.5–6.4) | 0.03 |
| Currant plantation | 2018–2019 | 92 | 6 | 0.96 b | 2.2 a (CI = 0.3–7.6) | 0.02 |
| Raspberry plantation | 2018–2019 | 7 | 3 | 0.96 b | 28.6 b (CI = 3.8–71.0) | 0.19 |
| Kitchen garden | 2019–2022 | 361 | 9 | 1.24 c | 0.8 a (CI = 0.2–2.4) | n/a |
| Homestead | 2019–2022 | 645 | 11 | 1.36 a | 0.9 a (CI = 0.3–2.0) | n/a |
| Farm | 2018, 2021–2022 | 289 | 6 | 1.26 c | 59.2 b (CI = 53.3–64.9) | 6.67 |
| Habitat | N | δ13C, ‰ | δ15N, ‰ | ||||
|---|---|---|---|---|---|---|---|
| Mean ± SE | Min–Max | Range | Mean ± SE | Min–Max | Range | ||
| Orchard and plantation | 10 | –16.55 ± 1.53 | −24.09–−11.42 | 12.67 | 10.84 ± 1.49 | 6.42–18.40 | 11.98 |
| Kitchen garden | 3 | –23.28 ± 3.11 | −28.51–−17.75 | 10.76 | 7.29 ± 0.76 | 5.93–8.55 | 2.62 |
| Homestead | 5 | –21.79 ± 1.70 | −26.88–−17.19 | 9.69 | 8.20 ± 0.67 | 6.49–9.67 | 3.18 |
| Farm | 86 | –23.42 ± 0.29 | −27.21–−13.89 | 13.32 | 8.37 ± 0.26 | 4.72–16.41 | 11.69 |
| Habitat | N | δ13C, ‰ | δ15N, ‰ | ||||
|---|---|---|---|---|---|---|---|
| Mean ± SE | Min–Max | Range | Mean ± SE | Min–Max | Range | ||
| Homestead | 56 | −25.91 ± 0.11 | −27.90–−23.98 | 3.92 | 5.94 ± 0.26 | 2.44–10.70 | 8.26 |
| Kitchen garden | 5 | −25.40 ± 0.28 | −27.43–−24.57 | 2.86 | 6.14 ± 0.26 | 4.78–7.06 | 2.28 |
| Orchard and plantation | 155 | −23.63 ± 0.08 | −26.00–−28.55 | 2.55 | 5.84 ± 0.16 | –1.62–11.30 | 2.28 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Balčiauskas, L.; Garbaras, A.; Stirkė, V.; Skipitytė, R.; Balčiauskienė, L. Isotopic Space of the House Mouse in the Gradient of Anthropogenic Habitats. Diversity 2023, 15, 173. https://doi.org/10.3390/d15020173
Balčiauskas L, Garbaras A, Stirkė V, Skipitytė R, Balčiauskienė L. Isotopic Space of the House Mouse in the Gradient of Anthropogenic Habitats. Diversity. 2023; 15(2):173. https://doi.org/10.3390/d15020173
Chicago/Turabian StyleBalčiauskas, Linas, Andrius Garbaras, Vitalijus Stirkė, Raminta Skipitytė, and Laima Balčiauskienė. 2023. "Isotopic Space of the House Mouse in the Gradient of Anthropogenic Habitats" Diversity 15, no. 2: 173. https://doi.org/10.3390/d15020173