
An Overview of Marine Non-Indigenous Species Found in Three Contrasting Biogeographic Metropolitan French Regions: Insights on Distribution, Origins and Pathways of Introduction

 , , , , , , , , , , , , , , , , , add
Show full author list
, , , , , , , , , , , , , , , , , add
Show full author list
Abstract
:1. Introduction
2. Methodology
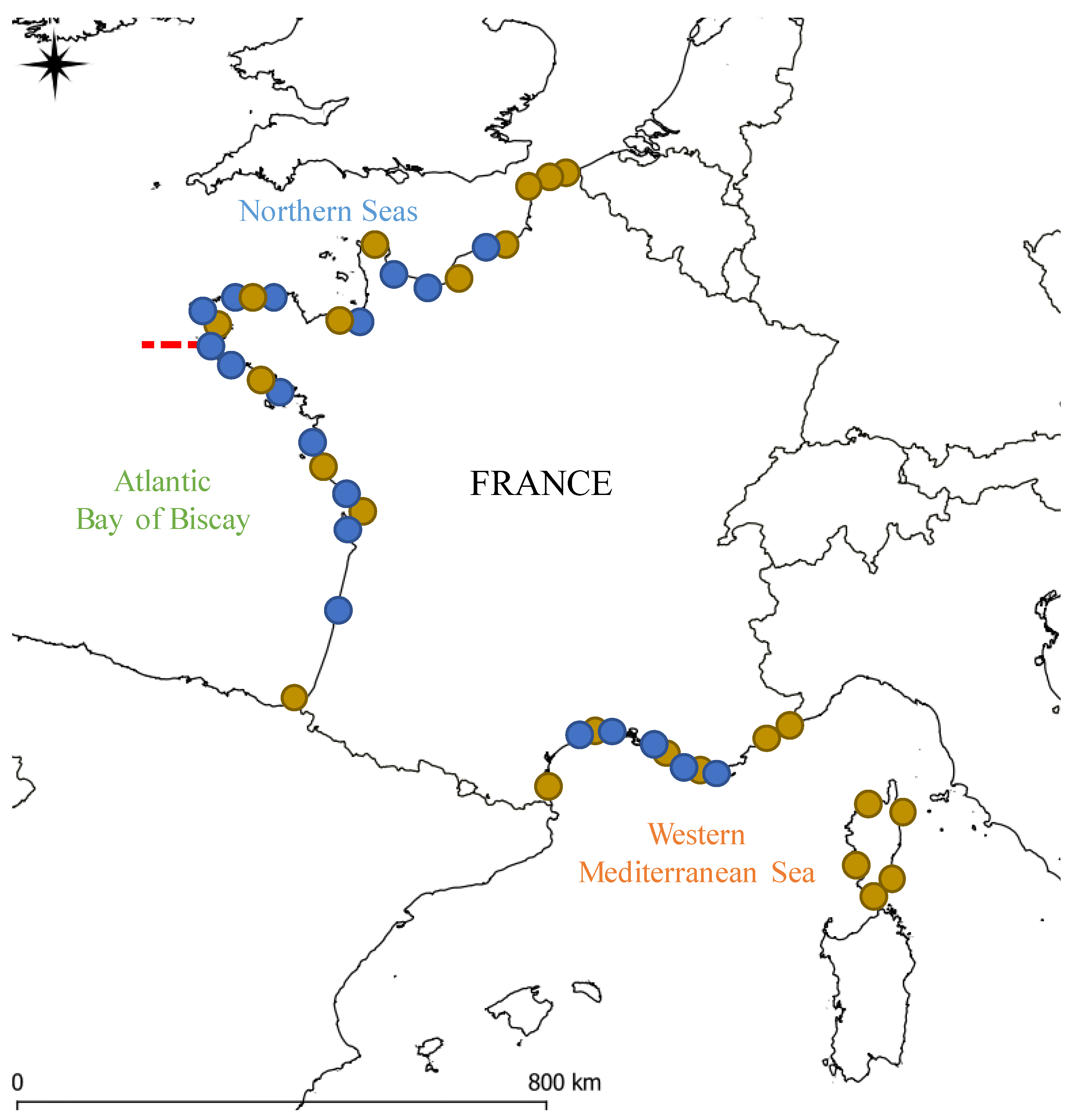
2.1. Geographic Coverage
2.2. Data Included
2.3. Species Attributes
3. Results
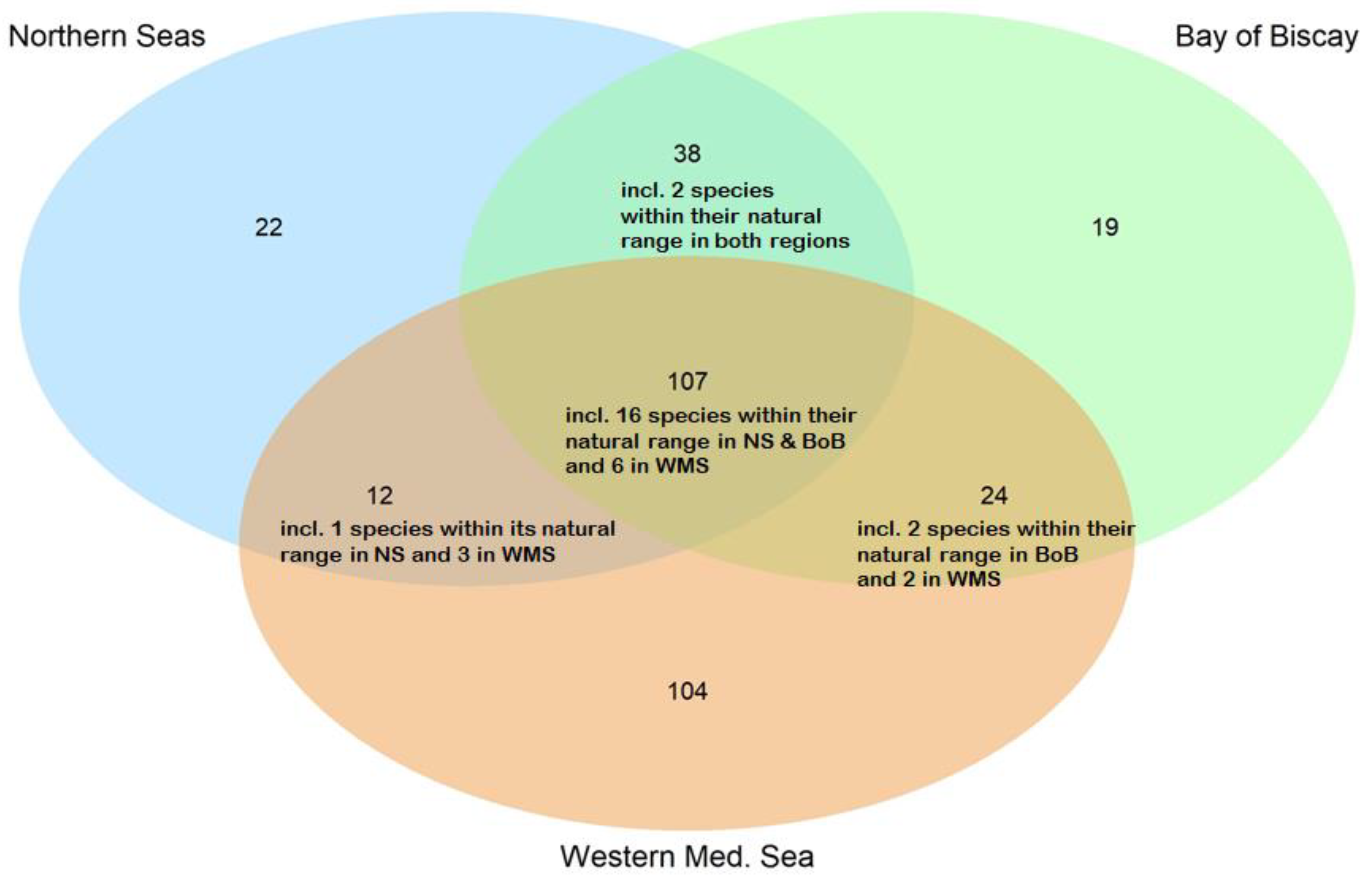
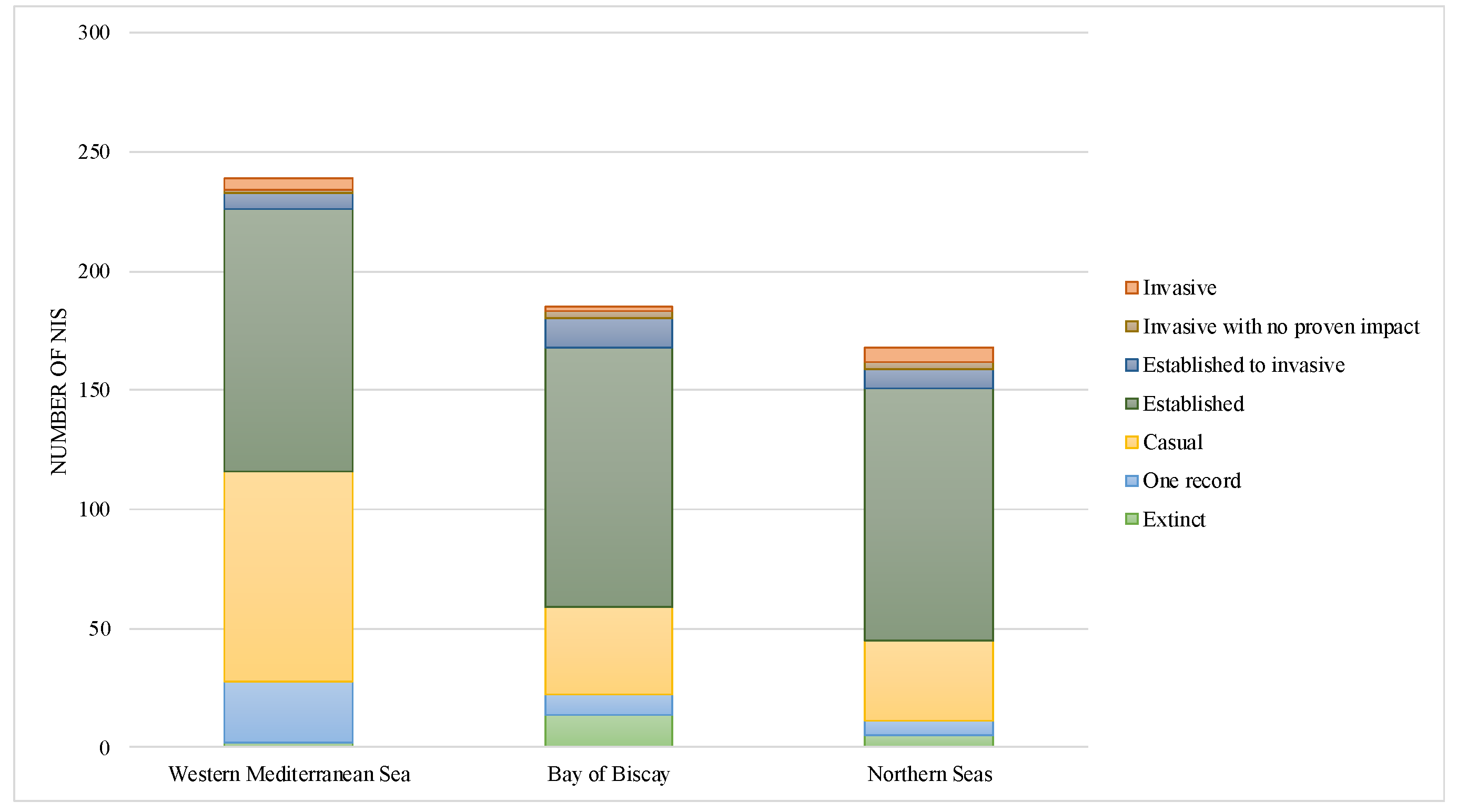
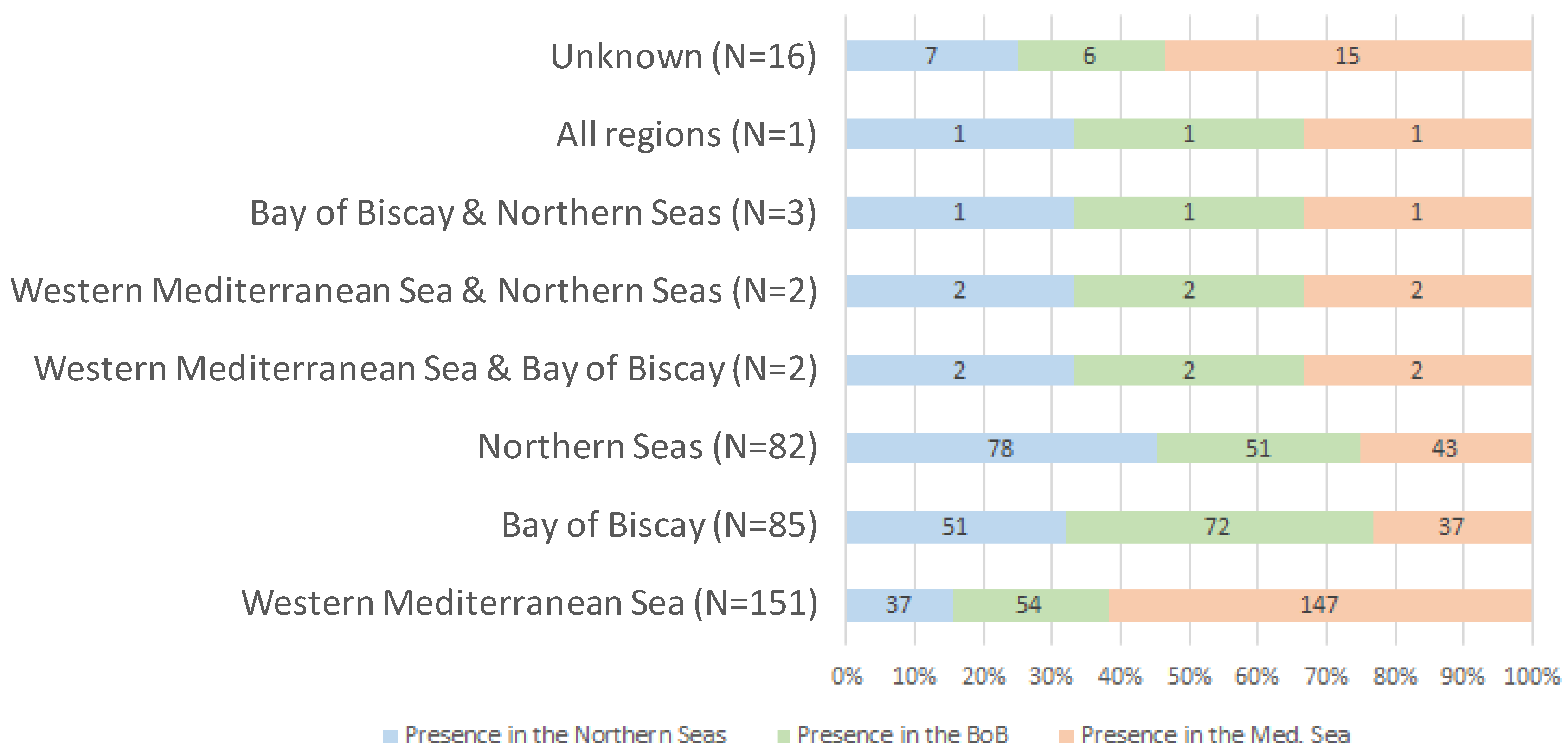
3.1. NIS Number, Taxonomic Identity (over France and per Region) and Status
- Western Mediterranean Sea
- Bay of Biscay
- Northern Seas
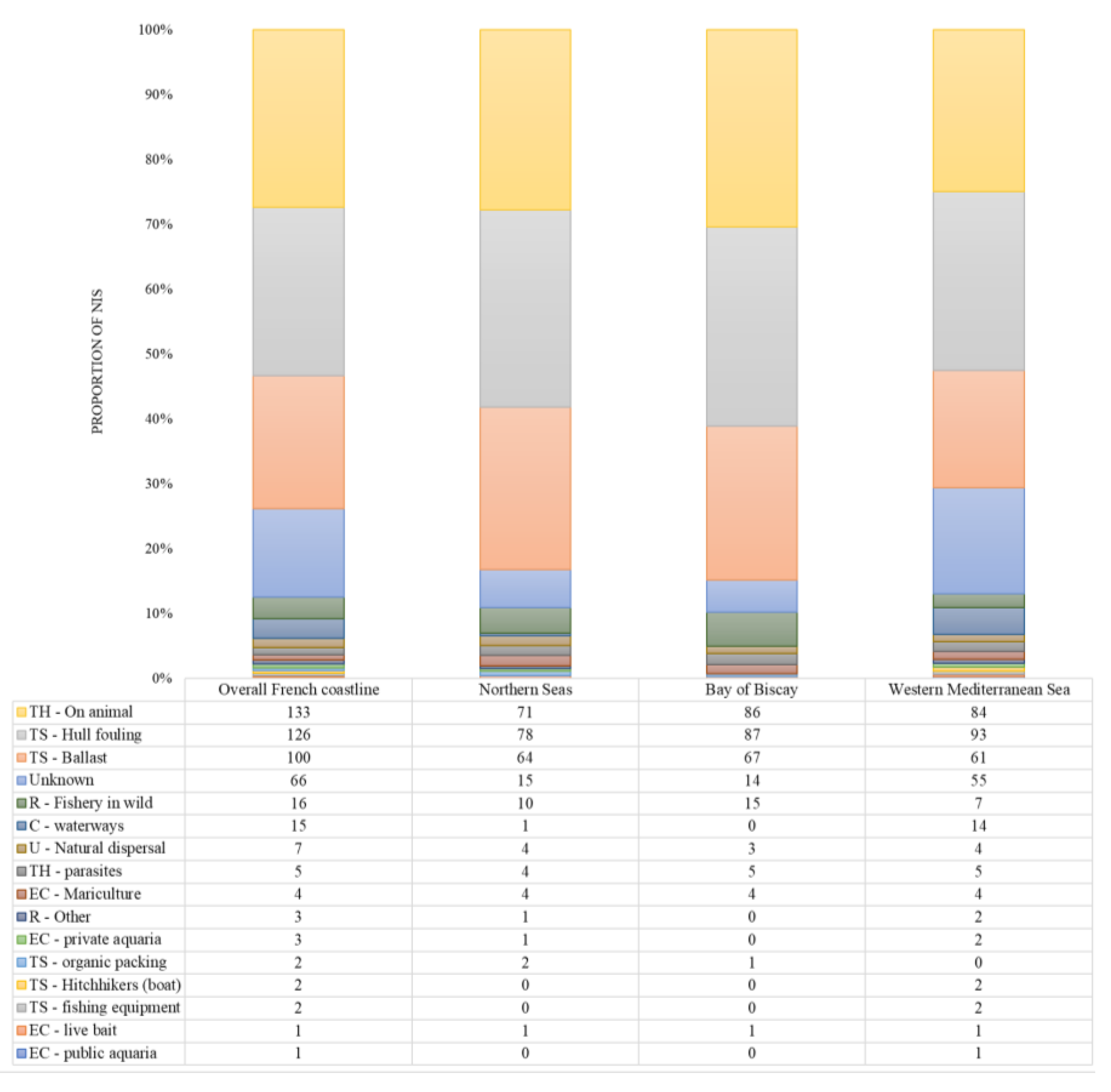
3.2. Reported Pathways and Vectors of Introduction
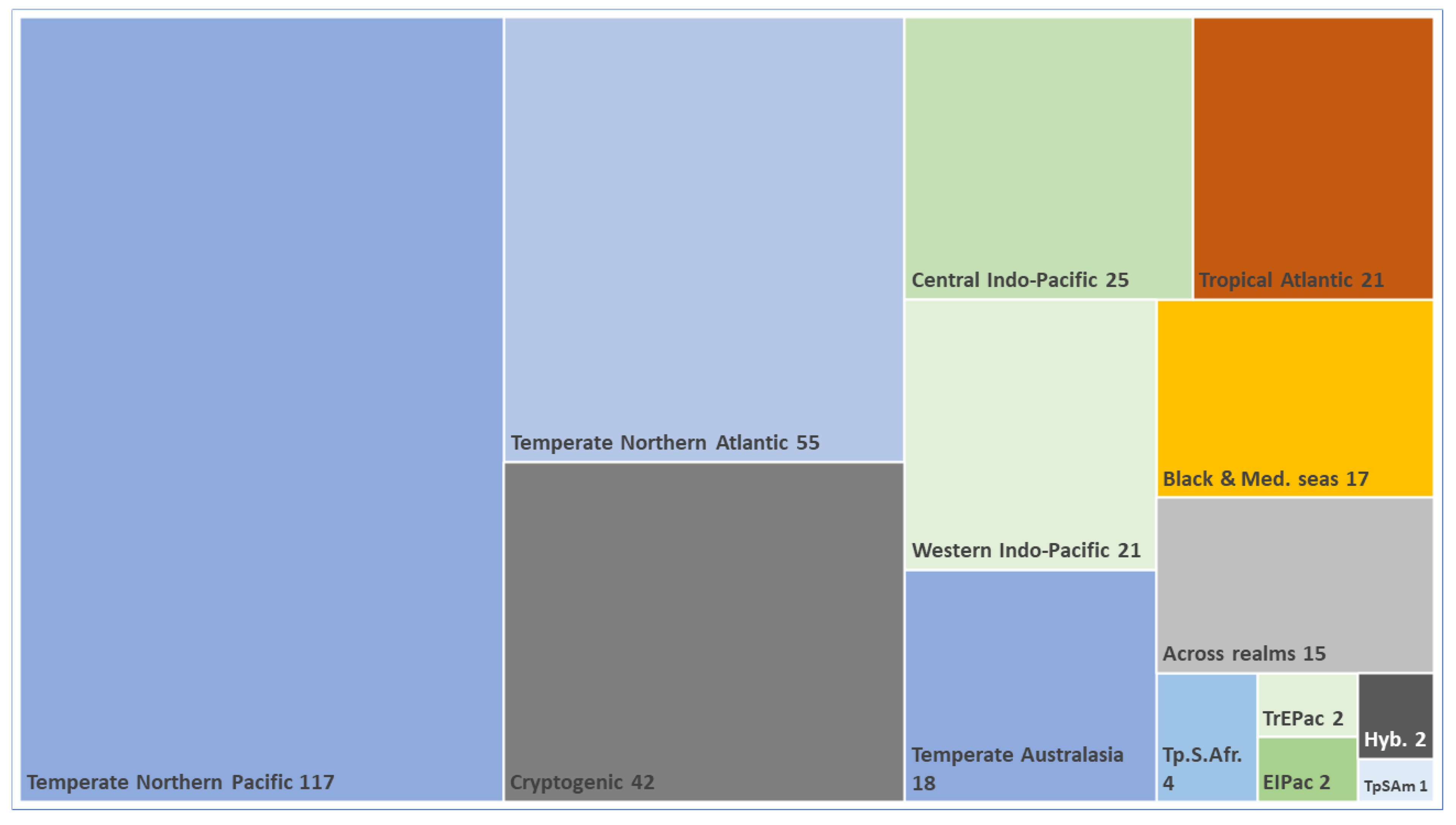
3.3. Biogeographic Origin of Introduced Species
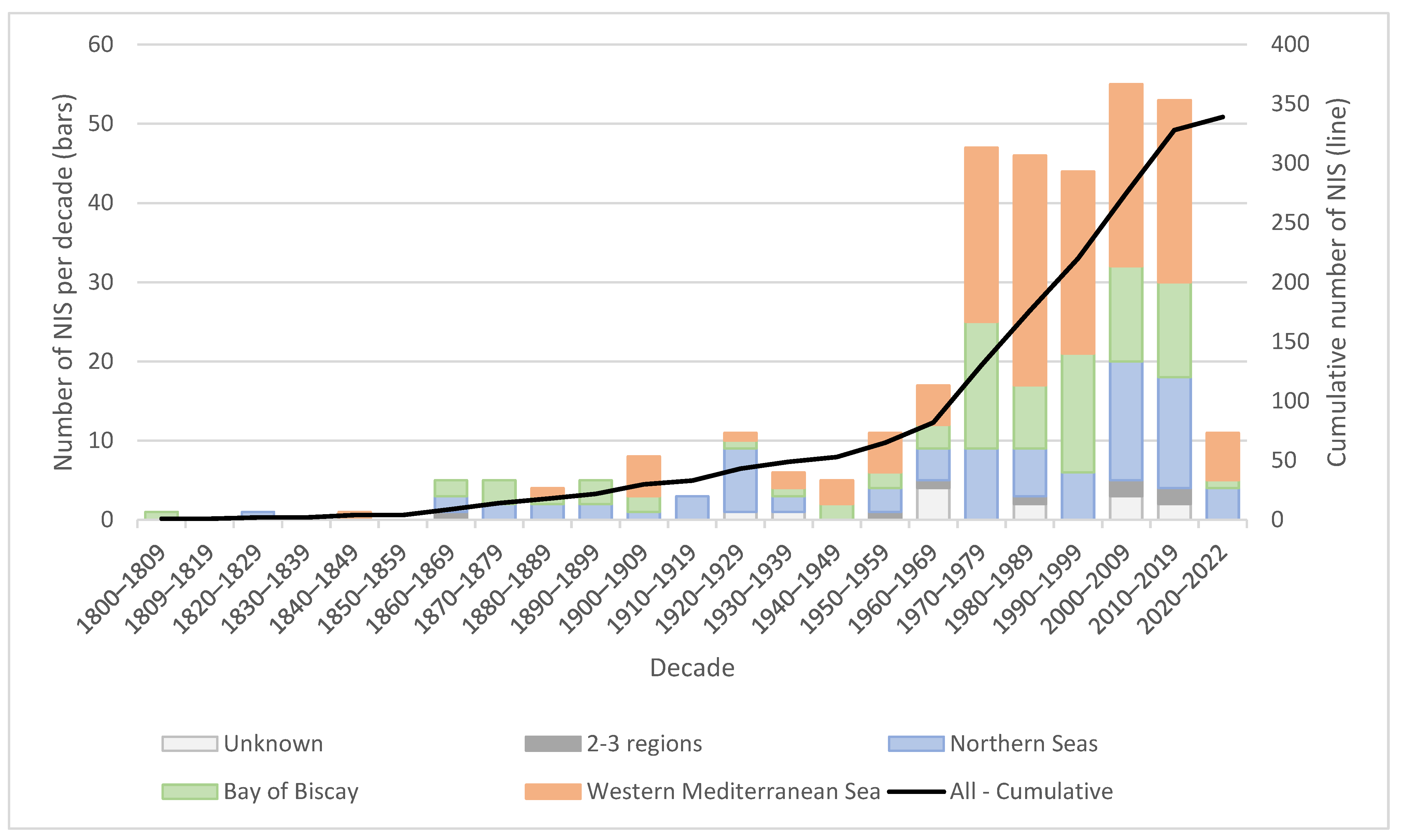
3.4. Temporal Trends in Primary Detections
4. Discussion
4.1. Update and Trends
4.2. Vectors
4.3. Exclusions and Uncertainties about Species Numbers and Identification
4.4. Available Tools to Limit this Invasion Phenomenon
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Seebens, H.; Bacher, S.; Blackburn, T.M.; Capinha, C.; Dawson, W.; Dullinger, S.; Genovesi, P.; Hulme, P.E.; Kleunen, M.; Kühn, I.; et al. Projecting the continental accumulation of alien species through to 2050. Glob. Chang. Biol. 2020, 27, 970–982. [Google Scholar] [CrossRef] [PubMed]
- Bailey, S.A.; Brown, L.; Campbell, M.L.; Canning-Clode, J.; Carlton, J.T.; Castro, N.; Chainho, P.; Chan, F.T.; Creed, J.C.; Curd, A.; et al. Trends in the detection of aquatic non-indigenous species across global marine, estuarine and freshwater ecosystems: A 50-year perspective. Divers. Distrib. 2020, 26, 1780–1797. [Google Scholar] [CrossRef]
- Capinha, C.; Essl, F.; Seebens, H.; Moser, D.; Pereira, H.M. The dispersal of alien species redefines biogeography in the Anthropocene. Science 2015, 348, 1248–1251. [Google Scholar] [CrossRef] [PubMed]
- Ruiz, G.M.; Rawlings, T.K.; Dobbs, F.C.; Drake, L.A.; Mullady, T.; Huq, A.; Colwell, R.R. Global spread of microorganisms by ships. Nature 2000, 408, 49–50. [Google Scholar] [CrossRef]
- Diagne, C.; Leroy, B.; Vaissière, A.-C.; Gozlan, R.E.; Roiz, D.; Jarić, I.; Salles, J.-M.; Bradshaw, C.J.A.; Courchamp, F. High and rising economic costs of biological invasions worldwide. Nature 2021, 592, 571–576. [Google Scholar] [CrossRef]
- IPBES; Brondizio, S.; Diaz, S.; Settele, J.; Ngo, H.T. Global Assessment Report on Biodiversity and Ecosystem Services of the Intergovernmental Science-Policy Platform on Biodiversity and Ecosystem Services; Zenodo: Bonn, Germany, 2019; p. 1144. [Google Scholar]
- Ojaveer, H.; Galil, B.S.; Carlton, J.T.; Alleway, H.; Goulletquer, P.; Lehtiniemi, M.; Marchini, A.; Miller, W.; Occhipinti-Ambrogi, A.; Peharda, M.; et al. Historical baselines in marine bioinvasions: Implications for policy and management. PLoS ONE 2018, 13, e0202383. [Google Scholar] [CrossRef] [Green Version]
- European Commission. Directive 2008/56/EC of the European Parliament and of the Council of 17 June 2008 Establishing a Framework for Community Action in the Field of Marine Environmental Policy (Marine Strategy Framework Directive) (Text with EEA Relevance); European Commission: Brussels, Belgium, 2008. [Google Scholar]
- Tsiamis, K.; Palialexis, A.; Connor, D.; Antoniadis, S.; Bartilotti, C.; Bartolo, G.A.; Berggreen, U.C.; Boschetti, S.; Buschbaum, C.; Canning-Clode, J.; et al. Marine Strategy Framework Directive, Descriptor 2, Non-Indigenous Species: Delivering Solid Recommendations for Setting Threshold Values for Non-Indigenous Species Pressure on European Seas; Publications Office of the European Union: Luxembourg, 2021; p. 36. [Google Scholar]
- Zenetos, A.; Tsiamis, K.; Galanidi, M.; Carvalho, N.; Bartilotti, C.; Canning-Clode, J.; Castriota, L.; Chainho, P.; Comas-González, R.; Costa, A.C.; et al. Status and Trends in the Rate of Introduction of Marine Non-Indigenous Species in European Seas. Diversity 2022, 14, 1077. [Google Scholar] [CrossRef]
- Spalding, M.D.; Fox, H.E.; Allen, G.R.; Davidson, N.; Ferdaña, Z.A.; Finlayson, M.; Halpern, B.S.; Jorge, M.A.; Lombana, A.; Lourie, S.A.; et al. Marine Ecoregions of the World: A Bioregionalization of Coastal and Shelf Areas. Bioscience 2007, 57, 573–583. [Google Scholar] [CrossRef] [Green Version]
- Tsiamis, K.; Palialexis, A.; Stefanova, K.; Gladan, N.; Skejić, S.; Despalatović, M.; Cvitković, I.; Dragičević, B.; Dulčić, J.; Vidjak, O.; et al. Non-indigenous species refined national baseline inventories: A synthesis in the context of the European Union's Marine Strategy Framework Directive. Mar. Pollut. Bull. 2019, 145, 429–435. [Google Scholar] [CrossRef]
- Le Fèvre, J. Aspects of the Biology of Frontal Systems. Adv. Mar. Biol. 1987, 23, 163–299. [Google Scholar] [CrossRef]
- Gallon, R.K.; Lavesque, N.; Grall, J.; Labrune, C.; Gremare, A.; Bachelet, G.; Blanchet, H.; Bonifácio, P.; Bouchet, V.M.; Dauvin, J.-C.; et al. Regional and latitudinal patterns of soft-bottom macrobenthic invertebrates along French coasts: Results from the RESOMAR database. J. Sea Res. 2017, 130, 96–106. [Google Scholar] [CrossRef] [Green Version]
- Boudouresque, C.F.; Klein, J.; Ruitton, S.; Verlaque, M. Biological Invasion: The Thau Lagoon, a Japanese Biological Island in the Mediterranean Sea. In Proceedings of the Global Change: Mankind-Marine Environment Interactions; Ceccaldi, H.-J., Dekeyser, I., Girault, M., Stora, G., Eds.; Springer: Dordrecht, The Netherlands, 2011; pp. 151–156. [Google Scholar]
- Nunes, A.L.; Katsanevakis, S.; Zenetos, A.; Cardoso, A.C. Gateways to alien invasions in the European seas. Aquat. Invasions 2014, 9, 133–144. [Google Scholar] [CrossRef]
- Ulman, A.; Ferrario, J.; Forcada, A.; Seebens, H.; Arvanitidis, C.; Occhipinti-Ambrogi, A.; Marchini, A. Alien species spreading via biofouling on recreational vessels in the Mediterranean Sea. J. Appl. Ecol. 2019, 56, 2620–2629. [Google Scholar] [CrossRef]
- Póvoa, A.A.; Skinner, L.F.; de Araújo, F.V. Fouling organisms in marine litter (rafting on abiogenic substrates): A global review of literature. Mar. Pollut. Bull. 2021, 166, 112189. [Google Scholar] [CrossRef]
- Johnson, L.E.; Brawley, S.H.; Adey, W.H. Secondary spread of invasive species: Historic patterns and underlying mechanisms of the continuing invasion of the European rockweed Fucus serratus in eastern North America. Biol. Invasions 2011, 14, 79–97. [Google Scholar] [CrossRef]
- Kelly, N.E.; Wantola, K.; Weisz, E.; Yan, N.D. Recreational boats as a vector of secondary spread for aquatic invasive species and native crustacean zooplankton. Biol. Invasions 2012, 15, 509–519. [Google Scholar] [CrossRef]
- Floc’h, J.-Y.; Pajot, R.; Mouret, V. Undaria pinnatifida (Laminariales, Phaeophyta) 12 Years after Its Introduction into the Atlantic Ocean. In Proceedings of the Fifteenth International Seaweed Symposium; Lindstrom, S.C., Chapman, D.J., Eds.; Springer: Dordrecht, The Netherlands, 1996; pp. 217–222. [Google Scholar]
- Epstein, G.; Smale, D.A. Undaria pinnatifida: A case study to highlight challenges in marine invasion ecology and management. Ecol. Evol. 2017, 7, 8624–8642. [Google Scholar] [CrossRef]
- Goulletquer, P.; Bachelet, G.; Sauriau, P.-G.; Noel, P. Open Atlantic Coast of Europe—A Century of Introduced Species into French Waters. In Invasive Aquatic Species of Europe; Leppäkoski, E., Gollash, S., Olenin, S., Eds.; Springer: Dordrecht, The Netherlands, 2002; pp. 276–290. [Google Scholar]
- Goulletquer, P. Guide Des Organismes Exotiques Marins; Éditions Belin: Paris, France, 2016; ISBN 978-2-7011-9020-4. [Google Scholar]
- Blanchard, M.; Goulletquer, P.; Hamon, D.; Le Mao, P.; Nezan, E.; Gentil, F.; Simon, N.; Viard, F.; Ar Gall, E.; Grall, J.; et al. Liste des Espèces Marines Introduites dans Les Eaux Bretonnes et des Espèces Introduites Envahissantes des Eaux Périphériques; Bretagne Environnement, France, 2010. 2010. Available online: https://www.researchgate.net/publication/235963754_Les_especes_marines_animales_et_vegetales_introduites_dans_le_Bassin_Artois-Picardie (accessed on 21 December 2022).
- Dewarumez, J.-M.; Gevaert, F.; Massé, C.; Foveau, A.; Desroy, N.; Grulois, D. Les Espèces Marines Animales et Végétales Introduites dans le Bassin Artois-Picardie; UMR CNRS 8187 LOG et Agence de l’Eau Artois-Picardie: Douai, France, 2011; pp. 140. [Google Scholar]
- Pezy, J.-P.; Baffreau, A.; Dauvin, J.-C. Records of two introduced Penaeidae (Crustacea: Decapoda) species from Le Havre Harbour, France, English Channel. Biol. Invasions Rec. 2017, 6, 363–367. [Google Scholar] [CrossRef]
- ICES Working Group on Introductions and Transfers of Marine Organisms (WGITMO); ICES Scientific Reports; ICES: Copenhagen, Denmark, 2022; Volume 4, pp. 209. [CrossRef]
- Bishop, J.D.D.; Roby, C.; Yunnie, A.L.E.; Wood, C.A.; Lévêque, L.; Turon, X.; Viard, F. The Southern Hemisphere ascidian Asterocarpa humilis is unrecognised but widely established in NW France and Great Britain. Biol. Invasions 2012, 15, 253–260. [Google Scholar] [CrossRef] [Green Version]
- Patarnello, T.; Volckaert, F.A.M.J.; Castilho, R. Pillars of Hercules: Is the Atlantic-Mediterranean transition a phylogeographical break? Mol. Ecol. 2007, 16, 4426–4444. [Google Scholar] [CrossRef]
- Harrower, C.; Scalera, R.; Pagad, S.; Schönrogge, K.; Roy, H. Guidance for Interpretation of the CBD Categories of Pathways for the Introduction of Invasive Alien Species; Publications Office of the European Union: Luxembourg, 2018; 79p. [Google Scholar]
- Katsanevakis, S.; Gatto, F.; Zenetos, A.; Cardoso, A.C. How many marine aliens in Europe? Manag. Biol. Invasions 2013, 4, 37–42. [Google Scholar] [CrossRef] [Green Version]
- Nellemann, C.; Hain, S.; Alder, J. Dead Water—Merging of Climate Change with Pollution, Over-Harvest, and Infestations in the World’s Fishing Grounds; United Nations Environment Programme, GRID-Arendal; UNEP/Earthprint: Oslo, Norway, 2008; 62p. [Google Scholar]
- Tsiamis, K.; Zenetos, A.; Deriu, I.; Gervasini, E.; Cardoso, A.C. The native distribution range of the European marine non-indigenous species. Aquat. Invasions 2018, 13, 187–198. [Google Scholar] [CrossRef]
- Jouffray, J.B.; Blasiak, R.; Norström, A.V.; Österblom, H.; Nyström, M. The Blue Acceleration: The Trajectory of Human Expansion into the Ocean. ONE Earth 2020, 2, 43–54. [Google Scholar] [CrossRef] [Green Version]
- Chainho, P.; Fernandes, A.; Amorim, A.; Ávila, S.P.; Canning-Clode, J.; Castro, J.J.; Costa, A.C.; Costa, J.L.; Cruz, T.; Gollasch, S.; et al. Non-indigenous species in Portuguese coastal areas, coastal lagoons, estuaries and islands. Estuar. Coast. Shelf Sci. 2015, 167, 199–211. [Google Scholar] [CrossRef]
- Minchin, D.; Cook, E.J.; Clark, P.F. Alien species in British brackish and marine waters. Aquat. Invasions 2013, 8, 3–19. [Google Scholar] [CrossRef]
- Crooks, J.A. Lag times and exotic species: The ecology and management of biological invasions in slow-motion1. Écoscience 2005, 12, 316–329. [Google Scholar] [CrossRef]
- Bock, D.G.; Caseys, C.; Cousens, R.D.; Hahn, M.A.; Heredia, S.M.; Hübner, S.; Turner, K.G.; Whitney, K.D.; Rieseberg, L.H. What we still don't know about invasion genetics. Mol. Ecol. 2015, 24, 2277–2297. [Google Scholar] [CrossRef]
- Anton, A.; Geraldi, N.R.; Lovelock, C.E.; Apostolaki, E.T.; Bennett, S.; Cebrian, J.; Krause-Jensen, D.; Marbà, N.; Martinetto, P.; Pandolfi, J.M.; et al. Global ecological impacts of marine exotic species. Nat. Ecol. Evol. 2019, 3, 787–800. [Google Scholar] [CrossRef] [Green Version]
- Labrune, C.; Amilhat, E.; Amouroux, J.-M.; Jabouin, C.; Gigou, A.; Noël, P. The arrival of the American blue crab, Callinectes sapidus Rathbun, 1896 (Decapoda: Brachyura: Portunidae), in the Gulf of Lions (Mediterranean Sea). BioInvasions Rec. 2019, 8, 876–881. [Google Scholar] [CrossRef]
- Boudouresque, C.-F.; Perret-Boudouresque, M.; Ruitton, S.; Thibault, D. The Invasive Thermophilic Red Alga Lophocladia lallemandii Reaches the Port-Cros National Park (Northwestern Mediterranean). Sci. Rep. Port-Cros Natl. Park 2022, 36, 59–66. [Google Scholar]
- Ruitton, S.; Blanfuné, A.; Boudouresque, C.-F.; Guillemain, D.; Michotey, V.; Roblet, S.; Thibault, D.; Thibaut, T.; Verlaque, M. Rapid Spread of the Invasive Brown Alga Rugulopteryx okamurae in a National Park in Provence (France, Mediterranean Sea). Water 2021, 13, 2306. [Google Scholar] [CrossRef]
- Sardain, A.; Sardain, E.; Leung, B. Global forecasts of shipping traffic and biological invasions to 2050. Nat. Sustain. 2019, 2, 274–282. [Google Scholar] [CrossRef]
- IMO International Convention for the Control and Management of Ships’ Ballast Water and Sediments (BWM); Adoption: 13 February 2004; Entry into Force: 8 September 2017; IMO: London, UK, 2017.
- Antajan, E. The invasive ctenophore Mnemiopsis leidyi A. Agassiz, 1865 along the English Channel and the North Sea French coasts: Another introduction pathway in northern European waters? Aquat. Invasions 2014, 9, 167–173. [Google Scholar] [CrossRef]
- Grizel, H.; Héral, M. Introduction into France of the Japanese oyster (Crassostrea gigas). ICES J. Mar. Sci. 1991, 47, 399–403. [Google Scholar] [CrossRef]
- Goulletquer, P.; Héral, M. Marine Molluscan Production Trends in France: From Fisheries to Aquaculture. NOAA Tech. Rep. NMFS 1997, 129, 137–164. [Google Scholar]
- Gruet, Y.; Héral, M.; Robert, J.-M. Premières Observations Sur l’introduction de La Faune Associée Au Naissain d’huîtres Japonaises Crassostrea gigas (Thunberg), Importé Sur La Côte Atlantique Française. Cah. Biol. Mar. 1976, 17, 173–184. [Google Scholar] [CrossRef]
- Essl, F.; Bacher, S.; Blackburn, T.M.; Booy, O.; Brundu, G.; Brunel, S.; Cardoso, A.-C.; Eschen, R.; Gallardo, B.; Galil, B.; et al. Crossing Frontiers in Tackling Pathways of Biological Invasions. Bioscience 2015, 65, 769–782. [Google Scholar] [CrossRef] [Green Version]
- Carlton, J.T.; Ruiz, G.M. 1. Anthropogenic Vectors of Marine and Estuarine Invasions: An Overview Framework. In Biological Invasions in Changing Ecosystems: Vectors, Ecological Impacts, Management and Predictions; Canning-Clode, J., Ed.; De Gruyter Open: Warsaw, Poland, 2015; pp. 24–36. [Google Scholar]
- Lockwood, J.L.; Cassey, P.; Blackburn, T.M. The more you introduce the more you get: The role of colonization pressure and propagule pressure in invasion ecology. Divers. Distrib. 2009, 15, 904–910. [Google Scholar] [CrossRef]
- Cassey, P.; Delean, S.; Lockwood, J.; Sadowski, J.S.; Blackburn, T.M. Dissecting the null model for biological invasions: A meta-analysis of the propagule pressure effect. PLoS Biol. 2018, 16, e2005987. [Google Scholar] [CrossRef]
- Williams, S.L.; Davidson, I.; Pasari, J.R.; Ashton, G.V.; Carlton, J.T.; Crafton, R.E.; Fontana, R.E.; Grosholz, E.D.; Miller, A.W.; Ruiz, G.M.; et al. Managing Multiple Vectors for Marine Invasions in an Increasingly Connected World. Bioscience 2013, 63, 952–966. [Google Scholar] [CrossRef] [Green Version]
- Rius, M.; Turon, X.; Bernardi, G.; Volckaert, F.A.M.; Viard, F. Marine invasion genetics: From spatio-temporal patterns to evolutionary outcomes. Biol. Invasions 2014, 17, 869–885. [Google Scholar] [CrossRef] [Green Version]
- Viard, F.; David, P.; Darling, J.A. Marine invasions enter the genomic era: Three lessons from the past, and the way forward. Curr. Zool. 2016, 62, 629–642. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Campbell, M.L.; King, S.; Heppenstall, L.D.; van Gool, E.; Martin, R.; Hewitt, C.L. Aquaculture and urban marine structures facilitate native and non-indigenous species transfer through generation and accumulation of marine debris. Mar. Pollut. Bull. 2017, 123, 304–312. [Google Scholar] [CrossRef] [PubMed]
- Carlton, J.; Fowler, A. Ocean rafting and marine debris: A broader vector menu requires a greater appetite for invasion biology research support. Aquat. Invasions 2018, 13, 11–15. [Google Scholar] [CrossRef]
- Fowler, A.E.; Blakeslee, A.M.H.; Canning-Clode, J.; Repetto, M.F.; Phillip, A.M.; Carlton, J.T.; Moser, F.C.; Ruiz, G.M.; Miller, A.W. Opening Pandora’s bait box: A potent vector for biological invasions of live marine species. Divers. Distrib. 2015, 22, 30–42. [Google Scholar] [CrossRef]
- Laing, I.; Gollasch, S. Coscinodiscus Wailesii—A Nuisance Diatom in European Waters. In Invasive Aquatic Species of Europe. Distribution, Impacts and Management; Leppäkoski, E., Gollasch, S., Olenin, S., Eds.; Springer: Dordrecht, The Netherlands, 2002; pp. 53–55. ISBN 978-94-015-9956-6. [Google Scholar]
- Fernandes, L.F.; Zehnder-Alves, L.; Bassfeld, J.C. The recently established diatom Coscinodiscus wailesii (Coscinodiscales, Bacillariophyta) in Brazilian waters. I: Remarks on morphology and distribution. Phycol. Res. 2001, 49, 89–96. [Google Scholar] [CrossRef]
- Bouchet, V.M.P.; Debenay, J.-P.; Sauriau, P.-G. First report of Quinqueloculina carinatastriata (wiesner, 1923) (foraminifera) along the french atlantic coast (marennes-oleron bay and ile de re). J. Foraminifer. Res. 2007, 37, 204–212. [Google Scholar] [CrossRef]
- Richirt, J.; Schweizer, M.; Bouchet, V.M.P.; Mouret, A.; Quinchard, S.; Jorissen, F.J. Morphological Distinction of Three Ammonia Phylotypes Occurring Along European Coasts. J. Foraminifer. Res. 2019, 49, 76–93. [Google Scholar] [CrossRef]
- Richirt, J.; Schweizer, M.; Mouret, A.; Quinchard, S.; Saad, S.A.; Bouchet, V.M.P.; Wade, C.M.; Jorissen, F.J. Biogeographic distribution of three phylotypes (T1, T2 and T6) of Ammonia (foraminifera, Rhizaria) around Great Britain: New insights from combined molecular and morphological recognition. J. Micropalaeontol. 2021, 40, 61–74. [Google Scholar] [CrossRef]
- Moodley, L.; Boschker, H.T.S.; Middelburg, J.; Pel, R.; Herman, P.; De Deckere, E.; Heip, C.H.R. Ecological significance of benthic foraminifera: 13C labelling experiments. Mar. Ecol. Prog. Ser. 2000, 202, 289–295. [Google Scholar] [CrossRef]
- Bouchet, V.; Seuront, L. Strength May Lie in Numbers: Intertidal Foraminifera Non-Negligible Contribution to Surface Sediment Reworking. Open J. Mar. Sci. 2020, 10, 131–140. [Google Scholar] [CrossRef]
- Deldicq, N.; Seuront, L.; Bouchet, V.M.P. Inter-specific and inter-individual trait variability matter in surface sediment reworking rates of intertidal benthic foraminifera. Mar. Biol. 2021, 168, 101. [Google Scholar] [CrossRef]
- Choquel, C.; Geslin, E.; Metzger, E.; Filipsson, H.L.; Risgaard-Petersen, N.; Launeau, P.; Giraud, M.; Jauffrais, T.; Jesus, B.; Mouret, A. Denitrification by benthic foraminifera and their contribution to N-loss from a fjord environment. Biogeosciences 2021, 18, 327–341. [Google Scholar] [CrossRef]
- Chomérat, N.; Antajan, E.; Auby, I.; Bilien, G.; Carpentier, L.; de Casamajor, M.-N.; Ganthy, F.; Hervé, F.; Labadie, M.; Méteigner, C.; et al. First Characterization of Ostreopsis cf. ovata (Dinophyceae) and Detection of Ovatoxins during a Multispecific and Toxic Ostreopsis Bloom on French Atlantic Coast. Mar. Drugs 2022, 20, 461. [Google Scholar] [CrossRef]
- Gully, F.; Cochu, M.; Delemarre, J.-L. Première Observation Du Gastéropode Nassarius corniculum (Olivi, 1792) En Bretagne. An Aod Cah. Nat. Obs. Mar. 2013, II, 29–33. [Google Scholar]
- Copp, G.; Vilizzi, L.; Tidbury, H.; Stebbing, P.; Tarkan, A.S.; Miossec, L.; Goulletquer, P. Development of a generic decision-support tool for identifying potentially invasive aquatic taxa: AS-ISK. Manag. Biol. Invasions 2016, 7, 343–350. [Google Scholar] [CrossRef] [Green Version]
- Essl, F.; Dullinger, S.; Genovesi, P.; Hulme, P.E.; Jeschke, J.M.; Katsanevakis, S.; Kühn, I.; Lenzner, B.; Pauchard, A.; Pyšek, P.; et al. A Conceptual Framework for Range-Expanding Species that Track Human-Induced Environmental Change. Bioscience 2019, 69, 908–919. [Google Scholar] [CrossRef]
- Sauriau, P.G. Spread of Cyclope neritea (Mollusca: Gastropoda) along the north-eastern Atlantic coasts in relation to oyster culture and to climatic fluctuations. Mar. Biol. 1991, 109, 299–309. [Google Scholar] [CrossRef]
- Simon-Bouhet, B.; Garcia, P.; Viard, F. Multiple introductions promote range expansion of the mollusc Cyclope neritea (Nassariidae) in France: Evidence from mitochondrial sequence data. Mol. Ecol. 2006, 15, 1699–1711. [Google Scholar] [CrossRef] [Green Version]
- Bachelet, G.; Simon-Bouhet, B.; Desclaux, C.; Garcia-Meunier, P.; Mairesse, G.; De Montaudouin, X.; Raigné, H.; Randriambao, K.; Sauriau, P.-G.; Viard, F. Invasion of the eastern Bay of Biscay by the nassariid gastropod Cyclope neritea: Origin and effects on resident fauna. Mar. Ecol. Prog. Ser. 2004, 276, 147–159. [Google Scholar] [CrossRef] [Green Version]
- Geller, J.B.; Darling, J.A.; Carlton, J.T. Genetic Perspectives on Marine Biological Invasions. Annu. Rev. Mar. Sci. 2010, 2, 367–393. [Google Scholar] [CrossRef] [PubMed]
- Viard, F.; Comtet, T. 18. Applications of DNA-based Methods for the Study of Biological Invasions. In Biological Invasions in Changing Ecosystems: Vectors, Ecological Impacts, Management and Predictions; Canning-Clode, J., Ed.; De Gruyter Open: Warsaw, Poland, 2015; pp. 411–435. [Google Scholar]
- Sato-Okoshi, W.; Okoshi, K.; Abe, H.; Dauvin, J.-C. Polydorid species (Annelida: Spionidae) associated with commercially important oyster shells and their shell infestation along the coast of Normandy, in the English Channel, France. Aquac. Int. 2022. [Google Scholar] [CrossRef]
- Tiralongo, F.; Crocetta, F.; Riginella, E.; Lillo, A.O.; Tondo, E.; Macali, A.; Mancini, E.; Russo, F.; Coco, S.; Paolillo, G.; et al. Snapshot of rare, exotic and overlooked fish species in the Italian seas: A citizen science survey. J. Sea Res. 2020, 164, 101930. [Google Scholar] [CrossRef]
- Zenetos, A.; Koutsogiannopoulos, D.; Ovalis, P.; Poursanidis, D. The Role Played by Citizen Scientists in Monitoring Marine Alien Species in Greece. Cah. Biol. Mar. 2013, 54, 419–426. [Google Scholar] [CrossRef]
- Mannino, A.M.; Balistreri, P. Citizen science: A successful tool for monitoring invasive alien species (IAS) in Marine Protected Areas. The case study of the Egadi Islands MPA (Tyrrhenian Sea, Italy). Biodiversity 2018, 19, 42–48. [Google Scholar] [CrossRef]
- Brodie, J.; Kunzig, S.; Agate, J.; Yesson, C.; Robinson, L. The Big Seaweed Search: Evaluating a citizen science project for a difficult to identify group of organisms. Aquat. Conserv. Mar. Freshw. Ecosyst. 2022, 33, 44–55. [Google Scholar] [CrossRef]
- Encarnação, J.; Teodósio, M.A.; Morais, P. Citizen Science and Biological Invasions: A Review. Front. Environ. Sci. 2021, 8, 602980. [Google Scholar] [CrossRef]
- Bishop, J.D.; Wood, C.A.; Lévêque, L.; Yunnie, A.L.; Viard, F. Repeated rapid assessment surveys reveal contrasting trends in occupancy of marinas by non-indigenous species on opposite sides of the western English Channel. Mar. Pollut. Bull. 2015, 95, 699–706. [Google Scholar] [CrossRef]
- Lambert, C.; Lambert, G. Persistence and differential distribution of nonindigenous ascidians in harbors of the Southern California Bight. Mar. Ecol. Prog. Ser. 2003, 259, 145–161. [Google Scholar] [CrossRef] [Green Version]
- Viard, F.; Roby, C.; Turon, X.; Bouchemousse, S.; Bishop, J. Cryptic Diversity and Database Errors Challenge Non-indigenous Species Surveys: An Illustration with Botrylloides spp. in the English Channel and Mediterranean Sea. Front. Mar. Sci. 2019, 6, 615. [Google Scholar] [CrossRef] [Green Version]
- Turon, X.; Casso, M.; Pascual, M.; Viard, F. Looks can be deceiving: Didemnum pseudovexillum sp. nov. (Ascidiacea) in European harbours. Mar. Biodivers. 2020, 50, 1–14. [Google Scholar] [CrossRef]
- Casso, M.; Turon, X.; Pascual, M. Single zooids, multiple loci: Independent colonisations revealed by population genomics of a global invader. Biol. Invasions 2019, 21, 3575–3592. [Google Scholar] [CrossRef]
- Lavesque, N.; Daffe, G.; Bonifácio, P.; Hutchings, P. A new species of the Marphysa sanguinea complex from French waters (Bay of Biscay, NE Atlantic) (Annelida, Eunicidae). Zookeys 2017, 716, 1–17. [Google Scholar] [CrossRef]
- Lavesque, N.; Hutchings, P.; Abe, H.; Daffe, G.; Gunton, L.; Glasby, C. Confirmation of the exotic status of Marphysa victori Lavesque, Daffe, Bonifácio & Hutchings, 2017 (Annelida) in French waters and synonymy of Marphysa bulla Liu, Hutchings & Kupriyanova, 2018. Aquat. Invasions 2020, 15, 355–366. [Google Scholar] [CrossRef]
- Pante, E.; Schoelinck, C.; Puillandre, N. From Integrative Taxonomy to Species Description: One Step Beyond. Syst. Biol. 2014, 64, 152–160. [Google Scholar] [CrossRef] [Green Version]
- Genovesi, P. Eradications of invasive alien species in Europe: A review. Biol. Invasions 2005, 7, 127–133. [Google Scholar] [CrossRef] [Green Version]
- Ojaveer, H.; Galil, B.S.; Campbell, M.L.; Carlton, J.T.; Canning-Clode, J.; Cook, E.J.; Davidson, A.D.; Hewitt, C.L.; Jelmert, A.; Marchini, A.; et al. Classification of non-indigenous species based on their impacts: Considerations for application in marine management. PLoS Biol. 2015, 13, e1002130. [Google Scholar] [CrossRef] [Green Version]
- Booy, O.; Mill, A.C.; Roy, H.E.; Hiley, A.; Moore, N.; Robertson, P.; Baker, S.; Brazier, M.; Bue, M.; Bullock, R.; et al. Risk management to prioritise the eradication of new and emerging invasive non-native species. Biol. Invasions 2017, 19, 2401–2417. [Google Scholar] [CrossRef] [Green Version]
- Reaser, J.K.; Burgiel, S.W.; Kirkey, J.; Brantley, K.A.; Veatch, S.D.; Burgos-Rodríguez, J. The early detection of and rapid response (EDRR) to invasive species: A conceptual framework and federal capacities assessment. Biol. Invasions 2019, 22, 1–19. [Google Scholar] [CrossRef] [Green Version]
- Ojaveer, H.; Galil, B.S.; Minchin, D.; Olenin, S.; Amorim, A.; Canning-Clode, J.; Chainho, P.; Copp, G.H.; Gollasch, S.; Jelmert, A.; et al. Ten recommendations for advancing the assessment and management of non-indigenous species in marine ecosystems. Mar. Policy 2014, 44, 160–165. [Google Scholar] [CrossRef]
- Wright, D.A. Compliance assessment for the ballast water convention: Time for a re-think? A U.K. case study. J. Mar. Eng. Technol. 2018, 20, 254–261. [Google Scholar] [CrossRef]
- Lister, J.; Poulsen, R.T.; Ponte, S. Orchestrating transnational environmental governance in maritime shipping. Glob. Environ. Chang. 2015, 34, 185–195. [Google Scholar] [CrossRef] [Green Version]
- European Commission. Regulation (EU) No 1143/2014 of the European Parliament and of the Council of 22 October 2014 on the Prevention and Management of the Introduction and Spread of Invasive Alien Species; European Commission: Brussels, Belgium, 2014. [Google Scholar]
- Acosta, H.; Forrest, B.M. The spread of marine non-indigenous species via recreational boating: A conceptual model for risk assessment based on fault tree analysis. Ecol. Model. 2009, 220, 1586–1598. [Google Scholar] [CrossRef]
- Raybaud, V.; Beaugrand, G.; Dewarumez, J.-M.; Luczak, C. Climate-induced range shifts of the American jackknife clam Ensis directus in Europe. Biol. Invasions 2014, 17, 725–741. [Google Scholar] [CrossRef] [Green Version]
- Schickele, A.; Guidetti, P.; Giakoumi, S.; Zenetos, A.; Francour, P.; Raybaud, V. Improving predictions of invasive fish ranges combining functional and ecological traits with environmental suitability under climate change scenarios. Glob. Chang. Biol. 2021, 27, 6086–6102. [Google Scholar] [CrossRef]
- Gimenez, L.; Rivera, R.; Brante, A. One step ahead of sea anemone invasions with ecological niche modeling: Potential distributions and niche dynamics of three successful invasive species. Mar. Ecol. Prog. Ser. 2022, 690, 83–95. [Google Scholar] [CrossRef]
- Tsiamis, K.; Azzurro, E.; Bariche, M.; Çinar, M.E.; Crocetta, F.; De Clerck, O.; Galil, B.; Gómez, F.; Hoffman, R.; Jensen, K.R.; et al. Prioritizing marine invasive alien species in the European Union through horizon scanning. Aquat. Conserv. Mar. Freshw. Ecosyst. 2020, 30, 794–845. [Google Scholar] [CrossRef]
- Copp, G.H.; Vilizzi, L.; Wei, H.; Li, S.; Piria, M.; Al-Faisal, A.J.; Almeida, D.; Atique, U.; Al-Wazzan, Z.; Bakiu, R.; et al. Speaking their language—Development of a multilingual decision-support tool for communicating invasive species risks to decision makers and stakeholders. Environ. Model. Softw. 2020, 135, 104900. [Google Scholar] [CrossRef]
- Vilizzi, L.; Copp, G.H.; Hill, J.E.; Adamovich, B.; Aislabie, L.; Akin, D.; Al-Faisal, A.J.; Almeida, D.; Azmai, M.A.; Bakiu, R.; et al. A global-scale screening of non-native aquatic organisms to identify potentially invasive species under current and future climate conditions. Sci. Total Environ. 2021, 788, 147868. [Google Scholar] [CrossRef]
- Tidbury, H.J.; Taylor, N.G.H.; Copp, G.H.; Garnacho, E.; Stebbing, P.D. Predicting and mapping the risk of introduction of marine non-indigenous species into Great Britain and Ireland. Biol. Invasions 2016, 18, 3277–3292. [Google Scholar] [CrossRef]
- Geraldi, N.R.; Anton, A.; Santana-Garcon, J.; Bennett, S.; Marbà, N.; Lovelock, C.E.; Apostolaki, E.T.; Cebrian, J.; Krause-Jensen, D.; Martinetto, P.; et al. Ecological effects of non-native species in marine ecosystems relate to co-occurring anthropogenic pressures. Glob. Chang. Biol. 2019, 26, 1248–1258. [Google Scholar] [CrossRef]
- Clavel, J.; Julliard, R.; Devictor, V. Worldwide decline of specialist species: Toward a global functional homogenization? Front. Ecol. Environ. 2010, 9, 222–228. [Google Scholar] [CrossRef] [Green Version]
- Wallingford, P.D.; Morelli, T.L.; Allen, J.; Beaury, E.M.; Blumenthal, D.M.; Bradley, B.A.; Dukes, J.S.; Early, R.; Fusco, E.J.; Goldberg, D.E.; et al. Adjusting the lens of invasion biology to focus on the impacts of climate-driven range shifts. Nat. Clim. Chang. 2020, 10, 398–405. [Google Scholar] [CrossRef]
- Sakai, A.K.; Allendorf, F.W.; Holt, J.S.; Lodge, D.M.; Molofsky, J.; With, K.A.; Baughman, S.; Cabin, R.J.; Cohen, J.E.; Ellstrand, N.C.; et al. The Population Biology of Invasive Species. Annu. Rev. Ecol. Syst. 2001, 32, 305–332. [Google Scholar] [CrossRef] [Green Version]
- Catford, J.A.; Jansson, R.; Nilsson, C. Reducing redundancy in invasion ecology by integrating hypotheses into a single theoretical framework. Divers. Distrib. 2008, 15, 22–40. [Google Scholar] [CrossRef] [Green Version]
- Hawkins, C.L.; Bacher, S.; Essl, F.; Hulme, P.E.; Jeschke, J.M.; Kühn, I.; Kumschick, S.; Nentwig, W.; Pergl, J.; Pyšek, P.; et al. Framework and guidelines for implementing the proposed IUCN Environmental Impact Classification for Alien Taxa (EICAT). Divers. Distrib. 2015, 21, 1360–1363. [Google Scholar] [CrossRef] [Green Version]
- Giakoumi, S.; Katsanevakis, S.; Albano, P.G.; Azzurro, E.; Cardoso, A.C.; Cebrian, E.; Deidun, A.; Edelist, D.; Francour, P.; Jimenez, C.; et al. Management priorities for marine invasive species. Sci. Total Environ. 2019, 688, 976–982. [Google Scholar] [CrossRef]
- Comtet, T.; Sandionigi, A.; Viard, F.; Casiraghi, M. DNA (meta)barcoding of biological invasions: A powerful tool to elucidate invasion processes and help managing aliens. Biol. Invasions 2015, 17, 905–922. [Google Scholar] [CrossRef]
- Darling, J.A.; Galil, B.S.; Carvalho, G.R.; Rius, M.; Viard, F.; Piraino, S. Recommendations for developing and applying genetic tools to assess and manage biological invasions in marine ecosystems. Mar. Policy 2017, 85, 54–64. [Google Scholar] [CrossRef]
- Von Ammon, U.; Wood, S.A.; Laroche, O.; Zaiko, A.; Tait, L.; Lavery, S.; Inglis, G.J.; Pochon, X. Combining morpho-taxonomy and metabarcoding enhances the detection of non-indigenous marine pests in biofouling communities. Sci. Rep. 2018, 8, 16290. [Google Scholar] [CrossRef] [Green Version]
- Azevedo, J.; Antunes, J.T.; Machado, A.M.; Vasconcelos, V.; Leão, P.N.; Froufe, E. Monitoring of biofouling communities in a Portuguese port using a combined morphological and metabarcoding approach. Sci. Rep. 2020, 10, 13461. [Google Scholar] [CrossRef] [PubMed]
- Westfall, K.M.; Therriault, T.W.; Abbott, C.L. A new approach to molecular biosurveillance of invasive species using DNA metabarcoding. Glob. Chang. Biol. 2019, 26, 1012–1022. [Google Scholar] [CrossRef] [PubMed]
- Couton, M.; Lévêque, L.; Daguin-Thiébaut, C.; Comtet, T.; Viard, F. Water eDNA metabarcoding is effective in detecting non-native species in marinas, but detection errors still hinder its use for passive monitoring. Biofouling 2022, 38, 367–383. [Google Scholar] [CrossRef] [PubMed]
- Rey, A.; Basurko, O.C.; Rodriguez-Ezpeleta, N. Considerations for metabarcoding-based port biological baseline surveys aimed at marine nonindigenous species monitoring and risk assessments. Ecol. Evol. 2020, 10, 2452–2465. [Google Scholar] [CrossRef] [Green Version]
- Zaiko, A.; Greenfield, P.; Abbott, C.; von Ammon, U.; Bilewitch, J.; Bunce, M.; Cristescu, M.E.; Chariton, A.; Dowle, E.; Geller, J.; et al. Towards reproducible metabarcoding data: Lessons from an international cross-laboratory experiment. Mol. Ecol. Resour. 2021, 22, 519–538. [Google Scholar] [CrossRef]
- Créach, V.; Derveaux, S.; Owen, K.R.; Pitois, S.; Antajan, E. Use of environmental DNA in early detection of Mnemiopsis leidyi in UK coastal waters. Biol. Invasions 2021, 24, 415–424. [Google Scholar] [CrossRef]
- Rondeau, S.; Davoult, D.; Lejeusne, C.; Kenworthy, J.; Bohner, O.; Loisel, S.; Gauff, R. Persistent dominance of non-indigenous species in the inner part of a marina highlighted by multi-year photographic monitoring. Mar. Ecol. Prog. Ser. 2022, 690, 15–30. [Google Scholar] [CrossRef]
- Spilmont, N.; Gothland, M.; Seuront, L. Exogenous control of the feeding activity in the invasive Asian shore crab Hemigrapsus sanguineus (De Haan, 1835). Aquat. Invasions 2015, 10, 327–332. [Google Scholar] [CrossRef] [Green Version]
- Chebaane, S.; Canning-Clode, J.; Ramalhosa, P.; Belz, J.; Castro, N.; Órfão, I.; Sempere-Valverde, J.; Engelen, A.H.; Pais, M.P.; Monteiro, J.G. From Plates to Baits: Using a Remote Video Foraging System to Study the Impact of Foraging on Fouling Non-Indigenous Species. J. Mar. Sci. Eng. 2022, 10, 611. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Type of Release | Pathway Categories | Vectors |
|---|---|---|
| Intentional | Release in nature | Fishery in the wild (including game fishing) Other intentional release |
| Escape from confinement | Aquaculture/mariculture Botanical garden/zoo/aquaria (excluding domestic aquaria) Live food and live bait | |
| Unintentional | Transport-Hitchhikers | Hitchhikers on animals (except parasites, species transported by host/vector) Parasites on animals (including species transported by host and vector) |
| Transport-Stowaway | Angling/fishing equipment Hitchhikers on ship/boat (excluding ballast water and hull fouling) Organic packing material, in particular, wood packaging Ship/boat ballast water Ship/boat hull fouling | |
| Corridor | Interconnected waterways/basins/seas | |
| Unaided | Natural secondary dispersal across borders of invasive alien species that have been introduced |
| Kingdom | Phylum | Overall French Coastline | Northern Seas | Bay of Biscay | Western Mediterranean |
|---|---|---|---|---|---|
| Plantae | Rhodophyta | 67 (20%) | 26 (15%) | 31 (17%) | 59 (24.7%) |
| Chlorophyta | 10 (3%) | 3 (2%) | 2 (1%) | 10 (4%) | |
| Tracheophyta | 5 (1%) | 3 (2%) | 4 (2%) | 2 (0.8%) | |
| Chromista | Ochrophyta | 20 (6%) | 6 (4%) | 6 (3%) | 18 (7.5%) |
| Animalia | Arthropoda | 69 (20%) | 44 (26%) | 47 (25%) | 38 (16%) |
| Mollusca | 44 (13%) | 23 (14%) | 29 (16%) | 25 (10%) | |
| Chordata | 43 (13%) | 18 (11%) | 21 (11%) | 36 (15%) | |
| Annelida | 38 (11%) | 22 (13%) | 22 (12%) | 18 (8%) | |
| Cnidaria | 23 (7%) | 9 (5%) | 8 (4%) | 17 (7%) | |
| Bryozoa | 16 (5%) | 9 (5%) | 10 (5%) | 11 (5%) | |
| Platyhelminthes | 2 (0.5%) | 2 (0.5%) | 2 (1%) | 1 (0.4%) | |
| Porifera | 2 (0.5%) | 1 (0.5%) | 1 (1%) | 1 (0.4%) | |
| Ctenophora | 1 (0%) | 1 (1%) | 1 (1%) | 1 (0.4%) | |
| Nematoda | 1 (0%) | 1 (1%) | 1 (1%) | 1 (0.4%) | |
| Echinodermata | 1 (0%) | 0 | 0 | 1 (0.4%) |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Massé, C.; Viard, F.; Humbert, S.; Antajan, E.; Auby, I.; Bachelet, G.; Bernard, G.; Bouchet, V.M.P.; Burel, T.; Dauvin, J.-C.; et al. An Overview of Marine Non-Indigenous Species Found in Three Contrasting Biogeographic Metropolitan French Regions: Insights on Distribution, Origins and Pathways of Introduction. Diversity 2023, 15, 161. https://doi.org/10.3390/d15020161
Massé C, Viard F, Humbert S, Antajan E, Auby I, Bachelet G, Bernard G, Bouchet VMP, Burel T, Dauvin J-C, et al. An Overview of Marine Non-Indigenous Species Found in Three Contrasting Biogeographic Metropolitan French Regions: Insights on Distribution, Origins and Pathways of Introduction. Diversity. 2023; 15(2):161. https://doi.org/10.3390/d15020161
Chicago/Turabian StyleMassé, Cécile, Frédérique Viard, Suzie Humbert, Elvire Antajan, Isabelle Auby, Guy Bachelet, Guillaume Bernard, Vincent M. P. Bouchet, Thomas Burel, Jean-Claude Dauvin, and et al. 2023. "An Overview of Marine Non-Indigenous Species Found in Three Contrasting Biogeographic Metropolitan French Regions: Insights on Distribution, Origins and Pathways of Introduction" Diversity 15, no. 2: 161. https://doi.org/10.3390/d15020161