
Shearwater Eggs in Lobos 3, a Holocene Site of Fuerteventura (Canary Islands)

 , , ,
, , , {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
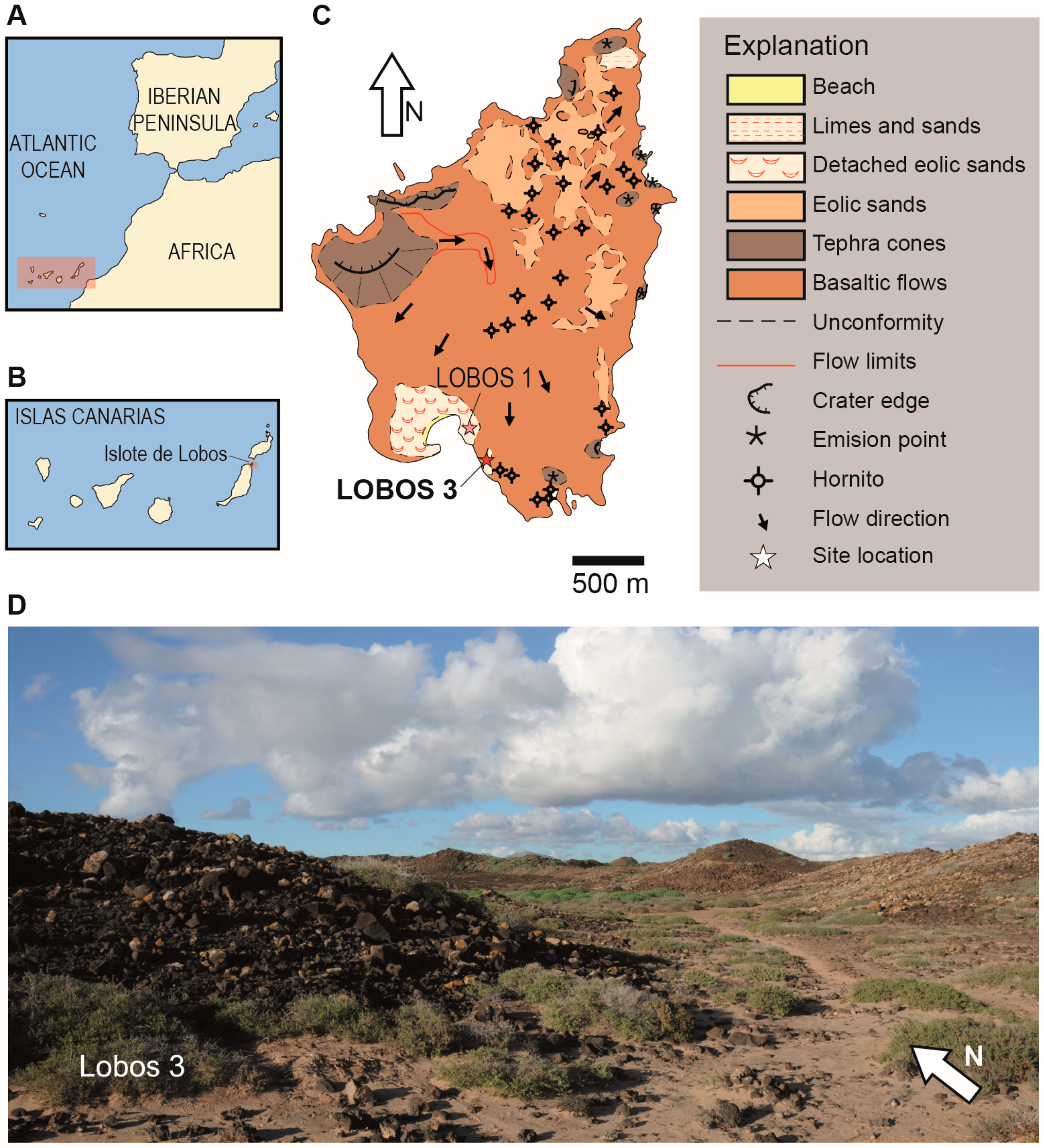
The Site of Lobos 3
2. Materials and Methods
2.1. Laboratory Work
2.2. Identification of the Eggs
3. Results
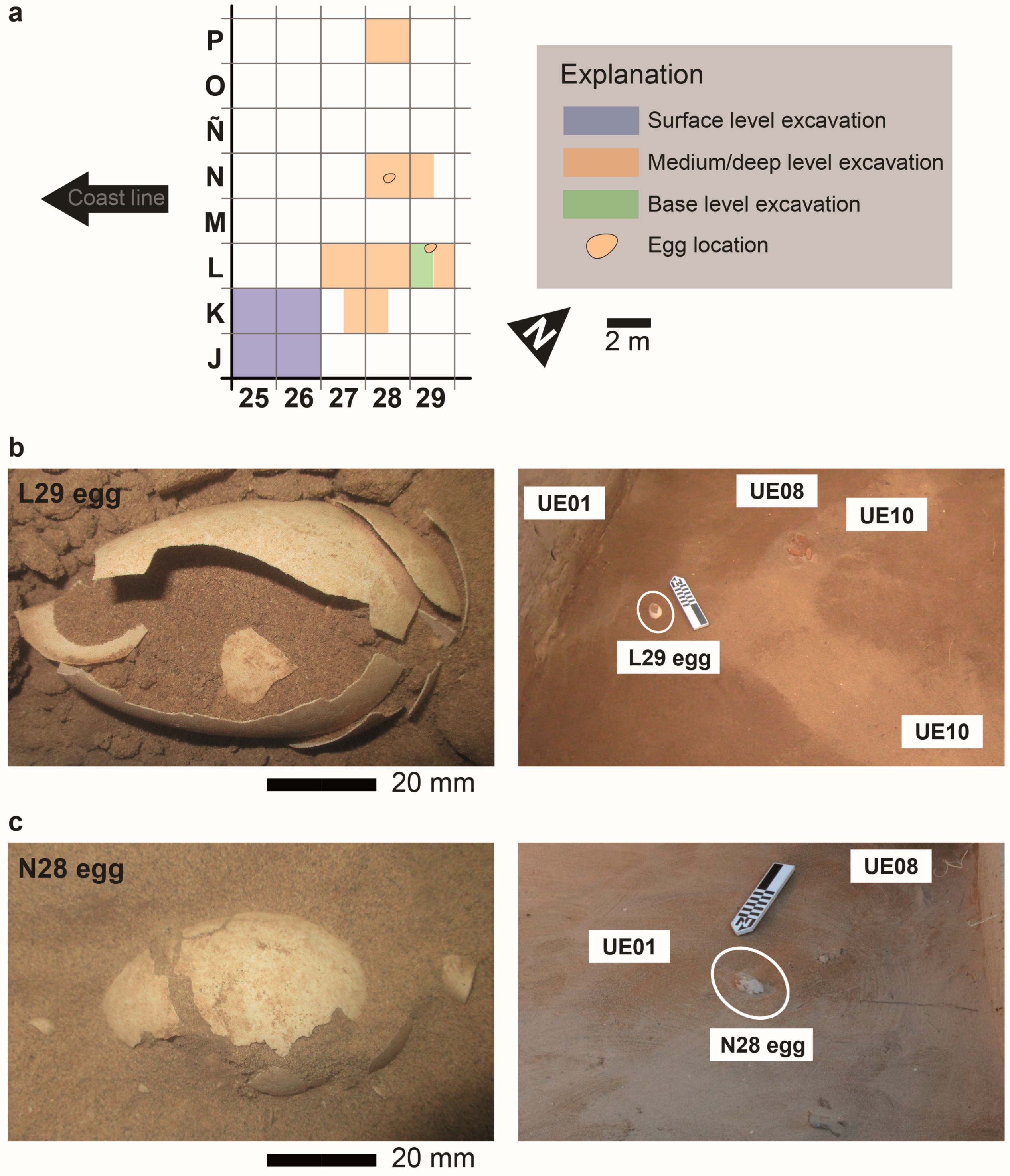
3.1. L29 Egg
3.2. N28 Egg
4. Discussion
4.1. Systematic Analysis
4.2. Origin of the Accumulation and Seasonality Pattern
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Del-Arco-Aguilar, M.C. De nuevo sobre el descubrimiento y colonización antiguos de Canarias. Reflexiones sobre aspectos teóricos y datos empíricos. Anu. Estud. Atlánticos 2021, 67, 1–27. [Google Scholar]
- Atoche-Peña, P.; Del-Arco-Aguilar, M.C. Las Islas Canarias en el contexto del Mediterráneo antiguo. In Homenaje a Paloma Cabrera Bonet; Carretero, A.., Ed.; Ministerio de Cultura y Deporte: Madrid, Spain, 2021; pp. 39–49. [Google Scholar]
- Carracedo, J.C.; Pérez Torrado, F.J.; Ancochea, E.; Meco, J.; Francisco, H.; Cuvas, C.R.; Casillas, E.; Rodríguez Badiola, E.; Ahijado, D. Cenozoic volcanism II: The Canary Islands. In The Geology of Spain; Gibbons, W., Moreno, T., Eds.; The Gological Society of London: London, UK, 2002. [Google Scholar]
- Balcells Herrera, R.; Barrera-Morate, J.L. Mapa geológico de la Hoja no. 1086I (Tostón-Corralejo). In Mapa Geológico de España E. 1:25.000. Segunda Serie (MAGNA), 1st ed.; IGME. M-4021-2004; Instituto Geológico y Minero de España: Madrid, Spain, 2004; ISBN 84-7840-501-1. [Google Scholar]
- Del-Arco-Aguilar, M.C.; Del-Arco-Aguilar, M.; Cebrián-Guimerá, R.; Garrido-Chacón, H.M.; Rodríguez-Fidel, D.A.; Siverio-Batista, C. Lobos 1: Una factoría de púrpura romana en el Atlántico centro-oriental (Fuerteventura, Islas Canarias). In Redefining Ancient Textile Handcraft: Structures, Tools and Production Processes PURPURAE VESTES VII; Bustamante-Álvarez, M., Sánchez López, E.H., Jiménez Ávila, J., Eds.; Universidad de Granada: Granada, Spain, 2020; pp. 95–117. [Google Scholar]
- Del-Arco, M.C.; Del-Arco, M. The romans in Canarias. A vision from the purple workshop of El Islote de Lobos (Fuerteventura). In Actualidad de la Investigación Arqueológica en España (2018-2019): Conferencias Impartidas en el Museo Arqueológico Nacional; Carretero-Pérez, A., Papí Rodes, C., Eds.; Museo Arqueológico Nacional, Ministerio de Cultura y Deporte: Madrid, España, 2020; pp. 451–567. [Google Scholar]
- Cebrián-Guimerá, R.; Del-Arco-Aguilar, M.C.; Del-Arco-Aguilar, M.; Bernal-Casasola, D.; Cantillo-Duarte, J.J.; Vargás-Girón, J.M. Púrpura romana en el Olivillo (Cádiz) y Lobos (Canarias): Comparativa de los patrones de fracturación y arqueología experimental. In Redefining Ancient Textile Handcraft: Structures, Tools and Production Processes PURPURAE VESTES VII; Bustamante-Álvarez, M., Sánchez López, E.H., Jiménez Ávila, J., Eds.; Universidad de Granada: Granada, Spain, 2020; pp. 109–117. [Google Scholar]
- Del-Arco-Aguilar, M.C.; Del-Arco-Aguilar, M.; Benito-Mateo, C.; Rosario Adrián, M.C. Un taller romano de púrpura en los límites de la Ecúmene: Lobos 1 (Fuerte Ventura, Islas Canarias), Primeros resultados. In Santa Cruz de Tenerife: Canarias Arqueologica, Monografías 6; Museos de Tenerife: Tenerife, Spain, 2016. [Google Scholar]
- Siverio-Batista, C. Estudio de las arqueofaunas terrestres en talleres de púrpura del Atlántico y Mediterráneo centro-occidental durante la Antigüedad. Arqueol. Territ. 2017, 14, 193–204. [Google Scholar]
- Castillo, C.; Martín-González, R.; Coello, J.J. Small vertebrate taphonomy of La Cueva del Llano, a volcanic cave on Fuerteventura (Canary Islands, Spain). Palaeoecological implications. Palaeogeogr. Palaeoclimatol. Palaeoecol. 2001, 166, 277–291. [Google Scholar] [CrossRef]
- Cruzado-Caballero, P.; Castillo-Ruiz, C.; Bolet, A.; Colmenero, J.R.; de la Nuez, J.; Casillas, R.; Llacer, S.; Bernardini, F.; Fortuny, J. First nearly complete skull of Gallotia auaritae (lower-middle Pleistocene, Squamata, Gallotiinae) and a morphological phylogenetic analysis of the genus Gallotia. Sci. Rep. 2018, 9, 16629. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Miller, J.P.; Cruzado-Cabalero, P.; de la Nuez, J.; Carrillo-Pacheco, M.; Castillo-Ruiz, C.P. Review of the Middle Pleistocene molluscan association from La Mancha de la Laja, Tenerife, Spain, with the description of two new species of Napaeus Albers, 1850 (Gastropoda: Enidae). Histor. Biol. 2022. [Google Scholar] [CrossRef]
- González-Dionis, J.; Castillo-Ruiz, C.; Cruzado Caballero, P.; David Melero, E.; Crespo, V.D. First study of the bat fossil record of the mid-Atlantic volcanic islands. Earth Environ. Sci. Trans. R. Soc. Edinburg 2022, 113, 13–27. [Google Scholar] [CrossRef]
- Palacios-García, S.; Cruzado-Caballero, P.; Casillas, R.; Castillo-Ruiz, C.R. Quaternary biodiversity of the giant fossil endemic lizards from the island of El Hierro (Canary Islands, Spain). Quat. Sci. Rev. 2021, 262, 106961. [Google Scholar] [CrossRef]
- Meco, J. Las Pardelas Pleistocenas de Hueso del Caballo, Jandía (Fuerteventura); Excelentísimo Cabildo Insular de Fuerteventura: Fuerteventura, Spain, 1994; ISBN 84-87461-25-5. [Google Scholar]
- Sánchez-Marco, A. New finding of Pleistocene marine birds from Fuerteventura (Canary Islands). Coloq. Paleontol. Vol. Extraordin. 2003, 1, 627–636. [Google Scholar]
- Sánchez-Marco, A. New data and overview of the past avifaunas from the Canary Islands. Ardeola 2010, 57, 13–40. [Google Scholar]
- Rando, J.C.; Alcover, J.A. Evidence for a second western Palearctic seabird extinction during the last Millenium: The Lava Shearwater Puffinus Olsoni. Ibis 2008, 150, 188–192. [Google Scholar] [CrossRef]
- Jaume, D.; McMinn, M.; Alcover, J.A. Fossil birds from the Bujero del Silo, La Gomera (Canary Islands), with a description of a new species of quail (Galliformes: Phasianidae). Bol. Mus. Munic. Funchal 1993, 2, 147–165. [Google Scholar]
- Alcover, J.A.; Florit, F. Una nueva especie de Carduelis (Fringillidae) de La Palma. Vieraea 1987, 17, 75–86. [Google Scholar]
- Rando, J.C.; Alcover, J.A.; Illera, J.C. Disentangling Ancient Interactions: A New Extinct Passerine Provides Insights on Character Displacement among Extinct and Extant Island Finches. PLoS ONE 2010, 5, e12956. [Google Scholar] [CrossRef] [Green Version]
- Rando, J.C.; López, M.; Seguí, B. A new species of extinct flightless passerine (Emberizidae: Emberiza) from the Canary Islands. Condor 1999, 101, 1–13. [Google Scholar] [CrossRef]
- McMinn, M.; Jaume, D.; Alcover, J.A. Puffinus olsoni n. sp.: Nova espècie de baldritja recentment extinguida provinent de depòsits espeleològics de Fuerteventura i Lanzarote (Illes Canàries, Atlàntic Oriental). Endins 1990, 16, 63–71. [Google Scholar]
- Walker, C.A.; Wragg, G.M.; Harrison, C.J.O. A new shearwater from the Pleistocene of the Canary Islands and its bearing on the evolution of certain Puffinus shearwaters. Hist. Biol. 1990, 3, 203–224. [Google Scholar] [CrossRef]
- Alcover, J.A.; McMinn, M. Fossil birds from the Canary Islands. Cour. Forsch. Senckenberg 1995, 181, 213–217. [Google Scholar]
- Sánchez-Marco, A. Aves fósiles de la Península Ibérica, Canarias y Baleares: Balance de los estudios realizados. Revista PH 2018, 94, 154–181. [Google Scholar] [CrossRef]
- Rando, J.C.; Alcover, J.A. On the extinction of the Dune Shearwater (Puffinus holeae) from the Canary Islands. J. Ornithol. 2010, 151, 365–369. [Google Scholar] [CrossRef]
- Illera, J.C.; Richardson, D.S.; Emerson, B.C. Age, origins and extinction of the avifauna of Macaronesia: A synthesis of phylogenetic and fossil information. Quat. Sci. Rev. 2012, 50, 14–22. [Google Scholar] [CrossRef] [Green Version]
- Illera, J.C.; Spurgin, L.G.; Rodríguez Expósito, E.; Nogales, M.; Rando, J.C. What are we learning about speciation and extinction from the Canary Islands? Ardeola 2016, 63, 15–33. [Google Scholar] [CrossRef] [Green Version]
- Sauer, G.F.; Rothe, P. Ratite eggshells from Lanzarote, Canary Islands. Science 1972, 176, 43–45. [Google Scholar] [CrossRef]
- Kai, A.; Miki, T.; Ikeya, M. ESR dating of teeth, bones and eggshells excavated at a Paleolithic site of Douara Cave, Syria. Quat. Sci. Rev. 1988, 7, 503–507. [Google Scholar] [CrossRef]
- Teixer, P.J.; Porraz, G.; Parkington, J.; Rigaud, J.P.; Poggenpoel, C.; Miller, C.; Tribolo, C.; Cartwright, C.; Coudennau, A.; Klein, R.; et al. A howiesons poort tradition of engraving ostrich eggshell containers dated to 60,000 years ago at Diepkloof Rock Shelter, South Africa. Proc. Natl. Acad. Sci. USA 2010, 107, 6180–6185. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fernández, F.J.; Del Papa, L.M.; Mange, E.; Teta, P.; Crivelli Montero, E.; Pardiñas, U.F.J. Human subsistence and environmental stability during the last 2200 years in Epullán Cave (Northwestern Patagonia, Argentina): A Perspective from zooarchaeological record. Quat. Int. 2016, 391, 38–50. [Google Scholar] [CrossRef]
- Clarke, S.J.; Miller, G.H.; Murray-Wallace, C.V.; David, B.; Pasveer, J.M. The geochronological potential of isoleucine epimerization in cassowary and megapode eggshells from archaeological sites. J. Archaeol. Sci. 2007, 34, 1051–1063. [Google Scholar] [CrossRef]
- Keepax, C.A. Avian egg-shell from archaeological sites. J. Archaeol. Sci. 1981, 8, 315–335. [Google Scholar] [CrossRef]
- Sidell, E.J. A methodology for the identification of avian eggshell from archaeological sites. Archaeofauna Archaeoornithol. 1993, 2, 45–51. [Google Scholar]
- Beacham, E.B.; Durand, S.R. Eggshell and the archaeological record: New insights into turkey husbandry in the American Southwest. J. Archeol. Sci. 2007, 34, 1610–1621. [Google Scholar] [CrossRef]
- Núñez-Lahuerta, C.; Moreno-Azanza, M.; Cuenca-Bescós, G. Avian eggshell remains in the human bearing level TD6 of the Gran Dolina site (Early Pleistocene, Atapuerca, Spain). Hist. Biol. 2021, 33, 660–671. [Google Scholar] [CrossRef]
- Quinn, B. Fossilized eggshell preparation. In Vertebrate Paleontological Techniques; Leigii, P., May, P., Eds.; Cambridge University Press: Cambridge, UK, 1994; Volume 1, pp. 146–153. [Google Scholar]
- Billerman, M.; Keeney, B.K.; Rodewald, P.G.; Schulenberg, T.S. Birds of the World; Cornell Laboratory of Ornithology: Ithaca, NY, USA, 2022. [Google Scholar]
- Mikhailov, K.E. Fossil and recent eggshell in amniotic vertebrates: Fine structure, comparative morphology and classification. Spec. Pap. Paleontol. 1997, 56, 1–76. [Google Scholar]
- Schneider, C.A.; Rasband, W.S.; Eliceri, K.W. NIH Image of ImageJ: 25 years of Image analysis. Nat. Methods 2012, 9, 671–675. [Google Scholar] [CrossRef]
- Mikhailov, K.E. Classification of fossil eggshells of Amniotic vertebrates. Acta Paleontol. Pol. 1991, 36, 193–238. [Google Scholar]
- Mikhailov, K.E. Avian Eggshells: An Atlas of Scanning Electron Micrographs; British Ornithologist´ Club: Newbury, UK, 1997. [Google Scholar]
- Brooke, M. Albatrosses and Petrels Across the World; Oxford University Press: Oxford, UK, 2004. [Google Scholar]
- Bond, A.L.; Hobson, K.A. Replaying propensity and characteristics of replacement clutches of Leach’s Storm-Petrels (Oceanodroma leucorhoa). Can. J. Zool. 2015, 93, 181–185. [Google Scholar] [CrossRef]
- Megyesi, J.L.; O’Daniel, D.L. Bulwer’s Petrel (Bulweria bulwerii), version 1.0. In Birds of the World; Billerman, S.M., Ed.; Cornell Lab of Ornithology: Ithaca, NY, USA, 2020. [Google Scholar] [CrossRef]
- Cuthbert, R.J. Breeding biology, chick growth and provisioning of Great Shearwaters (Puffinus gravis) at Gough Island, South Atlantic Ocean. Emu 2005, 105, 305–310. [Google Scholar] [CrossRef]
- Thibault, J.C.; Bretagnolle, V.; Rabouam, C. Calonectris diomedea Cory’s Shearwater. BWP Update 1997, 1, 75–98. [Google Scholar]
- Lee, D.S.; Haney, J.C.; Carboneras, C.; Jutglar, F.; Kirwan, G.M. Manx Shearwater (Puffinus puffinus), version 1.0. In Birds of the World; Billerman, S.M., Ed.; Cornell Lab of Ornithology: Ithaca, NY, USA, 2020. [Google Scholar] [CrossRef]
- Ranzoni, A.; Focardi, S.; Fossi, C.; Leonzio, C.; Mayol, J. Comparison between concentrations of mercury and other contaminants in eggs and tissues of Cory’s Shearwater Calonectris diomedea collected on Atlantic and Mediterranean Islands. Environ. Pollut. Ser. A 1986, 40, 17–35. [Google Scholar] [CrossRef]
- Schönwetter, S. Handbuch der Oologie: Band I; Akademie Verlag: Berlin, Germany, 1967. [Google Scholar]
- Núñez-Lahuerta, C.; Moreno-Azanza, M.; Pérez-Pueyo, M. First approach for a taphonomic key for fossil eggs and eggshells accumulations using optic microscopy: The case of Blasi-2B (Upper Cretaceous, Spain). Ciências Terra Procedia 2021, 1, 23–53. [Google Scholar] [CrossRef]
- Taivalkoski, A.; Holt, E. The effects of cooking on avian eggshell microstructure. J. Archaeol. Sci. Reports 2016, 6, 64–76. [Google Scholar] [CrossRef]





Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Núñez-Lahuerta, C.; Moreno-Azanza, M.; Pérez-Pueyo, M.; Del-Arco-Aguilar, M.d.C.; Del-Arco-Aguilar, M.; Siverio-Batista, C.; Castillo-Ruiz, C.; Cruzado-Caballero, P. Shearwater Eggs in Lobos 3, a Holocene Site of Fuerteventura (Canary Islands). Diversity 2023, 15, 144. https://doi.org/10.3390/d15020144
Núñez-Lahuerta C, Moreno-Azanza M, Pérez-Pueyo M, Del-Arco-Aguilar MdC, Del-Arco-Aguilar M, Siverio-Batista C, Castillo-Ruiz C, Cruzado-Caballero P. Shearwater Eggs in Lobos 3, a Holocene Site of Fuerteventura (Canary Islands). Diversity. 2023; 15(2):144. https://doi.org/10.3390/d15020144
Chicago/Turabian StyleNúñez-Lahuerta, Carmen, Miguel Moreno-Azanza, Manuel Pérez-Pueyo, M. del Carmen Del-Arco-Aguilar, Mercedes Del-Arco-Aguilar, Celia Siverio-Batista, Carolina Castillo-Ruiz, and Penélope Cruzado-Caballero. 2023. "Shearwater Eggs in Lobos 3, a Holocene Site of Fuerteventura (Canary Islands)" Diversity 15, no. 2: 144. https://doi.org/10.3390/d15020144