
An Eudromaeosaurian Theropod from Lo Hueco (Upper Cretaceous. Central Spain)


Abstract
:1. Introduction
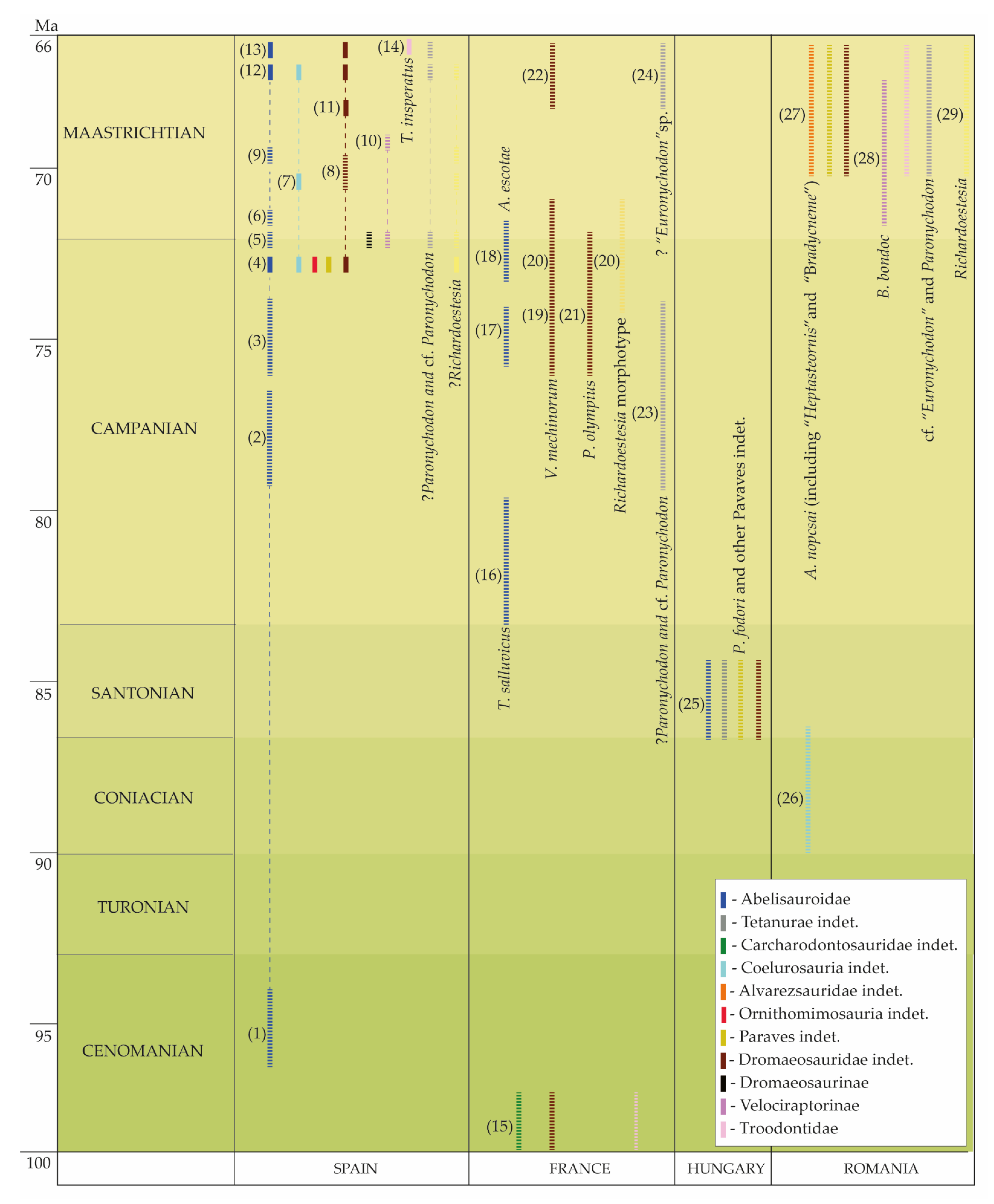
1.1. The Fossil Record of Late Cretaceous Theropod Dinosaurs in European Landmasses
1.1.1. Campanian to Lower Maastrichtian
1.1.2. Late Maastrichtian
2. Geographical and Geological Context
3. Materials and Methods
4. Discussion and Results
4.1. Systematic Paleontology
- Dinosauria Owen, 1842 [142]
- Theropoda Marsh, 1881 [143]
- Coelurosauria von Huene, 1914 [144]
- Eudromaeosauria Longrich and Currie, 2009 [30]
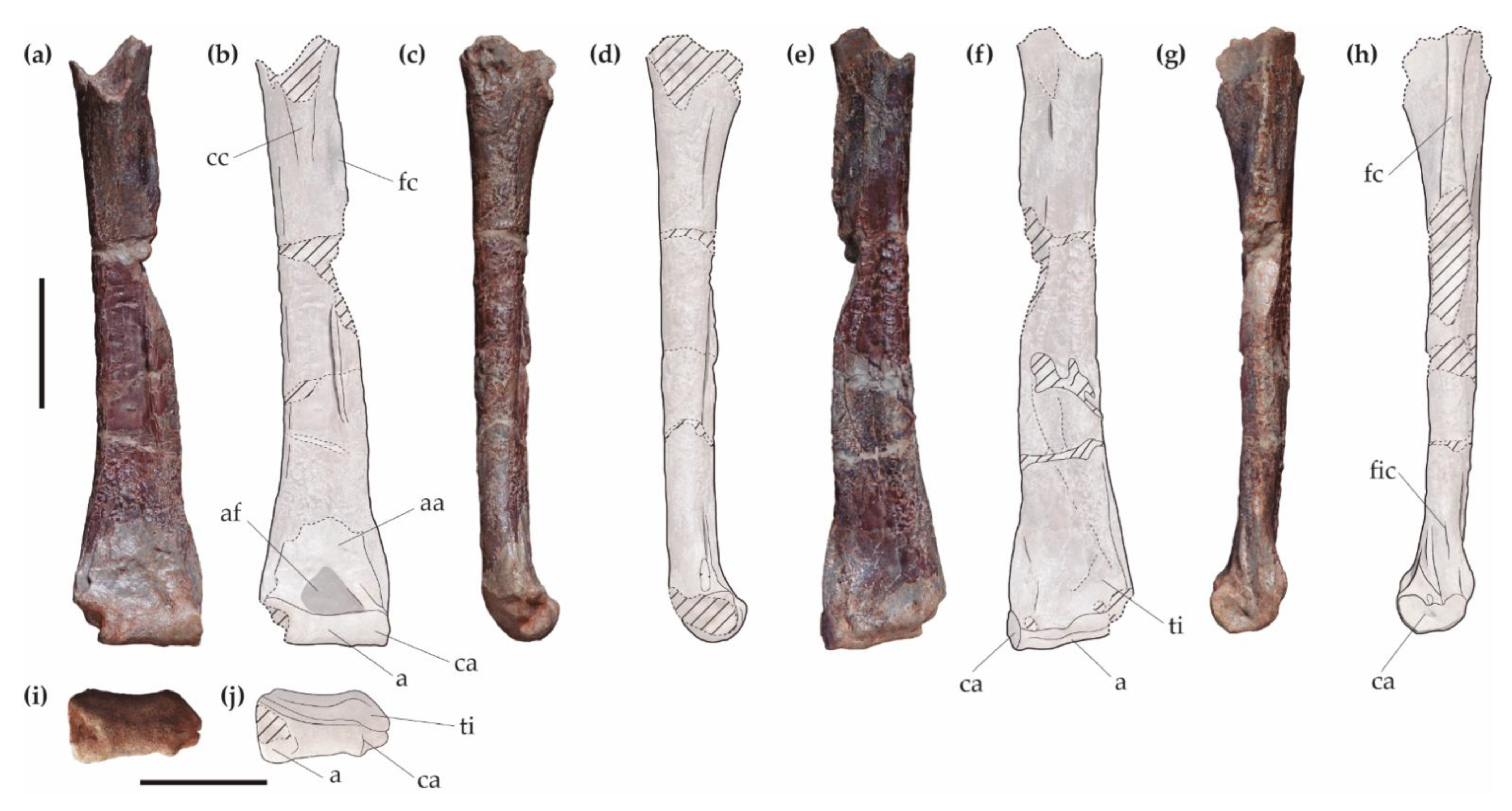
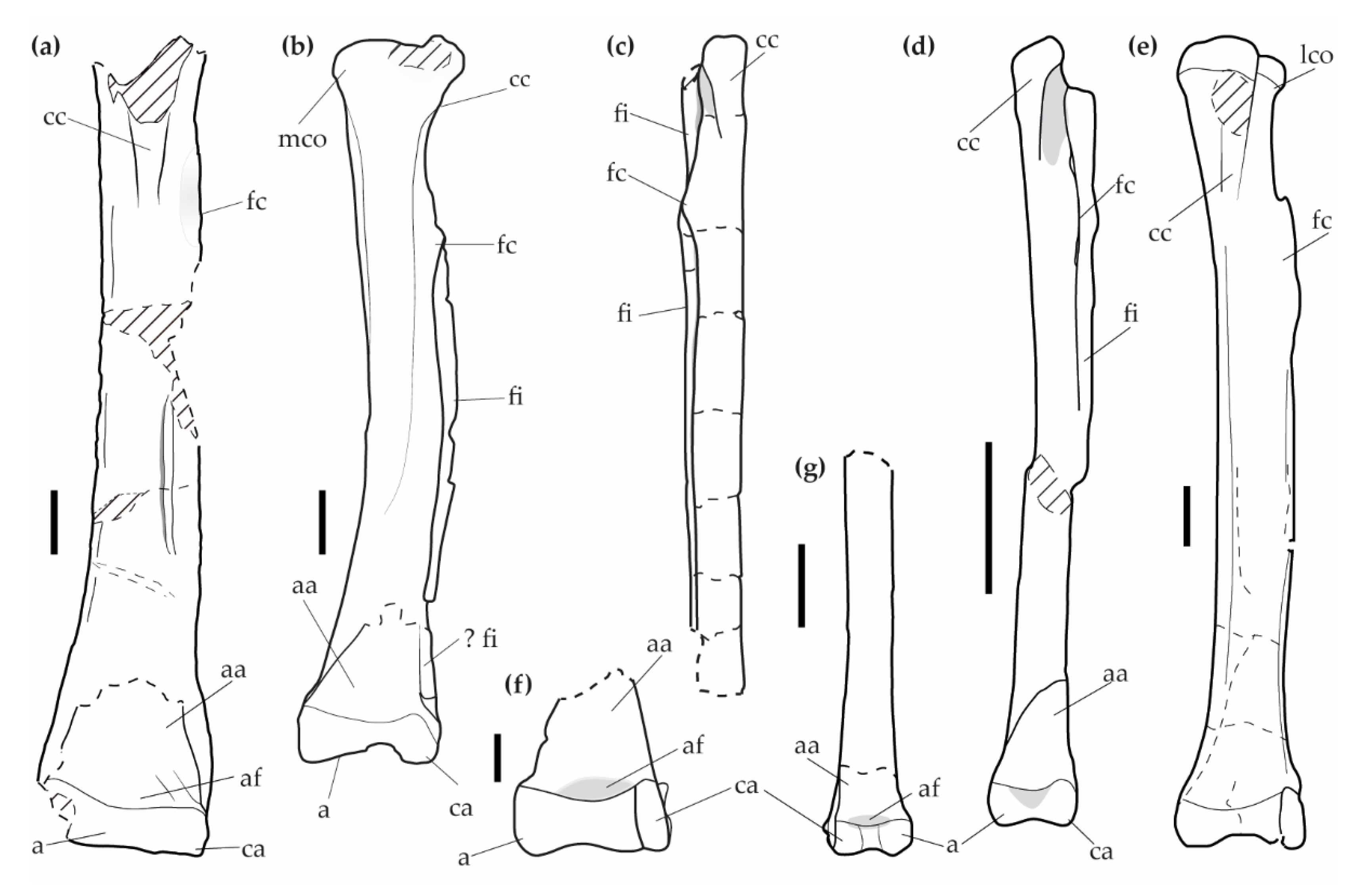
4.1.1. Description
4.1.2. Comparison
5. Discussion and Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Allain, A.; Suberbiola, X.P. Dinosaurs of France. C. R. Palevol. 2003, 2, 27–44. [Google Scholar] [CrossRef]
- Pereda-Suberbiola, X. Biogeographical affinities of Late Cretaceous continental tetrapods of Europe: A review. Bsgf Earth Sci. Bull. 2009, 180, 57–71. [Google Scholar] [CrossRef]
- Riera, V.; Oms, O.; Gaete, R.; Galobart, À. The end-Cretaceous dinosaur succession in Europe: The Tremp Basin record (Spain). Palaeogeogr. Palaeoclim. Palaeoecol. 2009, 283, 160–171. [Google Scholar] [CrossRef]
- Ősi, A.; Rabi, M.; Makádi, L.; Szentesi, Z.; Botfalvai, G.; Guly, P. The Late Cretaceous continental vertebrate fauna from Iharkút (western Hungary): A review. In Bernissart Dinosaurs and Early Cretaceous Terrestrial Ecosystems; Godefroit, P., Ed.; Indiana University Press: Bloomington, IN, USA, 2012; pp. 533–568. [Google Scholar]
- Csiki-Sava, Z.; Buffetaut, E.; Ősi, A.; Suberbiola, X.P.; Brusatte, S.L. Island life in the Cretaceous-faunal composition, biogeography, evolution, and extinction of land-living vertebrates on the Late Cretaceous European archipelago. ZooKeys 2015, 469, 1–161. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Suberbiola, X.P.; Corral, J.C.; Astibia, H.; Badiola, A.; Bardet, N.; Berreteaga, A.; Buffetaut, E.; Buscalioni, A.D.; Cappetta, H.; Cavin, L.; et al. Late Cretaceous continental and marine vertebrate assemblages of the Laño Quarry (Basque-Cantabrian Region, Iberian Peninsula): An update. J. Iber. Geol. 2015, 41, 101–124. [Google Scholar] [CrossRef] [Green Version]
- Canudo, J.I.; Oms, O.; Vila, B.; Galobart, A.; Fondevilla, V.; Puertolas-Pascual, E.; Selles, A.G.; Cruzado-Caballero, P.; Dinares-Turell, J.; Vicens, E.; et al. The upper Maastrichtian dinosaur fossil record from the southern Pyrenees and its contribution to the topic of the Cretaceous-Palaeogene mass extinction event. Cretac. Res. 2016, 57, 540–551. [Google Scholar] [CrossRef]
- Pérez-García, A.; Bardet, N.; Fregenal-Martínez, M.A.; Martín-Jiménez, M.; Mocho, P.; Narvaéz, I.; Torices, A.; Vullo, R.; Ortega, F. Cenomanian vertebrates from Algora (central Spain): New data on the establishment of the European Upper Cretaceous continental faunas. Cretac. Res. 2020, 115, 104566. [Google Scholar] [CrossRef]
- Antunes, M.T.; Pais, J. Notas sobre depósitos de Taveiro. Estratigrafia, Paleontologia, Idade, Paleoecologia. Ciências Terra 1978, 4, 109–128. [Google Scholar]
- Le Loeuff, J.; Buffetaut, E. Tarascosaurus salluvicus nov. gen., nov. sp., dinosaure théropode du Crétacé supérieur du Sud de la France. Geobios 1991, 24, 585–594. [Google Scholar] [CrossRef]
- Ősi, A.; Apesteguía, S.; Kowalewski, M. Non-avian theropod dinosaurs from the early Late Cretaceous of central Europe. Cretac. Res. 2010, 31, 304–320. [Google Scholar] [CrossRef]
- Tortosa, T.; Buffetaut, E.; Vialle, N.; Dutour, Y.; Turini, E.; Cheylan, G. A new abelisaurid dinosaur from the Late Cretaceous of southern France: Palaeobiogeographical implications. Ann. Paléontologie 2014, 100, 63–86. [Google Scholar] [CrossRef]
- Buffetaut, E.; Angst, D.; Mechin, P.; Mechin-Salessy, A. New remains of the giant bird Gargantuavis philoinos from the Late Cretaceous of Provence (south-eastern France). PalaeoVertebrata 2015, 39, e3. [Google Scholar] [CrossRef] [Green Version]
- Torices, A.; Currie, P.J.; Canudo, J.I.; Pereda-Suberbiola, X. Theropod dinosaurs from the Upper Cretaceous of the South Pyrenees Basin of Spain. Acta Palaeontol. Pol. 2015, 60, 611–626. [Google Scholar] [CrossRef] [Green Version]
- Vila, B.; Sellés, A.G.; Brusatte, S.L. Diversity and faunal changes in the latest Cretaceous dinosaur communities of southwestern Europe. Cretac. Res. 2016, 57, 552–564. [Google Scholar] [CrossRef]
- Fondevilla, V.; Dinarès-Turell, J.; Vila, B.; Le Loeuff, J.; Estrada, R.; Oms, O.; Galobart, À. Magnetostratigraphy of the Maastrichtian continental record in the Upper Aude Valley (northern Pyrenees, France): Placing age constraints on the succession of dinosaur-bearing sites. Cretac. Res. 2016, 57, 457–472. [Google Scholar] [CrossRef]
- Sellés, A.G.; Vila, B.; Brusatte, S.L.; Currie, P.J.; Galobart, À. A fast-growing basal troodontid (Dinosauria: Theropoda) from the latest Cretaceous of Europe. Sci. Rep. 2021, 11, 4855. [Google Scholar] [CrossRef]
- Isasmendi, E.; Torices, A.; Canudo, J.I.; Currie, P.J.; Pereda-Suberbiola, X. Upper Cretaceous European theropod palaeobiodiversity, palaeobiogeography and the intra-Maastrichtian faunal turnover: New contributions from the Iberian fossil site of Laño. Pap. Palaeontol. 2022, 8, e1419. [Google Scholar] [CrossRef]
- Barroso-Barcenilla, F.; Cambra-Moo, O.; Escaso, F.; Ortega, F.; Pascual, A.; Pérez-García, A.; Rodríguez-Lázaro, J.; Sanz, J.L.; Segura, M.; Torices, A. New and exceptional discovery in the Upper Cretaceous of the Iberian Peninsula: The palaeontological site of ‘‘Lo Hueco’’, Cuenca, Spain. Cretac. Res. 2009, 30, 1268–1278. [Google Scholar] [CrossRef]
- Narváez, I.; Brochu, C.A.; Escaso, F.; Pérez-García, A.; Ortega, F. New Spanish Late Cretaceous eusuchian reveals the synchronic and sympatric presence of two allodaposuchids. Cretac. Res. 2016, 65, 112–125. [Google Scholar] [CrossRef] [Green Version]
- Torices, A.; Diaz Berenguer, E.; Narvaéz, I.; Ortega, F.; Perez, S.; Serrano, H. Preliminary analysis of the microvertebrate fossils of “Lo Hueco” (Upper Cretaceous, Cuenca, Spain). In Mésogée-Bulletin du Muséum d’histoire Naturelle de Marseille 66, Proceedings of the 8ème Congrès de l’European Association of Vertebrate Palaeontology (EAVP), Aix-en-Provence, France, 7–12 June 2010; Muséum d’histoire naturelle: Marseille, France, 2012; Volume 74. [Google Scholar]
- Ortega, F.; Bardet, N.; Barroso-Barcenilla, F.; Callapez, P.M.; Cambra-Moo, O.; Daviero-Gómez, V.; Díez Díaz, V.; Domingo, L.; Elvira, A.; Escaso, F.; et al. The biota of the Upper Cretaceous site of Lo Hueco (Cuenca, Spain). J. Iber. Geol. 2015, 41, 83–99. [Google Scholar] [CrossRef] [Green Version]
- Díaz, V.D.; Mocho, P.; Páramo, A.; Escaso, F.; Marcos-Fernández, F.; Sanz, J.L.; Ortega, F. A new titanosaur (Dinosauria, Sauropoda) from the Upper Cretaceous of Lo Hueco (Cuenca, Spain). Cretac. Res. 2016, 68, 49–60. [Google Scholar] [CrossRef]
- Páramo, A.; Escaso, F.; Mocho, P.; Marcos-Fernandez, F.; Sanz, J.L.; Ortega, F. 3D geometric morphometrics of the hind limb in the titanosaur sauropods from Lo Hueco (Cuenca, Spain). Cretac. Res. 2022, 134, 105147. [Google Scholar] [CrossRef]
- Ortega, F.; Escaso, F.; Malafaia, E.; Coria, R.A. Aves gigantes en al Cretácico Superior de Lo Hueco (Fuentes, Cuenca). In Proceedings of the Lucas Mallada 23, Libro de Resúmenes de las XXXVI Jornadas de la Sociedad Española de Paleontología, Zaragoza, Spain, 29 September–2 October 2021; pp. 207–208. [Google Scholar]
- Ortega, F.; Malafaia, E.; Escaso, F.; Coria, R.A. New material of theropods (Abelisauroidea?) from Lo Hueco (Late Cretaceous. Cuenca, Central Spain). In Palaeovertebrata, Special Volume 1, Abstract book of the XIX Annual Conference of the European Association of Vertebrate Palaeontologists, Benevento/Pietraroja, Italy, 2–27 July 2022; Belvedere, M., Mecozzi, B., Amore, O., Sardella, R., Eds.; Association Palaeovertebrata: Montpellier, France, 2022; p. 147. [Google Scholar] [CrossRef]
- Csiki, Z.; Vremir, M.; Brusatte, S.L.; Norell, M.A. An aberrant island-dwelling theropod dinosaur from the Late Cretaceous of Romania. PNAS 2010, 107, 15357–15361. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brusatte, S.L.; Vremir, M.; Csiki-Sava, Z.; Turner, A.H.; Watanabe, A.; Erickson, G.M.; Norell, M.A. 2013. The osteology of Balaur bondoc, an island-dwelling dromaeosaurid (Dinosauria: Theropoda) from the Late Cretaceous of Romania. Bull. Am. Mu. Nat. Hist. 2013, 374, 1–100. [Google Scholar] [CrossRef]
- Csiki, Z.; Grigorescu, D. The “dinosaur island”-New interpretation of the Haţeg Basin vertebrate fauna after 110 years. Sargetia 2007, 20, 5–26. [Google Scholar]
- Longrich, N.R.; Currie, P.J. A microraptorine (Dinosauria–Dromaeosauridae) from the Late Cretaceous of North America. PNAS 2009, 106, 5002–5007. [Google Scholar] [CrossRef] [Green Version]
- Turner, A.H.; Makovicky, P.J.; Norell, M.A. A review of dromaeosaurid systematics and paravian phylogeny. Bull. Am. Mu. Nat. Hist. 2012, 371, 1–206. [Google Scholar] [CrossRef]
- Russell, D.A. The role of Central Asia in dinosaurian biogeography. Can. J. Earth Sci. 1993, 30, 2002–2012. [Google Scholar] [CrossRef]
- Evans, D.C.; Larson, D.W.; Currie, P.J. A new dromaeosaurid (Dinosauria: Theropoda) with Asian affinities from the latest Cretaceous of North America. Sci. Nat. 2013, 100, 1041–1049. [Google Scholar] [CrossRef]
- Powers, M.J.; Fabbri, M.; Doschak, M.R.; Bhullar, B.-A.S.; Evans, D.C.; Norell, M.A.; Currie, P.J. A new hypothesis of eudromaeosaurian evolution: CT scans assist in testing and constructing morphological characters. J. Vertebr. Paleontol. 2022, 41, e2010087. [Google Scholar] [CrossRef]
- Ortega, F.; Escaso, F.; Mocho, P.; Narvaéz, I.; Pérez-García, A. Eggs and bones: A preliminary comparison between the Upper Cretaceous faunas of the Poyos, Portilla and Lo Hueco sites (Villalba de la Sierra Formation. Central Spain). In Proceedings of the X Congreso Latinoamericano de Paleontología, El Salvador, Mexico, 4–8 February 2019. [Google Scholar]
- Buffetaut, E.; Mechin, P.; Mechin-Salessy, A. Un dinosaur théropode d’affinités gondwaniennes dans le Crétacé supérieur de Provence. C. R. Acad. Sci. Paris 1988, 306, 153–158. [Google Scholar]
- Pérez-García, A.; Ortega, F.; Bolet, A.; Escaso, F.; Houssaye, A.; Martínez-Salanova, J.; de Miguel Chaves, C.; Mocho, P.; Narvaéz, I.; Segura, M.; et al. A review of the upper Campanian vertebrate site of Armuña (Segovia Province, Spain). Cretac. Res. 2016, 57, 591–623. [Google Scholar] [CrossRef]
- Ősi, A.; Szabó, M.; Tóth, E.; Bodor, E.; Lobitzer, H.; Kvaček, J.; Svobodová, M.; Szente, I.; Wagreich, M.; Trabelsi, K.; et al. A brackish to non-marine aquatic and terrestrial fossil assemblage with vertebrates from the lower Coniacian (Upper Cretaceous) Gosau Group of the Tiefengraben locality near St. Wolfgang im Salzkammergut, Austria. Cretac. Res. 2021, 127, 104938. [Google Scholar] [CrossRef]
- Valentin, X.; Godefroit, P.; Tabuce, R.; Vianey-Liaud, M.; Wu, W.; Garcia, G. First late Maastrichtian (latest Cretaceous) vertebrate assemblage from Provence (Vitrolles-la-Plaine, southern France). In Bernissart Dinosaurs and Early Cretaceous Terrestrial Ecosystems; Godefroit, P., Ed.; Indiana University Press: Bloomington, IN, USA, 2012; pp. 582–597. [Google Scholar]
- López-Martínez, N.; Fernández-Marrón, N.T.; Valle, M.F. The succession of vertebrates and plants across the Cretaceous-Tertiary boundary in the Tremp Formation, Ager Valle (southcentral Pyrenees, Spain). Geobios 1999, 32, 617–627. [Google Scholar] [CrossRef] [Green Version]
- Le Loeuff, J.; Buffetaut, E.; Mechin, P.; Mechin-Salessy, A. The first record of dromaeosaurid dinosaurs (Saurischia, Theropoda) in the Maastrichtian of southern Europe: Palaeobiogeographical implications. Bull. Soc. Géol. France 1992, 163, 337–343. [Google Scholar]
- Andrews, C.W. On some bird remains from the Upper Cretaceous of Transylvania. Geol. Mag. 1913, 10, 193–196. [Google Scholar] [CrossRef] [Green Version]
- Harrison, C.J.O.; Walker, C.A. The Bradycnemidae, a new family of owls from the Upper Cretaceous of Romania. Palaeontology 1975, 18, 563–570. [Google Scholar]
- Pereda-Suberbiola, X.; Astibia, H.; Murelaga, X.; Elorza, J.J.; Gómez-Alday, J.J. Taphonomy of the Late Cretaceous dinosaur-bearing beds of the Laño Quarry (Iberian Peninsula). Palaeogeogr. Palaeoclimatol. Palaeoecol. 2000, 157, 247–275. [Google Scholar] [CrossRef]
- Ősi, A.; Főzy, I. A maniraptoran (Theropoda, Dinosauria) sacrum from the Upper Cretaceous of Haţeg Basin (Romania)-in search of the lost pterosaurs of Baron Franz Nopcsa. N. Jb. Geol. Paläontol. Abh. 2007, 246, 173–181. [Google Scholar] [CrossRef]
- Garcia, G.; Duffaud, S.; Feist, M.; Marandat, B.; Tambareau, Y.; Villatte, J.; Sigé, B. La Neuve, gisement à plantes, invertébrés et vertébrés du Bégudien (Sénonien supérieur continental) du bassin d’Aix-en-Provence. Geodiversitas 2000, 22, 325–348. [Google Scholar]
- Ősi, A. The first dinosaur remains from the Upper Cretaceous of Hungary (Csehbánya Formation, Bakony Mts). Geobios 2004, 37, 749–753. [Google Scholar] [CrossRef]
- Weishampel, D.B.; Jianu, C.-M. New theropod dinosaur material from the Haţeg Basin (Late Cretaceous, Western Romania). N. Jb. Geol. Paläontol. Abh. 1996, 200, 387–404. [Google Scholar] [CrossRef]
- Le Loeuff, J.; Buffetaut, E. A new dromaeosaurid theropod from the Upper Cretaceous of Southern France. Oryctos 1998, 1, 105–112. [Google Scholar]
- Allain, R.; Taquet, P. A new genus of Dromaeosauridae (Dinosauria, Theropoda) from the Upper Cretaceous of France. J. Vert. Paleontol. 2000, 20, 404–407. [Google Scholar] [CrossRef]
- Csiki, Z.; Grigorescu, D. Small theropods of the Late Cretaceous of the Haţeg Basin (Western Romania)-an unexpected diversity at the top of the food chain. Oryctos 1998, 1, 87–104. [Google Scholar]
- Ősi, A.; Szabó, M.; Kollmann, H.; Wagreich, M.; Kalmár, R.; Makádi, L.; Szentesi, Z.; Summesberger, H. Vertebrate remains from the Turonian (Upper Cretaceous) Gosau Group of Gams, Austria. Cretac. Res. 2019, 99, 190–208. [Google Scholar] [CrossRef]
- Seeley, H.G. The reptile fauna of the Gosau Formation preserved in the geological museum of the University of Vienna. Quart. J. Geol. Soc. London 1881, 37, 620–702. [Google Scholar] [CrossRef]
- Averianov, A.O.; Yarkov, A.A. Carnivorous Dinosaurs (Saurischia, Theropoda) from the Maastrichtian of the Volga–Don Interfluve, Russia. Paleontol. J. 2004, 38, 78–82. [Google Scholar]
- Buffetaut, E.; Le Loeuff, J. Late Cretaceous dinosaur faunas of Europe: Some correlation problems. Cretac. Res. 1991, 12, 159–176. [Google Scholar] [CrossRef]
- Garcia, G.; Pereda Suberbiola, X. A new species of Struthiosaurus (Dinosauria: Ankylosauria) from the Upper Cretaceous of Villeveyrac (southern France). J. Vert. Paleontol. 2003, 23, 156–165. [Google Scholar] [CrossRef]
- Buffetaut, E.; Escuillié, F.; Pohl, B. First theropod dinosaur from the Maastrichtian phosphates of Morocco. Kaupia 2005, 14, 3–8. [Google Scholar]
- Fondevilla, V.; Riera, V.; Vila, B.; Sellés, A.G.; Dinarès-Turell, J.; Vicens, E.; Gaete, R.; Oms, O.; Galobart, À. Chronostratigraphic synthesis of the latest Cretaceous dinosaur turnover in south-western Europe. Earth-Sci. Rev. 2019, 191, 168–189. [Google Scholar] [CrossRef]
- Buffetaut, E.; Le Loeuff, J.; Tong, H.; Duffaud, S.; Cavin, L.; Garcia, G.; Ward, D. Un nouveau gisement de vertébrés du Crétacé supérieur à Cruzy (Hérault, Sud de la France). C. R. Acad. Sci. Paris 1999, 328, 203–208. [Google Scholar]
- Buffetaut, E.; Marandat, B.; Sigé, B. Découverte de dents de Deinonychosaures (Saurischia, Theropoda) dans le Crétacé supérieur du Sud de la France. C. R. Acad. Sci. Paris II 1986, 303, 1393–1396. [Google Scholar]
- Chanthasit, P.; Buffetaut, E. New data on the Dromaeosauridae (Dinosauria: Theropoda) from the Late Cretaceous of southern France. Bull. Soc. Géol. France 2009, 180, 145–154. [Google Scholar] [CrossRef]
- Rauhut, O.W.M. The interrelationships and evolution of basal theropod dinosaurs. Spec. Pap. Palaeontol. 2003, 69, 1–213. [Google Scholar]
- Buffetaut, E.; Le Loeuff, J.; Mechin, P.; Mechin-Salessy, A. A large French Cretaceous bird. Nature 1995, 377, 110. [Google Scholar] [CrossRef]
- Buffetaut, E.; Le Loeuff, J. A new giant ground bird from the Upper Cretaceous of southern France. J. Geol. Soc. London 1998, 155, 1–4. [Google Scholar] [CrossRef]
- Buffetaut, E.; Angst, D. New evidence of a giant bird from the Late Cretaceous of France. Geol. Mag. 2013, 150, 173–176. [Google Scholar] [CrossRef] [Green Version]
- Angst, D.; Buffetaut, E.; Corral, J.C.; Pereda-Suberbiola, X. First record of the Late Cretaceous giant bird Gargantuavis philoinos from the Iberian Peninsula. Ann. Paléontol. 2017, 103, 135–139. [Google Scholar] [CrossRef]
- Mayr, G.; Codrea, V.; Solomon, A.; Bordeianu, M.; Smith, T. A well-preserved pelvis from the Maastrichtian of Romania suggests that the enigmatic Gargantuavis is neither an ornithurine bird nor an insular endemic. Cretac. Res. 2020, 106, 104271. [Google Scholar] [CrossRef]
- Buffetaut, E.; Angst, D. Gargantuavis is an insular basal ornithurine: A comment on Mayr et al, 2020, “A well-preserved pelvis from the Maastrichtian of Romania suggests that the enigmatic Gargantuavis is neither an ornithurine bird nor an insular endemic”. Cretac. Res. 2020, 112, 104438. [Google Scholar] [CrossRef]
- Walker, C.A.; Buffetaut, E.; Dyke, G.J. Large euenantiornithine birds from the Cretaceous of southern France, North America and Argentina. Geol. Mag. 2007, 144, 977–986. [Google Scholar] [CrossRef]
- Buffetaut, E. First evidence of enantiornithine birds from the Upper Cretaceous of Europe: Postcranial bones from Cruzy (Hérautt, southern France). Oryctos 1998, 1, 131–136. [Google Scholar]
- Buffetaut, E.; Mechin, P.; Mechin-Salessy, A. An archaic bird (Enantiornithes) from the Upper Cretaceous of Provence (southern France). C. R. Acad. Sci. Paris 2000, 331, 557–561. [Google Scholar] [CrossRef]
- Company, J.; Pereda Suberbiola, X.; Ruiz-Omeñaca, J.I.; Buscalioni, A.D. A new species of Doratodon (Crocodyliformes: Zhiphosuchia) from the Late Cretaceous of Spain. J. Vert. Paleontol. 2005, 25, 343–353. [Google Scholar] [CrossRef]
- Company, J.; Pereda Suberbiola, X.; Ruiz-Omeñaca, J.I. Nuevos restos fósiles del dinosaurio Lirainosaurus (Sauropoda, Titanosauria) en el Cretácico Superior (Campaniano-Maastrichtiano) de la Península Ibérica. Ameghiniana 2009, 46, 391–405. [Google Scholar]
- Company, J.; Torices, A.; Pereda-Suberbiola, X.; Ruiz-Omañaca, J. Theropod teeth from the Late Cretaceous of Chera (Valencia, Eastern Spain). J. Vert. Paleontol. 2009, 29, 81A. [Google Scholar]
- Peyrot, D.; Company, J.; Barrón, E. Palynological data of the Late Cretaceous vertebrate site of Chera (Southwestern Iberian Range, Valencia Province). In Proceedings of the Tenth International Symposium on Mesozoic Terrestrial Ecosystems and Biota, Teruel, Spain, 17–19 September 2009. [Google Scholar]
- Astibia, H.; Buffetaut, E.; Buscalioni, A.D.; Cappetta, H.; Corral, C.; Estes, R.; Garcia-Garmilla, F.; Jaeger, J.J.; Jimenez-Fuentes, J.; Le Loeuff, J.; et al. The fossil vertebrates from Laño (Basque Country, Spain); new evidence on the composition and affinities of the Late Cretaceous continental faunas of Europe. Terra Nova 1990, 2, 460–466. [Google Scholar] [CrossRef]
- Astibia, H.; Murelaga, X.; Suberbiola, X.P.; Elorza, J.J.; Gomez-Alday, J.J. taphonomy and palaeoecology of the Upper Cretaceous continental vertebrate-bearing beds of the Laño quarry (Iberian Peninsula). Est. Mus. Cienc. Nat. Alava 1999, 14, 43–104. [Google Scholar]
- Sanz, J.L.; Powell, J.E.; Le Loeuff, J.; Martínez, R.; Suberbiola, X.P. Sauropod remains from the Upper Cretaceous of Laño (northcentral Spain). Titanosaur phylogenetic relationships. Est. Mus. Cienc. Nat. Alava 1999, 14, 235–255. [Google Scholar]
- Díaz, V.D.; Suberbiola, X.P.; Sanz, J.L. Braincase anatomy of the titanosaurian sauropod Lirainosaurus astibiae from the Late Cretaceous of the Iberian Peninsula. Acta Palaeontol. Pol. 2011, 56, 521–533. [Google Scholar] [CrossRef] [Green Version]
- Isasmendi, E.; Torices, A.; Canudo, J.I.; Pereda-Suberbiola, X. Abelisaurid dinosaurs from the Upper Cretaceous Laño site (Iberian Peninsula). Ciências Terra Procedia 2021, 1, 38–41. [Google Scholar]
- Buffetaut, E.; Pereda-Suberbiola, X.; Corral, J.C. A bird-like tibiotarsus from the Late Cretaceous of Laño (Iberian Peninsula). In Proceedings of the 54th Symposium of Vertebrate Palaeontology and Comparative Anatomy, Paris, France, 13–16 September 2006; pp. 4–5. [Google Scholar]
- Sauvage, H.E. Les vertébrés fossiles du Portugal. Contributions à l’étude des poissons et des reptiles du Jurassique et du Crétacé. Mém. Commum. Serv. Géol. Portugal 1897, 1898, 1–46. [Google Scholar]
- Lapparent, A.F.; Zbyszewski, G. Les dinosauriens du Portugal. Mém. Serv. Géol. Portugal N. S. 1957, 2, 1–63. [Google Scholar]
- Antunes, M.T.; Sigogneau-Russell, D. Nouvelles données sur les dinosaures du Crétacé supérieur du Portugal. C. R. Acad. Sci. Paris II 1991, 313, 113–119. [Google Scholar]
- Galton, P.M. Notes on Dinosauria and Pterodactylia from the Cretaceous of Portugal. N. Jb. Geol. Paläont. Abh. 1994, 194, 253–267. [Google Scholar] [CrossRef]
- Galton, P.M. Notes on Dinosauria from the Upper Cretaceous of Portugal. N. Jb. Geol. Paläont. Mh. 1996, 2, 83–90. [Google Scholar] [CrossRef]
- Antunes, M.T.; Broin, F. Le Crétacé terminal de Beira Litoral, Portugal: Remarques stratigraphiques et écologiques; étude complémentaire de Rosasia soutoi (Chelonii, Bothremydidae). Ciências Terra 1988, 9, 153–200. [Google Scholar]
- Antunes, M.T.; Sigogneau-Russell, D. La faune de petits dinosaures du Crétacé terminal portugais. Comum. Serv. Geol. Portugal 1992, 78, 49–62. [Google Scholar]
- Rauhut, O.W.M. Dinosaur teeth from the Barremian of Uña, Province of Cuenca, Spain. Cretac. Res. 2002, 23, 255–263. [Google Scholar] [CrossRef]
- Sues, H.-D.; Averianov, A. Enigmatic teeth of small theropod dinosaurs from the Upper Cretaceous (Cenomanian–Turonian) of Uzbekistan. Can. J. Earth Sci. 2013, 50, 306–314. [Google Scholar] [CrossRef]
- Pérez-García, A.; Coria, R.A.; Gascó, F.; de la Horra, R.; Martín-Jiménez, M.; Mocho, P.; Narvaéz, I.; Ortega, F. El área de nidificación de dinosaurios del margen occidental del embalse de Buendía (Guadalajara y Cuenca): Relevancia del yacimiento del Cretácico Superior de Poyos (Sacedón). In Cuadernos del Museo Geominero 27, XXXIV Jornadas de Paleontología y IV Congreso Ibérico de Paleontología, Vila Real, Portugal, 26–29 September 2018; Vaz, N., Sá, A.A., Eds.; Instituto Geológico y Minero de España: Madrid, Spain, 2018; pp. 99–100. [Google Scholar]
- Baiano, M.A.; Ortega-Coloma, F.; Coria, R.A. Un metatarsiano de un neotheropod basal del yacimiento de Lo Hueco, Formación “Margas, Arcillas y Yesos de Villalba de la Sierra” (Campaniano-Maastrichtiano, Cretácico Superior, Cuenca. España). In Proceedings of the Libro de Resúmenes de las XXX Jornadas Argentinas de Paleontología de Vertebrados, Buenos Aires, Argentina, 17–20 May 2016; p. 12. [Google Scholar]
- Mateus, O.; Dyke, G.J.; Motchurova-Dekova, N.; Kamenov, G.D.; Ivanov, P. The first record of a dinosaur from Bulgaria. Lethaia 2010, 43, 88–94. [Google Scholar] [CrossRef]
- Wellnhofer, P. Ein Dinosaurier (Hadrosauridae) aus der Oberkreide (Maastricht, Helvetikum-Zone) des bayerischen Alpenvorlandes. Mitt. Bayer. Staatssamml. Für Paläontologie Hist. Geol. 1994, 34, 221–238. [Google Scholar]
- Debeljak, I.; Košir, A.; Buffetaut, E.; Otoničar, B. The Late Cretaceous dinosaurs and crocodiles of Kozina (SW Slovenia): A preliminary study. Mem. Soc. Geol. It. 2002, 57, 193–205. [Google Scholar]
- Godefroit, P.; Motchurova-Dekova, N. Latest Cretaceous hadrosauroid (Dinosauria: Ornithopoda) remains from Bulgaria. C. R. Palevol 2010, 9, 163–169. [Google Scholar] [CrossRef]
- Averianov, A.O.; Lopatin, A.V. Dinosaur fossils from the Upper Cretaceous of Crimea. Paleontol. J. 2019, 53, 398–410. [Google Scholar] [CrossRef]
- Gierliński, G.; Lockley, M. First report of probable therizinosaur (cf. Macropodosaurus) tracks from North America, with notes on the neglected vertebrate ichnofauna of the Ferron Sandstone (Late Cretaceous) of central Utah. In At the Top of the Grand Staircase: The Late Cretaceous of Southern Utah; Titus, A.L., Loewen, M.A., Eds.; Indiana University Press: Bloomington, IN, USA, 2013; pp. 530–535. [Google Scholar]
- Gierliński, G.; Lockley, M. A trackmaker for Saurexallopus: Ichnological evidence for oviraptorosaurian tracks from the Upper Cretaceous of western North America. In At the Top of the Grand Staircase: The Late Cretaceous of Southern Utah; Titus, A.L., Loewen, M.A., Eds.; Indiana University Press: Bloomington, IN, USA, 2013; pp. 526–529. [Google Scholar]
- Laurent, Y.; Bilotte, M.; Le Loeuff, J. Late Maastrichtian continental vertebrates from southwestern France: Correlation with marine fauna. Palaeogeogr. Palaeoclimatol. Palaeoecol. 2002, 187, 121–135. [Google Scholar] [CrossRef]
- López-Martínez, N.; Canudo, J.I.; Ardèvol, L.; Pereda-Suberbiola, X.; Orue-Etxebarria, X.; Cuenca Bescós, G.; Ruiz-Omeñaca, J.I.; Murelaga, X.; Feist, M. New dinosaur sites correlated with Upper Maastrichtian pelagic deposits in the Spanish Pyrenees: Implications for the dinosaur extinction pattern in Europe. Cretac. Res. 2001, 22, 41–61. [Google Scholar] [CrossRef] [Green Version]
- Pereda-Suberbiola, X.; Canudo, J.I.; Company, J.; Cruzado-Caballero, P.; Ruiz-Omeñaca, J.I. Hadrosauroid dinosaurs from the Latest Cretaceous of the Iberian Peninsula. J. Vert. Paleontol. 2009, 29, 946–951. [Google Scholar] [CrossRef]
- Pereda-Suberbiola, X.; Canudo, J.I.; Cruzado-Caballero, P.; Barco, J.L.; López-Martínez, N.; Oms, O.; Ruiz-Omeñaca, J.I. The last hadrosaurid dinosaurs of Europe: A new lambeosaurine from the uppermost Cretaceous of Arén (Huesca, Spain). C. R. Palevol 2009, 8, 559–571. [Google Scholar] [CrossRef] [Green Version]
- Cruzado-Caballero, P.; Pereda-Suberbiola, X.; Ruiz-Omeñaca, J.I. Blasisaurus canudoi gen. et sp. nov., a new lambeosaurine dinosaur (Hadrosauridae) from the Latest Cretaceous of Arén (Huesca, Spain). Can. J. Earth Sci. 2010, 47, 1507–1517. [Google Scholar] [CrossRef]
- Cruzado-Caballero, P.; Ruiz-Omeñaca, J.I.; Canudo, J.I. Evidencias de la coexistência de hadrosaurinos y lambeosaurinos en el Maastrichtiano superior de la Península Ibérica (Arén, Huesca, España). Ameghiniana 2010, 47, 153–164. [Google Scholar] [CrossRef]
- Cruzado-Caballero, P.; Canudo, J.I.; Moreno-Azanza, M.; Ruiz-Omeñaca, J.I. New material and phylogenetic position of Arenysaurus ardevoli, a lambeosaurine dinosaur from the late Maastrichtian of Arén (northern Spain). J. Vert. Paleontol. 2013, 3, 1367–1384. [Google Scholar] [CrossRef]
- Vila, B.; Galobart, A.; Canudo, J.I.; Le Loeuff, J.; Dinares-Turell, J.; Riera, V.; Oms, O.; Tortosa, T.; Gaete, R. The diversity of sauropod dinosaurs and their first taxonomic succession from the latest Cretaceous of southwestern Europe: Clues to demise and extinction. Palaeogeogr. Palaeoclimatol. Palaeoecol. 2012, 350–352, 19–38. [Google Scholar] [CrossRef]
- Vila, B.; Oms, O.; Fondevilla, V.; Gaete, R.; Galobart, A.; Riera, V.; Canudo, J.I. The latest succession of dinosaur tracksites in Europe: Hadrosaur ichnology, track production and paleoenvironments. PLoS ONE 2013, 8, e72579. [Google Scholar] [CrossRef] [Green Version]
- Puértolas-Pascual, E.; Arenillas, I.; Arz, J.A.; Calvín, P.; Ezquerro, L.; García-Vicente, C.; Pérez-Pueyo, M.; Sánchez-Moreno, E.M.; Villalaín, J.J.; Canudo, J.I. Chronostratigraphy and new vertebrate sites from the upper Maastrichtian of Huesca (Spain), and their relation with the K/Pg boundary. Cretac. Res. 2018, 89, 36–59. [Google Scholar] [CrossRef] [Green Version]
- Sellés, A.G.; Vila, B.; Galobart, A. Spheroolithus europaeus, oosp. nov. (Late Maastrichtian, Catalonia), the youngest oological record of hadrosauroids in Eurasia. J. Vert. Paleontol. 2014, 34, 725–729. [Google Scholar] [CrossRef]
- Blanco, A.; Méndez, J.M.; Marmi, J. The fóssil record of the uppermost Maastrichtian Reptile Sandstone (Tremp Formation, northeastern Iberian Peninsula. Spanish J. Palaeontol. 2015, 30, 147–160. [Google Scholar] [CrossRef]
- Torices, A.; Ruiz-Omenaca, J.I.; Canudo, J.I.; López-Martínez, N. Nuevos datos sobre los dinosaurios terópodos (Saurischia: Theropoda) del Cretácico superior de los Pirineos Sur-Centrales (Huesca y Lleida). Geo-Temas 2004, 6, 71–74. [Google Scholar]
- Pérez-Pueyo, M.; Cruzado-Caballero, P.; Moreno-Azanza, M.; Vila, B.; Castanera, D.; Gasca, J.M.; Puértolas-Pascual, E.; Bádenas, B.; Canudo, J.I. The tetrapod fossil record from the uppermost Maastrichtian of the Ibero-Armorican Island: An integrative review based on the outcrops of the western Tremp Syncline (Aragón, Huesca Province, NE Spain). Geosciences 2021, 11, 162. [Google Scholar] [CrossRef]
- Pérez-Pueyo, M.; Puértolas-Pascual, E.; Moreno-Azanza, M.; Cruzado-Caballero, P.; Gasca, J.M.; Núñez-Lahuerta, C.; Canudo, J.I. First record of a giant bird (Ornithuromorpha) from the uppermost Maastrichtian of the Southern Pyrenees, northeast Spain. J. Vert. Paleontol. 2021, 41, e1900210. [Google Scholar] [CrossRef]
- Choi, S.; Moreno-Azanza, M.; Csiki-Sava, Z.; Prondvai, E.; Lee, Y.-N. Comparative crystallography suggests maniraptoran theropod affinities for latest Cretaceous European “Geckoid” eggshell. Pap. Palaeontol. 2020, 6, 265–292. [Google Scholar] [CrossRef]
- Baiano, M.A.; Galobart, À.; Dalla Vecchia, F.M.; Vila, B. First evidence of velociraptorine theropods (Dinosaiuria, Dromaeosauridae) from the Upper Maastrichtian of the Iberian Peninsula. In Proceedings of the Paleontologia I evolució. Field Trip Guide and Abstracts Book Reconstructing the Terrestrial End-Cretaceous Palaeoenvironments in Europe, Tremp, Spain, 16–20 September 2014; p. 54. [Google Scholar]
- Grigorescu, D.; Kessler, E. A new specimen of Elopteryx nopcsai Andrews from the dinosaurian beds of Haţeg Basin. Rev. Roum. Géol. Géogr. Geologie 1980, 24, 171–175. [Google Scholar]
- Elzanowski, A. Birds in Cretaceous ecosystems. Acta Palaeont. Polonica 1983, 28, 75–92. [Google Scholar]
- Kessler, E.; Grigorescu, D.; Csiki, Z. Elopteryx revisited-a new bird-like specimen from the Maastrichtian of the Haţeg Basin (Romania). Acta Palaeontol. Romaniae 2005, 5, 249–258. [Google Scholar]
- Makovicky, P.; Norell, M.A. Troodontidae. In The Dinosauria, 2nd ed.; Weishampel, D.B., Dodson, P., Osmólska, H., Eds.; California University Press: Berkeley, CA, USA, 2004; pp. 184–195. [Google Scholar]
- Martin, L.D. The origin and early radiation of birds. In Perspectives in Ornithology, Essays Presented for the Centennial of the American Ornithologists’ Union 29; Brush, A.H., Clam, G.A., Eds.; Cambridge Univ. Press: Cambridge, UK, 1986; pp. 291–338. [Google Scholar]
- Naish, D.; Dyke, G.J. Heptasteornis was no ornithomimid, troodontid, dromaeosaurid or owl-the first alvarezsaurid (Dinosauria, Theropoda) from Europe. N. Jb. Geol. Paläont. Mh. 2004, 7, 385–401. [Google Scholar] [CrossRef]
- Grigorescu, D.; Venczel, M.; Csiki, Z.; Limberea, R. New latest Cretaceous microvertebrate fossil assemblages from the Haţeg Basin (Romania). Geol. Mijnb. 1999, 78, 301–314. [Google Scholar] [CrossRef]
- Codrea, V.; Smith, T.; Dica, P.; Folie, A.; Garcia, G.; Godefroit, P.; Van Itterbeeck, J. Dinosaur egg nests, mammals and other vertebrates from a new Maastrichtian site of the Haţeg Basin (Romania). C. R. Palevol 2002, 1, 173–180. [Google Scholar] [CrossRef]
- Codrea, V.; Vremir, M.; Jipa, C.; Godefroit, P.; Csiki, Z.; Smith, T.; Fărcaş, C. More than just Nopcsa’s Transylvanian dinosaurs: A look outside the Haţeg Basin. Palaeogeogr. Palaeoclimatol. Palaeoecol. 2010, 293, 391–405. [Google Scholar] [CrossRef]
- Codrea, V.; Godefroit, P.; Smith, T. First discovery of Maastrichtian (latest Cretaceous) terrestrial vertebrates in Rusca Montană Basin (Romania). In Bernissart Dinosaurs and Early Cretaceous Terrestrial Ecosystems; Godefroit, P., Ed.; Indiana University Press: Bloomington, IN, USA, 2012; pp. 570–581. [Google Scholar]
- Smith, T.; Codrea, V.; Săsăran, E.; Van Itterbeck, J.; Bultynck, P.; Csiki, Z.; Dica, P.; Fărcaş, C.; Folie, A.; Garcia, G.; et al. A new exceptional vertebrate site from the Late Cretaceous of the Haţeg Basin (Romania). Stud. Univ. Babeş-Bolyai Geol. 2022, 2002, 321–330. [Google Scholar]
- Csiki, Z.; Ionescu, A.; Grigorescu, D. The Budurone microvertebrate site from the Maastrichtian of the Haţeg Basin—flora, fauna, taphonomy and paleoenvironment. Acta Palaeontol. Rom. 2008, 6, 49–66. [Google Scholar]
- Vasile, Ş. A new microvertebrate site from the Upper Cretaceous (Maastrichtian) deposits of the Haţeg Basin. Sargetia. Acta Mus. Dev. Ser. Sci. Nat. 2008, 21, 5–15. [Google Scholar]
- Vasile, Ş.; Csiki, Z. New Maastrichtian microvertebrates from the Rusca Montană Basin (Romania). Oltenia. Studii şi comunicări. Ştiinţele Naturii 2011, 27, 221–230. [Google Scholar]
- Vasile, Ş.; Grigorescu, D.; Csiki-Sava, Z. Maastrichtian continental microvertebrates from Fărcădeana (Rusca Montană Basin, Romania). In Proceedings of the Fundamental 20, 10th Annual Meeting of the European Association of Vertebrate Palaeontologists, Teruel, Spain, 19–24 June 2012; Royo-Torres, R., Gascó, F., Alcalá, L., Eds.; Fundación Conjunto Paleontológico de Teruel: Teruel, Spain, 2012; pp. 271–273. [Google Scholar]
- Godefroit, P.; Cau, A.; Dong-Yu, H.; Escuillié, F.; Wenhao, W.; Dyke, G. A Jurassic avialan dinosaur from China resolves the early phylogenetic history of birds. Nature 2013, 498, 359–362. [Google Scholar] [CrossRef]
- Foth, C.; Tischlinger, H.; Rauhut, O.W.M. New specimen of Archaeopteryx provides insights into the evolution of pennaceous feathers. Nature 2014, 511, 79–82. [Google Scholar] [CrossRef]
- Cau, A.; Brougham, T.; Naish, D. The phylogenetic affinities of the bizarre Late Cretaceous Romanian theropod Balaur bondoc (Dinosauria, Maniraptora): Dromaeosaurid or flightless bird? PeerJ 2015, 3, e1032. [Google Scholar] [CrossRef] [Green Version]
- von Huene, F. Die fossile Reptil-Ordnung Saurischia. Ihre Entwicklung und Geschichte. Monograph. Geol. Palaeontol. 1932, 1, 1–361. [Google Scholar]
- Russell, D.A. Ostrich dinosaurs from the Late Cretaceous of Western Canada. Can. J. Earth Sci. 1972, 9, 375–402. [Google Scholar] [CrossRef]
- Carrano, M.T.; Benson, R.B.J.; Sampson, S.D. The phylogeny of Tetanurae (Dinosauria: Theropoda). J. Syst. Palaeontol. 2012, 10, 211–300. [Google Scholar] [CrossRef]
- Carrano, M.T.; Sampson, S.D. The phylogeny of Ceratosauria (Dinosauria: Theropoda). J. Syst. Palaeontol. 2008, 6, 183–236. [Google Scholar] [CrossRef] [Green Version]
- Dyke, G.J.; Dortang, R.W.; Jagt, J.W.M.; Mulder, E.W.A.; Schulp, A.S.; Chiappe, L.M. Europe’s last Mesozoic bird. Sci. Nat. 2002, 89, 408–411. [Google Scholar] [CrossRef]
- Dyke, G.J.; Schulp, A.S.; Jagt, J.W.M. Bird remains from the Maastrichtian type area (Late Cretaceous). Neth. J. Geosci. 2008, 87, 353–358. [Google Scholar] [CrossRef] [Green Version]
- Sereno, P.C. A rationale for phylogenetic definitions, with application to the higher-level taxonomy of Dinosauria. N. Jb. Geol. Paläont. Abh. 1998, 210, 41–83. [Google Scholar] [CrossRef]
- Owen, R. Report on British fossil reptiles. Rep. Br. Assoc. Adv. Sci. 1887, 58, 698–699. [Google Scholar]
- Marsh, O.C. Principal characters of American Jurassic dinosaurs. Part V. Am. J. Sci. 1881, 3, 417–423. [Google Scholar] [CrossRef]
- von Huene, F. Vollständige Osteologie eines Plateosauriden aus dem schwäbischen Keuper. Geol. Palaeontol. 1926, 15, 139–179. [Google Scholar]
- Gauthier, J.A. Saurischian monophyly and the origin of birds. Mem. Calif. Acad. Sci. 1986, 8, 1–55. [Google Scholar]
- Colbert, E.H.; Russell, D.A. The small Cretaceous dinosaur Dromaeosaurus. Am. Mus. Novit. 1969, 2380, 1–49. [Google Scholar]
- Matthew, W.D.; Brown, B. The family Deinodontidae, with notice of a new genus from the Cretaceous of Alberta. Bull. Am. Mu. Nat. Hist. 1922, 46, 367–385. [Google Scholar]
- Barsbold, R. Carnivorous dinosaurs from the Late Cretaceous of Mongolia. Sovmest. Sov.-Mong. Paleontol. Ekspiditsiya Tr. 1983, 19, 1–119, In Russian. [Google Scholar]
- Currie, P.J. New information on the anatomy and relationships of Dromaeosaurus albertensis (Dinosauria: Theropoda). J. Vert. Paleontol. 1995, 15, 576–591. [Google Scholar] [CrossRef]
- Makovicky, P.J.; Apesteguía, S.; Gianechini, F.A. A new coelurosaurian theropod from the La Buitrera fossil locality of Río Negro, Argentina. Fieldiana Life Earth Sci. 2012, 5, 90–98. [Google Scholar] [CrossRef]
- Sereno, P.C. Early Cretaceous ornithomimosaurs (Dinosauria: Coelurosauria) from Africa. Ameghiniana 2017, 54, 576–616. [Google Scholar] [CrossRef]
- Carpenter, K.; Miles, C.; Ostrom, J.H.; Cloward, K. Redescription of the small maniraptoran theropods Ornitholestes and Coelurus from the Upper Jurassic Morrison Formation of Wyoming. In The Carnivorous Dinosaurs Carpenter; Indiana University Press: Bloomington, IN, USA, 2005; pp. 49–71. [Google Scholar]
- Currie, P.J.; Chen, P. Anatomy of Sinosauropteryx prima from Liaoning, northeastern China. Can. J. Earth Sci. 2001, 38, 1705–1727. [Google Scholar] [CrossRef]
- Peyer, K. A reconsideration of Compsognathus from the Upper Tithonian of Canjuers, southeastern France. J. Vert. Paleontol. 2006, 26, 879–896. [Google Scholar] [CrossRef]
- Altangarel, P.; Chiappe, L.M.; Barsbold, R.; Clark, J.M.; Norell, M.A. Skeletal morphology of Mononykus olecranus (Theropoda: Avialae) from the Late Cretaceous of Mongolia. Am. Mus. Novit. 1994, 3105, 1–29. [Google Scholar]
- Lee, S.; Park, J.-Y.; Lee, Y.-M.; Kim, S.-H.; Lü, J.; Barsbold, R.; Tsogtbaatar, K. A new alvarezsaurid dinosaur from the Nemegt Formation of Mongolia. Sci. Rep. 2019, 9, 15493. [Google Scholar] [CrossRef] [Green Version]
- Zanno, L.E.; Gillette, D.D.; Albright, L.B.; Titus, A.L. A new North American therizinosaurid and the role of herbivory in “predatory” dinosaur evolution. Proc. R. Soc. B Boil. Sci. 2009, 276, 3505–3511. [Google Scholar] [CrossRef] [Green Version]
- Novas, F.E.; Ezcurra, M.D.; Agnolini, F.L.; Pol, D.; Ortíz, R. New Patagonian Cretaceous theropod sheds light about the early radiation of Coelurosauria. Rev. Mus. Argentino Cienc. Nat. 2012, 14, 57–81. [Google Scholar] [CrossRef] [Green Version]
- Gianechini, F.A.; Makovicky, P.J.; Apesteguía, S.; Cerda, I. Postcranial skeletal anatomy of the holotype and referred specimens of Buitreraptor gonzalezorum Makovicky, Apesteguía and Agnolín 2005 (Theropoda, Dromaeosauridae), from the Late Cretaceous of Patagonia. PeerJ 2018, 6, e4558. [Google Scholar] [CrossRef] [PubMed]
- Chiappe, L.M.; Norell, M.A.; Clark, J.M. The skull of a relative of the stem-group bird Mononykus. Nature 1998, 392, 275–278. [Google Scholar] [CrossRef]
- Chiappe, L.M.; Shu’an, J.; Qiang, J.; Norell, M.A. Anatomy and systematics of the Confuciusornithidae (Theropoda: Aves) from the Late Mesozoic of Northeastern China. Bull. Am. Mus. Nat. Hist. 1999, 242, 1–89. [Google Scholar]
- Forster, C.A.; O’Connor, P.M.; Chiappe, L.M.; Alan, T.H. The osteology of the Late Cretaceous paravian Rahonavis ostromi from Madagascar. Palaeontol. Electronica 2020, 23, a31. [Google Scholar] [CrossRef] [PubMed]
- Gatesy, S.M. Guineafowl hind limb function. I: Cineradiographic analysis and speed effects. J. Morphol. 1999, 240, 115–125. [Google Scholar] [CrossRef]
- Hwang, S.H.; Norell, M.A.; Qiang, J.; Kequin, G. New Specimens of Microraptor zhaoianus (Theropoda: Dromaeosauridae) from Northeastern China. Am. Mus. Novit. 2002, 3381, 1–44. [Google Scholar] [CrossRef]
- Xu, X.; Wang, X.-L. A New dromaeosaur (Dinosauria: Theropoda) from the Early Cretaceous Yixian Formation of western Liaoning. Vert. PalAs. 2004, 42, 111–119. [Google Scholar]
- Xu, X.; Choiniere, J.N.; Pittman, M.; Tan, Q.; Dong, X.; Zhiquan, L.; Tan, L.; Clark, J.M.; Norell, M.A.; Hone, D.W.E.; et al. A new dromaeosaurid (Dinosauria: Theropoda) from the Upper Cretaceous Wulansuhai Formation of Inner Mongolia, China. Zootaxa 2010, 2403, 1–9. [Google Scholar] [CrossRef] [Green Version]
- Gao, C.; Morschhauser, E.M.; Varricchio, D.J.; Liu, J.; Zhao, B. A second soundly sleeping dragon: New anatomical details of the Chinese troodontid Mei long with implications for phylogeny and taphonomy. PLoS ONE 2012, 7, e45203. [Google Scholar] [CrossRef] [Green Version]
- Turner, A.H.; Pol, D.; Norell, M.A. Anatomy of Mahakala omnogovae (Theropoda: Dromaeosauridae), Tögrögiin Shiree, Mongolia. Am. Mus. Novit. 2011, 3722, 1–66. [Google Scholar] [CrossRef]
- Turner, A.H.; Pol, D.; Clarke, J.A.; Erickson, G.M.; Norell, M.A. A basal dromaeosaurid and size evolution preceding avian flight. Science 2007, 317, 1378–1381. [Google Scholar] [CrossRef] [Green Version]
- DePalma, R.A.; Burnham, D.A.; Martin, L.D.; Larson, P.L.; Bakker, R.T. The first giant raptor (Theropoda: Dromaeosauridae) from the Hell Creek Formation. Paleontol. Contrib. 2015, 14, 1–16. [Google Scholar]
- Novas, F.E.; Agnolín, F.L.; Matias, J.; Motta, M.J.; Brisson Egli, F.B. Osteology of Unenlagia comahuensis (Theropoda, Paraves, Unenlagiidae) from the Late Cretaceous of Patagonia. Anat Rec. 2021, 304, 2741–2788. [Google Scholar] [CrossRef]
- Novas, F.E.; Pol, D.; Canale, J.I.; Porfiri, J.D.; Calvo, J.O. A bizarre Cretaceous theropod dinosaur from Patagonia and the evolution of Gondwanan dromaeosaurids. Proc. R. Soc. B Boil. Sci. 2009, 276, 1101–1107. [Google Scholar] [CrossRef]
- Holtz, T.R., Jr. Dinosaurs: The Most Complete, Up-to-Date Encyclopedia for Dinosaur Lovers of All Ages; Random House Books for Young Readers: Toronto, ON, Canada, 2008; p. 432. [Google Scholar]
- Kirkland, J.I.; Gaston, R.; Burge, D. A large dromaeosaur (Theropoda) from the Lower Cretaceous of eastern Utah. Hunteria 1993, 2, 1–16. [Google Scholar]
- Ostrom, J.H. A new theropod dinosaur from the Lower Cretaceous of Montana. Postilla 1969, 128, 1–17. [Google Scholar]
- Parsons, W.L.; Parsons, K.M. Morphological Variations within the Ontogeny of Deinonychus antirrhopus (Theropoda, Dromaeosauridae). PLoS ONE 2015, 10, e0121476. [Google Scholar] [CrossRef] [Green Version]
- Brunham, D.A.; Derstler, K.L.; Currie, P.J.; Bakker, R.T.; Zhou, Z.; Ostrom, J.H. Remarkable new birdlike dinosaur (Theropoda: Maniraptora) from the Upper Cretaceous of Montana. Paleontol. Contrib. New Ser. 2000, 13, 1–14. [Google Scholar]
- Norell, M.A.; Makovicky, P.J. Important features of the dromaeosaurid skeleton II: Information from newly collected specimens of Velociraptor mongoliensis. Am. Mus. Novit. 1999, 3282, 1–45. [Google Scholar]
- Perle, A.; Norell, M.; Clark, J. A new maniraptoran Theropod-Achillobator giganticus (Dromaeosauridae)-from the Upper Cretaceous of Burkhant, Mongolia. Contrib. Mong. Am. Mus. Paleontol. Proj. 1999, 101, 1–104. [Google Scholar]
- Novas, F.E.; Egli, F.B.; Agnolin, F.L.; Gianechini, F.A.; Cerda, I. Postcranial osteology of a new specimen of Buitreraptor gonzalezorum (Theropoda, Coelurosauria). Cretac. Res. 2018, 83, 127–167. [Google Scholar] [CrossRef]
- Agnolín, F.L.; Novas, F.E. Avian ancestors: A review of the phylogenetic relationships of the theropods Unenlagiidae, Microraptoria, Anchiornis and Scansoriopterygidae. In SpringerBriefs South America and the Southern Hemisphere; Lohmann, G., Mysak, L.A., Notholt, J., Rabassa, J., Unnithan, V., Eds.; SpringerBriefs in Earth System Science; Springer: Berlin/Heidelberg, Germany, 2013; pp. 1–96. [Google Scholar]
- Hartman, S.; Mortimer, M.; Wahl, W.R.; Lomax, D.R.; Lippincott, J.; Lovelace, D.M. A new paravian dinosaur from the Late Jurassic of North America supports a late acquisition of avian flight. PeerJ 2019, 7, e7247. [Google Scholar] [CrossRef] [PubMed]
- Motta, M.J.; Agnolín, F.L.; Egli, F.B.; Novas, F.E. New theropod dinosaur from the Upper Cretaceous of Patagonia sheds light on the paravian radiation in Gondwana. Sci. Nat. 2020, 107, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Vullo, R.; Neraudeau, D.; Lenglet, T. Dinosaur teeth from the Cenomanian of Charentes, Western France; evidence for a mixed Laurasian-Gondwanan assemblage. J. Vert. Paleontol. 2007, 27, 931–943. [Google Scholar] [CrossRef]
- Vullo, R.; Néraudeau, D. Cenomanian vertebrate assemblages from southwestern France: A new insight into the European mid-Cretaceous continental fauna. Cretac. Res. 2008, 29, 930–935. [Google Scholar] [CrossRef] [Green Version]
- Ősi, A.; Buffetaut, E. Additional non-avian theropod and bird remains from the early Late Cretaceous (Santonian) of Hungary and a review of the European abelisauroid record. Ann. Paléontol. 2011, 97, 35–49. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| Taxa | Cenomanian | Turonian to Early Santonian | Santonian to Early Campanian | Middle Campanian to Late Maastrichtian | ||||
|---|---|---|---|---|---|---|---|---|
| Spain | France | Austria | France | Hungary | Spain | France | Romania | |
| Abelisauroidea indet. | [8] | [11] | [35] | [36] | ||||
| Tarascosaurus | [10] | ? [10] | ||||||
| Arcovenator | ? [18,37] | [12] | ||||||
| Tetanurae indet. | [38] | [11] | ||||||
| Carcharodontosauridae indet. | [39,40] | |||||||
| Coelurosauria indet. | [14] | |||||||
| Elopteryx | ? [41] | [42] | ||||||
| Bradycneme | [43] | |||||||
| “Heptasteornis” | [43] | |||||||
| Maniraptora indet. | [38] | |||||||
| Ornithomimosauria indet. | ? [44] | |||||||
| Paraves indet. | [11] | [18] | [45] | |||||
| Pneumatoraptor | [11] | |||||||
| Dromaeosauridae indet. | [39,40] | [46] | [47] | [14,18] | [48] | |||
| Variraptor | [49] | |||||||
| Pyroraptor | [50] | |||||||
| Dromaeosaurinae indet. | [22] | |||||||
| Velociraptorinae indet. | [22] | ? [49] | [51] | |||||
| Balaur | [27] | |||||||
| Troodontidae indet. | [39,40] | ? [51] | ||||||
| Tamarro | [17] | |||||||
| Paronychodon | [52] | [14,22] | [46] | ? [51] | ||||
| Richardoestesia | [14,22] | |||||||
| Specimen | L | W (mL) | W (ap) | W (mL)/L | Wd (mL) | Wd (ap) | Wd (mL)/L |
|---|---|---|---|---|---|---|---|
| HUE-02696 | 244 * | 31.7 | 18 | 0.12 | 54 | 29 | 0.21 |
| 260 (a) | |||||||
| Mahakala omnogovae (IGM 100/1033) [168,169] | 110 | - | - | - | - | - | - |
| Dakotaraptor steini (PBMNH.P.10.113.T) [170] | 673 | 68.4 (1) | - | 0.10 | - | - | - |
| Unenlagia comahuesis (MCF PVPH 78) [171] | 418 * | - | - | - | 63 * | - | 0.15 |
| Buitreraptor gonzalezorum (MPCA 238 and MPCA 245) [159,172] | >149 | 12.2(1) | 8.5(1) | 0.08 | - | - | - |
| Austroraptor cabazai (MML–195) [172] | 565 | - | - | - | 140 | - | 0.25 |
| Rahonavis ostromi (UA 8656) [162,173] | 118 | 5 | 5 | 0.04 | 10 | 4 | 0.08 |
| Graciliraptor lujiatunensis (IVPP V 13474) [28,168] | 115 | - | - | - | - | - | - |
| Utahraptor ostromaysi (CEU 184v.260) [173,174] | 505 | 76.5 (1) | 47.7 (1) | 0.15 | 145 | - | 0.29 |
| Deinonychus antirrhopus (AMNH 3015) [28,173,175,176] | 312 | - | 18 | - | 63.3 * | - | 0.20 |
| Deinonychus antirrhopus (MCZ 4371) [28,175] | 368 | - | 28.5 | - | 73 | - | 0.20 |
| Saurornitholestes langstoni (MOR 660) [28,169] | 301 | - | - | - | - | - | - |
| Bambiraptor feinbergorum (FIP 001) [30,31,177] | 168 | - | - | - | - | - | - |
| Balaur bondoc (EME PV.313) [27,28] | 153 | 11 | 12 | 0.07 | 25 | - | 0.16 |
| Velociraptor mongoliensis (IGM 100/986) [178] | 255 | 20 (1) | 17.8 (1) | 0.07 | 38.7 | 20 | 0.15 |
| Achillobator giganticus (MNUFR 15) [31,179] | 490 | 60.4 (1) | 39.4 (1) | 0.12 | 113 | 22.4 | 0.23 |
| Linheraptor exquisitus (IVPP V16923) [166] | 255 | - | - | - | - | - | - |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Malafaia, E.; Escaso, F.; Coria, R.A.; Ortega, F. An Eudromaeosaurian Theropod from Lo Hueco (Upper Cretaceous. Central Spain). Diversity 2023, 15, 141. https://doi.org/10.3390/d15020141
Malafaia E, Escaso F, Coria RA, Ortega F. An Eudromaeosaurian Theropod from Lo Hueco (Upper Cretaceous. Central Spain). Diversity. 2023; 15(2):141. https://doi.org/10.3390/d15020141
Chicago/Turabian StyleMalafaia, Elisabete, Fernando Escaso, Rodolfo A. Coria, and Francisco Ortega. 2023. "An Eudromaeosaurian Theropod from Lo Hueco (Upper Cretaceous. Central Spain)" Diversity 15, no. 2: 141. https://doi.org/10.3390/d15020141