
Diversity of the Piscicola Species (Hirudinea, Piscicolidae) in the Eastern Palaearctic with a Description of Three New Species and Notes on Their Biogeography †

Abstract
:1. Introduction
2. Materials and Methods
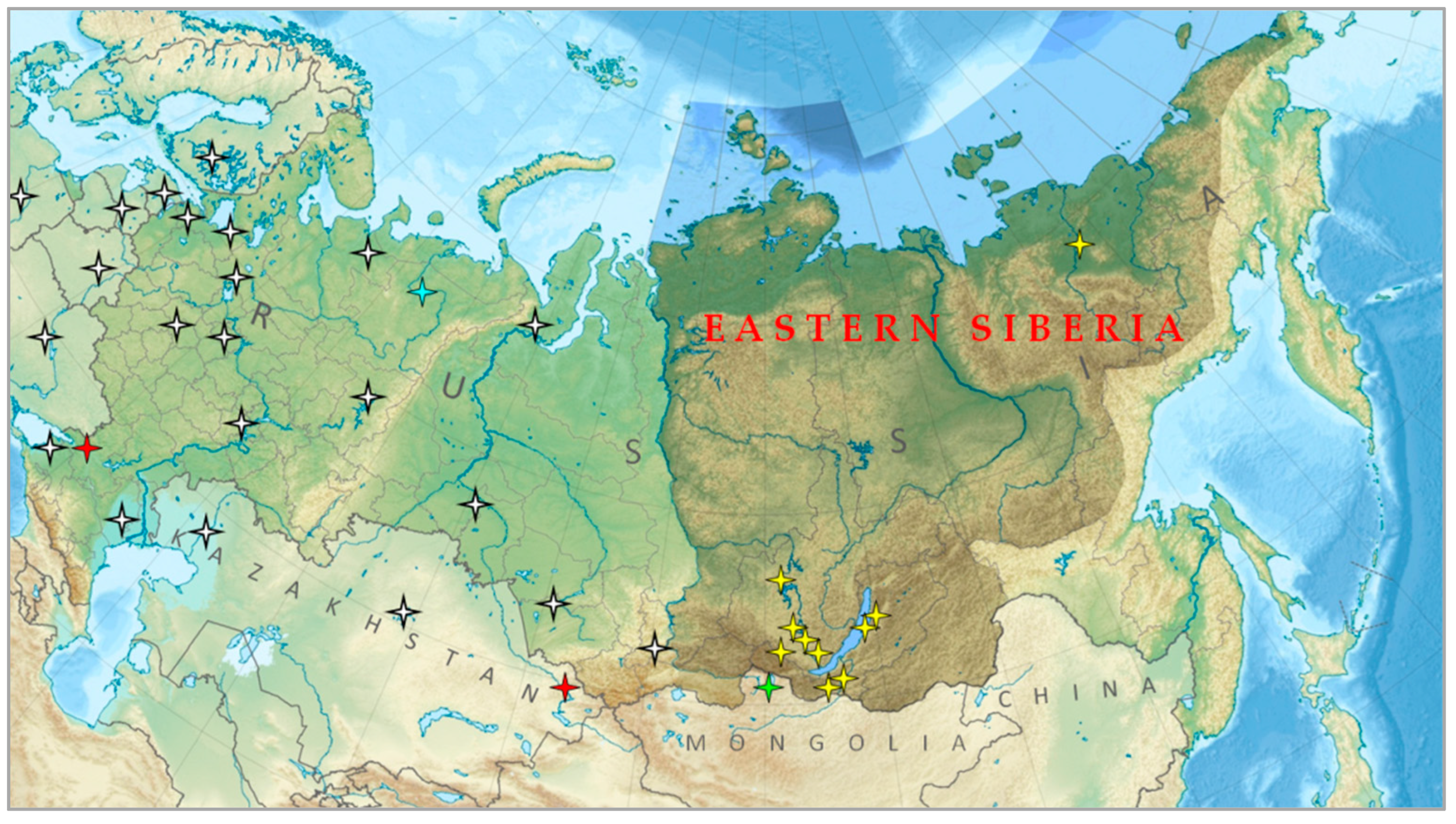
2.1. Study Area
2.2. Sample Collection
2.3. Morphological Analysis
2.4. DNA-Based Species Identification
3. Results
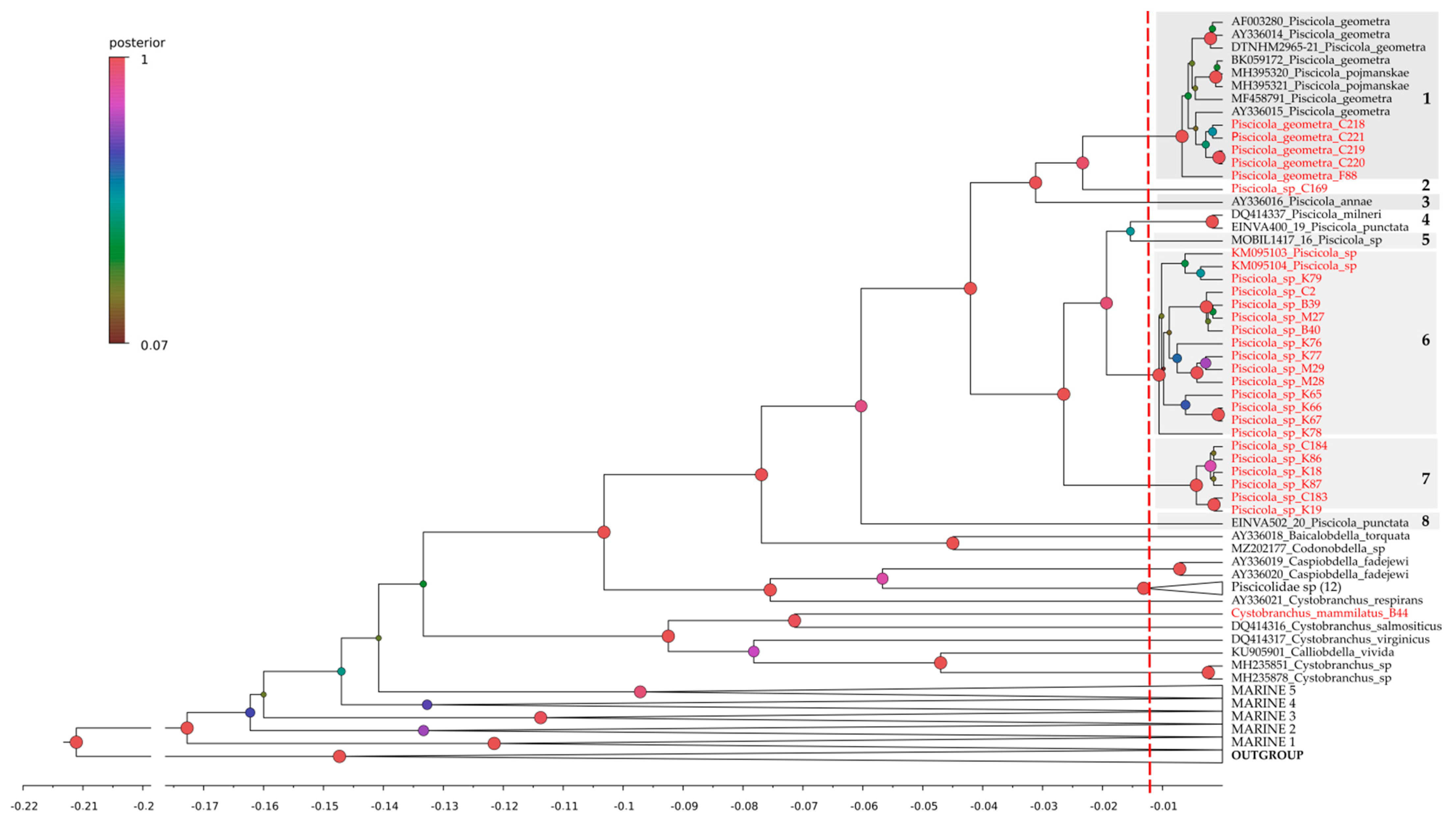
3.1. DNA Identification of Leeches
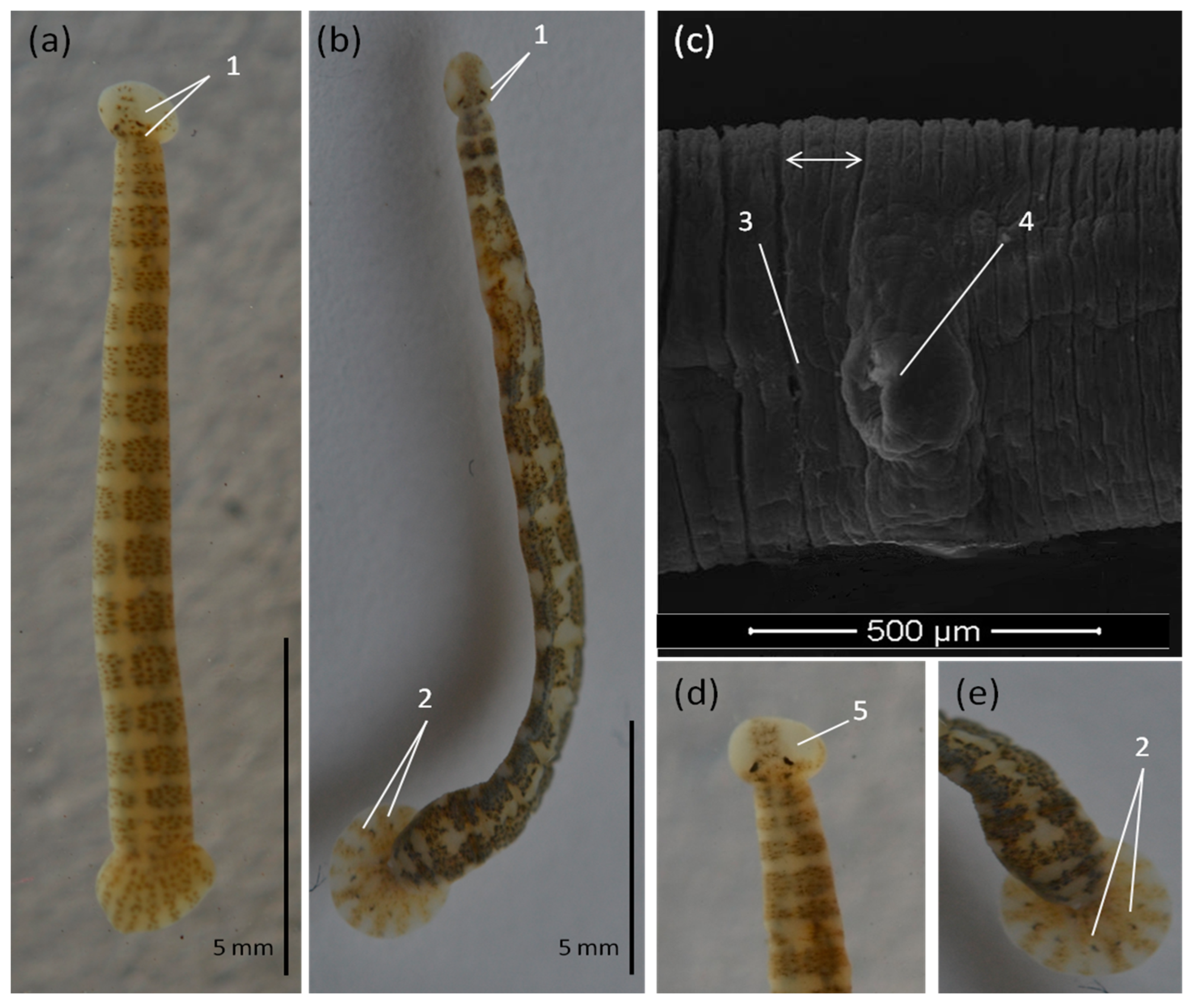
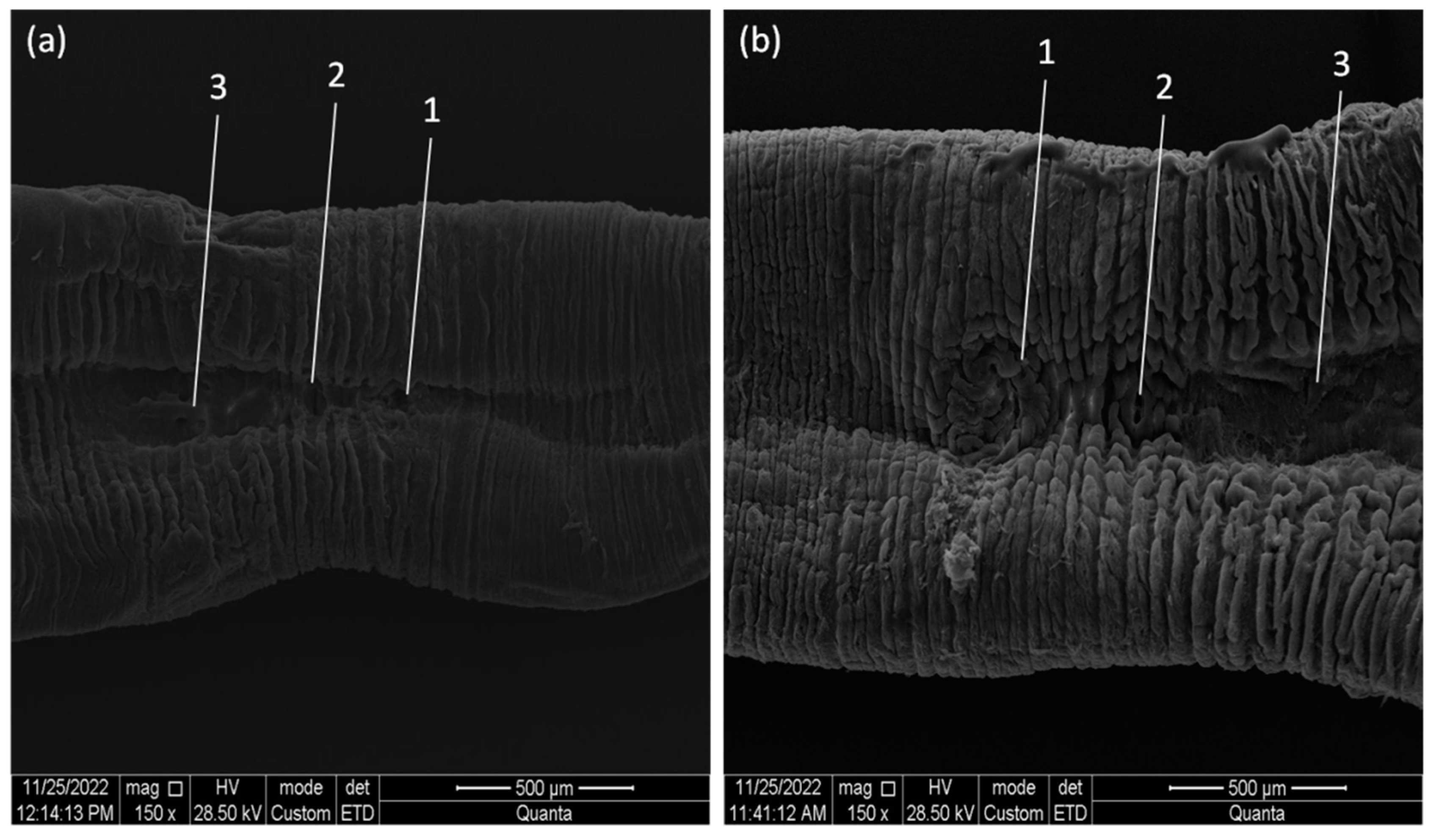
3.2. Morphological Identification of Piscine Leeches
3.3. Taxonomy of Studied Piscicola and Description of New Species
| Phylum: | Annelida Lamarck, 1809 |
| Classis: | Clitellata Michaelsen, 1919 |
| Subclassis: | Hirudinea Lamarck, 1818 |
| Order: | Rhynchobdellida (Blanchard, 1894) |
| Family: | Piscicolidae Johnston, 1865 (syn. Icthyobdellidae Leuckart, 1863) |
| Genus: | Piscicola de Blainville, 1818 |
| Type species: | Piscicola geometra (Linnaeus, 1761) |
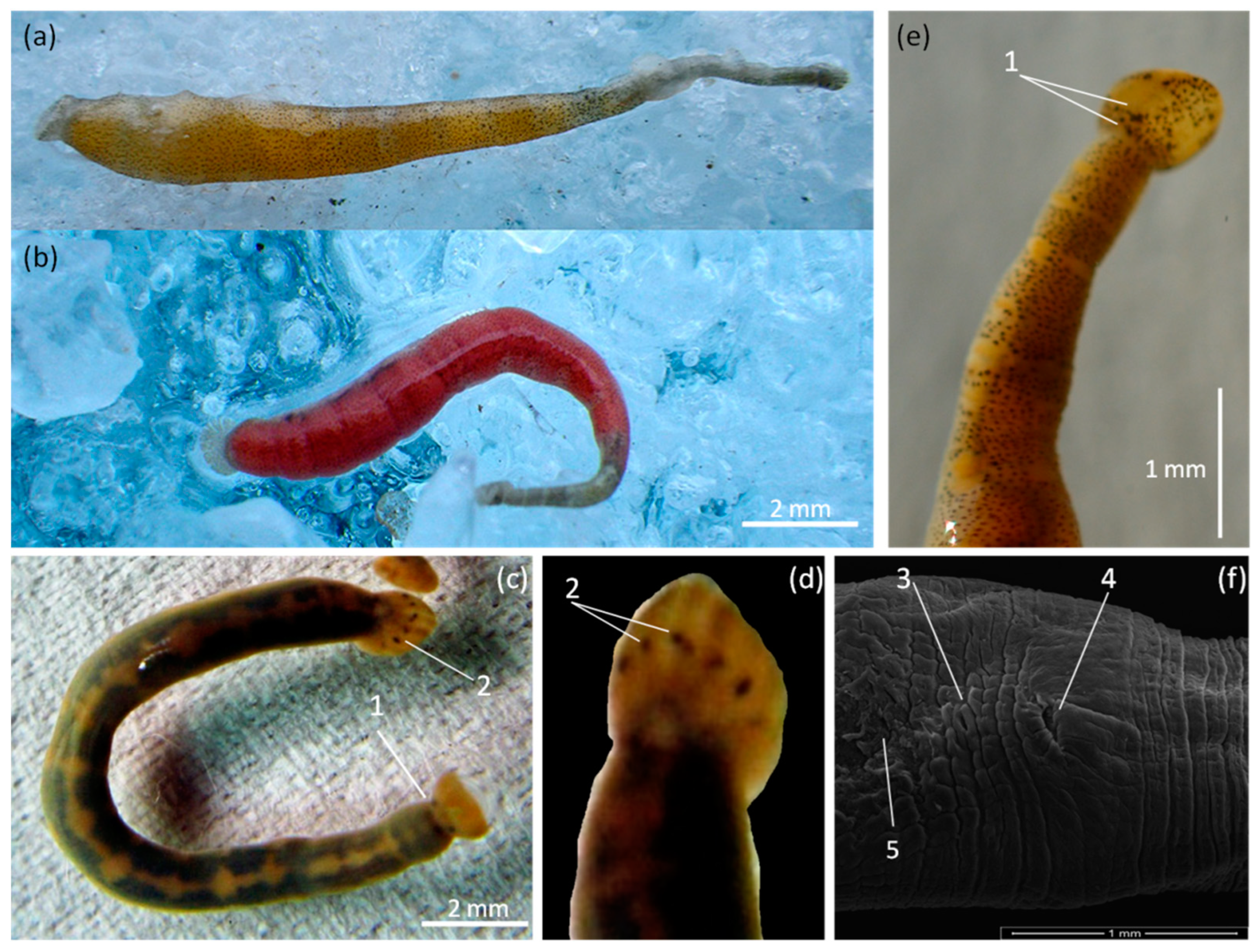
| Species: | Piscicola geometra (Linnaeus, 1758) (Figure 3) |
- Species:
- Piscicola nordica sp. nov. Kaygorodova (Figure 4)
- Species:
- Species:
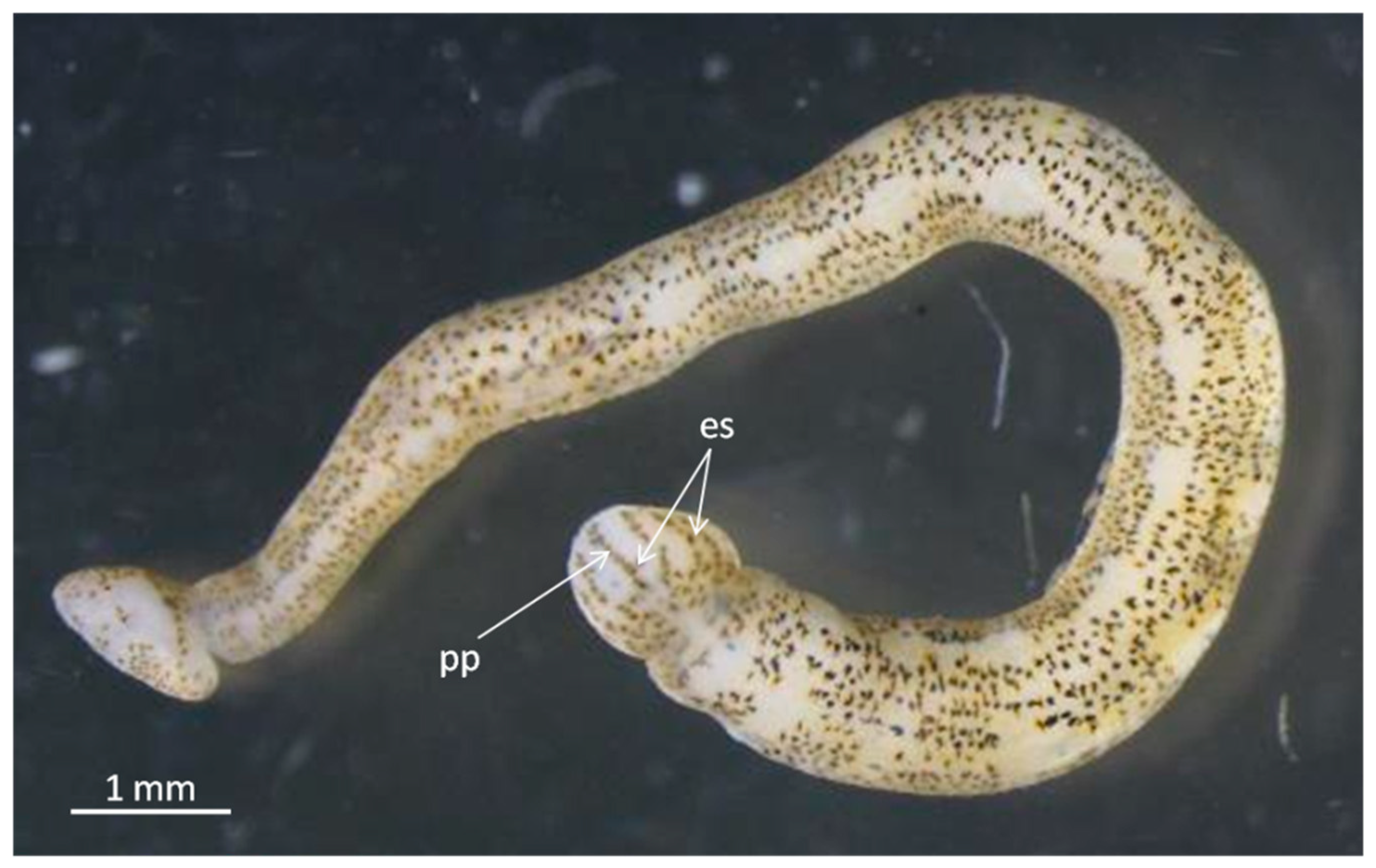
- Piscicola khubsugulensis sp. nov. Kaygorodova (Figure 7)
4. Discussion
4.1. Adjustments in the Piscicola Species Diversity
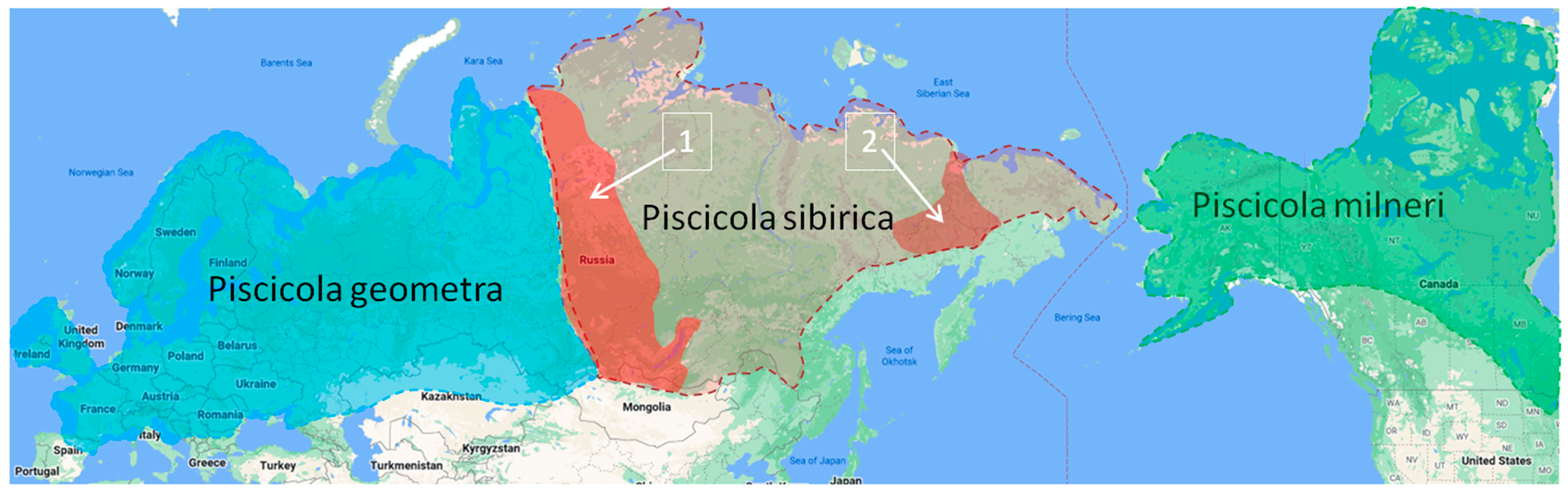
4.2. Biogeography of the Piscicola Fauna
5. Conclusions
- Three new species of the genus Piscicola from the Eastern Palaearctic (P. nordica sp. nov., P. sibirica sp. nov. and P. khubsugulensis sp. nov.) were first discovered using DNA-based identification and were then morphologically differentiated and described.
- The species Piscicola pojmanskae Bielecki 1994 was reduced to a synonym for Piscicola geometra (L., 1761) since they are genetically, morphologically and ecologically identical; the species diversity of the Piscicolidae fauna from the Western Palaearctic is most likely redundant and requires revision.
- Due to these findings, the description of Piscicola geometra (L., 1761) required revision and clarification, which eventually led to the establishment of the neotype.
- The geographic range of Piscicola geometra (L., 1761), previously known as a trans-Palaearctic species, turned out to be limited in the east by the Yenisei River.
- The species Piscicola geometra and Piscicola sibirica sp. nov., the most widespread species in the Palaearctic, as well as the Nearctic Piscicola milneri are geographic vicariates of each other.
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Klemm, D. Leeches (Annelida: Hirudinea) of North America; U.S. Government Printing Office: Cincinnati, OH, USA, 1982; 177p.
- Sket, B. Piscicola hadzii sp. n. (Piscicolidae, Hirudinea), a probably endemic species of leeches from Hercegovina, Yugoslavia. Biol. Vestnik 1985, 33, 89–94. [Google Scholar]
- Epstein, V.M. Leeches. In Key of the Freshwater Fish Parasites of the USSR Fauna; Skarlato, O.A., Ed.; Nauka: Leningrad, USSR, 1987; Volume 3, pp. 340–372. [Google Scholar]
- Nesemann, H.; Neubert, E. Annelida, Clitellata: Branchiobdellida, Acanthobdellea, Hirudinea, Susswasserfauna von Mitteleuropa; Spectrum Akademischer Verlag: Heidelberg, Germany, 1999; 178p. [Google Scholar]
- Bielecki, A. Fish leeches of Poland in relation to the Palaearctic piscicolines (Hirudinea: Piscicolidae: Piscicolinae). Genus. Int. J. Inverteb Taxon. 1997, 8, 223–375. [Google Scholar]
- Bielecki, A. Piscicola brylinskae, a new leech species from the Netherlands (Hirudinea: Piscicolidae). Wiadomości Parazytol. 2001, 47, 119–126. [Google Scholar]
- Bielecki, A.; Cios, S.; Cichocka, J.M.; Pakulnicka, J. Piscicola siddalli n. sp., a Leech species from the United Kingdom (Clitellata: Hirudinida: Piscicolidae). Comp. Parasitol. 2012, 79, 219–230. [Google Scholar] [CrossRef]
- Bielecki, A.; Cichocka, J.M.; Świątek, P.; Gorzel, M. A new leech species (Clitellata: Hirudinida: Piscicolidae) from the Łyna River near Olsztyn, Poland. J. Parasitol. 2013, 99, 467–474. [Google Scholar] [CrossRef] [PubMed]
- Yang, T. Fauna Sinica: Annelida Hirudinea; Science Press: Beijing, China, 1996; 260p. [Google Scholar]
- Suyanti, E.; Mahasri, G.; Lokapirnasari, W.P. Marine leech Zeylanicobdella arugamensis infestation as a predisposing factor for Vibrio alginolyticus infection on the hybrid grouper “Cantang” (Epinephelus fuscoguttatus × Epinephelus lanceolatus) from traditional ponds in the Kampung Kerapu Lamongan, East Java, Indonesia. IOP Conf. Ser. Earth Environ. Sci. 2021, 718, 012035. [Google Scholar] [CrossRef]
- Faisal, M.; Schulz, C.A. Detection of viral hemorrhagic septicemia virus (VHSV) from the leech Myzobdella lugubris Leidy, 1851. Parasit. Vectors 2009, 2, 45. [Google Scholar] [CrossRef] [Green Version]
- Demshin, N.I. Oligochaeta and Hirudinea as Intermediate Hosts of Helminthes; Nauka: Novosibirsk, USSR, 1975; pp. 1–190. [Google Scholar]
- Pyrka, E.; Kanarek, G.; Zaleśny, G.; Hildebrand, J. Leeches as the intermediate host for strigeid trematodes: Genetic diversity and taxonomy of the genera Australapatemon Sudarikov, 1959 and Cotylurus Szidat, 1928. Parasit. Vectors 2021, 14, 44. [Google Scholar] [CrossRef]
- Smit, N.J.; Joubert, A.; Lawton, S.P.; Hayes, P.M.; Cook, C.A. Morphological and molecular characterization of an African freshwater fish trypanosome, including its development in a leech vector. Int. J. Parasitol. 2020, 50, 921–929. [Google Scholar] [CrossRef]
- Ellis, J.; Barratt, J.; Kaufer, A.; Pearn, L.; Armstrong, B.; Johnson, M.; Park, Y.; Downey, L.; Cao, M.; Neill, L.; et al. A new subspecies of Trypanosoma cyclops found in the Australian terrestrial leech Chtonobdella bilineata. Parasitology 2021, 148, 1125–1136. [Google Scholar] [CrossRef]
- Linnaeus, C. Fauna Svecica, 2nd ed.; Sumtu & Literis Direct; Laurentii Salvii: Stockholm, Sweden, 1761; p. 506. [Google Scholar]
- Sket, B.; Trontelj, P. Global diversity of leeches (Hirudinea) in freshwater. Hydrobiologia 2008, 595, 129–137. [Google Scholar] [CrossRef]
- Lukin, E.I. Leeches of Fresh and SalineWaters. In Fauna of the USSR; Nauka: Leningrad, USSR, 1976; pp. 303–351. [Google Scholar]
- Kaygorodova, I.A.; Fedorova, L.I.; Bukin, Y.S. Comparative Analysis of Hirudofauna in the Upper Irtysh Reservoirs Cascade (Eastern Kazakhstan). Proc. Irk. State University. Ser. Biology. Ecol. 2015, 14, 57–64. [Google Scholar]
- Fedorova, L.I.; Kaygorodova, I.A. The present state of the leech fauna (Annelida, Hirudinea) in the Upper Irtysh cascade of water reservoirs. ZooKeys 2016, 596, 1–12. [Google Scholar] [CrossRef] [Green Version]
- Kaygorodova, I.A.; Fedorova, L.I. The first data on species diversity of leeches (Hirudinea) the Irtysh River Basin, East Kazakhstan. Zootaxa 2016, 4144, 287–290. [Google Scholar] [CrossRef] [PubMed]
- Fedorova, L.I.; Kaygorodova, I.A.; Bukin, Y.S. Ecological and Faunistic Characteristics of the Leech Fauna in the Bukhtarma Reservoir, Eastern Kazakhstan. Rus. J. Ecol. 2017, 48, 170–177. [Google Scholar] [CrossRef]
- Kaygorodova, I.A.; Dzyuba, E.V.; Sorokovikova, N.V. First records of potamic leech fauna of Eastern Siberia. Dataset Pap. Sci. 2013, 2013, 362683. [Google Scholar] [CrossRef] [Green Version]
- Kaygorodova, I. A revised checklist of the Lake Baikal Hirudinida fauna. Lauterbornia 2012, 75, 49–62. [Google Scholar]
- Kaygorodova, I. An Illustrated Checklist of Leech Species from Lake Baikal (Eastern Siberia, Russia). Dataset Pap. Biol. 2013, 2013, 261521. [Google Scholar] [CrossRef] [Green Version]
- Kaygorodova, I.A.; Pronin, N.M. New Records of Lake Baikal Leech Fauna: Species Diversity and Spatial Distribution in Chivyrkuy Gulf. Sci. World J. 2013, 2013, 206590. [Google Scholar] [CrossRef] [Green Version]
- Kaygorodova, I.A. Annotated checklist of the leech species diversity in the Maloe More Strait of Lake Baikal, Russia. ZooKeys 2015, 545, 37–52. [Google Scholar] [CrossRef]
- Samojlova, G. Eastern Siberia. In Great Russian Encyclopedia; Osipov, Y.S., Ed.; BRE: Moscow, Russia, 2016; Volume 5, pp. 752–753. [Google Scholar]
- Bielecki, A. Piscicola pojmanskae, a new leech species from Poland (Hirudinea: Piscicolidae). Genus. Int. J. Inverteb. Taxon. 1994, 5, 411–421. [Google Scholar]
- Hebert, P.; Cywinska, A.; Ball, S.; de Waard, J. Biological identifications through DNA barcodes. Proc. R. Soc. B 2003, 70, 313–321. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hebert, P.; Gregory, T. The promise of DNA barcoding for taxonomy. Syst. Biol. 2005, 54, 852–859. [Google Scholar] [CrossRef] [PubMed]
- Hebert, P.; Hollingsworth, P.; Hajibabaei, M. From writing to reading the encyclopedia of life. Phil. Trans. R. Soc. B. 2016, 371, 20150321. [Google Scholar] [CrossRef] [Green Version]
- Folmer, O.; Black, M.; Hoeh, W.; Lutz, R.; Vrijenhoek, R. DNA primers for amplification of mitochondrial cytochrome c oxidase subunit I from diverse metazoan invertebrates. Mol. Mar. Biol. Biotechnol. 1994, 3, 294–299. [Google Scholar]
- Tamura, K.; Stecher, G.; Kumar, S. MEGA11: Molecular Evolutionary Genetics Analysis version 11. Mol. Biol. Evol. 2021, 38, 3022–3027. [Google Scholar] [CrossRef]
- Camacho, C.; Coulouris, G.; Avagyan, V.; Ma, N.; Papadopoulos, J.; Bealer, K.; Madden, T.L. BLAST+: Architecture and applications. BMC Bioinform. 2009, 10, 421. [Google Scholar] [CrossRef]
- Fujisawa, T.; Barraclough, T.G. Delimiting Species Using Single-Locus Data and the Generalized Mixed Yule Coalescent Approach: A Revised Method and Evaluation on Simulated Data Sets. Syst. Biol. 2013, 62, 707–724. [Google Scholar] [CrossRef] [Green Version]
- Bouckaert, R.; Vaughan, T.; Barido-Sottani, J.; Duchêne, S.; Fourment, M.; Gavryushkina, S.; Heled, J.; Jones, G.; Kühnert, D.; De Maio, N.; et al. BEAST 2.5: An advanced software platform for Bayesian evolutionary analysis. PLoS Comput. Biol. 2019, 15, e1006650. [Google Scholar] [CrossRef] [Green Version]
- Darriba, D.; Taboada, G.L.; Doallo, R.; Posada, D. jModelTest 2: More models, new heuristics and parallel computing. Nat. Methods 2012, 9, 772. [Google Scholar] [CrossRef] [Green Version]
- Drummond, A.; Ho, S.; Phillips, M.; Rambaut, A. Relaxed phylogenetics and dating with confidence. PLoS ONE 2006, 4, E88. [Google Scholar] [CrossRef] [PubMed]
- Rambaut, A.; Drummond, A.; Xie, D.; Baele, G.; Suchard, M. Posterior summarisation in Bayesian phylogenetics using Tracer 1.7. Syst. Biol. 2018, 67, 901–904. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Molecular Evolution, Phylogenetics and Epidemiology: FigTree. Available online: http://tree.bio.ed.ac.uk/software/figtree/ (accessed on 11 November 2022).
- Sawyer, R.T. Leech Biology and Behaviour; Clarendon Press: Oxford, UK, 1986; Volume 2, pp. 646–706. [Google Scholar]
- Bely, A.; Weisblat, D. Lessons from leeches: A call for DNA barcoding in the lab. Evol. Dev. 2006, 8, 491–501. [Google Scholar] [CrossRef] [PubMed]
- Kutschera, U. The golden gate leech Helobdella californica (Hirudinea: Glossiphoniidae): Occurrence and DNA-based taxonomy of a species restricted to San Francisco. Int. Rev. Hydrobiol. 2011, 96, 286–295. [Google Scholar] [CrossRef]
- Nakano, T. A new species of Orobdella (Hirudinida, Arhynchobdellida, Orobdellidae) and rediscription of O. kawakatsuorum from Hokkaido, Japan with the phylogenetic position of the new species. ZooKeys 2012, 169, 9–30. [Google Scholar] [CrossRef] [PubMed]
- Reyes-Prieto, M.; Oceguera-Figuera, A.; Snell, S.; Negredo, A.; Barba, E.; Fernandez, L.; Moya, A.; Latorre, A. DNA barcodes reveal the presence of the introduced freshwater leech Helobdella europaea in Spain. Mitochondrial DNA 2014, 25, 387–395. [Google Scholar] [CrossRef]
- Kaygorodova, I.; Bolbat, N.; Bolbat, A. Species delimitation through DNA barcoding of freshwater leeches of the Glossiphonia genus (Hirudinea: Glossiphoniidae) from Eastern Siberia, Russia. J. Zool. Syst. Evol. 2020, 58, 1437–1446. [Google Scholar] [CrossRef]
- Kaygorodova, I.; Matveenko, E.; Dzyuba, E. Unexpected discovery of an ectoparasitic invasion first detected in the Baikal coregonid fish population. Fishes 2022, 7, 298. [Google Scholar] [CrossRef]
- Pons, J.; Barraclough, T.; Gomez-Zurita, J.; Cardoso, A.; Duran, D.; Hazell, S.; Kamoun, S.; Sumlin, W.; Vogler, A. Sequence-based species delimitation for the DNA taxonomy of undescribed insects. Syst. Biol. 2006, 55, 595–609. [Google Scholar] [CrossRef] [Green Version]
- Kapli, P.; Lutteropp, S.; Zhang, J.; Kobert, K.; Pavlidis, P.; Stamatakis, A.; Flouri, T. Multi-rate Poisson tree processes for single-locus species delimitation under maximum likelihood and Markov chain Monte Carlo. Bioinformatics 2017, 33, 1630–1638. [Google Scholar] [CrossRef] [Green Version]
- Puillandre, N.; Brouillet, S.; Achaz, G. ASAP: Assemble species by automatic partitioning. Mol. Ecol. Resour. 2021, 21, 609–620. [Google Scholar] [CrossRef] [PubMed]
- Kendzierska, H.; Dąbrowska, A.; Cichocka, J.; Janas, U.; Bielecki, A. The first record of Piscicola pojmanskae Bielecki, 1994 in the Gulf of Gdańsk (the southern Baltic Sea) with species characteristics distinguishing it from Piscicola geometra (Linnaeus, 1758). Oceanol. Hydrobiol. Stud. 2014, 43, 324–327. [Google Scholar] [CrossRef]
- Mann, K.H. Leeches (Hirudinea): Their Structure, Physiology, Ecology, and Embryology; Pergamon Press: New York, NY, USA, 1962; p. 161. [Google Scholar]
- Koperski, P. Taxonomic, phylogenetic and functional diversity of leeches (Hirudinea) and their suitability in biological assessment of environmental quality. Knowl. Manag. Aquat. Ecosyst. 2017, 418, 49. [Google Scholar] [CrossRef] [Green Version]
- Bogolepov, K.; Votakh, O. Precambrian Continents: Ancient Platforms of Eurasia; Nauka: Novosibirsk, USSR, 1977; 312p. [Google Scholar]
- Ernst, R.; Hamilton, M.; Söderlund, U.; Hanes, J.A.; Gladkochub, D.P.; Okrugin, A.V.; Kolotilina, T.; Mekhonoshin, A.S.; Bleeker, W.; LeCheminant, A.N.; et al. Long-lived connection between southern Siberia and northern Laurentia in the Proterozoic. Nat. Geosci. 2016, 9, 464–469. [Google Scholar] [CrossRef]
- Marincovich, L.; Gladenkov, A. Evidence for an early opening of the Bering Strait. Nature 1999, 397, 149–151. [Google Scholar] [CrossRef]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Species | Voucher | Locality 1 | GenBank Accession |
|---|---|---|---|
| Piscicola geometra (L., 1761) | C218-220 | EU: Temernik River, RR, Russia; 47.276107, 39.718591 | OQ152291-3 |
| KM3-11 | 8 | ||
| C221 | EU: Temernik River, RR, Russia; 47.286388, 39.723242 | OQ152294 | |
| F88 | WS: Bukhtarma reservoir, Kazakhstan; 49.616111, 83.525833 | OQ152290 | |
| L1-L5 | 5 | ||
| L6-L7 | WS: Bukhtarma reservoir, Kazakhstan; 49.616944, 83.571667 | 2 | |
| L8-L11 | WS: Bukhtarma reservoir, Kazakhstan; 49.627500, 83.455833 | 4 | |
| Piscicola nordica sp. nov. | C169 | EU: Lake Golovka, RK, Russia; 67.493504, 64.050113 | OQ152314 |
| Piscicola sibirica sp. nov. | C2 | ES: Angara River, IR, Russia; 52.432133, 104.109404 | OQ152295 |
| B39-40 | ES: Angara River, IR, Russia; 52.535392, 103.964607 | OQ152304-5 | |
| B38, A1-22 | 23 | ||
| BS76-77 | ES: Bratsk reservoir, IR, Russia; 53.095889, 103.356420 | OQ152296-7 | |
| BS31-33 | 3 | ||
| M27-29 | ES: Irkutsk reservoir, IR, Russia; 51.998870, 104.664175 | OQ152301-3 | |
| BR1-8 | ES: Irkutsk reservoir, IR, Russia; 51.962708, 104.713775 | 8 | |
| BR10-20 | ES: Irkutsk reservoir, IR, Russia; 52.118985, 104.481527 | 11 | |
| C175 | Olkha River, IR, Russia; 52.138078, 104.073423 | 1 | |
| M1 | Irkut River, IR, Russia; 51.919409, 103.697399 | 1 | |
| C222 | Irkut River, IR, Russia; 52.280077, 104.209310 | 1 | |
| C140 | Belaya River, IR, Russia; 52.866246, 103.310471 | 1 | |
| G80 | ES: Lake Gusinoe, RB, Russia; 51.116221, 106.363094 | KM095104 | |
| KM77-78 C137 | 3 | ||
| G81 | ES: Lake Tsaidam, RB, Russia; 50.564094, 105.788088 | KM095103 | |
| K65-67 | ES: Ina River, RB, Russia; 53.654650, 109.840142 | OQ152298-300 | |
| K79-80 | ES: Lake Baikal, RB, Russia; 51.953118, 106.127163 | OQ152306-7 | |
| K81-K82 | 2 | ||
| P1-P7 | ES: Lake Baikal, RB, Russia; 53.745792, 109.110156 | 7 | |
| S1-S4 | ES: Yana River, RS, Russia; 67.553945, 133.367242 | 4 | |
| Piscicola khubsugulensis sp. nov. | K86-87 | ES: Lake Khovsgol, Mongolia; 51.080021, 100.477723 | OQ152308-9 |
| C183-184 | OQ152310-1 | ||
| K18-19 | ES: Lake Khovsgol, Mongolia; 50.441656, 100.181546 | OQ152312-3 | |
| X494-496 | 4 | ||
| Cystobranchus mammilatus (Malm, 1863) 2 | B44 | EU: Pechora River, RK, Russia; 65.427151, 52.134787 | OQ152315 |
| KM1-2 | 2 |
| Description | Max Score | Query Cover | Identity | Length, bp | Accession |
|---|---|---|---|---|---|
| Piscicola milneri | 1120 | 100% | 97.84% | 665 | DQ414337 |
| Piscicola geometra | 976 | 100% | 93.83% | 15135 | OX030972 |
| Piscicola geometra | 976 | 100% | 93.83% | 14788 | BK059172 |
| Piscicola geometra | 976 | 100% | 93.83% | 651 | AF003280 |
| Piscicola pojmanskae | 924 | 95% | 93.68% | 617 | MH395321 |
| Piscicola pojmanskae | 924 | 95% | 93.68% | 617 | MH395320 |
| Piscicola geometra | 939 | 96% | 93.63% | 634 | MF458791 |
| Piscicola annae | 922 | 95% | 93.53% | 629 | AY336016 |
| Piscicola geometra | 920 | 95% | 93.53% | 631 | AY336014 |
| Piscicola geometra | 913 | 95% | 93.35% | 628 | AY336015 |
| No. | Species Group | P | SE | 1 | 2 | 3 | 4 | 5 | 6 | 7 | 8 | 9 |
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 1 | P. annae | n/c | n/c | 0.0083 | 0.0092 | 0.0085 | 0.0105 | 0.0098 | 0.0094 | 0.0070 | 0.0081 | |
| 2 | P. geometra | 0.0043 | 0.0014 | 0.0491 | 0.0090 | 0.0022 | 0.0104 | 0.0094 | 0.0090 | 0.0089 | 0.0072 | |
| 3 | P. milneri | 0.0015 | 0.0015 | 0.0598 | 0.0577 | 0.0093 | 0.0104 | 0.0048 | 0.0054 | 0.0068 | 0.0086 | |
| 4 | P. pojmanskae | 0 | 0 | 0.0487 | 0.0047 | 0.0559 | 0.0104 | 0.0096 | 0.0046 | 0.0093 | 0.0078 | |
| 5 | P. punctata | n/c | n/c | 0.0734 | 0.0754 | 0.0677 | 0.0697 | 0.0112 | 0.0104 | 0.0107 | 0.0107 | |
| 6 | Piscicola sp. | n/c | n/c | 0.0637 | 0.0594 | 0.0152 | 0.0570 | 0.0734 | 0.0058 | 0.0067 | 0.0093 | |
| 7 | P. sibirica * | 0.0102 | 0.0020 | 0.0653 | 0.0640 | 0.0253 | 0.0628 | 0.0749 | 0.0259 | 0.0074 | 0.0090 | |
| 8 | P. khubsugulensis * | 0.0022 | 0.0013 | 0.0615 | 0.0565 | 0.0308 | 0.0556 | 0.0726 | 0.0265 | 0.0392 | 0.0090 | |
| 9 | P. nordica * | n/c | n/c | 0.0447 | 0.0386 | 0.0546 | 0.0389 | 0.0738 | 0.0581 | 0.0607 | 0.0565 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kaygorodova, I.; Matveenko, E. Diversity of the Piscicola Species (Hirudinea, Piscicolidae) in the Eastern Palaearctic with a Description of Three New Species and Notes on Their Biogeography. Diversity 2023, 15, 98. https://doi.org/10.3390/d15010098
Kaygorodova I, Matveenko E. Diversity of the Piscicola Species (Hirudinea, Piscicolidae) in the Eastern Palaearctic with a Description of Three New Species and Notes on Their Biogeography. Diversity. 2023; 15(1):98. https://doi.org/10.3390/d15010098
Chicago/Turabian StyleKaygorodova, Irina, and Ekaterina Matveenko. 2023. "Diversity of the Piscicola Species (Hirudinea, Piscicolidae) in the Eastern Palaearctic with a Description of Three New Species and Notes on Their Biogeography" Diversity 15, no. 1: 98. https://doi.org/10.3390/d15010098