


Marine Noise Effects on Juvenile Sparid Fish Change among Species and Developmental Stages
 , , , , , , ,
, , , , , , ,
Abstract
:1. Introduction
2. Materials and Methods
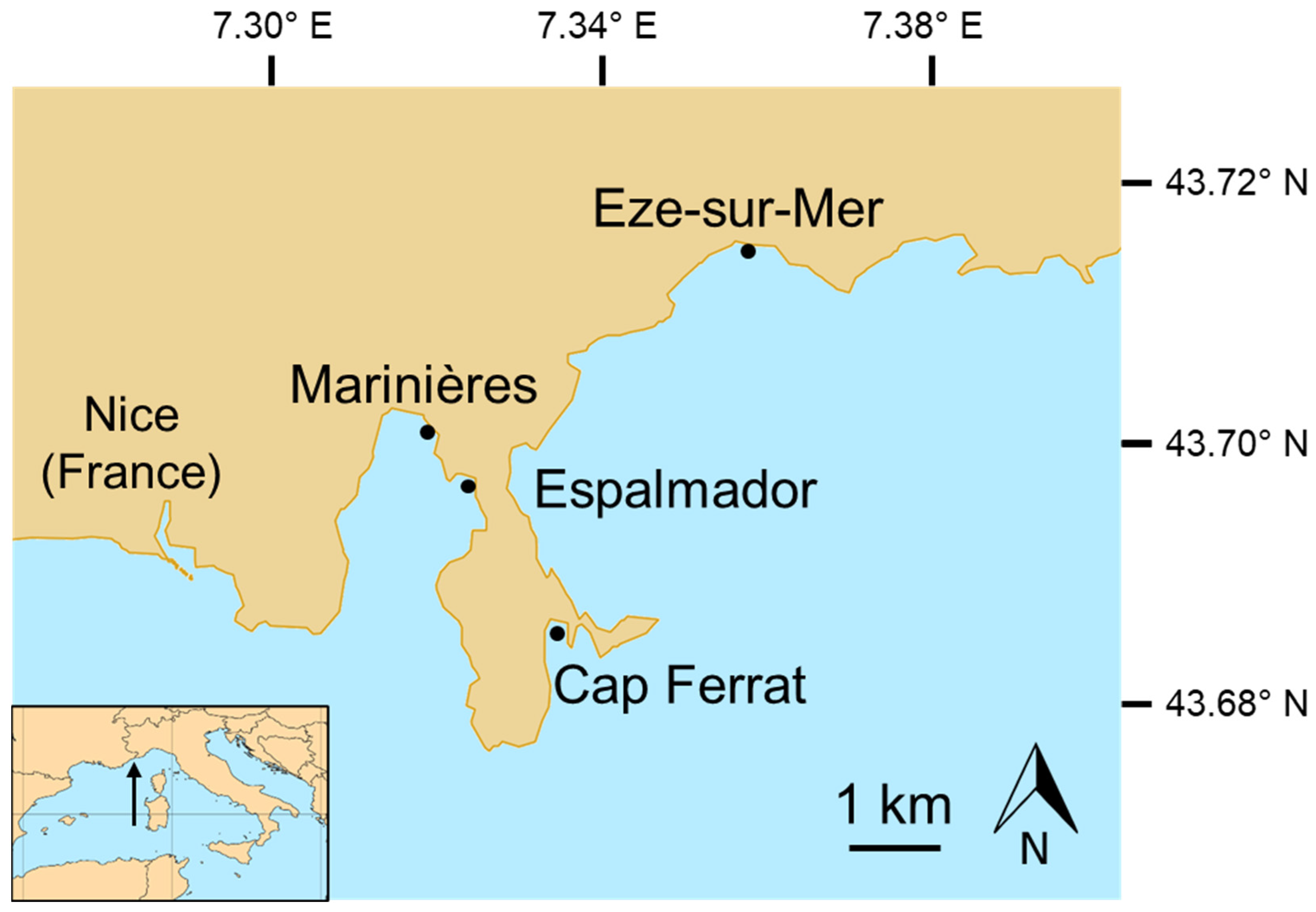
2.1. Study Area
2.2. Noise Measures to Characterize the Sampling Locations
2.3. Assessment of Juvenile Fish Assemblages
2.4. Statistical Data Analyses
3. Results
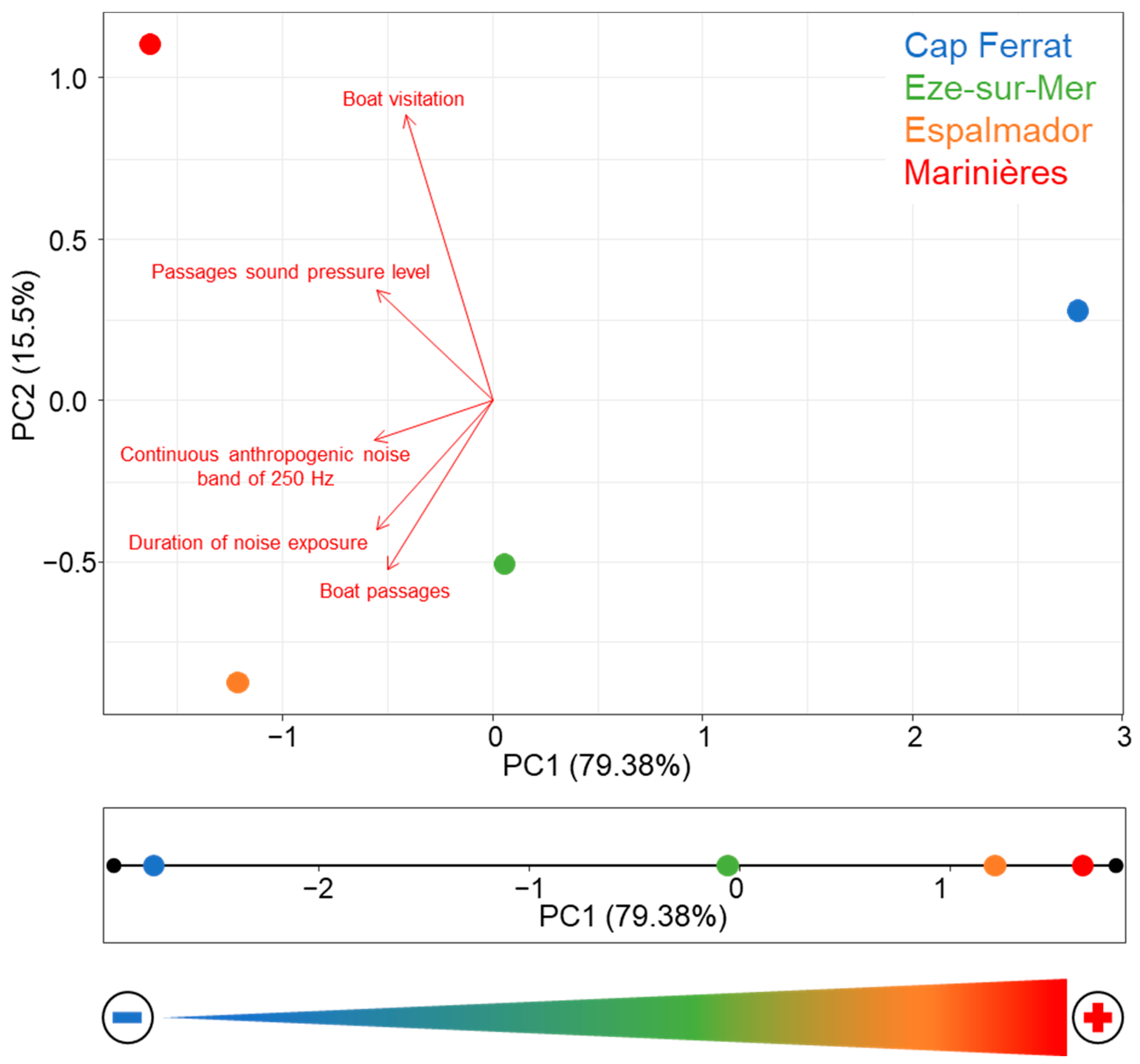
3.1. Noise Measures
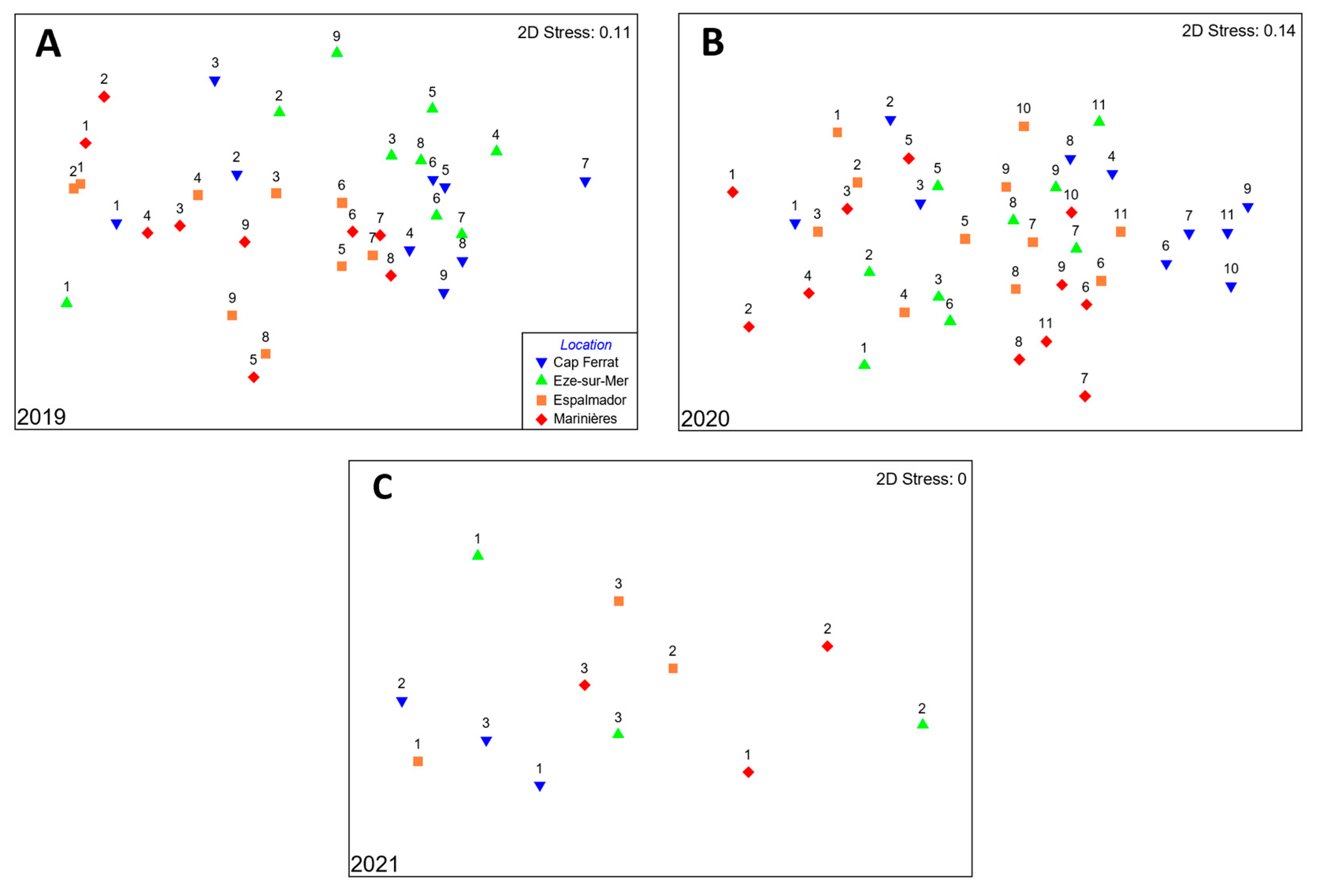
3.2. Juvenile Sparid Assemblage Structure
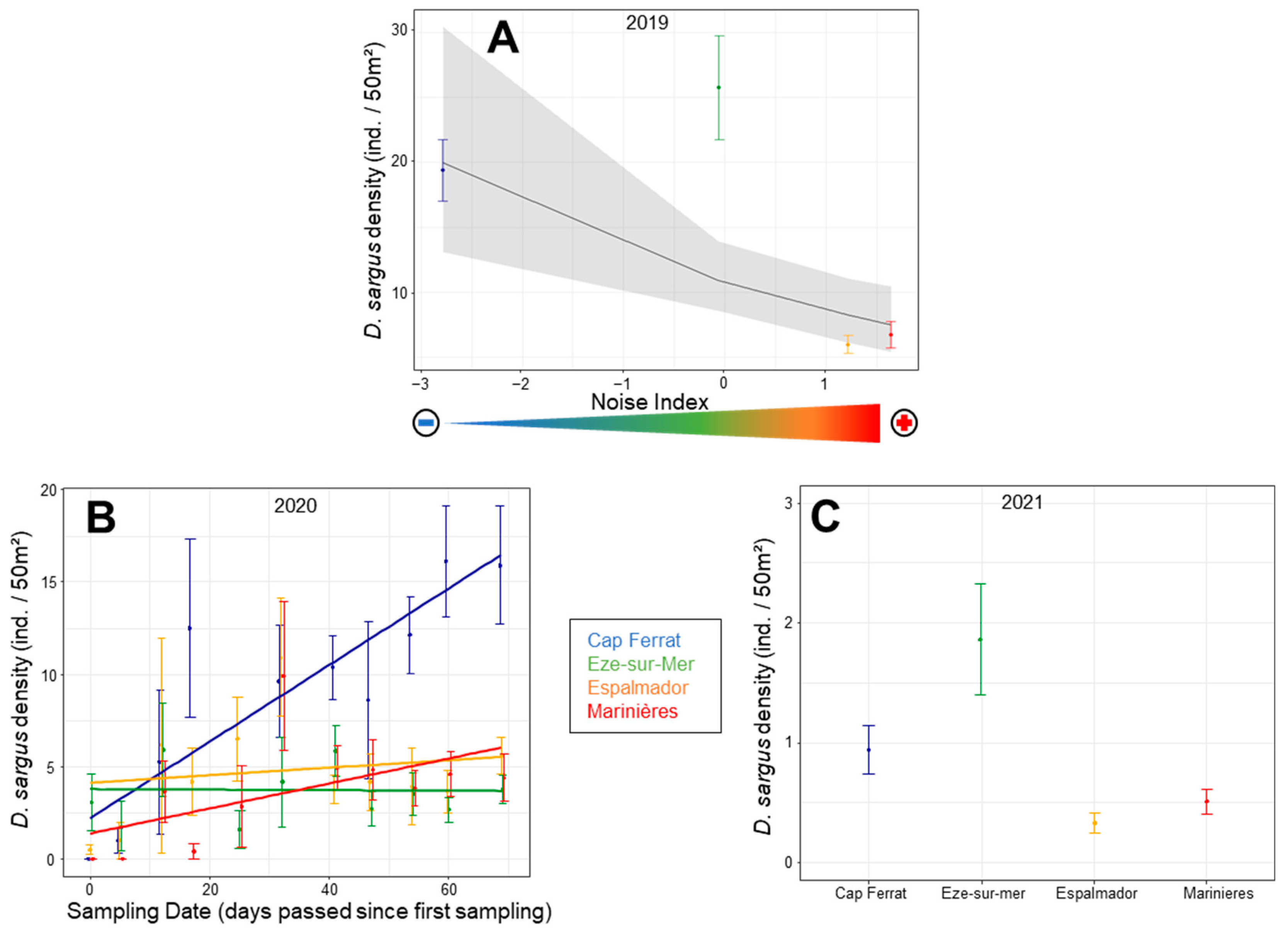
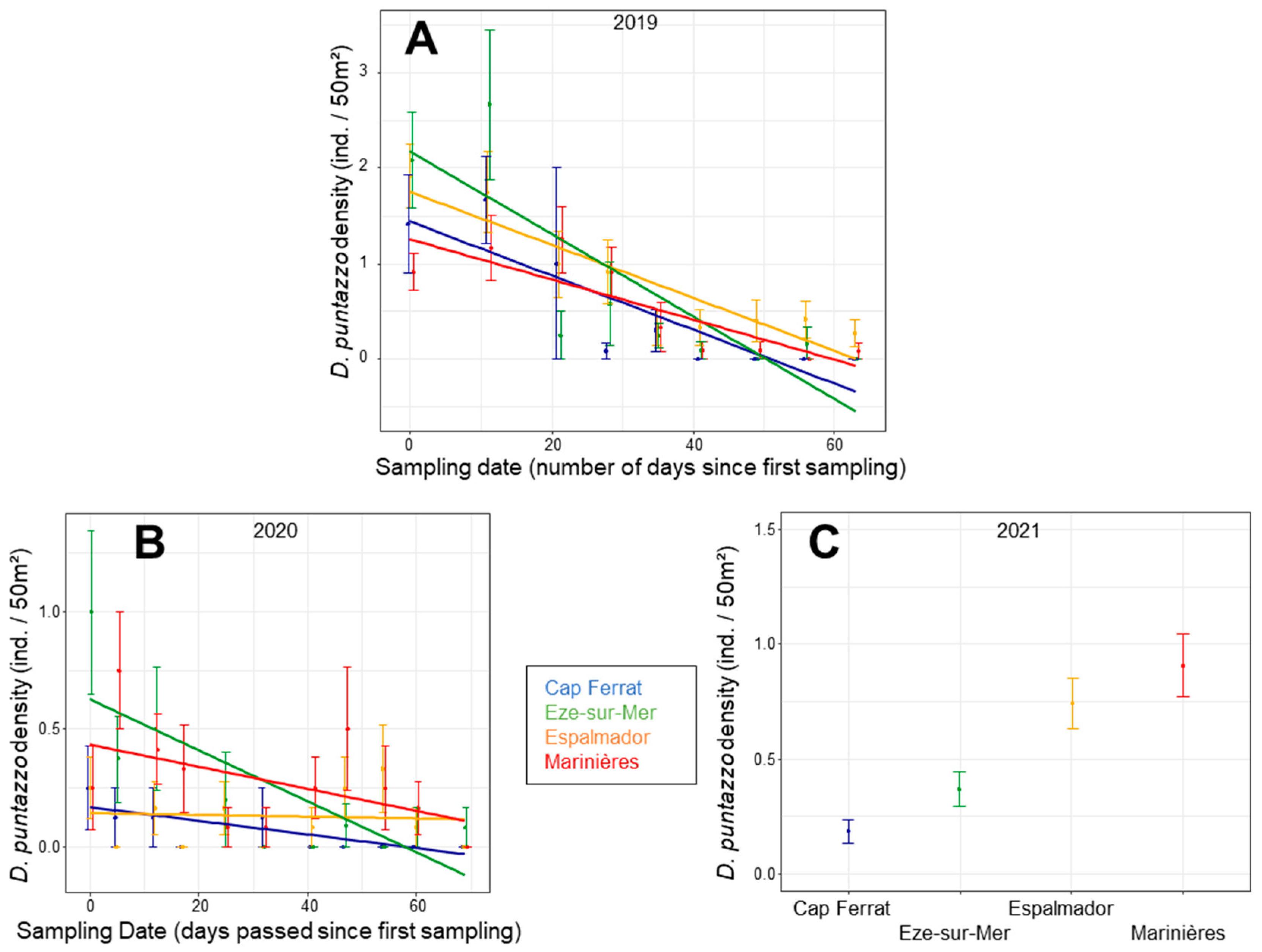
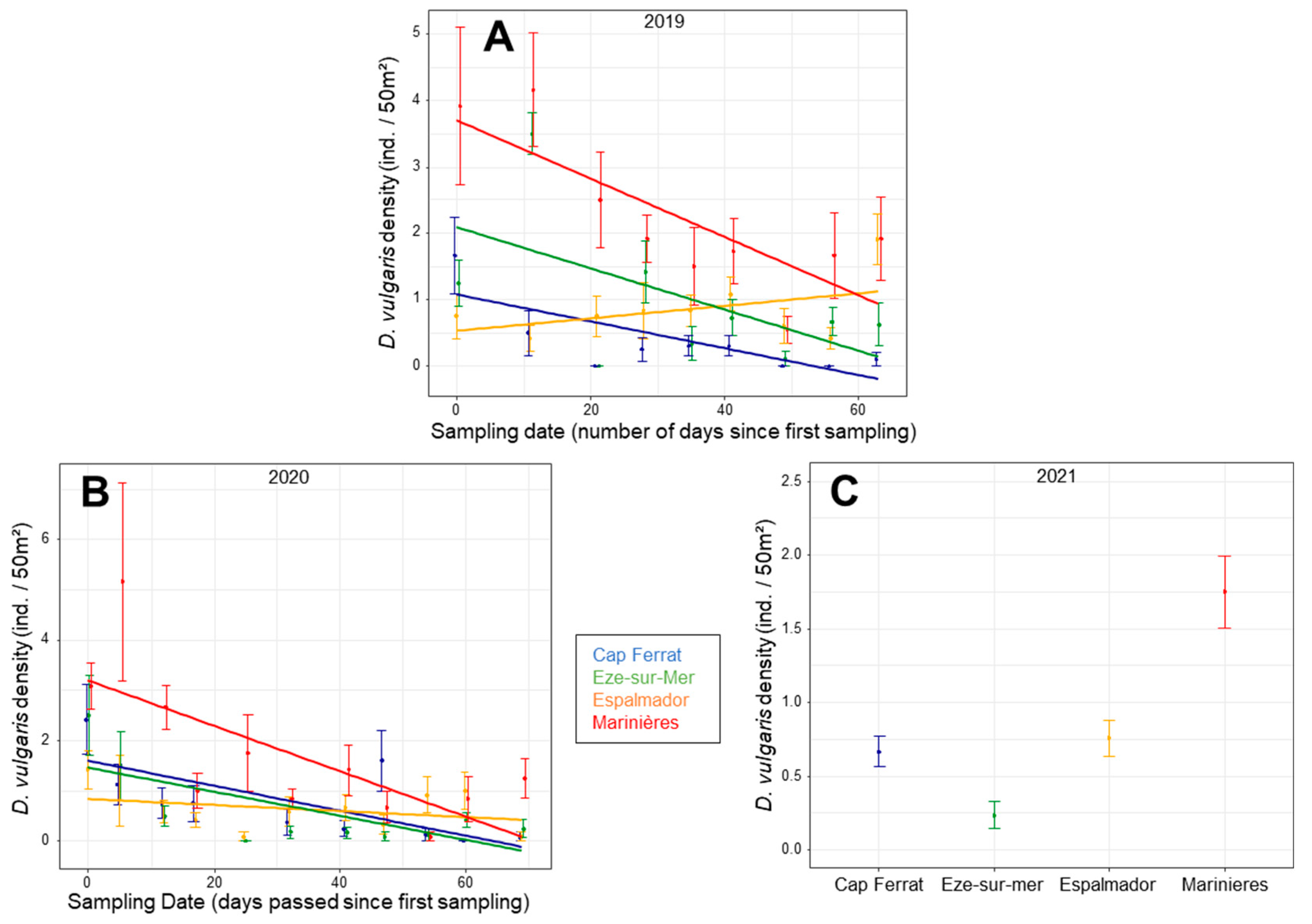
3.3. Single Species Density Patterns
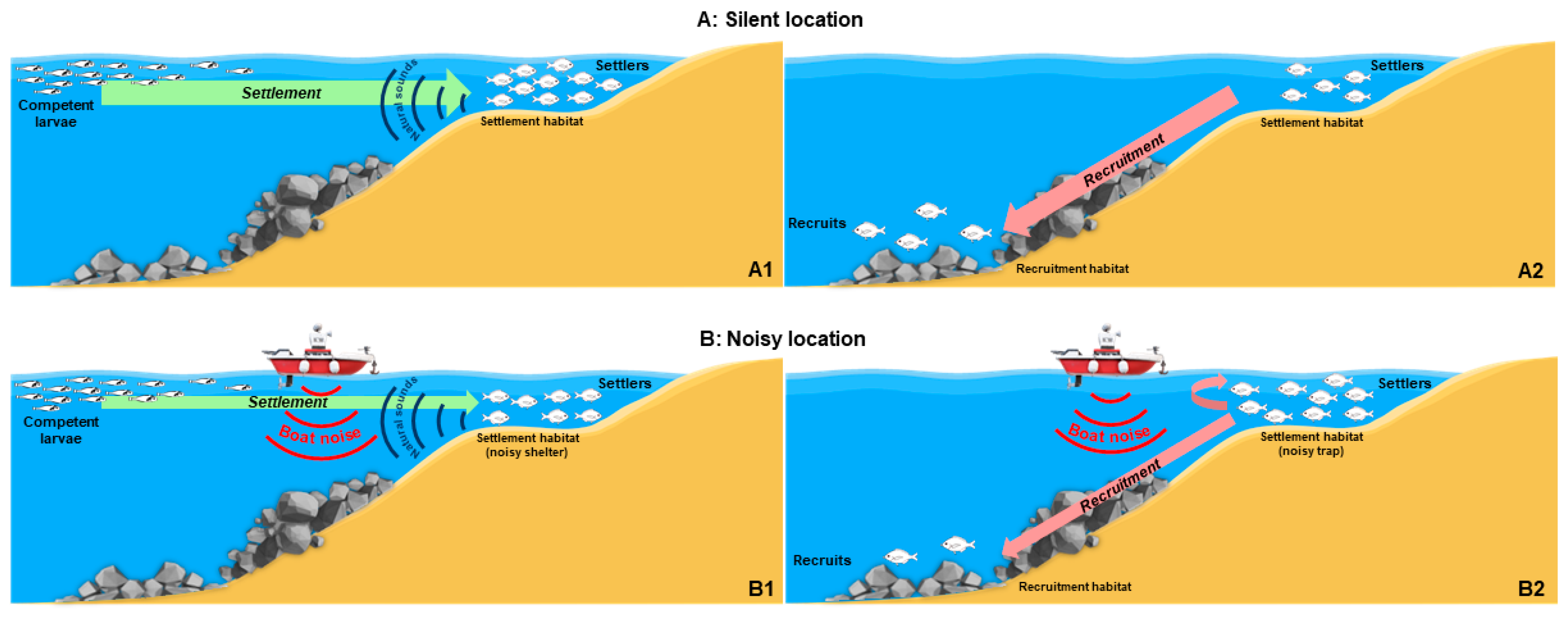
4. Discussion and Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Pijanowski, B.C.; Villanueva-Rivera, L.J.; Dumyahn, S.L.; Farina, A.; Krause, B.L.; Napoletano, B.M.; Gage, S.H.; Pieretti, N. Soundscape Ecology: The Science of Sound in the Landscape. BioScience 2011, 61, 203–216. [Google Scholar] [CrossRef] [Green Version]
- Sueur, J.; Farina, A. Ecoacoustics: The Ecological Investigation and Interpretation of Environmental Sound. Biosemiotics 2015, 8, 493–502. [Google Scholar] [CrossRef]
- Mooney, T.A.; Di Iorio, L.; Lammers, M.; Lin, T.-H.; Nedelec, S.L.; Parsons, M.; Radford, C.; Urban, E.; Stanley, J. Listening Forward: Approaching Marine Biodiversity Assessments Using Acoustic Methods. R. Soc. Open Sci. 2020, 7, 201287. [Google Scholar] [CrossRef] [PubMed]
- Tolimieri, N.; Jeffs, A.; Montgomery, J. Ambient Sound as a Cue for Navigation by the Pelagic Larvae of Reef Fishes. Mar. Ecol. Prog. Ser. 2000, 207, 219–224. [Google Scholar] [CrossRef]
- Tyack, P.L.; Clark, C.W. Communication and Acoustic Behavior of Dolphins and Whales. In Hearing by Whales and Dolphins; Au, W.W.L., Fay, R.R., Popper, A.N., Eds.; Springer Handbook of Auditory Research; Springer: New York, NY, USA, 2000; Volume 12, pp. 156–224. ISBN 978-1-4612-7024-9. [Google Scholar]
- Stanley, J.A.; Radford, C.A.; Jeffs, A.G. Location, Location, Location: Finding a Suitable Home among the Noise. Proc. R. Soc. B Biol. Sci. 2012, 279, 3622–3631. [Google Scholar] [CrossRef]
- Ladich, F. Sound Communication in Fishes, 1st ed.; Animal Signals and Communication; Springer: Vienna, Austria, 2015; ISBN 978-3-7091-1846-7. [Google Scholar]
- Popper, A.N.; Hawkins, A. The Effects of Noise on Aquatic Life II; Advances in Experimental Medicine and Biology; Springer: New York, NY, USA, 2016; Volume 875, ISBN 978-1-4939-2980-1. [Google Scholar]
- Cox, K.; Brennan, L.P.; Gerwing, T.G.; Dudas, S.E.; Juanes, F. Sound the Alarm: A Meta-Analysis on the Effect of Aquatic Noise on Fish Behavior and Physiology. Global Chang. Biol. 2018, 24, 3105–3116. [Google Scholar] [CrossRef] [PubMed]
- Di Franco, E.; Pierson, P.; Di Iorio, L.; Calò, A.; Cottalorda, J.M.; Derijard, B.; Di Franco, A.; Galvé, A.; Guibbolini, M.; Lebrun, J.; et al. Effects of Marine Noise Pollution on Mediterranean Fishes and Invertebrates: A Review. Mar. Pollut. Bull. 2020, 159, 111450. [Google Scholar] [CrossRef]
- Solé, M.; Lenoir, M.; Fontuño, J.M.; Durfort, M.; van der Schaar, M.; André, M. Evidence of Cnidarians Sensitivity to Sound after Exposure to Low Frequency Noise Underwater Sources. Sci. Rep. 2016, 6, 37979. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Weilgart, L. The Impact of Ocean Noise Pollution on Fish and Invertebrates. Doctoral Dissertation, Oceancare & Dalhousie University, Dalhousie, NS, Camada, 2018. Volume 34. [Google Scholar]
- Duarte, C.M.; Chapuis, L.; Collin, S.P.; Costa, D.P.; Devassy, R.P.; Eguiluz, V.M.; Erbe, C.; Gordon, T.A.C.; Halpern, B.S.; Harding, H.R.; et al. The Soundscape of the Anthropocene Ocean. Science 2021, 371, eaba4658. [Google Scholar] [CrossRef]
- Levin, P.S. Fine-Scale Temporal Variation in Recruitment of a Temperate Demersal Fish: The Importance of Settlement versus Post-Settlement Loss. Oecologia 1994, 97, 124–133. [Google Scholar] [CrossRef]
- Macpherson, E. Ontogenetic Shifts in Habitat Use and Aggregation in Juvenile Sparid Fishes. J. Exp. Mar. Biol. Ecol. 1998, 220, 127–150. [Google Scholar] [CrossRef]
- Leis, J.M.; McCormick, M.I. The Biology, Behavior, and Ecology of the Pelagic, Larval Stage of Coral Reef Fishes. In Coral Reef Fishes; Elsevier: Amsterdam, The Netherlands, 2002; pp. 171–199. ISBN 978-0-12-615185-5. [Google Scholar]
- Bussotti, S.; Guidetti, P. Timing and Habitat Preferences for Settlement of Juvenile Fishes in the Marine Protected Area of Torre Guaceto (South-Eastern Italy, Adriatic Sea). Ital. J. Zool. 2011, 78, 243–254. [Google Scholar] [CrossRef]
- Simpson, S.D.; Radford, A.N.; Tickle, E.J.; Meekan, M.G.; Jeffs, A.G. Adaptive Avoidance of Reef Noise. PLoS ONE 2011, 6, e16625. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Simpson, S.D.; Radford, A.N.; Holles, S.; Ferarri, M.C.O.; Chivers, D.P.; McCormick, M.I.; Meekan, M.G. Small-Boat Noise Impacts Natural Settlement Behavior of Coral Reef Fish Larvae. In The Effects of Noise on Aquatic Life II; Popper, A.N., Hawkins, A., Eds.; Springer: New York, NY, USA, 2016; Volume 875, pp. 1041–1048. ISBN 978-1-4939-2980-1. [Google Scholar]
- Holles, S.; Simpson, S.; Radford, A.; Berten, L.; Lecchini, D. Boat Noise Disrupts Orientation Behaviour in a Coral Reef Fish. Mar. Ecol. Prog. Ser. 2013, 485, 295–300. [Google Scholar] [CrossRef] [Green Version]
- Simpson, S.; Meekan, M.; McCauley, R.; Jeffs, A. Attraction of Settlement-Stage Coral Reef Fishes to Reef Noise. Mar. Ecol. Prog. Ser. 2004, 276, 263–268. [Google Scholar] [CrossRef] [Green Version]
- Kingsford, M.J.; Leis, J.M.; Shanks, A.; Lindeman, K.C.; Morgan, S.G.; Pineda, J. Sensory Environments, Larval Abilities and Local Self-Recruitment. Bull. Mar. Sci. 2002, 70, 309–340. [Google Scholar]
- Gordon, T.A.C.; Harding, H.R.; Wong, K.E.; Merchant, N.D.; Meekan, M.G.; McCormick, M.I.; Radford, A.N.; Simpson, S.D. Habitat Degradation Negatively Affects Auditory Settlement Behavior of Coral Reef Fishes. Proc. Natl. Acad. Sci. USA 2018, 115, 5193–5198. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Beets, J. Effects of a Predatory Fish on the Recruitment and Abundance of Caribbean Coral Reef Fishes. Mar. Ecol. Prog. Ser. 1997, 148, 11–21. [Google Scholar] [CrossRef]
- Fontes, J.; Caselle, J.E.; Afonso, P.; Santos, R.S. Multi-Scale Recruitment Patterns and Effects on Local Population Size of a Temperate Reef Fish. J. Fish Biol. 2009, 75, 1271–1286. [Google Scholar] [CrossRef]
- Gordon, T.A.C.; Radford, A.N.; Davidson, I.K.; Barnes, K.; McCloskey, K.; Nedelec, S.L.; Meekan, M.G.; McCormick, M.I.; Simpson, S.D. Acoustic Enrichment Can Enhance Fish Community Development on Degraded Coral Reef Habitat. Nat. Commun. 2019, 10, 5414. [Google Scholar] [CrossRef] [Green Version]
- Airoldi, L.; Beck, M.W.; Firth, L.B.; Bugnot, A.B.; Steinberg, P.D.; Dafforn, K.A. Emerging Solutions to Return Nature to the Urban Ocean. Annu. Rev. Mar. Sci. 2021, 13, 445–477. [Google Scholar] [CrossRef]
- Bevilacqua, S.; Airoldi, L.; Ballesteros, E.; Benedetti-Cecchi, L.; Boero, F.; Bulleri, F.; Cebrian, E.; Cerrano, C.; Claudet, J.; Colloca, F.; et al. Mediterranean Rocky Reefs in the Anthropocene: Present Status and Future Concerns. In Advances in Marine Biology; Elsevier: Amsterdam, The Netherlands, 2021; Volume 89, pp. 1–51. ISBN 978-0-12-824623-8. [Google Scholar]
- Coll, M.; Piroddi, C.; Albouy, C.; Ben Rais Lasram, F.; Cheung, W.W.L.; Christensen, V.; Karpouzi, V.S.; Guilhaumon, F.; Mouillot, D.; Paleczny, M.; et al. The Mediterranean Sea under Siege: Spatial Overlap between Marine Biodiversity, Cumulative Threats and Marine Reserves: The Mediterranean Sea under Siege. Glob. Ecol. Biogeogr. 2012, 21, 465–480. [Google Scholar] [CrossRef]
- Micheli, F.; Halpern, B.S.; Walbridge, S.; Ciriaco, S.; Ferretti, F.; Fraschetti, S.; Lewison, R.; Nykjaer, L.; Rosenberg, A.A. Cumulative Human Impacts on Mediterranean and Black Sea Marine Ecosystems: Assessing Current Pressures and Opportunities. PLoS ONE 2013, 8, e79889. [Google Scholar] [CrossRef] [Green Version]
- Froese, R.; Pauly, D. FishBase 2021. Available online: https://www.fishbase.org (accessed on 1 January 2023).
- Vigliola, L.; Harmelin-Vivien, M. Post-Settlement Ontogeny in Three Mediterranean Reef Fish Species of the Genus Diplodus. Bull. Mar. Sci. 2001, 68, 16. [Google Scholar]
- Di Franco, A.; Calò, A.; Pennetta, A.; De Benedetto, G.; Planes, S.; Guidetti, P. Dispersal of Larval and Juvenile Seabream: Implications for Mediterranean Marine Protected Areas. Biol. Conserv. 2015, 192, 361–368. [Google Scholar] [CrossRef]
- Isnard, E.; Tournois, J.; McKenzie, D.J.; Ferraton, F.; Bodin, N.; Aliaume, C.; Darnaude, A.M. Getting a Good Start in Life? A Comparative Analysis of the Quality of Lagoons as Juvenile Habitats for the Gilthead Seabream Sparus aurata in the Gulf of Lions. Estuaries Coasts 2015, 38, 1937–1950. [Google Scholar] [CrossRef]
- Vandenbussche, P.S.P.; Spennato, G.; Pierson, P.M. Juvenile Oblada melanura (L. 1758) Otolith Shape Is Impacted near Recreational Harbours or Due to Settlement Position in Nearby Sites. Cont. Shelf Res. 2020, 208, 104239. [Google Scholar] [CrossRef]
- Di Franco, A.; Di Lorenzo, M.; Guidetti, P. Spatial Patterns of Density at Multiple Life Stages in Protected and Fished Conditions: An Example from a Mediterranean Coastal Fish. J. Sea Res. 2013, 76, 73–81. [Google Scholar] [CrossRef]
- Biagi, F.; Gambaccini, S.; Zazzetta, M. Settlement and Recruitment in Fishes: The Role of Coastal Areas. Ital. J. Zool. 1998, 65, 269–274. [Google Scholar] [CrossRef]
- Vigliola, L.; Harmelin-Vivien, M.; Biagi, F.; Galzin, R.; Garcia-Rubies, A.; Harmelin, J.; Jouvenel, J.; Le Direach-Boursier, L.; Macpherson, E.; Tunesi, L. Spatial and Temporal Patterns of Settlement among Sparid Fishes of the Genus Diplodus in the Northwestern Mediterranean. Mar. Ecol. Prog. Ser. 1998, 168, 45–56. [Google Scholar] [CrossRef] [Green Version]
- Di Franco, A.; Guidetti, P. Patterns of Variability in Early-Life Traits of Fishes Depend on Spatial Scale of Analysis. Biol. Lett. 2011, 7, 454–456. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Harmelin-Vivien, M.L.; Harmelin, J.G.; Leboulleux, V. Microhabitat Requirements for Settlement of Juvenile Sparid Fishes on Mediterranean Rocky Shores. Hydrobiologia 1995, 300–301, 309–320. [Google Scholar] [CrossRef]
- Meinesz, A.; Blanfuné, A.; Chancollon, O.; Javel, F.; Longepierre, S.; Markovic, L.; Vaugelas de, J.; Garcia, D. Côtes Méditerranéennes Françaises: Inventaire et Impacts des Aménagements Gagnés sur la mer; Lab. ECOMERS, Université de Nice Sophia Antipolis: Nice, France, 2013; p. 156, Et Pubblication Electronique. [Google Scholar]
- Maglio, A.; Soares, C.; Bouzidi, M.; Zabel, F.; Souami, Y.; Pavan, G. Mapping Shipping Noise in the Pelagos Sanctuary (French Part) through Acoustic Modelling to Assess Potential Impacts on Marine Mammals. Sci. Rep. Port-Cros. Natl. Park 2015, 185, 167–185. [Google Scholar]
- Ourmieres, Y.; Mansui, J.; Molcard, A.; Galgani, F.; Poitou, I. The Boundary Current Role on the Transport and Stranding of Floating Marine Litter: The French Riviera Case. Cont. Shelf Res. 2018, 155, 11–20. [Google Scholar] [CrossRef] [Green Version]
- Di Franco, A.; Gianni, F.; Guidetti, P. Mismatch in Early Life Traits between Settlers and Recruits in a Mediterranean Fish: Clue of the Relevance of the Settlement Tail? Acta Ichthyol. Et Piscat. 2015, 45, 153–159. [Google Scholar] [CrossRef] [Green Version]
- Wright, K.J.; Higgs, D.M.; Cato, D.H.; Leis, J.M. Auditory Sensitivity in Settlement-Stage Larvae of Coral Reef Fishes. Coral Reefs 2010, 29, 235–243. [Google Scholar] [CrossRef]
- Ainslie, M.A. Standard for Measurement and Monitoring of Underwater Noise, Part I; Physical Quantities and Their Units; 2011; Retrieved Online on 10 January 2023. Available online: https://www.noordzeeloket.nl/publish/pages/122320/standard_for_measurement_and_monitoring_of_underwater_noise_part_i_648.pdf (accessed on 8 January 2023).
- Colleye, O.; Kéver, L.; Lecchini, D.; Berten, L.; Parmentier, E. Auditory Evoked Potential Audiograms in Post-Settlement Stage Individuals of Coral Reef Fishes. J. Exp. Mar. Biol. Ecol. 2016, 483, 1–9. [Google Scholar] [CrossRef]
- Oksanen, J.; Simpson, G.; Blanchet, F.; Kindt, R.; Legendre, P.; Minchin, P.; O’Hara, R.; Solymos, P.; Stevens, M.; Szoecs, E.; et al. Vegan: Community Ecology Package_. R Package Version 2.6-4. 2022. Available online: https://CRAN.R-project.org/package=vegan (accessed on 8 January 2023).
- Wang, Y.; Naumann, U.; Wright, S.; Warton, D.I. Mvabund: An R Package for Model-Based Analysis of Multivariate Abundance Data. Methods Ecol. Evol. 2012, 3, 471–474. [Google Scholar] [CrossRef]
- Anderson, M.J.; Gorley, R.N.; Clarke, K.R. Permanova+ for Primer: Guide to Software and Statistical Methods. J. Oceanogr. 2008, 67, 589–599. [Google Scholar]
- Brooks, M.E.; Kristensen, K.; Benthem, K.J.; van Magnusson, A.; Berg, C.W.; Nielsen, A.; Skaug, H.J.; Mächler, M.; Bolker, B.M. GlmmTMB Balances Speed and Flexibility Among Packages for Zero-Inflated Generalized Linear Mixed Modeling. R J. 2017, 9, 378. [Google Scholar] [CrossRef] [Green Version]
- Fox, J.; Weisberg, S. An R Companion to Applied Regression, 2nd ed.; Sage Publications: Thousand Oaks, CA, USA, 2011. [Google Scholar]
- R Core Team. R: A Language and Environment for Statistical Computing; R Core Team: Vienna, Austria, 2014. [Google Scholar]
- Chahouri, A.; Elouahmani, N.; Ouchene, H. Recent Progress in Marine Noise Pollution: A Thorough Review. Chemosphere 2022, 291, 132983. [Google Scholar] [CrossRef]
- Caraka, R.E.; Yusra, Y.; Toharudin, T.; Chen, R.-C.; Basyuni, M.; Juned, V.; Gio, P.U.; Pardamean, B. Did Noise Pollution Really Improve during COVID-19? Evidence from Taiwan. Sustainability 2021, 13, 5946. [Google Scholar] [CrossRef]
- Ryan, J.P.; Joseph, J.E.; Margolina, T.; Hatch, L.T.; Azzara, A.; Reyes, A.; Southall, B.L.; DeVogelaere, A.; Peavey Reeves, L.E.; Zhang, Y.; et al. Reduction of Low-Frequency Vessel Noise in Monterey Bay National Marine Sanctuary During the COVID-19 Pandemic. Front. Mar. Sci. 2021, 8, 656566. [Google Scholar] [CrossRef]
- Bertucci, F.; Lecchini, D.; Greeven, C.; Brooker, R.M.; Minier, L.; Cordonnier, S.; René-Trouillefou, M.; Parmentier, E. Changes to an Urban Marina Soundscape Associated with COVID-19 Lockdown in Guadeloupe. Environ. Pollut. 2021, 289, 117898. [Google Scholar] [CrossRef] [PubMed]
- Karkarey, R.; Theo, A.H. Homeward Bound: Fish Larvae Use Dispersal Corridors When Settling on Coral Reefs. Front. Ecol. Environ. 2016, 14, 569–570. [Google Scholar] [CrossRef]
- Azzurro, E.; Matiddi, M.; Fanelli, E.; Guidetti, P.; Mesa, G.L.; Scarpato, A.; Axiak, V. Sewage Pollution Impact on Mediterranean Rocky-Reef Fish Assemblages. Mar. Environ. Res. 2010, 69, 390–397. [Google Scholar] [CrossRef] [Green Version]
- Lee, J.-W.; Choi, H.; Hwang, U.-K.; Kang, J.-C.; Kang, Y.J.; Kim, K.I.; Kim, J.-H. Toxic Effects of Lead Exposure on Bioaccumulation, Oxidative Stress, Neurotoxicity, and Immune Responses in Fish: A Review. Environ. Toxicol. Pharmacol. 2019, 68, 101–108. [Google Scholar] [CrossRef] [PubMed]
- Fortibuoni, T.; Giovanardi, O.; Pranovi, F.; Raicevich, S.; Solidoro, C.; Libralato, S. Analysis of Long-Term Changes in a Mediterranean Marine Ecosystem Based on Fishery Landings. Front. Mar. Sci. 2017, 4, 33. [Google Scholar] [CrossRef] [Green Version]
- Kantoussan, J.; Laë, R.; Tine, M. Review of the Fisheries Indicators for Monitoring the Impacts of Fishing on Fish Communities. Rev. Fish. Sci. Aquac. 2018, 26, 460–478. [Google Scholar] [CrossRef]
- Edgar, G.J.; Stuart-Smith, R.D.; Willis, T.J.; Kininmonth, S.; Baker, S.C.; Banks, S.; Barrett, N.S.; Becerro, M.A.; Bernard, A.T.F.; Berkhout, J.; et al. Global Conservation Outcomes Depend on Marine Protected Areas with Five Key Features. Nature 2014, 506, 216–220. [Google Scholar] [CrossRef] [Green Version]
- Giakoumi, S.; Scianna, C.; Plass-Johnson, J.; Micheli, F.; Grorud-Colvert, K.; Thiriet, P.; Claudet, J.; Di Carlo, G.; Di Franco, A.; Gaines, S.D.; et al. Ecological Effects of Full and Partial Protection in the Crowded Mediterranean Sea: A Regional Meta-Analysis. Sci. Rep. 2017, 7, 8940. [Google Scholar] [CrossRef] [Green Version]
- Sievert, N.A.; Paukert, C.P.; Tsang, Y.-P.; Infante, D. Development and Assessment of Indices to Determine Stream Fish Vulnerability to Climate Change and Habitat Alteration. Ecol. Indic. 2016, 67, 403–416. [Google Scholar] [CrossRef] [Green Version]
- Sabetian, A.; Zhang, J.; Campbell, M.; Walter, R.; Allen, H.; Reid, M.; Wijenayake, K.; Lilkendey, J. Fish Nearshore Habitat-Use Patterns as Ecological Indicators of Nursery Quality. Ecol. Indic. 2021, 131, 108225. [Google Scholar] [CrossRef]
- Guidetti, P.; Baiata, P.; Ballesteros, E.; Di Franco, A.; Hereu, B.; Macpherson, E.; Micheli, F.; Pais, A.; Panzalis, P.; Rosenberg, A.A.; et al. Large-Scale Assessment of Mediterranean Marine Protected Areas Effects on Fish Assemblages. PLoS ONE 2014, 9, e91841. [Google Scholar] [CrossRef]
- Di Franco, E.; Di Franco, A.; Calò, A.; Di Lorenzo, M.; Mangialajo, L.; Bussotti, S.; Bianchi, C.N.; Guidetti, P. Inconsistent Relationships among Protection, Benthic Assemblage, Habitat Complexity and Fish Biomass in Mediterranean Temperate Rocky Reefs. Ecol. Indic. 2021, 128, 107850. [Google Scholar] [CrossRef]
- Vargas, C.A.; Lagos, N.A.; Lardies, M.A.; Duarte, C.; Manríquez, P.H.; Aguilera, V.M.; Broitman, B.; Widdicombe, S.; Dupont, S. Species-Specific Responses to Ocean Acidification Should Account for Local Adaptation and Adaptive Plasticity. Nat. Ecol. Evol. 2017, 1, 0084. [Google Scholar] [CrossRef] [Green Version]
- Audzijonyte, A.; Richards, S.A.; Stuart-Smith, R.D.; Pecl, G.; Edgar, G.J.; Barrett, N.S.; Payne, N.; Blanchard, J.L. Fish Body Sizes Change with Temperature but Not All Species Shrink with Warming. Nat. Ecol. Evol. 2020, 4, 809–814. [Google Scholar] [CrossRef] [PubMed]
- Jones, E.R.; Simning, D.; Serafin, J.; Sepúlveda, M.S.; Griffitt, R.J. Acute Exposure to Oil Induces Age and Species-Specific Transcriptional Responses in Embryo-Larval Estuarine Fish. Environ. Pollut. 2020, 263, 114325. [Google Scholar] [CrossRef] [PubMed]
- Becker, A.; Whitfield, A.K.; Cowley, P.D.; Järnegren, J.; Næsje, T.F. Does Boat Traffic Cause Displacement of Fish in Estuaries? Mar. Pollut. Bull. 2013, 75, 168–173. [Google Scholar] [CrossRef]
- Engås, A.; Løkkeborg, S.; Ona, E.; Soldal, A.V. Effects of Seismic Shooting on Local Abundance and Catch Rates of Cod (Gadus Morhua) and Haddock (Melanogrammus Aeglefinus). Can. J. Fish. Aquat. Sci. 1996, 53, 2238–2249. [Google Scholar] [CrossRef]
- Paxton, A.B.; Taylor, J.C.; Nowacek, D.P.; Dale, J.; Cole, E.; Voss, C.M.; Peterson, C.H. Seismic Survey Noise Disrupted Fish Use of a Temperate Reef. Mar. Policy 2017, 78, 68–73. [Google Scholar] [CrossRef]
- Slotte, A.; Hansen, K.; Dalen, J.; Ona, E. Acoustic Mapping of Pelagic Fish Distribution and Abundance in Relation to a Seismic Shooting Area off the Norwegian West Coast. Fish. Res. 2004, 67, 143–150. [Google Scholar] [CrossRef]
- Amorim, M.C.P.; Vieira, M.; Meireles, G.; Novais, S.C.; Lemos, M.F.L.; Modesto, T.; Alves, D.; Zuazu, A.; Lopes, A.F.; Matos, A.B.; et al. Boat Noise Impacts Lusitanian Toadfish Breeding Males and Reproductive Outcome. Sci. Total Environ. 2022, 830, 154735. [Google Scholar] [CrossRef]
- Faria, A.; Fonseca, P.J.; Vieira, M.; Alves, L.M.F.; Lemos, M.F.L.; Novais, S.C.; Matos, A.B.; Vieira, D.; Amorim, M.C.P. Boat Noise Impacts Early Life Stages in the Lusitanian Toadfish: A Field Experiment. Sci. Total Environ. 2022, 811, 151367. [Google Scholar] [CrossRef]
- Jain-Schlaepfer, S.; Fakan, E.; Rummer, J.L.; Simpson, S.D.; McCormick, M.I. Impact of Motorboats on Fish Embryos Depends on Engine Type. Conserv. Physiol. 2018, 6, coy014. [Google Scholar] [CrossRef] [Green Version]
- Nedelec, S.L.; Radford, A.N.; Gatenby, P.; Davidson, I.K.; Velasquez Jimenez, L.; Travis, M.; Chapman, K.E.; McCloskey, K.P.; Lamont, T.A.C.; Illing, B.; et al. Limiting Motorboat Noise on Coral Reefs Boosts Fish Reproductive Success. Nat. Commun. 2022, 13, 2822. [Google Scholar] [CrossRef] [PubMed]
- Voellmy, I.K.; Purser, J.; Flynn, D.; Kennedy, P.; Simpson, S.D.; Radford, A.N. Acoustic Noise Reduces Foraging Success in Two Sympatric Fish Species via Different Mechanisms. Anim. Behav. 2014, 89, 191–198. [Google Scholar] [CrossRef]
- Fakan, E.P.; McCormick, M.I. Boat Noise Affects the Early Life History of Two Damselfishes. Mar. Pollut. Bull. 2019, 141, 493–500. [Google Scholar] [CrossRef] [PubMed]
- Radford, A.N.; Lèbre, L.; Lecaillon, G.; Nedelec, S.L.; Simpson, S.D. Repeated Exposure Reduces the Response to Impulsive Noise in European Seabass. Glob. Chang. Biol. 2016, 22, 3349–3360. [Google Scholar] [CrossRef] [Green Version]
- Mauro, M.; Pérez-Arjona, I.; Perez, E.J.B.; Ceraulo, M.; Bou-Cabo, M.; Benson, T.; Espinosa, V.; Beltrame, F.; Mazzola, S.; Vazzana, M.; et al. The Effect of Low Frequency Noise on the Behaviour of Juvenile Sparus aurata. J. Acoust. Soc. Am. 2020, 147, 3795–3807. [Google Scholar] [CrossRef] [PubMed]
- O’Connor, J.J.; Lecchini, D.; Beck, H.J.; Cadiou, G.; Lecellier, G.; Booth, D.J.; Nakamura, Y. Sediment Pollution Impacts Sensory Ability and Performance of Settling Coral-Reef Fish. Oecologia 2016, 180, 11–21. [Google Scholar] [CrossRef]
- Cuadros, A.; Basterretxea, G.; Cardona, L.; Cheminée, A.; Hidalgo, M.; Moranta, J. Settlement and Post-Settlement Survival Rates of the White Seabream (Diplodus sargus) in the Western Mediterranean Sea. PLoS ONE 2018, 13, e0190278. [Google Scholar] [CrossRef] [PubMed]
- Simpson, S.D.; Radford, A.N.; Nedelec, S.L.; Ferrari, M.C.O.; Chivers, D.P.; McCormick, M.I.; Meekan, M.G. Anthropogenic Noise Increases Fish Mortality by Predation. Nat. Commun. 2016, 7, 10544. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cheminée, A.; Francour, P.; Harmelin-Vivien, M. Assessment of Diplodus spp. (Sparidae) Nursery Grounds along the Rocky Shore of Marseilles (France, NW Mediterranean). Sci. Mar. 2011, 75, 181–188. [Google Scholar] [CrossRef]
- Ventura, D.; Jona Lasinio, G.; Ardizzone, G. Temporal Partitioning of Microhabitat Use among Four Juvenile Fish Species of the Genus Diplodus (Pisces: Perciformes, Sparidae). Mar. Ecol. 2015, 36, 1013–1032. [Google Scholar] [CrossRef]
- Di Franco, A.; Gillanders, B.M.; De Benedetto, G.; Pennetta, A.; De Leo, G.A.; Guidetti, P. Dispersal Patterns of Coastal Fish: Implications for Designing Networks of Marine Protected Areas. PLoS ONE 2012, 7, e31681. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Spiga, I.; Aldred, N.; Caldwell, G.S. Anthropogenic Noise Compromises the Anti-Predator Behaviour of the European Seabass, Dicentrarchus labrax (L.). Mar. Pollut. Bull. 2017, 122, 297–305. [Google Scholar] [CrossRef]
- Kok, A.C.M.; van Hulten, D.; Timmerman, K.H.; Lankhorst, J.; Visser, F.; Slabbekoorn, H. Interacting Effects of Short-Term and Long-Term Noise Exposure on Antipredator Behaviour in Sand Gobies. Anim. Behav. 2021, 172, 93–102. [Google Scholar] [CrossRef]
- Filiciotto, F.; Giacalone, V.M.; Fazio, F.; Buffa, G.; Piccione, G.; Maccarrone, V.; Di Stefano, V.; Mazzola, S.; Buscaino, G. Effect of Acoustic Environment on Gilthead Sea Bream (Sparus aurata): Sea and Onshore Aquaculture Background Noise. Aquaculture 2013, 414–415, 36–45. [Google Scholar] [CrossRef]
- Nichols, T.A.; Anderson, T.W.; Širović, A. Intermittent Noise Induces Physiological Stress in a Coastal Marine Fish. PLoS ONE 2015, 10, e0139157. [Google Scholar] [CrossRef] [Green Version]
- Celi, M.; Filiciotto, F.; Maricchiolo, G.; Genovese, L.; Quinci, E.M.; Maccarrone, V.; Mazzola, S.; Vazzana, M.; Buscaino, G. Vessel Noise Pollution as a Human Threat to Fish: Assessment of the Stress Response in Gilthead Sea Bream (Sparus aurata, Linnaeus 1758). Fish Physiol. Biochem. 2016, 42, 631–641. [Google Scholar] [CrossRef] [PubMed]
- Underwood, A.J. Experiments in Ecology: Their Logical Design and Interpretation Using Analysis of Variance. J. Mar. Biol. Ass. 1997, 77, 572. [Google Scholar] [CrossRef]
- Verling, E.; Miralles Ricós, R.; Bou-Cabo, M.; Lara, G.; Garagouni, M.; Brignon, J.-M.; O’Higgins, T. Application of a Risk-Based Approach to Continuous Underwater Noise at Local and Subregional Scales for the Marine Strategy Framework Directive. Mar. Policy 2021, 134, 104786. [Google Scholar] [CrossRef]
- Sala, E.; Boudouresque, C.F.; Harmelin-Vivien, M. Fishing, Trophic Cascades, and the Structure of Algal Assemblages: Evaluation of an Old but Untested Paradigm. Oikos 1998, 82, 425. [Google Scholar] [CrossRef]
- Guidetti, P. Marine Reserves Reestablish Lost Predatory Interactions And Cause Community Changes In Rocky Reefs. Ecol. Appl. 2006, 16, 963–976. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Location | Boat Visitation (Boats/Picture) | Boat Passages (Passages/Hour) | Passages Sound Pressure Level (dB re 1 µPa) | Continuous Anthropogenic Noise Band of 250 Hz (dB re 1 µPa) | Duration of Noise Exposure (%) |
|---|---|---|---|---|---|
| Cap Ferrat | 8.3 | 2.4 | 88.2 | 78.2 | 31.5 |
| Eze-sur-Mer | 7.0 | 4.8 | 91.8 | 84.1 | 63.8 |
| Espalmador | 25.3 | 8.6 | 91.9 | 84.4 | 78.2 |
| Marinières | 63.0 | 6.0 | 94.3 | 85.1 | 68.6 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Di Franco, E.; Rossi, F.; Di Iorio, L.; Sdiri, K.; Calò, A.; Di Franco, A.; Harmelin-Vivien, M.; Spennato, G.; Cottalorda, J.-M.; Derijard, B.; et al. Marine Noise Effects on Juvenile Sparid Fish Change among Species and Developmental Stages. Diversity 2023, 15, 92. https://doi.org/10.3390/d15010092
Di Franco E, Rossi F, Di Iorio L, Sdiri K, Calò A, Di Franco A, Harmelin-Vivien M, Spennato G, Cottalorda J-M, Derijard B, et al. Marine Noise Effects on Juvenile Sparid Fish Change among Species and Developmental Stages. Diversity. 2023; 15(1):92. https://doi.org/10.3390/d15010092
Chicago/Turabian StyleDi Franco, Eugenio, Francesca Rossi, Lucia Di Iorio, Khalil Sdiri, Antonio Calò, Antonio Di Franco, Mireille Harmelin-Vivien, Guillaume Spennato, Jean-Michel Cottalorda, Benoit Derijard, and et al. 2023. "Marine Noise Effects on Juvenile Sparid Fish Change among Species and Developmental Stages" Diversity 15, no. 1: 92. https://doi.org/10.3390/d15010092