
“Lepidoptera Flies”, but Not Always…Interactions of Caterpillars and Chrysalis with Soil

Laboratoire Ecologie Fonctionnelle et Environnement, Université Paul Sabatier, CNRS, 31062 Toulouse, France
Diversity 2023, 15(1), 27; https://doi.org/10.3390/d15010027
Submission received: 9 November 2022
/
Revised: 9 December 2022
/
Accepted: 21 December 2022
/
Published: 24 December 2022
(This article belongs to the Special Issue Global Diversity of Lepidopteras)
Abstract
:Lepidoptera, an order of insects traditionally linked to the aerial habitat, are much more diverse in their living environment than the clichéd image we may have of them. The imago stage, which is the most visible in these insects, is not the one that has the most interaction with the environment (usually caterpillars) nor the one that lasts the longest (very often chrysalises). These two stages are often directly related to litter and soil, although only the interaction at the pupal stage seems to follow a phylogenetic logic with two independent evolutionary events for the preference with soil: Use of litter and the upper “O” horizon as protection against predation for the evolutionarily oldest Lepidoptera families, pupation at greater depths (up to 60 centimetres in extreme cases) for the most derived Lepidoptera families; this probably to take advantage of the thermal and moisture buffer provided by the soil. An estimate suggests that about 25% of lepidopteran species worldwide have more or less obligatory interactions with soil.
1. Introduction
Soil is divided into several horizons [1], more or less suitable to host fauna, but even if some organisms like some anecitic annelids can descend to 3 to 4 m in extreme cases [2], the highest density of soil fauna is found in the first 50 cm of the O-horizons and the upper part of the A-horizon.
Excluding annelids, the greatest diversity, density, and biomass are arthropods, mainly springtails, and mites (about 60/70% of the density). Nevertheless, many other arthropods, chelicerates, and myriapods can be found here [3].
Although soil is an important compartment of life [4,5], except for beetles [6] there seems to be no literature that directly links the role of soil on the phylogeny of the different arthropod classes. Only attempts to count species within the different classes [5,7] have been made in relation to evolution.
The soil is a world of darkness, but several factors can explain the high density of organisms present despite the absence of light. Three factors seem most important: The quantity of resources; in tropical forests, it has been shown that 40% of biomass is located between 0 and −1 m [5,8,9]; lower risk of predation [10,11]; and finally, buffering of temperature and humidity variations [12]. The soil therefore represents a rich and above all relatively stable compartment for certain nutrients. The balance of the food web is also more stable due to a buffering effect after a few centimetres of soil or even already within the litter. Predation is less active or at least less diversified than in the above-ground environment. On the other hand, this type of environment requires adaptations to the absence (or low quantity) of light and possibilities of movement limited by its nature [4,5,6,7,8,9,10,11,12].
Among the largest orders of insects, Lepidoptera, with more than 157,000 described species and at least 200,000 estimated [13], are key players in terrestrial ecosystems [14]. The Trichoptera are the evolutionary ancestors of the Lepidoptera with about 13,000 species known. All larvae are aquatic and make a protective sheath of various debris. Their main food is algae growing at very shallow depths [15]. When the larvae are mature, they pupate below the water surface by attaching to a solid object and closing the sheaths. To ensure oxygenation, these nymphs are very mobile creating a constant movement of water [15]. Despite this aquatic origin, almost no Lepidoptera are related to water. Most are herbivores, pollinators, and prey for many birds and small mammals. They actively participate in the food web of terrestrial ecosystems [16].
The cliché image is, of course, represented by the light and fragile winged creature that everyone knows, but the diversity of interactions of these insects with the natural environment is largely unknown. In most terrestrial environments, Lepidopterans are ecologically important because they transform large amounts of plant material into animal biomass [17]. They have a major impact on humans as agricultural pests [18], but also provide important model systems for scientific research [19]. Lepidopterans live on all continents except Antarctica, but three species are endemic to the French Austral Islands which have an extreme cold climate [20]. There are many species in almost every environment, from arid deserts and high mountain peaks to tropical rainforests [21]. Most have adapted to living in relatively specialized ecological niches and may be restricted to a single or a small group of host plants, and even to a single plant part [22,23].
Lepidopterans have a holometabolous cycle in 4 distinct states: egg (static), caterpillar (moderately mobile, main feeding stage), pupa (static), and imago (dispersal and reproductive stage) [24]. The feeding habits of lepidopteran larvae are extremely diverse, depending on the species adaptations to climate, environment, type of food plant, feeding mode, etc. The vast majority of food plants are conifers and flowering plants, but primitive plants such as mosses, liverworts and ferns, and some lichens are consumed by a few families or sub-families [25].
All parts of the plant can be consumed by the larvae. Flowers are consumed in particular by some moths and several Lycaenidae [26]. Cones and fruits [27], as well as their seeds [28], are consumed by other species. Some seed-eaters, such as the flour moth, have become domestic pests, feeding on stored grain and cereals [29]. Buds or stems are widely used [30]. Several families dig deep tunnels in the wood of ligneous plants [31]. Many microlepidopteras feed on dead or decaying plant material, although it is not always clear whether the source of nutrients is the plants or the fungi that grow on them during decomposition [32]. Some caterpillars feed directly from roots in the soil [33]. Finally, a very limited number of species have fully or partly carnivorous caterpillars [34].
This review aims to compare the evolutionary history/phylogeny of Lepidoptera according to the interactions that the different subdivisions of this order have with the soil. Lepidopteran eggs are systematically laid on or above plants and imagos have an open-air existence. It is therefore the caterpillars and chrysalids that will interest us. Considering imagos, males of many species use dilute mineral salts in wetland sludge to increase sperm mobility and thus increase reproductive success [35,36]. However, this interaction with the soil only exists in males and is not obligatory for the ontogeny of the species observed on these muds and are therefore not considered in this work.
The main question will be to verify whether the use of the soil by Lepidoptera is related and/or congruent with the evolutionary history of this order. Interaction with the soil is ultimately a relatively vague concept. Moreover, this interaction is different depending on whether one considers caterpillars where the soil components are food or chrysalis whether the soil is merely the site of the chrysalis. In the light of the results and in order to refine this work, a discussion is proposed to determine whether it is justified to separate the soil into two main zones/horizons for the two stages considered: the superficial soil including litter (“O” horizon) and the deeper soil (from the “OF” horizon); see the material and methods section for more details. An estimate of the number of soil-related species will also be presented for both caterpillars and pupae.
2. Materials and Methods
To select the articles used in this work, the following logic was applied in order to follow the most objective criteria possible.
The first filter was a search for each of the subdivisions. Articles that are too specialised on an agro-economic theme or monospecific studies are eliminated (except in a few cases where the total number of articles per subdivision was very limited). The second filter was a search for each subdivision + the keywords soil and litter. For some subfamilies the number of articles was very limited (3 or 4) and a criterion of number of species treated and date of publication (taking the most recent also to avoid taxonomical problems) was applied. For subdivisions where a large number of articles (>5) was found, a filtering was carried out to take into account all or at least the majority of the subfamilies or tribes of each subdivision. The previous steps allowed a selection of about 400/450 items. A final filtering by date of publication and citation of the majority of the 450 selected articles is carried out to retain a bibliography limited to the 150 articles cited in this work.
The phylogeny base used is the one performed by [37] which, even if slight changes and corrections have been made since that date, remains the most recent phylogeny including all Lepidoptera families. A review of the literature was carried out and for both stages: caterpillar and pupae, discrete values were assigned for each subdivision of the phylogeny used (note that in this phylogeny, heterogeneous subdivisions are considered: superfamilies, families, subfamilies and even a genus).
The matrices used for caterpillars and pupae are based on two criteria: The estimated number of species interacting with the soil per subdivision and the estimated number of species per subdivision described per environmental use and/or food type. It should be noted that the latter two criteria are difficult to separate strictly due to the lack of precise ecological data for most species. Absence: 0, Exceptional (one or two species of the subdivision): 1, from 5 to 40% of the subdivision: 2 and finally for occurrences above 40%: 3. A more general matrix following the same logic is generated for the 81 subdivisions and the following seven characters: leaf use (including leaf-miner caterpillars), stem and trunk use (mainly caterpillars, strictly xylophagous or species digging galleries in the stems), species found in the superficial litter (horizon “OL” according to the nomenclature used by [1]), species feeding on fungi (note: for some species, it is not clear if they use the mycelium that can be found in the “O” horizon, if in doubt, all these subdivisions are considered as belonging to the litter category), species that can live in the soil (from the “OF” to the “A” horizons), species with at least part of the carnivorous larval cycle (mainly entomophagous) and finally species with part of their cycle in fresh water.
Treatment was carried out using the Paup* 4.0b10 software [38]:
A Wagner-ordered parsimony cladistic approach, where not only the differences of characters 0 to 3 are considered, but that these characters are progressive 0 > 1 > 2 > 3; a Goloboff constant (GK = 4) is applied to reduce the effects of homoplasy.
3. Results
3.1. Generalities
3.1.1. Caterpillars
The majority of caterpillars (89%) feed on leaves (Figure 1), whether whole or very often as leaf miners in the case of small species, but many species may have alternative food sources. For about a quarter of the species/subdivisions, leaves are not the exclusive food. Another food source, but still vegetal, are the internal parts of plants, mainly stems, and there are wood-eating species in the strict sense for 21% of the species/subdivisions. Only six subdivisions (7%) feed partly or exclusively on dead plants, mainly leaves, but some species have specialised in feeding on dry flowers. The unambiguously mycophagous species/subdivisions represent 11% of the Lepidoptera. Root-feeding caterpillars in the soil (horizons OF to A) are few: 6%. In many cases, it is very difficult to determine with certainty whether the actual food of the species using litter or found in the soil is not ultimately consuming the mycelium, and therefore the percentage of mycophagous species could be higher at the expense of the other categories. Some species (6%) have a very modified diet compared to the other species and are predators or at least parasites of other insects, mainly as “cuckoo” species towards ants or direct predators towards hemipterans. Finally, a very limited number of litter species have adapted to the freshwater environment.
3.1.2. Chrysalids
While many caterpillars have aposematic colours indicating physical or usually chemical protection, chrysalids, which are static, are homochromatic to the environment and are classically found in places that are more difficult for predators to access including litter and soil. Nevertheless, the majority (64%) of chrysalids or cocoons, for sericogenic species, are found hidden on the host plant or nearby on perennial aerial support such as stone or anthropogenic elements (walls, windows, under roofs, etc.), (Figure 1). About 21% are even more protected inside the dry stems of tall grasses or rushes but also under the bark of trees. The litter, even if it is a dynamic and temporary environment, remains in place long enough to allow the pupation of 20% of lepidopteran species. Fungi, on the other hand, due to their rapid cycle, especially the aerial parts of these organisms, are not used for the pupation of Lepidoptera. On the other hand, the soil represents a refuge zone for chrysalises with 22.7% of the species that can be found there. The chrysalis stage being static, it is clear that there are no carnivorous species nevertheless, some species specifically pupate below ground in ant hills. Finally, there do not seem to be any pupae adapted to aquatic respiration, even among species whose caterpillars feed underwater.
It is possible to roughly assess the changes in the use of the different subdivisions between the caterpillar and chrysalis stages. It can be seen that litter and soil are used comparatively much more by pupae than by caterpillars. On the other hand, the ephemeral aspect of the areal phase of the fungi leads to an absence of chrysalises on this type of support (Figure 2).
3.2. Details Subdivisions
3.2.1. Caterpillars
In all of the following families, no species were found that develop in soil, litter, or use the mycelium of fungi: Agathiphagidae [39], Heterobathmiidae [40], Eriocraniidae [41], Lophocoronidae [42,43,44], Prototheoridae [44], Acanthopteroctetidae [45], Opostegidae [46,47], Nepticulidae [47,48], Andesianidae [49], Cecidosidae [50], Prodoxidae [51,52,53], Incurvariidae [54,55], Nematopogoninae [56], Heliozelidae [57], Tischeriidae [58], Palaephatidae [59], Yponomeutidae [60,61], Urodidae [62], Douglasiidae [63], Schreckensteiniidae [64], Choreutidae [65], Millieriidae [66], Immidae [67], Tortricidae [68,69,70], Heliocosmidae [71], Galacticidae [72,73], Zygaenoids [74,75,76], Lacturidae [77], Himantopteridae [78,79], Aididae [80], Megalopygidae [81,82], Limacodidae [83,84]), Dalceridae [81,85], Cyclotornidae [86,87], Dudgeoneidae [86], Cossidae [88,89], Cossulinae [90], Brachodidae [91], Epipyropidae [86,92], Zeuzerinae [93], Castniidae [94,95], Metarbelinae [88,96], Hypoptinae [97], Pseudurgis [98], Thyrididae [99,100], Hyblaeidae [101], Callidulidae [102], Alucitidae [103], Epermeniidae [104], Carposinidae [105], Copromorphidae [106], Pterophoridae [107], Papilionidae [22,108,109], Hesperiidae [110,111], Hedylidae [112,113], Pieridae [114], Lycaenidae [115,116,117,118], Nymphalidae [119,120], Drepanidae [121], Cimeliidae [122], Doidae [123], Mimallonidae [124], Epicopeiidae [125], Sematuridae [126], Uraniidae [127], Lasiocampidae [128,129], Bombycoidea [130,131], Figure 3.
The following families have caterpillars that feed in the soil, in the litter, and/or on fungal mycelium, Figure 3.
Micropterigidae: About 140 species worldwide, they are small, generally diurnal butterflies with mandibles and not a proboscis such as most lepidopterans. The larvae feed on the dead residues of liverworts (Ranunculaceae) thalli, for the few species whose biology is known, and probably on other litter elements such as decaying bryophytes [132].
Mnesarchaeidae: A very small family endemic to New Zealand of 14 species. They are a small nocturnal species but diurnal activity is documented. The larvae appear to be polyphagous and feed on fungi, algae, mosses, liverworts, and fern sporangia. They weave delicate silk tunnels through which they move to feed [133].
Paleosetidae: Another small family of seven species widely distributed from South America, India, Taiwan, and Australia. The food of the caterpillars is not very clear, but they seem to use mosses (but in this case the development is not complete), mycelium, and possibly roots [134].
Hepialidae: A family of about 400 species distributed throughout the world with some very large species exceeding 20 cm wingspan. Many species feed strictly in the soil on roots, others consume the mycelium and a more limited number develop in the stems or trunks of various woody plants [135].
Adelidae: A family of about 300 species distributed throughout the world except for New Zealand. The major characteristic is the length of the antennae which especially in males can exceed three times the body size of the insect. Caterpillars build silken cocoons that the larva carries and there are documented species that feed in the litter [136].
Tineidae: A huge family of over 3000 species, still largely underestimated. Few species consume aerial parts of plants and food items are very diverse: decaying plant material, fungi (aerial part and mycelium), lichens, and even animal tissues such as wool, horns, etc. [137,138,139].
Psychidae: A family that exceeds 1000 species. In general, females are apterous and do not leave the cocoon made by the caterpillars using various fragments of dead plant material but also some mineral elements. Several species are described as mycophagous [140].
Eriocottidae: A small family of about 210 species from the Old World. These are very a poorly known species of small, generally unadorned moths. All documented species have caterpillars in the soil, which presumably feed on roots or fragments of the upper “O” horizon [141].
Sesiidae: A family of about 1000 species worldwide that are characterised by their hymenopteran mimicry. Most species are diurnal. Caterpillars live inside plants or galls, and some species specialised in roots [86,142].
Gelechioidea: A very large family spread all over the world with 4500 to 5000 species. They are small moths, generally not very colourful except for some tropical species. Some species form cocoons and live in litter, probably feeding on mycelium, but most live in stems, roots, or even seeds [138,143,144,145].
Pyraloidea: One of the largest superfamilies of butterflies with, according to sources, between 6000 and 10,000 species. They are found on all continents and at all latitudes, including the coldest. In general, they are small species but some have a wingspan of over 6 cm. Many species consume seeds on the plant but also in the soil where they can build dense silk tunnels. Some species even have aquatic larvae [29,146,147,148].
Geometridae: A huge family with perhaps over 15,000 species worldwide. Many species are grey but some are very colourful. Most species have a wingspan of 2 or 3 cm but some are larger, up to 9 cm. The caterpillars are mostly aerial but some feed on fungi and probably on litter. A few rare species in Hawaii are carnivorous [149,150,151].
Noctuoidea: the largest super-family of Lepidoptera with a low estimate of at least 20,000 species. This superfamily, which includes the Noctuidae, Erebidae, and Notodontidae, is distributed worldwide and almost all feeding types are described, except perhaps the carnivore. The caterpillars feed on foliage, dead leaves, lichens, and fungi. They are also found consuming roots or miners in stems [152,153,154,155,156,157,158,159].
3.2.2. Chrysalids
In all the following families, no species were found that develop in soil, litter, or using the mycelium of fungi.
Agathiphagidae [39], Heterobathmiidae [40], Prototheoridae [44], Andesianidae [49], Incurvariidae [54,55], Nematopogoninae [56], Adelidae [136], Heliozelidae [57], Tischeriidae [58], Palaephatidae [59], Psychidae [140], Yponomeutidae [60], Urodidae [72], Douglasiidae [73,160], Schreckensteiniidae [64], Choreutidae [65], Millieriidae [66], Immidae [67], Tortricidae [68,69,70], Heliocosmidae [71], Galacticidae [72,73], Zygaenoids [74,76], Lacturidae [77], Himantopteridae [78,79], Aididae [80], Megalopygidae [81,82], Limacodidae [83,84], Dalceridae [81,85], Cyclotornidae [86,87], Sesiidae [86,142], Dudgeoneidae [86], Cossidae [88,89], Cossulinae [90], Brachodidae [91], Epipyropidae [92], Zeuzerinae [93], Castniidae [94,95], Metarbelinae [88,96], Hypoptinae [97], Hyblaeidae [101], Callidulidae [102], Copromorphidae [106], Hesperiidae [110,111], Hedylidae [112,113], Pieridae [114], Lycaenidae [115,116,117,118], Nymphalidae [119,120], Drepanidae [121], Doidae [123], Mimallonidae [124], Epicopeiidae [125], Lasiocampidae [128,129], Figure 4.
The following families have chrysalids in the soil, in the litter, and/or on fungal mycelium, Figure 4.
Micropterigidae: Pupation takes place under decomposed bryophytes [132].
Mnesarchaeidae: Pupation takes place in silk tunnels in the upper part of the litter [133].
Eriocraniidae: A small family of 29 species with primitive features such as the subsistence of mandibles. They are small diurnal species whose caterpillars often live on trees (Betulaceae, Fagaceae). However, for all documented species, pupation takes place underground probably to provide a thermal buffer [41].
Lophocoronidae: A very small family of six Australian species discovered only in 1973. Very little data is available on the biology, but it appears that the caterpillars are aerial while pupation occurs underground as for Eriocraniidae [42].
Paleosetidae: Pupation is assumed to take place in the soil, but there is no verified data on this [134].
Hepialidae: Pupation occurs most often at the feeding site and thus, often in plant roots [135].
Acanthopteroctetidae: A very small family of three to five species that appears to be restricted to the Rocky Mountains from Canada to the USA. Pupation is assumed to be under the litter [45].
Opostegidae: A small family of about 200 species well represented on all continents. Caterpillars are leaf miners, but pupation takes place below the litter [46,47].
Nepticulidae: A family comprising the smallest known Lepidoptera with the largest species not exceeding a centimetre wingspan. Caterpillars mine on the aerial parts of plants, but the pupae are found in litter or shallow soil [47,48].
Cecidosidae: A small family of about 15 species distributed in the Southern Hemisphere, Southern America, South Africa, and New Zealand. The caterpillars feed on galls induced by the oviposition of females or live under the bark of woody plants. Pupation occurs on the soil surface or in the top few centimetres [50].
Prodoxidae: A family of about 100 species mainly distributed in tropical areas. Some genera have developed a very high level of sophistication in plant-insect interaction, particularly on Yuccas, with which they are obligate pollinators and exclusive consumers. Pupation takes place in the first few centimetres of soil at the foot of the host plant [51,52,53].
Tineidae: Several species of this large family pupate in litter or silk tunnels dug in the soil [137,138,139].
Eriocottidae: Pupation takes place just below the litter zone in the upper soil zone in a loose cocoon of silks [141].
Pseudurgis: A genus of about ten species highly localised in east and southern Africa. Caterpillars make shelters from silks and plant fragments on the aerial part of the host plant (Liliaceae). Pupation takes place in the first few centimetres of soil [98].
Thyrididae: A family of 750 small species, some of which are diurnal. Most are tropical and only one flies in Europe. More than 30 host plant families are described for the caterpillars. The cocoons are most often found in the litter at the foot of plants [99,100].
Alucitidae: A family of about 250 species distributed in temperate and subtropical regions. The species are very homogeneous in appearance with characteristic feathery wings. The caterpillars are miners of flowers, fruits, and stems of Caprifoliaceae. Pupation takes place below the litter [103].
Epermeniidae: A family of about 100 small species. Most have diurnal activity and the caterpillars feed mainly on Apiaceae. Pupae are found on soil, at the base of plants, and in flexible silken cocoons [104].
Carposinidae: A family of 290 small, fairly homogeneous species distributed throughout the world. Caterpillars are often fruit miners. Pupation occurs at shallow depths in the soil [105].
Pterophoridae: A large family of some 1300 species distributed worldwide. Adults have a characteristic ‘T’ shape at rest. Caterpillars feed on many plant families including ferns. Pupae are on the plant or in the first few centimetres of soil [107].
Papilionidae: An iconic family of butterflies. Approximately 600 species are spread all over the world. All species have their caterpillars and chrysalids in the open air (sometimes below rocks) except for the most primitive taxon of this family (lineage age exceeding 70 million years); Baronia brevicornis, a species endemic to the Pacific slope of Central Mexico. Chrysalids were found around 20 cm in the ground but up to 60 cm and are hard as stones [22,108,109].
Pyraloidea: Considering the number of species in this super-family, all cases exist with pupae on plants, in stems, in the litter, and in soil [17,146,147,148].
Cimeliidae: A very small family of six Mediterranean species. Adults are very colourful and caterpillars live on Euphorbiaceae. Pupae are on the ground in the litter [122].
Sematuridae: Despite the medium to the large size of these species and the fact that some are diurnal, very little biology is documented. There are reportedly about 40 species in tropical America and one isolated species in South Africa. The pupae are in the litter at the foot of the host plant or slightly in the soil [126].
Uraniidae: Among the most beautiful lepidopterans in the world. There are about 100 species in the tropics, and many species are diurnal. Most caterpillars live on Euphorbiaceae. Pupae are most often found in the litter [127].
Geometridae: This huge family has all pupation modes, and many species use the litter, but some species bury themselves in the soil while staying mostly in the top 10 cm [149,150,151].
Noctuoidea: As for the Geometridae, all cases are found. However, the percentage of species that pupate in the soil is much higher (>60%). Some species pupate quite deeply, up to 15 cm. Some species, such as procession moths, pupate in compact groups of several hundred individuals just below the soil surface [152,153,154,155,156,157,158,159].
Bombycoidea: This superfamily includes the largest known Lepidoptera. In the Saturniidae, the vast majority of species pupate in tough silk cocoons directly on the host plant. In contrast, in the Sphingidae, which are the most recent evolutionary lineage among Lepidoptera, chrysalidation takes place in the soil in large lodges dug into the ground. Most species occur between 5 and 15 cm in depth, but records exist up to 50 cm [130,131].
3.3. Estimation of the Number of Species Having a Relationship with the Soil
For some subdivisions, 100% of the species are soil related, but this proportion can drop to 6/100,000 in the case of the Papilionoidea (which includes the ex-Rhopalocera + Hedylidae), Table 1.
Finally, in light of this literature search, only 2% of caterpillars are strictly subterranean. Knowing that most of the species whose caterpillars are related to the soil (7.6%) also have their chrysalises in litter and/or soil (22.7%), the total is not equivalent to the sum of these two values (30.3%), and an approximate total of 25% of Lepidoptera worldwide are in some way related to this life compartment, Table 1.
4. Discussion
The number of lepidopteran species estimated at the global level varies greatly between sources, but a minimum number would be 200,000 [5], personal estimates (data not shown) give values around 400,000. Clearly, the vast majority of species yet to be discovered/described belong to the microlepidopterans, which are often most closely related to litter or soil.
Lepidopterans are by definition aerial insects and their relationship with the soil is limited [33]. Nevertheless, contrary to popular belief, the most important stage in terms of ecosystem interaction and ecological function is the larval stage [25]. While the adults of many species are visible, the caterpillars, and even more so the chrysalids, are rarely observed even by specialists. As a result, the feeding habits of caterpillars are still very poorly known, and much data on their associations with the environment and plants are incomplete or need to be verified [16,17]. One of the reasons for this lack of knowledge is also that most professional or amateur lepidopterists are not botanists at all, and as a result, there are many errors in the use of plants in the literature. In addition, mentions of soil-dwelling species are rare as most information on these insects comes from amateur lepidopterists who are only interested in adults [130,135,142]. Finally, the greatest source of information on caterpillars, especially those of micro-heterocerans, comes from collections made during soil arthropod studies and are incidental [3,5].
4.1. Caterpillars
Caterpillars are the main feeding stage for Lepidoptera so it is more the trophic component that will be important and not the habitat aspect as such. Nevertheless, the ground may represent a less-exposed foraging area than the aerial zone and thus explain in two different ways the presence of certain species in this environment. Many caterpillars may not be found directly in the soil but in the litter or they feed on dead plant material which in terms of food is less energetic than the living aerial plant: Adelidae [136]; Psychidae [140]; Pyraloidea [29,146,147,148]; Noctuidae [152,153,154,155,156,157,158,159]. Finally, the main question is whether litter species are not ultimately strict consumers of fungal mycelium. Among the subdivisions whose food is probably more mycelium-based than litter-based are: Micropterigidae [132]; Mnesarchaeidae [133]; Paleosetidae [134]; Tineidae [137,138,139]. There do not appear to be any caterpillars with modified burrowing legs, so most species found in the soil are only found in the upper soft soil horizons. In addition, the body of the caterpillars is not very sclerotic and the slightest wound leads to the loss of hemolymph and quickly to the death of the caterpillar, which does not have an efficient coagulation system [161]. These factors mean that few caterpillars can forage in the soil. Four families are root feeders, but only the Eriocottidae have all their species documented developing in the soil in the upper part of the “O” horizon [141]. The caterpillars of the Hepialidae and Sesiidae almost all live under the bark of trees or in the soil and are the only ones to have a reinforced head capsule which allows them to have some adaptation to environments where they have to find their way into closed areas.
The soil/lepidopteran caterpillar relationship is well-verified for some species or families but does not seem to follow evolutionary logic. We can nevertheless note that the Hepialidae and the very small family Paleosetidae belong to the same clade. It is in the large clade Cossoidea/Sesioidea that we find species with a relatively tough integument that could allow adaptation to the soil, but apart from some Sesia species, all others develop in the stems of plants or under the bark of trees. However, the life cycle of more than 80% of these species is unknown and it cannot be excluded that some additive species are subterranean.
Caterpillars therefore have a limited contribution with soil, with about 12,000 species (7.6%) worldwide interacting with soil. Most species are soil litter degraders. As far as true soil caterpillars are concerned, only the Hepialidae (moths) [135] and a few Noctuidae (mainly of the genus Agrotis and Euxoa) [162] and Pyralidae are considered to be pests of crops and may therefore represent an important food source for soil predators due to their local densities.
4.2. Chrysalids
Chrysalids are the most vulnerable static stage during lepidopteran ontogeny and many strategies have been developed to limit the risk of predation: most are homochromatic to the environment and/or are protected by a more or less resistant silken cocoon. As with caterpillars, two broad categories of strategies can be distinguished: species that pupate in the litter, often in the same area as caterpillar feeding sites or at the foot of host plants, and species that burrow into the soil and make a niche several centimetres deep to pupate. Paradoxically, there is more verified evidence of these pupae occurring in or just above the ground than for caterpillars, probably because there are more species in these environments and of course, this stage is not mobile.
Litter is an environment where it is easy to make loose silken cocoons between two dead leaves and thus camouflage from predators. Most chrysalids or their cocoons are homochromatic with the environment which increases their protection: Micropterigidae [132]; Mnesarchaeidae [133]; Opostegidae [46,47]; Nepticulidae [47,48]; Cecidosidae [103]; Tineidae [137,138,139]; Gelechioidea [138,143,144,145]; Thyrididae [99,100]; Alucitidae [103]; Pyraloidea [29,146,147,148]; Cimeliidae [122]; Sematuridae [126]; Uraniidae [127]; and Geometridae [149,150,151].
Many species use the soil for pupation. The soil may represent an area of lower predation, although this is debatable [163], the most likely reason is the thermal buffering effect of the soil, which from 10 to 40 centimetres depth limits day/night extremes and averages monthly temperatures [164]. In addition, the amplitudes of moisture variation are reduced the deeper one goes into the soil. Thus, some caterpillars of large moths such as the Sphingidae may form chrysalidation lodges between 30 and 50 centimetres deep, allowing migratory tropical species of this family to complete their full cycle in mid-Europe, whereas a temperature of −3/−4 °C is lethal for these insects [120]. Only one species of Papilionoidea in the world, the most evolutionary primitive; Baronia brevicornis pupates in the soil [22]. In this case, this chrysalidation, which can be very deep (perhaps an absolute record depth of 60 cm), would be more to ensure the persistence of moisture during the 7-month dry season for this species, which is a strict specialist of the driest tropical forests of Mexico [19].
The subdivisions whose caterpillars mainly pupate in the soil are the following: Eriocraniidae [41]; Lophocoronidae [42]; Palaeosetidae [134]; Hepialidae [23]; Prodoxidae [51,52,53]; Eriocottidae [141]; Tineidae [137,138,139]; Pseudurgis [98]; Epermeniidae [104]; Carposinidae [105]; Noctuoidea [152,153,154,155,156,157,158,159]; Bombycoidea [130,131].
The variety of families/subdivisions using soil for chrysalising is more structured than for caterpillars with two main groups: the evolutionarily oldest families from Eriocraniidae to Prodoxidae (Figure 1) and the most recent families from Gelechioidea to Bombycoidea with the notable exception of the day lepidopterans (Papilionoidea including the nocturnal Hedylidae) where there is a clear reversion from the use of litter and/or soil for pupation for aerial substrate or directly on the plant (the only exception being B. brevicornis, the oldest species of this clade, mentioned earlier). Some species of Lycaenidae which pupate in ant hills benefit from the triple protection of soil, thermal and humidity buffer, and ants themselves as antipredator [115].
The ability to bury oneself at depth is very different between these two large groups; for the oldest, apart from the Hepialidae, which have caterpillars capable of foraging on roots [23], pupation takes place in the first 5 cm of the soil, whereas for the group of more derived families, there is an ability to pupate at depth, even if the record values (Sphingidae, B. brevicornis) do not seem to be able to exceed 60/70 cm. Nevertheless, we must remain cautious, given that more than 80% of the complete life cycles of Lepidoptera are still unknown to us.
The contribution of chrysalises is more important than caterpillars first because of the number of species considered (around 35,500 species in interaction with the soil). Chrysalids are particularly nutritious (lipids: 25/30%; proteins: 30/40%; carbohydrates: 10/15%) and therefore represent an important food source for soil predators but are also industrially used as livestock feed [165]. Chrysalids are also used industrially as fertilizers, which demonstrates their potential ecological role in soil nitrate balance [165].
4.3. General Findings
Compared to the insect order with the largest number of taxa, Coleoptera, interactions with soil are not as strong for Lepidoptera [6]. On the other hand, for the families (or tribes) specialised in deep soil, in general almost all species that share the interaction with soil are included for both Lepidoptera (Sphingidae, Hepialidae) and Coleoptera (Anillina, Leptotyphlini, Osoriini, Torneumatini, Anommatini, Pselaphidae and Scydmaenidae) [6]. For the lineages using the superficial soil and/or bedding, whether for beetles or lepidopterans, few families are purely specialised. However, some apterous arthropods such as springtails and many Staphylinidae beetles are almost entirely dependent on the upper layers of the soil, reflecting a relationship with the evolutionary history of these species [166].
Therefore, the estimate of the number of soil-related species presented in Section 3.3. should be taken with caution and represents only a snapshot of our current knowledge of Lepidoptera biology. Nevertheless, our value of 25% of species using the soil at either the caterpillar or chrysalis stage is consistent with some studies on the soil as a habitat, where 23% of invertebrate species are thought to be in direct contact with the soil at least at some stage of their cycle [5,7]. This rate of 25% drops to 12.5% if the soil dependence of Lepidoptera is strictly considered. All lepidopteran ontogeny stages are important and the contribution of faeces mixed with litter or directly deposited in the soil as well as the organic intake of exuviae that remain at soil level have an important role in soil balance and contribute to better plant growth [167].
Funding
This research received no external funding.
Acknowledgments
This review would never have existed without the help and complicity of Charles Gers, who suggested that I write this work and who, for many years, helped me discover the complexity of “living soil”.
Conflicts of Interest
The author is section editor-in-chief of this journal.
References
- Jabiol, B.; Brêthes, A.; Brun, J.J.; Ponge, J.F.; Toutain, F.; Zanella, A.; Aubert, M.; Bureau, F. Typologie des Formes d’Humus Forestières (Sous Climats Tempérés); Référentiel Pédologique 2008, Quae, Collection Savoir-Faire; Association Française pour l’Étude du Sol: Versailles, France, 2009; pp. 327–355. [Google Scholar]
- Yadav, Y.; Shukla, V. Role of Earthworms in Soil Ecotoxicology Assessment Studies. In Trends in Technology for Agriculture, Food, Environment and Health; Agrobios International: Jodhpur, India, 2021; pp. 433–442. [Google Scholar]
- Coleman, D.C.; Wall, D.H. Chapter 5—Soil Fauna: Occurrence, Biodiversity, and Roles in Ecosystem Function. In Soil Microbiology, Ecology and Biochemistry, 4th ed.; Academic Press: Cambridge, MA, USA, 2015; pp. 111–149. [Google Scholar]
- Gobat, J.M.; Aragno, M.; Matthey, W. Le Sol Vivant: Bases de Pédologie, Biologie des Sols, 3rd ed.; PPUR Presses Polytechniques: Neuchatel, Switzerland, 2010; pp. 415–515. [Google Scholar]
- Decaëns, T.; Jiménez, J.J.; Gioia, C.; Measey, G.J.; Lavelle, P. The values of soil animals for conservation biology. Eur. J. Soil Biol. 2006, 42, S23–S38. [Google Scholar] [CrossRef]
- Andujar, C.; Arribas, P.; Ruzicka, F.; Crampton-Platt, A.; Timmermans, M.J.T.N.; Vogler, A.P. Phylogenetic community ecology of soil biodisversity using mitochondrial metagenomics. Mol. Ecol. 2015, 24, 3603–3617. [Google Scholar] [CrossRef] [PubMed]
- Lavelle, P.; Decaëns, T.; Aubert, M.; Barot, S.; Blouin Bureau, M.F.; Margerie, P.; Mora, P.; Rossi, J.P. Soil invertebrates and ecosystem services. Eur. J. Soil Biol. 2006, 42, S3–S15. [Google Scholar] [CrossRef]
- Mayer, M.; Prescott, C.E.; Abaker, W.E.A.; Augusto, L.; Cécillon, L.; Ferreira, G.W.D.; James, J.; Jandl, R.; Katzensteiner, K.; Laclau, J.P.; et al. Tamm Review: Influence of forest management activities on soil organic carbon stocks: A knowledge synthesis. For. Ecol. Manag. 2020, 466, 118127. [Google Scholar] [CrossRef]
- Teuben, A. Nutrient availability and interactions between soil arthropods and microorganisms during decomposition of coniferous litter: A mesocosm study. Biol. Fertil. Soils 1991, 10, 256–266. [Google Scholar] [CrossRef]
- Hoekstha, J.M.; Bell, R.T.; Launer, A.E.; Murphy, D.D. Soil arthropod abundance in coast redwood forest: Effect of selective timber harvest. Environ. Entomol. 1995, 24, 246–252. [Google Scholar] [CrossRef]
- Staudacher, K.; Rennstam Rubbmark, O.; Birkhofer, K.; Malsher, G.; Sint, D.; Jonsson, M.; Traugott, M. Habitat heterogeneity induces rapid changes in the feeding behaviour of generalist arthropod predators. Funct. Ecol. 2018, 32, 809–819. [Google Scholar] [CrossRef] [Green Version]
- Jensen, A.; Alemu, T.; Alemneh, T.; Pertoldi, C.; Bahrndorff, S. Thermal acclimation and adaptation across populations in a broadly distributed soil arthropod. Funct. Ecol. 2019, 33, 833–845. [Google Scholar] [CrossRef]
- Van Nieukerken, E.J.; Kaila, L.; Kitching, I.; Kristensen, N.P.; Lees, D.C.; Minet, J.; Mitter, C.; Mutanen, M.; Regier, J.C.; Simonsen, T.J.; et al. Order Lepidoptera Linnaeus, 1758. In Animal Biodiversity: An Outline of Higher Classification and Survey of Taxonomic Richness; Zhang, Z.Q., Ed.; Magnolia Press: Auckland, New Zealand, 2011; Volume 3148, pp. 212–221. [Google Scholar]
- Scoble, M.J. The Lepidoptera. Form, Function and Diversity; Natural History Museum Publications (RU); Oxford University Press: Oxford, UK, 1992; pp. 1–404. [Google Scholar]
- Holzenthal, R.W. Tricoptera (Caddisflies). In Encyclopedia of Inland Waters; Likens, G.E., Ed.; Academic Press: Cambridge, MA, USA, 2009; pp. 456–467. [Google Scholar]
- López-Carretero, A.; Díaz-Castelazo, C.; Boege, K.; Rico-Gray, V. Evaluating the spatio-temporal factors that structure network parameters of plant-herbivore interactions. PLoS ONE 2014, 9, e110430. [Google Scholar] [CrossRef]
- Powell, J.A.; Mitter, C.; Farrell, B. 20. Evolution of Larval Food Preferences in Lepidoptera. In Evolution, Systematics, and Biogeography; Kükenthal, W., Ed.; De Gruyter: Berlin, Germany; Boston, MA, USA, 2013; Volume 1, pp. 403–422. [Google Scholar]
- Pichon, A.; Arvanitakis, L.; Roux, O.; Kirk, A.; Alauzet, C.; Bordat, D.; Legal, L. Genetic differentiation among various populations of the diamondback moth, Plutella xylostella (Lepidoptera: Plutellidae). Bull. Entomol. Res. 2006, 96, 137–144. [Google Scholar] [CrossRef]
- Legal, L.; Valet, M.; Dorado, O.; Jesus-Almonte, J.M.; Lopez, K.; Céréghino, R. Lepidoptera are relevant bioindicators of passive regeneration in tropical dry forests. Diversity 2020, 12, 231. [Google Scholar] [CrossRef]
- Enderlein, G. Beitrag zur Kenntnis der antarktischen Landarthropoden. Pringleoptera, eine neue Schmetterlingsgattung aus dem antarktischen Gebiet. Zool. Anz. 1905, 29, 119–125. [Google Scholar]
- New, T.R. Are Lepidoptera an effective ‘umbrella group’ for biodiversity conservation? J. Insect Conserv. 1997, 1, 5–12. [Google Scholar] [CrossRef]
- Legal, L.; Dorado, O.; Machkour-M’Rabet, S.; Leberger, R.; Albre, J.; Mariano, N.A.; Gers, C. Ecological constraints and distribution of the primitive and enigmatic endemic Mexican butterfly Baronia brevicornis (Lepidoptera: Papilionidae). Can. Entomol. 2015, 147, 71–88. [Google Scholar] [CrossRef]
- Grehan, J.R.; Mielke, C.G. Evolutionary biogeography and tectonic history of the ghost moth families Hepialidae, Mnesarchaeidae, and Palaeosetidae in the Southwest Pacific (Lepidoptera: Exoporia). Zootaxa 2018, 4415, 243–275. [Google Scholar] [CrossRef]
- Loof, A.; Schoofs, L. Calcitox-Metamorphosis in Insects: The Calcium (Ca2+)-Homeostasis System as the Integrated Primordial Receptor System for both Juvenile Hormone and Ecdysteroids. Life Excit. Biol. 2020, 7, 41–81. [Google Scholar]
- Common, I.F.B. Moths of Australia; Brill: Leyde, The Netherlands, 1990; pp. 1–535. [Google Scholar]
- Vargas, H.A. Lycaenid caterpillars (Lepidoptera, Lycaenidae) eating flowers of Dalea pennellii var. chilensis (Fabaceae) in the northern Chilean Andes. Rev. Bras. Entomol. 2014, 58, 309–312. [Google Scholar] [CrossRef] [Green Version]
- Brown, J.W. Patterns of Lepidoptera herbivory on conifers in the New World. J. Asia Pac. Biodivers. 2018, 11, 1–10. [Google Scholar] [CrossRef]
- Valdés, A.; Ehrlén, J. Caterpillar seed predators mediate shifts in selection on flowering phenology in their host plant. Ecology 2017, 98, 228–238. [Google Scholar] [CrossRef] [Green Version]
- Nouri, B.; Fotouhi, K.; Mohtasebi, S.S.; Nasiri, A.; Goldansaaz, S.H. Detection of different densities of Ephestia kuehniella pest on white flour at different larvae instar by an electronic nose system. J. Stored Prod. Res. 2019, 84, 101522. [Google Scholar] [CrossRef]
- Naino-Jika, A.K.; Le Ru, B.; Capdevielle-Dulac, C.; Chardonnet, F.; Silvain, J.F. Population genetics of the Mediterranean corn borer (Sesamia nonagrioides) differs between wild and cultivated plants. PLoS ONE 2020, 15, e0230434. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pasqualini, E.; Natale, D. Zeuzera pyrina and Cossus cossus (Lepidoptera: Cossidae) control by pheromones: Four years advances in Italy. IOBC WPRS Bull. 1999, 22, 115–124. [Google Scholar]
- Nishida, K.; Robbins, R.K. One side makes you taller: A mushroom–eating butterfly caterpillar (Lycaenidae) in Costa Rica. Neotrop. Biol. Conserv. 2020, 15, 463. [Google Scholar] [CrossRef]
- Potocký, P.; Bartoňová, A.; Beneš, J.; Zapletal, M.; Konvička, M. Life-history traits of Central European moths: Gradients of variation and their association with rarity and threats. Insect Conserv. Divers. 2018, 11, 493–505. [Google Scholar] [CrossRef]
- Mota, L.L.; Kaminski, L.A.; Freitas, A.V. The tortoise caterpillar: Carnivory and armoured larval morphology of the metalmark butterfly Pachythone xanthe (Lepidoptera: Riodinidae). J. Nat. Hist. 2020, 54, 309–319. [Google Scholar] [CrossRef]
- Molleman, F. Puddling: From natural history to understanding how it affects fitness. Entomol. Exp. Appl. 2010, 134, 107–113. [Google Scholar] [CrossRef]
- Legal, L.; Dorado, O.; Albre, J.; Bermudez-Torres, K.; Lopez, K. Mariposas Diurnas, Reserva de la Biosfera Sierra de Huautla, Estado de Morelos, México; Tropico Seco Ediciones: Cuernavaca, Mexico, 2017; pp. 1–330. [Google Scholar]
- Regier, J.C.; Mitter, C.; Kristensen, N.P.; Davis, D.R.; Van Nieukerken, E.J.; Rota, J.; Simonsen, T.J.; Mitter, K.T.; Kawahara, A.Y.; Yen, S.H.; et al. A molecular phylogeny for the oldest (nonditrysian) lineages of extant Lepidoptera, with implications for classification, comparative morphology and life-history evolution. Syst. Entomol. 2015, 40, 671–704. [Google Scholar] [CrossRef]
- Swofford, D.L. PAUP-Phylogenetic Analysis Using Parsimony, Version 4.0b10; Sinauer Associates: Sunderland, MA, USA, 2001.
- Dumbleton, L. A new genus of seed-infesting micropterygid moths. Pac. Sci. 1952, 6, 17–29. [Google Scholar]
- Heppner, J.B. Valdivian Archaic Moths (Lepidoptera: Heterobathmiidae). In Encyclopedia of Entomology; Capinera, J.L., Ed.; Springer: Dordrecht, The Netherlands, 2008; p. 653. [Google Scholar]
- Powell, J.A. Chapter 151—Lepidoptera: Moths, Butterflies. In Encyclopedia of Insects, 2nd ed.; Resh, V.H., Cardé, R.T., Eds.; Academic Press: Cambridge, MA, USA, 2009; pp. 559–587. [Google Scholar]
- Nielsen, E.S.; Kristensen, N.P. The Australian moth family Lophocoronidae and the basal phylogeny of the Lepidoptera Glossata. Invertebr. Taxon 1996, 10, 1199–1302. [Google Scholar] [CrossRef]
- Faucheux, M.J. Antennal sensilla of male Lophocorona pediasia Common 1973 and their phylogenetic implications (Lepidoptera: Lophocoronidae). Ann. Soc. Entomol. Fr. 2006, 42, 113–118. [Google Scholar] [CrossRef]
- Mey, W.; Léger, T.; Lien, V.V. Extant and fossil records of currently Australian primitive moths in South-East Asia and their biogeographic significance (Lepidoptera, Micropterigidae, Agatiphagidae, Lophocoronidae). Nota Lepidopterol. 2021, 44, 29–56. [Google Scholar] [CrossRef]
- Minet, J. Proposal of an infraordinal name for the Acanthopteroctetidae (Lepidoptera). Bull. Soc. Entomol. France 2002, 107, 222. [Google Scholar] [CrossRef]
- Davis, D.R. Generic revision of the Opostegidae, with a synoptic catalog of the world’s species (Lepidoptera: Nepticuloidea). Smithson. Contrib. Zool. 1989, 478, 1–97. [Google Scholar] [CrossRef]
- Van Nieukerken, E.J.; Doorenweerd, C.; Hoare, R.; Davis, D. Revised classification and catalogue of global Nepticulidae and Opostegidae (Lepidoptera, Nepticuloidea). ZooKeys 2016, 628, 65–246. [Google Scholar] [CrossRef] [PubMed]
- Shin, Y.M.; Lim, J.; Lee, B.W.; Byun, B.K. Three species of the genus Stigmella Schrank (Lepidoptera: Nepticulidae) feeding Quercus (Fagaceae) new to Korea. J. Asia Pac. Biodivers. 2020, 13, 599–604. [Google Scholar] [CrossRef]
- Davis, D.R.; Gentile, P. Andesianidae, a new family of monotrysian moths (Lepidoptera: Andesianoidea) from austral South America. Invertebr. Syst. 2003, 17, 15–26. [Google Scholar] [CrossRef] [Green Version]
- Moreira, G.R.P.; Eltz, R.P.; Pase, R.B.; Silva, G.T.; Bordignon, S.A.L.; Mey, W.; Gonçalves, G.L. Cecidonius pampeanus, gen. et sp. n.: An overlooked and rare, new gall-inducing micromoth associated with Schinus in southern Brazil (Lepidoptera, Cecidosidae). ZooKeys 2017, 695, 37–74. [Google Scholar] [CrossRef] [Green Version]
- Davis, D.R. A Revision of the Moths of the Subfamily Prodoxinae (Lepidoptera: Incurvariidae); Bulletin of the United States National Museum; Smithsonian Institution: Washington, DC, USA, 1967; Volume 255, pp. 1–170. [Google Scholar]
- Powell, J.A. Interrelationships of yuccas and yucca moths. Trends Ecol. Evol. 1992, 7, 10–15. [Google Scholar] [CrossRef]
- Pellmyr, O.; Thompson, J.N.; Brown, J.; Harrison, R.G. Evolution of pollination and mutualism in the yucca moth lineage. Am. Nat. 1996, 148, 827–847. [Google Scholar] [CrossRef]
- Heppner, J.B. Leafcutter Moths (Lepidoptera: Incurvariidae). In Encyclopedia of Entomology; Capinera, J.L., Ed.; Springer: Dordrecht, The Netherlands, 2008; p. 656. [Google Scholar]
- Hirowatari, T.; Yagi, S.; Ohshima, I.; Huang, G.H.; Wang, M. Review of the genus Vespina (Lepidoptera, Incurvariidae) with two new species from China and Japan. Zootaxa 2021, 4927, 209–233. [Google Scholar] [CrossRef]
- Heppner, J.B. Longhorned Fairy Moths (Lepidoptera: Adelidae). In Encyclopedia of Entomology; Capinera, J.L., Ed.; Springer: Dordrecht, The Netherlands, 2008; p. 2226. [Google Scholar]
- Van Nieukerken, E.J.; Eiseman, C.S. Splitting the leafmining shield-bearer moth genus Antispila Hübner (Lepidoptera, Heliozelidae): North American species with reduced venation placed in Aspilanta new genus, with a review of Heliozelid morphology. ZooKeys 2020, 957, 105–161. [Google Scholar] [CrossRef] [PubMed]
- Stonis, J.R.; Diškus, A.; Remeikis, A.; Solis, M.A.; Katinas, L. Exotic-looking Neotropical Tischeriidae (Lepidoptera) and their host plants. ZooKeys 2020, 970, 117–158. [Google Scholar] [CrossRef] [PubMed]
- Davis, D.R. A New Family of Monotrysian Moth from Austral South America (Lepidoptera: Palaephatidae), with a Phylogenetic Review of the Monotrysia; Smithsonian Contributions to Zoology; Smithsonian Institution: Washington, DC, USA, 1986; Volume 434, pp. 1–202. [Google Scholar]
- Heppner, J.B. Ermine Moths (Lepidoptera: Yponomeutidae). In Encyclopedia of Entomology; Capinera, J.L., Ed.; Springer: Dordrecht, The Netherlands, 2008; pp. 1360–1361. [Google Scholar]
- Sohn, J.C. Molecular Phylogenetics, Biodiversity and Life History Evolution of Yponomeutoidea (Lepidoptera: Ditrysia), with a Catalog and an Overview of the Lepidopteran Fossils. Ph.D. Thesis, University of Maryland, College Park, MD, USA, 2013. [Google Scholar]
- Liu, T.; Wang, S.X. First report of the family Urodidae from China, with descriptions of the immature stages of Wockia magna Sohn, 2014 (Lepidoptera: Urodoidea). SHILAP Rev. Lepidopterol. 2015, 43, 499–505. [Google Scholar]
- Poinar, G., Jr. A new genus of moths (Lepidoptera: Gracillarioidea: Douglasiidae) in Myanmar amber. Hist. Biol. 2019, 31, 898–902. [Google Scholar] [CrossRef]
- Sam, K.; Ctvrtecka, R.; Miller, S.E.; Rosati, M.E.; Molem, K.; Damas, K.; Gewa, B.; Novotny, V. Low host specificity and abundance of frugivorous Lepidoptera in the lowland rain forests of Papua New Guinea. PLoS ONE 2017, 12, e0171843. [Google Scholar] [CrossRef] [PubMed]
- Rota, J.; Aguiar, A.; Karsholt, O. Choreutidae of Madeira: Review of the known species and description of the male of Anthophila threnodes (Walsingham, 1910) (Lepidoptera). Nota Lepidopterol. 2014, 37, 91–103. [Google Scholar] [CrossRef] [Green Version]
- Kawahara, A.Y.; Plotkin, D.; Hamilton, C.A.; Gough, H.; St Laurent, R.; Owens, H.L.; Homziak, N.T.; Barber, J.B. Diel behavior in moths and butterflies: A synthesis of data illuminates the evolution of temporal activity. Org. Divers. Evol. 2018, 18, 13–27. [Google Scholar] [CrossRef]
- Heppner, J.B. Imma Moths (Lepidoptera: Immidae). In Encyclopedia of Entomology; Capinera, J.L., Ed.; Springer: Dordrecht, The Netherlands, 2008; p. 1921. [Google Scholar]
- Brown, J.W.; Basset, Y.; Panmeng, M.; Putnaul, S.; Miller, S.E. Host records for Tortricidae (Lepidoptera) reared from seeds and fruits in a Thailand rainforest. Proc. Entomol. Soc. Wash. 2019, 121, 544–556. [Google Scholar] [CrossRef]
- Basoalto, A.; Ramírez, C.C.; Lavandero, B.; Devotto, L.; Curkovic, T.; Franck, P.; Fuentes-Contreras, E. Population genetic structure of codling moth, Cydia pomonella (L.) (Lepidoptera: Tortricidae), in different localities and host plants in Chile. Insects 2020, 11, 285. [Google Scholar] [CrossRef]
- Liu, Y.; Li, X.; Yan, X.; Li, G. Effect of Different Host Plants on Growth, Development and Reproduction of Yellow Tortrix Moth Acleris fimbriana (Lepidoptera: Tortricidae). Pak. J. Zool. 2021, 53, 475–481. [Google Scholar] [CrossRef]
- Fagua, G.; Condamine, F.L.; Horak, M.; Zwick, A.; Sperling, F.A. Diversification shifts in leafroller moths linked to continental colonization and the rise of angiosperms. Cladistics 2017, 33, 449–466. [Google Scholar] [CrossRef] [PubMed]
- Sohn, J.C.; Adamski, D. A New Species of Wockia Heinemann, 1890 (Lepidoptera: Urodidae) from Korea. Proc. Entomol. Soc. Wash. 2008, 110, 556–561. [Google Scholar] [CrossRef] [Green Version]
- Van Driesche, R.; LaForest, J.H.; Bargeron, C.T.; Reardon, R.C.; Herlihy, M. Forest Pest Insects in North America: A Photographic Guide; USDA Forest Service: Morgantown, WV, USA, 2013.
- Tremewan, W.G. A Bibliography of the Zygaenidae (Lepidoptera: Zygaenidae); Brill: Leiden, The Netherlands, 1996; pp. 1–188. [Google Scholar]
- De Castro, E.C.; Musgrove, J.; Bak, S.; McMillan, W.O.; Jiggins, C.D. Phenotypic plasticity in chemical defence of butterflies allows usage of diverse host plants. Biol. Lett. 2021, 17, 20200863. [Google Scholar] [CrossRef] [PubMed]
- Naveena, K.; Chinniah, C.; Shanthi, M. Cyanogenic glycosides and plant-herbivore interactions. J. Entomol. Zool. Stud. 2021, 9, 1345–1350. [Google Scholar] [CrossRef]
- Matson, T.A.; Wagner, D.L.; Miller, S.E. A Revision of North American Lactura (Lepidoptera, Zygaenoidea, Lacturidae). ZooKeys 2019, 846, 75. [Google Scholar] [CrossRef] [PubMed]
- Niehuis, O.; Naumann, C.M.; Misof, B. Phylogenetic analysis of Zygaenoidea small-subunit rRNA structural variation implies initial oligophagy on cyanogenic host plants in larvae of the moth genus Zygaena (Insecta: Lepidoptera). Zool. J. Linn. Soc. 2006, 147, 367–381. [Google Scholar] [CrossRef] [Green Version]
- Symondson, W.O.; Holloway, J.D.; Goossens, B.; Müller, C.T. Bornean caterpillar (Lepidoptera) constructs cocoon from Vatica rassak (Dipterocarpaceae) resin containing multiple deterrent compounds. J. Nat. Hist. 2015, 49, 553–560. [Google Scholar] [CrossRef]
- Specht, A.; Corseuil, E.; Formentini, A.C. Lepidópteros de importância médica ocorrentes no Rio Grande do Sul. II. Aididae e Limacodidae. Biociencias 2005, 13, 89–94. [Google Scholar]
- Diniz, I.R.; Morais, H.C. Lepidopteran caterpillar fauna of cerrado host plants. Biodiv. Cons. 1997, 6, 817–836. [Google Scholar] [CrossRef]
- Hall, D.W. Puss Caterpillar (Larva), Southern Flannel Moth (Adult), Megalopyge opercularis (JE Smith 1797), (Insecta: Lepidoptera: Zygaenoidea: Megalopygidae). EDIS. 2013. Available online: https://entnemdept.ufl.edu/creatures/misc/moths/puss.htm (accessed on 23 December 2022).
- Wu, C.S.; Solovyev, A.V. A review of the genus Miresa Walker in China (Lepidoptera: Limacodidae). J Insect Sci. 2011, 11, 1–16. [Google Scholar] [CrossRef] [Green Version]
- Stoepler, T.M.; Lill, J.T.; Murphy, S.M. Intraplant movement of generalist slug caterpillars (Limacodidae: Lepidoptera): Effects of host plant and light environment. Environ. Entomol. 2014, 43, 1561–1573. [Google Scholar] [CrossRef] [PubMed]
- Lin, Y.C.; Lin, R.J.; Braby, M.F.; Hsu, Y.F. Evolution and losses of spines in slug caterpillars (Lepidoptera: Limacodidae). Ecol. Evol. 2019, 9, 9827–9840. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Heppner, J.B. Mexican Lepidoptera biodiversity. Insecta Mundi 2002, 16, 171–190. [Google Scholar]
- Yen, S.H.; Robinson, G.S.; Quicke, D.L. The phylogenetic relationships of Chalcosiinae (Lepidoptera, Zygaenoidea, Zygaenidae). Zool. J. Linn. Soc. 2005, 143, 161–341. [Google Scholar] [CrossRef]
- Yakovlev, R.V.; Sulak, H.; Witt, T.J. Preliminary list of Cossidae sensu str. (Lepidoptera, Cossoidea: Cossidae: Cossinae & Zeuzerinae) of the Republic of Angola with description of a new Strigocossus species. Zootaxa 2019, 4586, 445–460. [Google Scholar]
- Rishi, R.R.; Sundararaj, R. Incidence of bark feeding borer, Indarbela quadrinotata Walker (Lepidoptera: Cossidae) on Sonneratia apetala in the Mangroves of Maharashtra, India. J. Entomol. Zool. Stud. 2021, 9, 207–209. [Google Scholar] [CrossRef]
- Montes-Rodríguez, J.M. Three new registers of stem-borer insects in Terminalia ivorensis A. Chev. Rev. Mex. Cienc. For. 2017, 8, 27–38. [Google Scholar]
- Bella, S.; Bartsch, D.; Laštůvka, Z. Bibliographic summary and new records of the Brachodidae and Sesiidae of Sicily, with an updated list and some comments on the distribution of Italian species (Lepidoptera, Cossoidea). Spixiana 2017, 40, 139–156. [Google Scholar]
- Dyar, H.G. A lepidopterous larva on a leaf-hooper. (Epipyros barberiana n. sp.). Proc. Ent. Soc. Wash. 1902, 5, 43–45. [Google Scholar]
- Hashim, S.M. Natural Host Rearing and some Biological Aspects of the Leopard Moth Zeuzera pyrina L. (Lepidoptera: Cossidae) on Poplar Trees (Populus Sp.) (Malpighiales Salicaceae) in Egypt. J. Plant Prot. Pathol. 2019, 10, 317–319. [Google Scholar] [CrossRef]
- González, J.M.; López, G.B.; Huerta, P.J.P.; Miller, J.Y. A new genus of Castniinae (Lepidoptera Castniidae) with comments on comparative morphology and bionomics of its assigned species. Zootaxa 2019, 4668, 89–104. [Google Scholar] [CrossRef] [PubMed]
- Williams, A.A.; Williams, M.R.; Bishop, C.L.; Gamblin, T.; Fissioli, J.; Coppen, R.A. Notes on the biology, ecology, life history and conservation of the graceful sun-moth ‘Synemon gratiosa’ westwood, 1877 (Lepidoptera: Castniidae). Aust. Entomol. 2021, 48, 17–32. [Google Scholar]
- Martinez, J.I.; Crane, J.H.; Wasielewski, J.; Miller, J.Y.; Carrillo, D. Lepidoptera pests of sapodilla (Manilkara zapota (L.) van Royen) in south Florida, with some comments on life history and natural control. Insecta Mundi 2019, 0739, 1–26. [Google Scholar]
- Scaccini, D.; Ruzzier, E.; Daane, K.M. Givira ethela (Neumoegen and Dyar, 1893) (Lepidoptera: Cossidae), A Previously Unidentified Pest on Vitis vinifera (L.). Insects 2021, 12, 239. [Google Scholar] [CrossRef] [PubMed]
- Taylor, J.S. Notes on Lepidoptera in the Eastern Cape Province (Part III). J. Entomol. Soc. S. Afr. 1954, 16, 166–167. [Google Scholar]
- Aiello, A. Life history of Dysodia sica (Lepidoptera: Thyrididae) in Panama. Psyche 1980, 87, 121–130. [Google Scholar] [CrossRef] [Green Version]
- Darling, D.C. Morphology and behavior of the larva of Calindoea trifascialis (Lepidoptera: Thyrididae), a chemically defended retreat-building caterpillar from Vietnam. Zootaxa 2003, 225, 1–16. [Google Scholar] [CrossRef] [Green Version]
- Baksha, M.W.; Crawley, M.J. Relative preference of different host plants to teak defoliator, Hyblaea puera Cram. (Hyblaeidae: Lepidoptera) in Bangladesh. Bangladesh J. For. Sci. 1995, 24, 21–25. [Google Scholar]
- Yen, S.H.; Wu, S. Biota Taiwanica: Calliduloidea, Callidulidae, Callidulinae; National Sun Yat-Sen University: Kaohsiung, Taiwan, 2009; pp. 1–25. [Google Scholar]
- Moreira, G.R.P.; Pereira, C.M.; Becker, V.O.; Specht, A.; Gonçalves, G.L. A new cecidogenous species of many-plumed moth (Alucitidae) associated with Cordiera A. Rich. ex DC. (Rubiaceae) in the Brazilian Cerrado. Zoologia 2019, 36, e34604. [Google Scholar] [CrossRef] [Green Version]
- Gaedike, R. New data on the taxonomy, distribution and host plants of Australian Epermeniidae (Lepidoptera: Epermenioidea). Zootaxa 2018, 4524, 33–50. [Google Scholar] [CrossRef]
- Wang, Y.Z.; Li, B.Y.; Hoffmann, A.A.; Cao, L.J.; Gong, Y.J.; Song, W.; Wei, S.J. Patterns of genetic variation among geographic and host-plant associated populations of the peach fruit moth Carposina sasakii (Lepidoptera: Carposinidae). BMC Evol. Biol. 2017, 17, 265. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- De Benedictis, J.A. On the taxonomic position of Ellabella Busck, with descriptions of the larva and pupa of E. bayensis (Lepidoptera: Copromorphidae). J. Res. Lepid. 1984, 23, 74–82. [Google Scholar] [CrossRef]
- Bigot, L.; Etienne, J. Les Pterophoridae de l’île de la Guadeloupe (Lepidoptera). Bull. Soc. Entomol. France 2009, 114, 463–467. [Google Scholar] [CrossRef]
- Miller, J.S. Host-plant relationships in the Papilionidae (Lepidoptera): Parallel cladogenesis or colonization? Cladistics 1987, 3, 105–120. [Google Scholar] [CrossRef]
- Aubert, J.; Legal, L.; Descimon, H.; Michel, F. Molecular phylogeny of swallowtail butterflies of the tribe Papilionini (Papilionidae, Lepidoptera). Mol. Phylogenet. Evol. 1999, 12, 156–167. [Google Scholar] [CrossRef]
- Nakasuji, F. Egg size of skippers (Lepidoptera: Hesperiidae) in relation to their host specificity and to leaf toughness of host plants. Ecol. Res. 1987, 2, 175–183. [Google Scholar] [CrossRef]
- Warren, A.D.; Ogawa, J.R.; Brower, A.V.Z. Revised classification of the family Hesperiidae (Lepidoptera: Hesperioidea) based on combined molecular and morphological data. Syst. Entomol. 2009, 34, 467–523. [Google Scholar] [CrossRef]
- Janz, N.; Nylin, S. Butterflies and plants: A phylogenetic study. Evolution 1998, 52, 486–502. [Google Scholar]
- Ferrer-Paris, J.R.; Sanchez-Mercado, A.; Viloria, A.L.; Donaldson, J. Congruence and diversity of butterfly-host plant associations at higher taxonomic levels. PLoS ONE 2013, 8, e63570. [Google Scholar] [CrossRef] [Green Version]
- Chew, F.S.; Renwick, J.A.A. Host plant choice in Pieris butterflies. In Chemical Ecology of Insects 2; Springer: Boston, MA, USA, 1995; pp. 214–238. [Google Scholar]
- Koubínová, D.; Dincă, V.; Dapporto, L.; Voda, R.; Suchan, T.; Vila, R.; Alvarez, N. Genomics of extreme ecological specialists: Multiple convergent evolution but no genetic divergence between ecotypes of Maculinea alcon butterflies. Sci. Rep. 2017, 7, 13752. [Google Scholar] [CrossRef] [Green Version]
- Terblanche, R.F. Anoplolepis steingroeveri (Forel, 1894) (Hymenoptera: Formicidae), a new associated ant species record for myrmecophilous butterflies in Africa and new hostplant records for Crudaria (Wallengren, 1875) (Lepidoptera: Lycaenidae: Aphnaeinae). Metamorphosis 2017, 28, 41–48. [Google Scholar]
- Carleial, S.; Maurel, N.; van Kleunen, M.; Stift, M. Oviposition by the Mountain Alcon Blue butterfly increases with host plant flower number and host ant abundance. Basic Appl. Ecol. 2018, 28, 87–96. [Google Scholar] [CrossRef]
- Ohsaki, N.; Ohata, M.; Sato, Y.; Rausher, M.D. Host Plant Choice Determined by Reproductive Interference between Closely Related Butterflies. Am. Nat. 2020, 196, 512–523. [Google Scholar] [CrossRef] [PubMed]
- Hardman, J.E.; Smith, M. How important are olfactory cues for host-plant detection by migrating Danaus chrysippus (Linnaeus, 1758) (Lepidoptera: Nymphalidae, Danainae) in Cyprus? Entomol. Gaz. 2019, 70, 223–238. [Google Scholar]
- Ramos, R.R.; Francini, R.B.; Habib, M.E.E.D.M.; Freitas, A.V.L. Seasonal Patterns of Host Plant Use in an Assemblage of Heliconiini Butterflies (Lepidoptera: Nymphalidae) in a Neotropical forest. Neotrop. Entomol. 2021, 50, 358–365. [Google Scholar] [CrossRef] [PubMed]
- Yadav, C.; Yack, J.E. Immature stages of the masked birch caterpillar, Drepana arcuata (Lepidoptera: Drepanidae) with comments on feeding and shelter building. J. Insect Sci. 2018, 18, 1–9. [Google Scholar] [CrossRef]
- León, Y.M.; Baquero, A.I.; Farino, T.; Jiménez, R.E.; Petty, S.J. Aportaciones a la distribución, ecología e identificación de la oruga de Axia margarita (Hübner, 1813) en España (Lepidoptera: Cimeliidae). Bol. SEA 2018, 63, 297–300. [Google Scholar]
- Miller, J.S.; Wagner, D.L.; Opler, P.A.; Lafontaine, J.D. Fascicle 22.1A: Drepanoidea, Doidae; Noctuoidea, Notodontidae (Part): Pygaerinae, Notodontinae, Cerurinae, Phalerinae, Periergosinae, Dudusinae, Hemiceratinae. In The Moths of America North of Mexico; Wedge Entomological Research Foundation: Bakersfield, CA, USA, 2018; pp. 1–348. [Google Scholar]
- St Laurent, R.A.; Wagner, D.L.; Reeves, L.E.; Kawahara, A.Y. Notes on the larva and natural history of Lacosoma arizonicum Dyar (Mimallonoidea, Mimallonidae) with new host and parasitoid records. J. Lepid. Soc. 2017, 71, 177–181. [Google Scholar] [CrossRef]
- Huang, S.Y.; Zhang, Y.; Wang, M.; Fan, X.L. First record of the genus Schistomitra Butler, 1881 (Lepidoptera, Epicopeiidae) from China, with the description of a new species. ZooKeys 2019, 878, 145. [Google Scholar] [CrossRef]
- Minet, J.; Scoble, M.J. The Drepanoid/Geometroid Assemblage. In Lepidoptera, Moths and Butterflies. Volume 1, Evolution, Systematics, and Biogeography. Handbook of Zoology, vol. IV, Arthropoda: Insecta, Part 35; Kristensen, N.P., Ed.; Walter de Gruyter: Berlin, Germany; New York, NY, USA, 1999; pp. 301–320. [Google Scholar]
- Cock, M.J.; Shaw, R.H. The biology of Epiplema albida (Hampson, 1891) (Lepidoptera, Uraniidae, Epipleminae) on Sri Lankan Privet, Ligustrum robustum (Roxb.) Blume subsp. walkeri (Decne.) PS Green (Oleaceae). Trop. Lepid. Res. 2017, 27, 96–100. [Google Scholar]
- Kadej, M.; Zając, K.; Tarnawski, D. Oviposition site selection of a threatened moth Eriogaster catax (Lepidoptera: Lasiocampidae) in agricultural landscape—Implications for its conservation. J. Insect Conserv. 2018, 22, 29–39. [Google Scholar] [CrossRef] [Green Version]
- Raath, M.J.; Le Roux, P.C.; Veldtman, R.; Greve, M. Incorporating biotic interactions in the distribution models of African wild silk moths (Gonometa species, Lasiocampidae) using different representations of modelled host tree distributions. Aus. Ecol. 2018, 43, 316–327. [Google Scholar] [CrossRef] [Green Version]
- Shimoda, M.; Kiuchi, M. Effect of chilling of diapause pupa on adult emergence in the sweet potato hornworm, Agrius convolvuli (Linné) (Lepidoptera; Sphingidae). Appl. Entomol. Zool. 1997, 32, 617–624. [Google Scholar] [CrossRef] [Green Version]
- Mejia, L.B.; Arnal, P.; Hallwachs, W.; Haxaire, J.; Janzen, D.; Kitching, I.J.; Rougerie, R. A global food plant dataset for wild silkmoths and hawkmoths and its use in documenting polyphagy of their caterpillars (Lepidoptera: Bombycoidea: Saturniidae, Sphingidae). Biodivers. Data J. 2020, 8, e60027. [Google Scholar] [CrossRef] [PubMed]
- Imada, Y.; Kawakita, A.; Makoto, K. Allopatric distribution and diversification without niche shift in a bryophyte-feeding basal moth lineage (Lepidoptera: Micropterigidae). Proc. R. Soc. B 2011, 278, 3026–3033. [Google Scholar] [CrossRef] [Green Version]
- Gibbs, G.W.; Kristensen, N.P. Mnesarchaeidae (Insecta: Lepidoptera: Hepialoidea). Fauna N. Z. 2019, 78. Available online: https://www.biotaxa.org/fnz/article/view/fnz.78 (accessed on 21 September 2021).
- Heppner, J.; Balcázar, L.M.A.; Wang, H.Y. Larval morphology of Ogygioses caliginosa from Taiwan (Lepidoptera: Palaeosetidae). Trop. Lep. 1995, 6, 149–154. [Google Scholar]
- Grehan, J. Larval feeding habits of the Hepialidae (Lepidoptera). J. Nat. Hist. 1989, 23, 803–824. [Google Scholar] [CrossRef]
- Ahola, M.; Davis, D.; Itämies, J.; Leinonen, R.; Mutanen, M. Description of immature stages of Nemophora bellela (Walker, 1863) (Lepidoptera: Adelidae). Entomol. Fenn. 2017, 28, 49–56. [Google Scholar] [CrossRef] [Green Version]
- Raman, A.; Suryanarayanan, T.S. Fungus–plant interaction influences plant-feeding insects. Fungal Ecol. 2017, 29, 123–132. [Google Scholar] [CrossRef]
- Jaworski, T. Diversity of saproxylic Lepidoptera. In Saproxylic Insects; Zoological Monographs; Ulyshen, M., Ed.; Springer: Cham, Switzerland, 2018; pp. 319–338. [Google Scholar]
- Metz, M.A.; Davis, D.R.; Davis, M.M. A new species of Niditinea (Tineidae: Tineinae) with a preference for bird nests, and the known larval habitats of the species in the United States. Proc. Entomol. Soc. Wash. 2018, 120, 153–166. [Google Scholar] [CrossRef]
- Bourgogne, J. Observations sur l’instinct des chenilles de Psychidae [Lep.]. Bull. Soc. Entomol. France 1949, 54, 49–52. [Google Scholar] [CrossRef]
- Heppner, J.B. Old World Spiny-Winged Moths (Lepidoptera: Eriocottidae). In Encyclopedia of Entomology; Capinera, J.L., Ed.; Springer: Dordrecht, The Netherlands, 2008; p. 1358. [Google Scholar]
- Scott, J.K.; Sagliocco, J.L. Host-specificity of a root borer, Bembecia chrysidiformis [Lep.: Sesiidae], a potential control agent for Rumex sp. [Polygonaceae] in Australia. Entomophaga 1991, 36, 235–244. [Google Scholar] [CrossRef]
- Bruzzese, D.J.; Wagner, D.L.; Harrison, T.; Jogesh, T.; Overson, R.P.; Wickett, N.J.; Skogen, K.A. Diversification in the microlepidopteran genus Mompha (Lepidoptera: Gelechioidea: Momphidae) is explained more by tissue specificity than host plant family. BioRxiv 2018. [Google Scholar] [CrossRef] [Green Version]
- Kirichenko, N.; Triberti, P.; Akulov, E.; Ponomarenko, M.; Gorokhova, S.; Sheiko, V.; Lopez-Vaamonde, C. Exploring species diversity and host plant associations of leaf-mining micromoths (Lepidoptera: Gracillariidae) in the Russian Far East using DNA barcoding. Zootaxa 2019, 4652, 1–55. [Google Scholar] [CrossRef]
- Sruoga, V.; Kaila, L.; Rocienė, A. The Elachistinae (Lepidoptera: Gelechioidea, Elachistidae) of Thailand, with description of eight new species. Eur. J. Taxon. 2019, 574, 1–34. [Google Scholar] [CrossRef]
- Pabis, K. What is a moth doing under water? Ecology of aquatic and semi-aquatic Lepidoptera. Knowl. Manag. Aquat. Ecosyst. 2018, 419, 42–128. [Google Scholar] [CrossRef] [Green Version]
- Tooker, J.F.; Giron, D. The evolution of endophagy in herbivorous insects. Front. Plant Sci. 2020, 11, 581816. [Google Scholar]
- Léger, T.; Mally, R.; Neinhuis, C.; Nuss, M. Refining the phylogeny of Crambidae with complete sampling of subfamilies (Lepidoptera, Pyraloidea). Zool. Scr. 2021, 50, 84–99. [Google Scholar] [CrossRef]
- Montgomery, S. Carnivorous caterpillars: The behaviour, biogeography and conservation of Eupithecia (Lepidoptera: Geometridae) in the Hawaiian islands. Geojournal 1983, 7, 549–556. [Google Scholar] [CrossRef]
- Bodner, F.; Brehm, G.; Homeier, J.; Strutzenberger, P.; Fiedler, K. Caterpillars and host plant records for 59 species of Geometridae (Lepidoptera) from a montane rainforest in southern Ecuador. J. Insect Sci. 2010, 10, 1–22. [Google Scholar] [CrossRef] [PubMed]
- Strutzenberger, P.; Brehm, G.; Gottsberger, B.; Bodner, F.; Seifert, C.L.; Fiedler, K. Diversification rates, host plant shifts and an updated molecular phylogeny of Andean Eois moths (Lepidoptera: Geometridae). PLoS ONE 2017, 12, e0188430. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Basso, A.; Negrisolo, E.; Zilli, A.; Battisti, A.; Cerretti, P. A total evidence phylogeny for the processionary moths of the genus Thaumetopoea (Lepidoptera: Notodontidae: Thaumetopoeinae). Cladistics 2017, 33, 557–573. [Google Scholar] [CrossRef] [PubMed]
- Narem, D.M.; Meyer, M.H. Native prairie graminoid host plants of Minnesota and associated Lepidoptera: A literature review. J. Lepidop. Soc. 2017, 71, 225–235. [Google Scholar] [CrossRef]
- Sedighi, L.; Ranjbar Aghdam, H.; Imani, S.; Shojai, M. Age-stage two-sex life table analysis of Sesamia nonagrioides (Lep.: Noctuidae) reared on different host plants. Arch. Phytopathol. Pflanzenschutz 2017, 50, 438–453. [Google Scholar] [CrossRef]
- Fetzner, J.W., Jr.; Fetzner, J.J. Notes on the Life History of the Delicate Silver Y, Autographa pseudogamma (Grote, 1875) (Lepidoptera: Noctuoidea: Noctuidae: Plusiinae). Ann. Carnegie Mus. 2018, 85, 5–23. [Google Scholar] [CrossRef]
- De Freina, J.J.; Mecenero, S.; Morton, A. Notes on the life history of Epitoxis namaqua de Freina & Mey, 2011 (Lepidoptera: Erebidae: Arctiinae: Syntomini). Metamorphosis 2020, 31, 15–19. [Google Scholar]
- Husain, A.; Hasan, W. New record of Common Owlet Moth Spirama helicina (Hubner, 1831) (Lepidoptera: Erebidae: Catocalinae) from Aligarh (Uttar Pradesh) with systematic account, distribution, host plants and biological control. Int. J. Agric. Appl. Sci. 2020, 1, 37–41. [Google Scholar] [CrossRef]
- Goldstein, P.Z.; Janzen, D.H.; Hallwachs, W. A Novel Origin of Pteridivory among the New World Noctuoidea: Fern-Feeding “Litter Moths” (Erebidae: Herminiinae). Proc. Entomol. Soc. Wash. 2021, 123, 314–333. [Google Scholar] [CrossRef]
- Salem, A. Revision of Lepidoptera of Egypt, Superfamily Noctuoidea Part II: Erebidae, Nolidae and Euteliidae. Egypt. Acad. J. Biol. Sci. 2021, 14, 59–142. [Google Scholar] [CrossRef]
- Heppner, J.B. Douglas Moths (Lepidoptera: Douglasiidae). In Encyclopedia of Entomology; Capinera, J.L., Ed.; Springer: Dordrecht, The Netherlands, 2008; p. 1241. [Google Scholar]
- Muttkowski, R.A. Studies on the blood of insects III. The coagulation and clotting of insect blood. Bull. Brooklyn Entomol. Soc. 1924, 19, 128–144. [Google Scholar]
- He, F.; Sun, S.; Tan, H.; Qin, C.; Ji, S.; Li, X.; Zhang, J.; Jiang, X. Chlorantraniliprole against the black cutworm Agrotis ipsilon (Lepidoptera: Noctuidae): From biochemical/physiological to demographic responses. Sci. Rep. 2019, 9, 10328. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Melguizo-Ruiz, N.; Jiménez-Navarro, G.; De Mas, E.; Pato, J.; Scheu, S.; Austin, A.T.; Wise, D.H.; Moya-Laraño, J. Field exclusion of large soil predators impacts lower trophic levels and decreases leaf-litter decomposition in dry forests. J. Anim. Ecol. 2020, 89, 334–346. [Google Scholar] [CrossRef] [PubMed]
- Werner, W.J.; Sanderman, J.; Melillo, J.M. Decreased soil organic matter in a long term soil warming experiment lowers soil water holding capacity and affects soil thermal and hydrological buffering. JGR Biogeosci. 2020, 125, e2019JG005158. [Google Scholar] [CrossRef]
- Reddy, R.; Jiang, Q.; Aramwit, P.; Reddy, N. Litter to leaf: The unexplored potential of silk byproducts. Trends Biotechnol. 2021, 39, 706–718. [Google Scholar] [CrossRef]
- Deharveng, L.; Dalens, H.; Drugmand, D.; Simon-Benito, J.C.; Da Gama, M.M.; Sousa, P.; Gers, C.; Bedos, A. Endemism mapping and biodiversity conservation in western Europe: An Arthropod perspective. Belg. J. Entomol. 2000, 2, 59–75. [Google Scholar]
- Barragán-Fonseca, K.Y.; Nurfikari, A.; Van De Zande, E.M.; Wantulla, M.; Van Loon, J.J.; De Boer, W.; Dicke, M. Insect frass and exuviae to promote plant growth and health. Trends Plant Sci. 2022, 27, 646–654. [Google Scholar] [CrossRef]
Figure 1.
Phylogeny of Lepidoptera (Separated into “A”, “B” for readability) modified from [37] and the main environmental uses of caterpillars (left) and pupae (right). For each category, the intensity of the colour is related to the number of species using a particular resource (Deep colour: majority of species, pastel colour: less than 40% of species). Green: leaf use, yellow: stems or wood, orange: litter, purple/pink: fungi, brown: soil, red: carnivores or nymphs in anthills, and blue: freshwater.
Figure 1.
Phylogeny of Lepidoptera (Separated into “A”, “B” for readability) modified from [37] and the main environmental uses of caterpillars (left) and pupae (right). For each category, the intensity of the colour is related to the number of species using a particular resource (Deep colour: majority of species, pastel colour: less than 40% of species). Green: leaf use, yellow: stems or wood, orange: litter, purple/pink: fungi, brown: soil, red: carnivores or nymphs in anthills, and blue: freshwater.

Figure 2.
Main changes in environmental or food use between caterpillars and pupae. The width of the arrows is relative to the number of switches. Green: leaf use, yellow: stems or wood, orange: litter, purple: fungi, brown: soil, red: carnivores or pupation in anthills, and blue: fresh water.
Figure 2.
Main changes in environmental or food use between caterpillars and pupae. The width of the arrows is relative to the number of switches. Green: leaf use, yellow: stems or wood, orange: litter, purple: fungi, brown: soil, red: carnivores or pupation in anthills, and blue: fresh water.

Figure 3.
Caterpillars; cladistic classification (see material and methods for parameters) and grouping of subdivisions of the phylogeny of [37] according to the occupation of various living environments. Green: leaf use, yellow: stems or wood, orange: litter, purple: fungi, brown: soil, red: carnivores, and blue: freshwater.
Figure 3.
Caterpillars; cladistic classification (see material and methods for parameters) and grouping of subdivisions of the phylogeny of [37] according to the occupation of various living environments. Green: leaf use, yellow: stems or wood, orange: litter, purple: fungi, brown: soil, red: carnivores, and blue: freshwater.

Figure 4.
Chrysalis; cladistic classification (see material and methods for parameters) and grouping of subdivisions of the phylogeny of [37] according to the occupation of various living environments. Green: leaf use, yellow: stems or wood, orange: litter, brown: soil, red: pupation in ant hills, and blue: freshwater.
Figure 4.
Chrysalis; cladistic classification (see material and methods for parameters) and grouping of subdivisions of the phylogeny of [37] according to the occupation of various living environments. Green: leaf use, yellow: stems or wood, orange: litter, brown: soil, red: pupation in ant hills, and blue: freshwater.


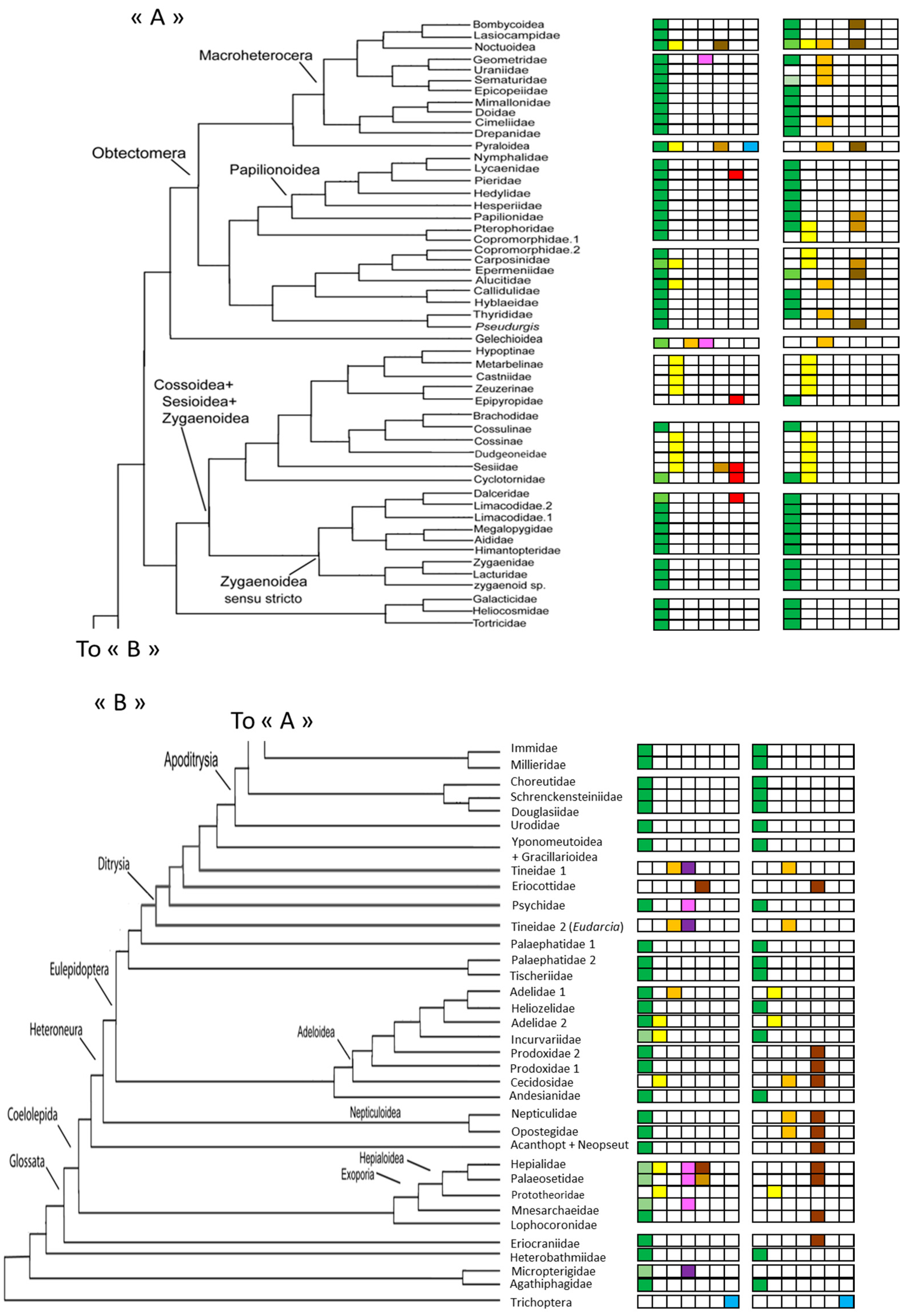{kind=link}
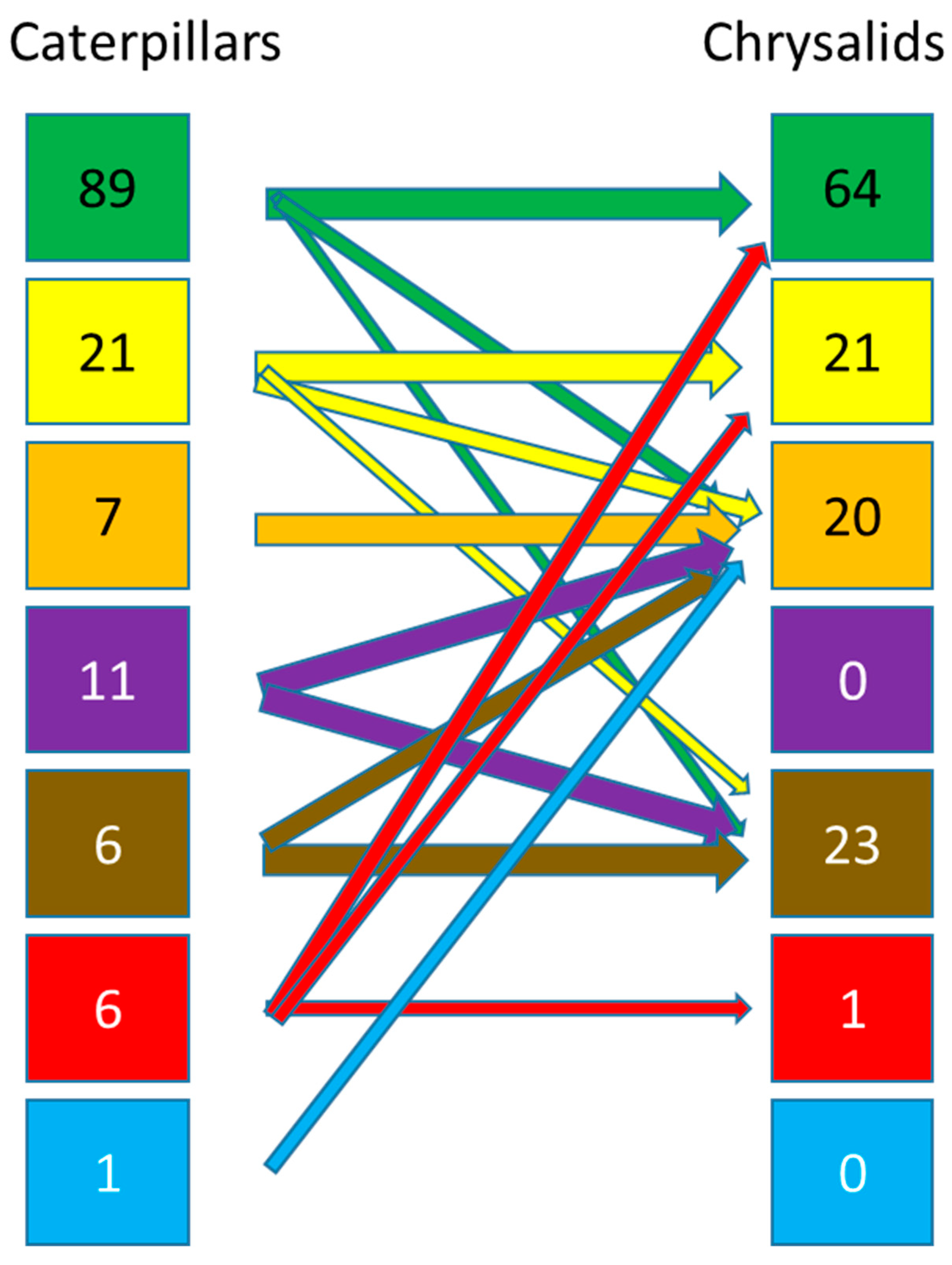{kind=link}
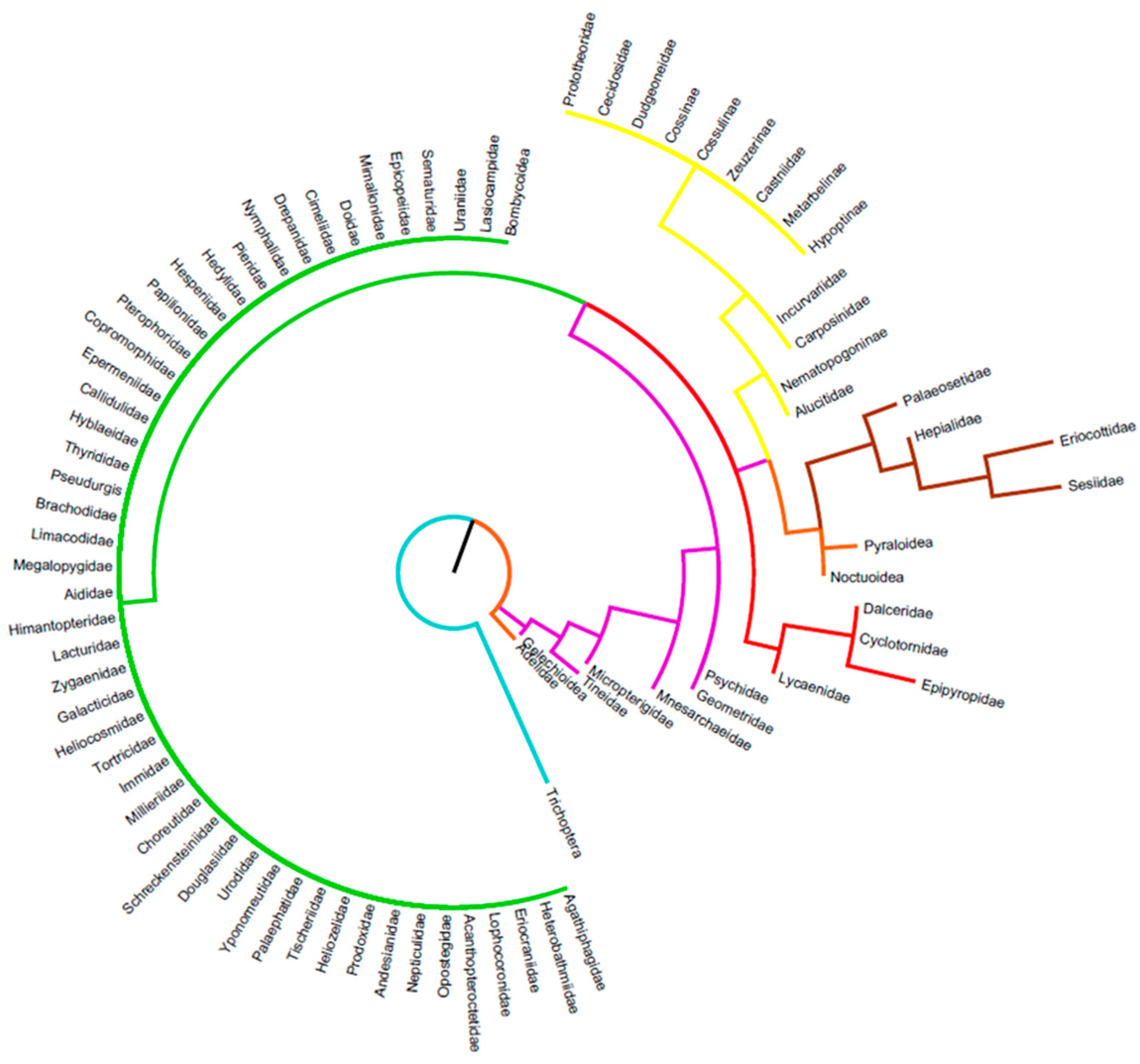{kind=link}
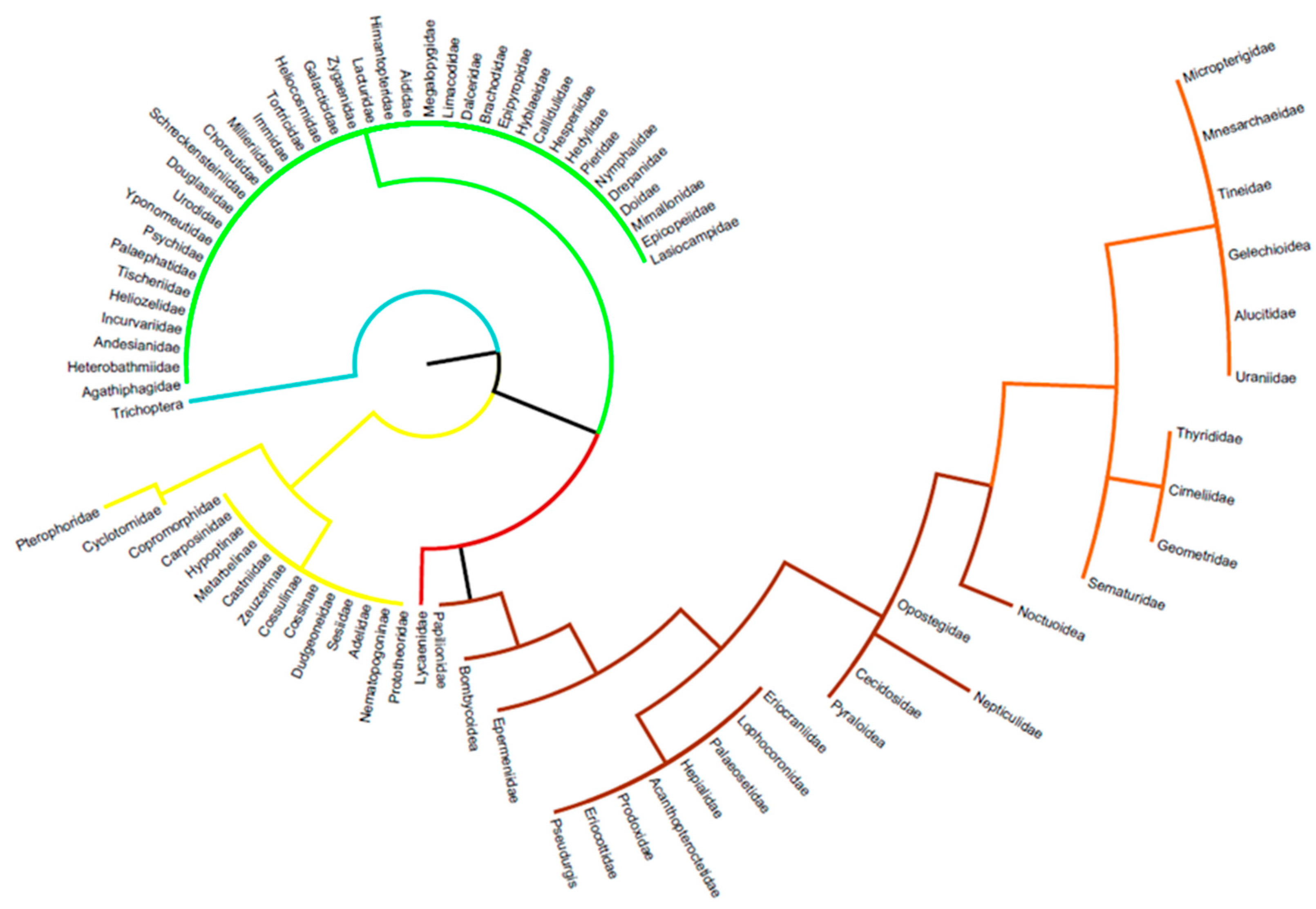{kind=link}
Table 1.
Rough estimates of the number of soil-related Lepidoptera species. For each subdivision used in this work, an estimate of the number of currently described species (i.e., based on 157,000 species) was made and only those subdivisions where at least one species has a relationship with soil are included (i.e., a total of 64,484 species = 41% of the total species). In each of the subdivisions where there is at least one soil-related species, an estimate of the number of species related to the soil is given. The results are presented separately for caterpillars and chrysalises. For caterpillars, those that feed on litter and fungi have been separated from those that feed on roots. Similarly, for chrysalises, a separation was made between species that nymph in the litter and those in the soil. Column 2: Number of species for each family with at least one species described to be related to soil. Column 3–5: Number of species feeding on litter or fungi and those feeding on roots + overall percentage per family of soil-using caterpillars. Column 6–8: Number of species nymphalizing in litter or soil + overall percentage by family of species pupating in the soil.
Table 1.
Rough estimates of the number of soil-related Lepidoptera species. For each subdivision used in this work, an estimate of the number of currently described species (i.e., based on 157,000 species) was made and only those subdivisions where at least one species has a relationship with soil are included (i.e., a total of 64,484 species = 41% of the total species). In each of the subdivisions where there is at least one soil-related species, an estimate of the number of species related to the soil is given. The results are presented separately for caterpillars and chrysalises. For caterpillars, those that feed on litter and fungi have been separated from those that feed on roots. Similarly, for chrysalises, a separation was made between species that nymph in the litter and those in the soil. Column 2: Number of species for each family with at least one species described to be related to soil. Column 3–5: Number of species feeding on litter or fungi and those feeding on roots + overall percentage per family of soil-using caterpillars. Column 6–8: Number of species nymphalizing in litter or soil + overall percentage by family of species pupating in the soil.
| Number of Species | Caterpillars | % per Family | Chrysalids | % per Family | |||
|---|---|---|---|---|---|---|---|
| Litter/Fungi | Roots | Litter | Soil | ||||
| Micropterigidae | 140 | 140 | 0 | 100 | 140 | 0 | 100 |
| Eriocraniidae | 29 | 0 | 0 | 0 | 0 | 29 | 100 |
| Lophocoronidae | 6 | 0 | 0 | 0 | 0 | 6 | 100 |
| Mnesarchaeidae | 14 | 14 | 0 | 100 | 14 | 0 | 100 |
| Palaeosetidae | 7 | 3 | 4 | 100 | 0 | 7 | 100 |
| Hepialidae | 400 | 0 | 200 | 50 | 0 | 200 | 50 |
| Acanthopteroctetidae | 5 | 0 | 0 | 0 | 5 | 0 | 100 |
| Opostegidae | 200 | 0 | 0 | 0 | 200 | 0 | 100 |
| Adelidae | 300 | 100 | 0 | 33 | 100 | 0 | 33 |
| Nepticulidae | 862 | 0 | 0 | 0 | 862 | 0 | 100 |
| Cecidosidae | 15 | 0 | 0 | 0 | 0 | 15 | 100 |
| Prodoxidae | 100 | 0 | 0 | 0 | 0 | 100 | 100 |
| Tineidae | 3000 | 1000 | 0 | 33 | 1000 | 500 | 50 |
| Psychidae | 1000 | 600 | 0 | 60 | 0 | 0 | 0 |
| Eriocottidae | 210 | 0 | 200 | 95 | 0 | 200 | 95 |
| Sesiidae | 1000 | 0 | 150 | 15 | 0 | 150 | 15 |
| Gelechioidea | 4750 | 1000 | 500 | 32 | 3000 | 0 | 63 |
| Pseudurgis | 10 | 0 | 0 | 0 | 0 | 10 | 100 |
| Thyrididae | 750 | 0 | 0 | 0 | 750 | 0 | 100 |
| Alucitidae | 250 | 0 | 0 | 0 | 250 | 0 | 100 |
| Epermeniidae | 100 | 0 | 0 | 0 | 0 | 100 | 100 |
| Carposinidae | 290 | 0 | 0 | 0 | 0 | 290 | 100 |
| Pterophoridae | 1300 | 0 | 0 | 0 | 0 | 700 | 54 |
| Papilionidea | 600 | 0 | 0 | 0 | 0 | 1 | 0.2 |
| Pyraloidea | 8000 | 1500 | 0 | 19 | 2500 | 1500 | 50 |
| Cimeliidae | 6 | 0 | 0 | 0 | 6 | 0 | 100 |
| Sematuridae | 40 | 0 | 0 | 0 | 30 | 10 | 100 |
| Uraniidae | 100 | 0 | 0 | 0 | 90 | 0 | 90 |
| Geometridae | 15,000 | 2000 | 0 | 13 | 6000 | 2000 | 53 |
| Noctuoidea | 20,000 | 3000 | 1500 | 22.5 | 1000 | 12,000 | 65 |
| Bombycoidea | 6000 | 0 | 0 | 0 | 0 | 1500 | 25 |
| Total species | 64,484 | 9357 | 2554 | 18.5 | 15,947 | 19,318 | 54.7 |
| base 157,000 species | 41% | 6% | 2% | 10% | 12% | ||
| 7.60% | 22.70% | ||||||
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2022 by the author. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Legal, L. “Lepidoptera Flies”, but Not Always…Interactions of Caterpillars and Chrysalis with Soil. Diversity 2023, 15, 27. https://doi.org/10.3390/d15010027
AMA Style
Legal L. “Lepidoptera Flies”, but Not Always…Interactions of Caterpillars and Chrysalis with Soil. Diversity. 2023; 15(1):27. https://doi.org/10.3390/d15010027
Chicago/Turabian StyleLegal, Luc. 2023. "“Lepidoptera Flies”, but Not Always…Interactions of Caterpillars and Chrysalis with Soil" Diversity 15, no. 1: 27. https://doi.org/10.3390/d15010027
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.