

The Species Diversity Assessment of Azygia Looss, 1899 (Digenea: Azygiidae) from the Volga, Ob, and Artyomovka Rivers Basins (Russia), with Description of A. sibirica n. sp.
 ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Sampling
2.2. Morphological Examination
2.3. Multivariate Analysis
2.4. DNA Extraction, Amplification, and Sequencing
2.5. Alignment and Phylogenetic Analyses
3. Results
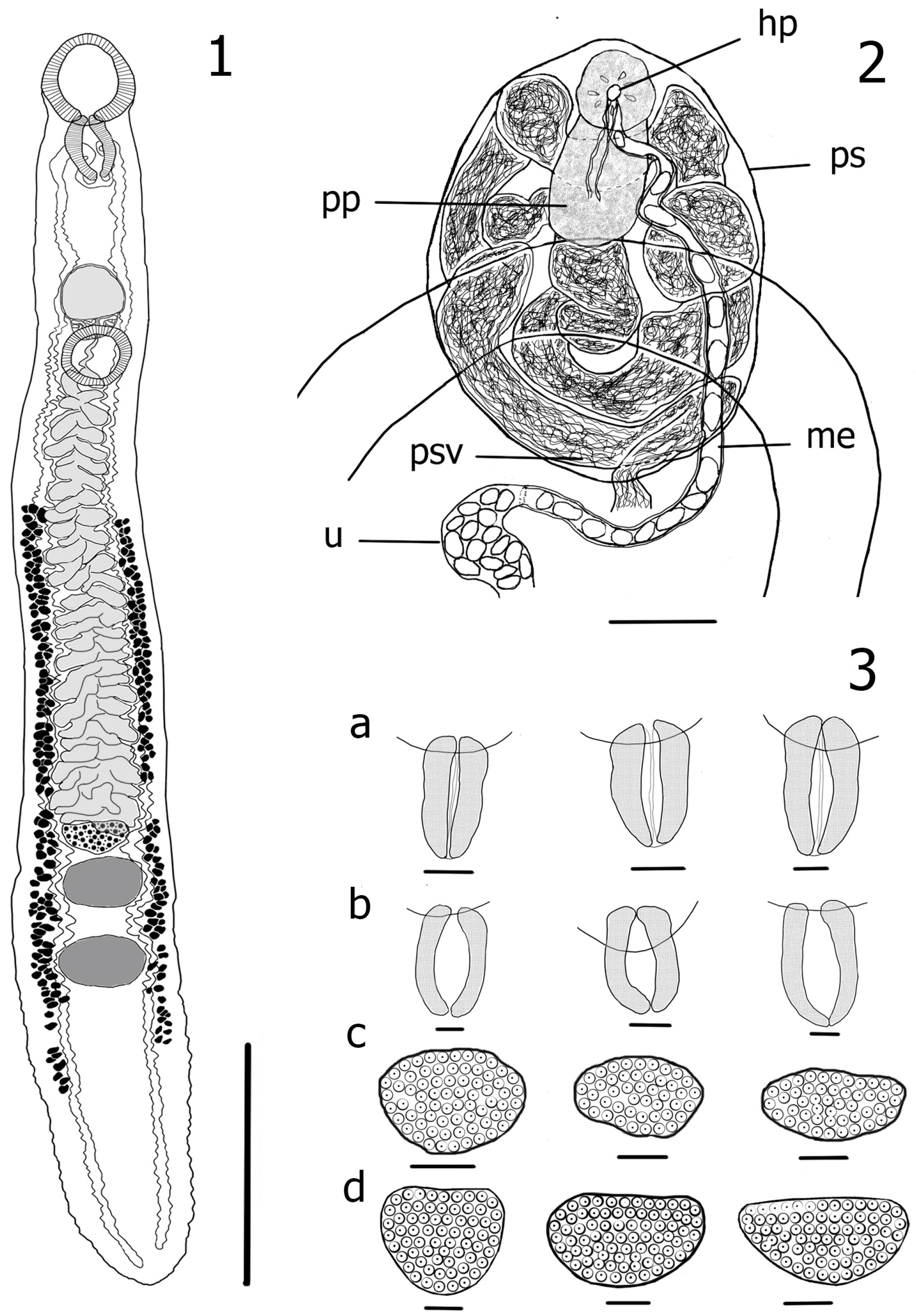
3.1. Redescription of Azygia Lucii (Müller, 1776) (Figure 1)
Taxonomic Summary

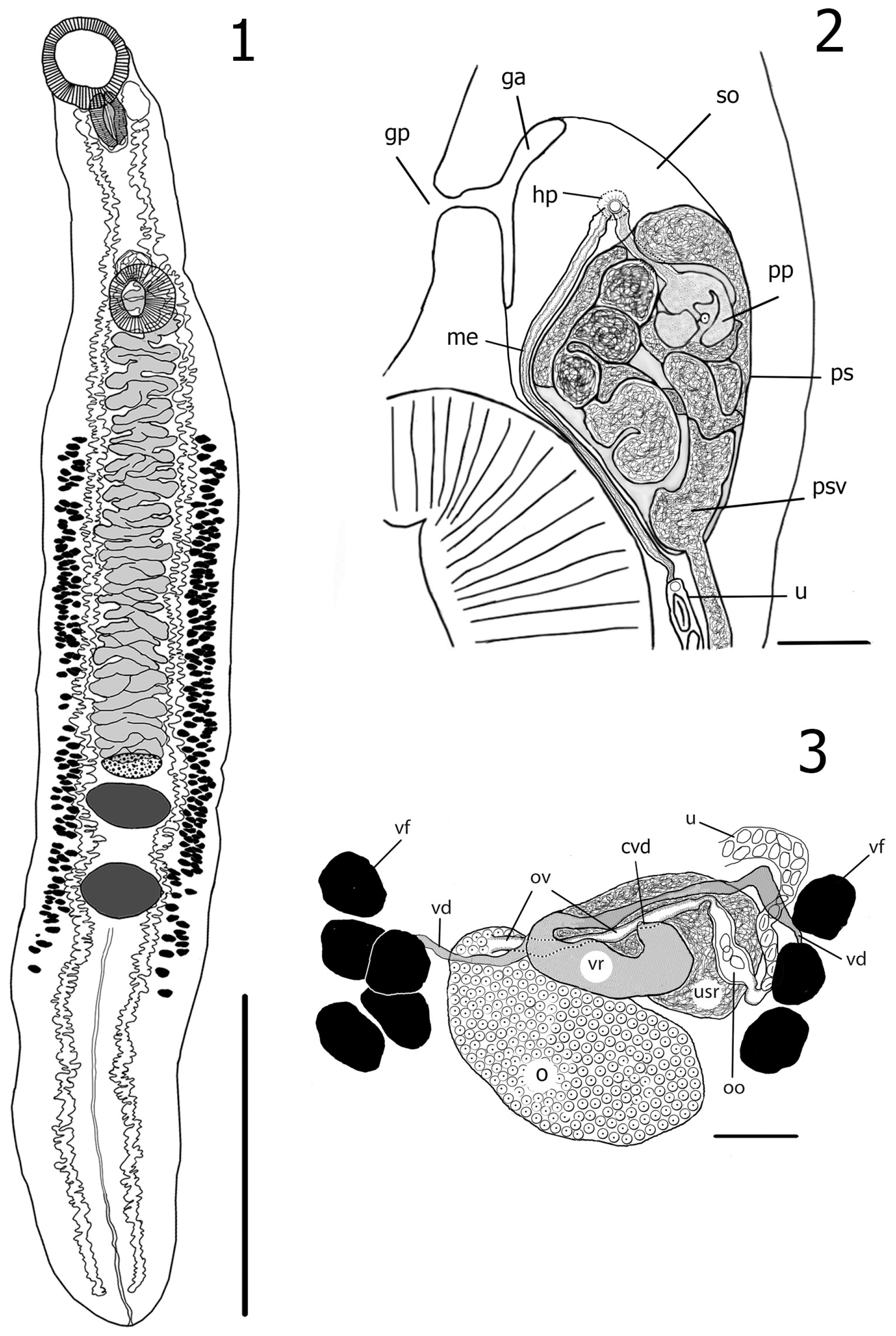
3.2. Description of Azygia Sibirica Vainutis, Voronova et Zhokhov, n. sp. (Figure 2)
Taxonomic Summary

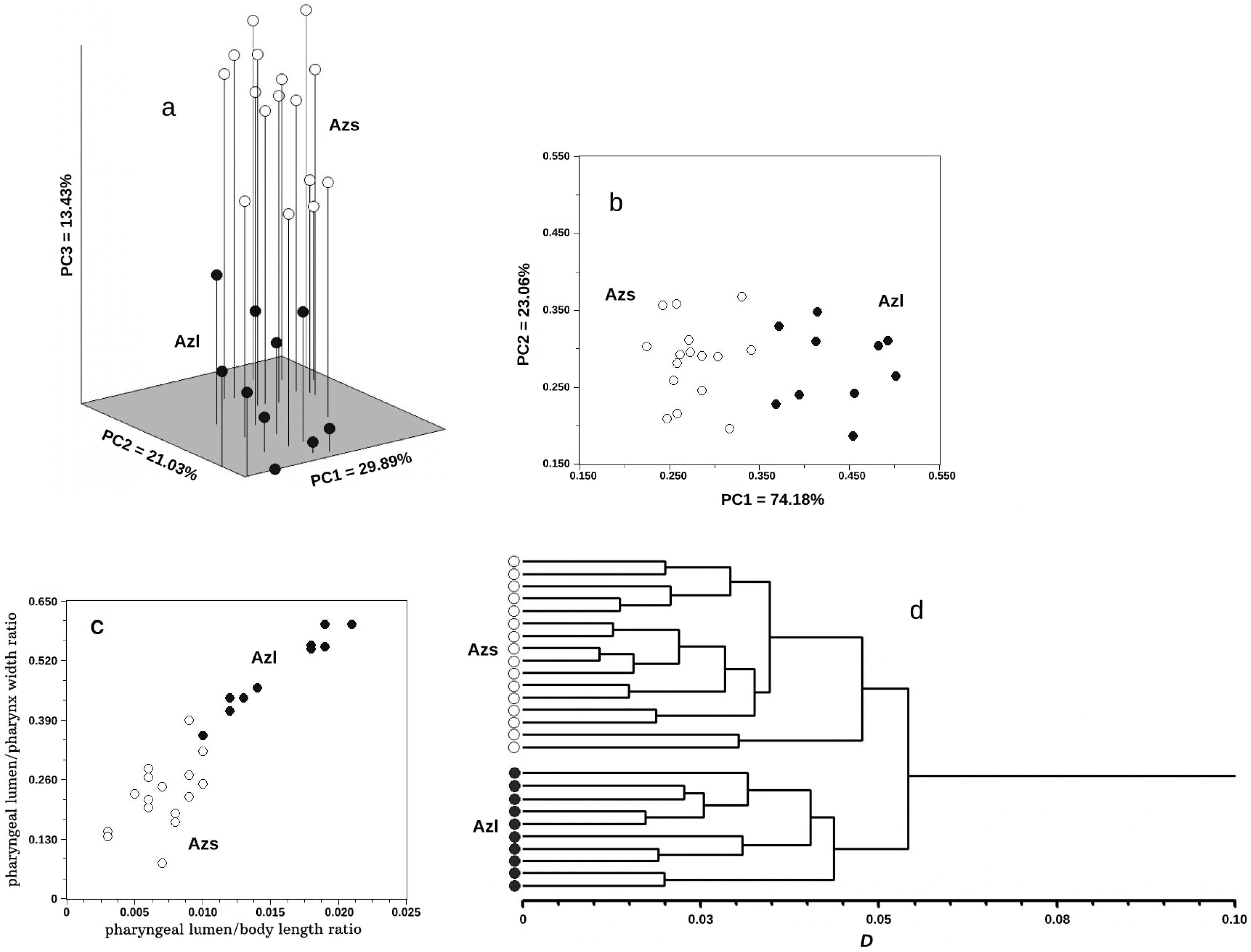
3.3. Multivariate Analysis
3.4. Molecular Genetic Analyses
3.4.1. 28S rRNA Gene
3.4.2. cox1 mtDNA Gene
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Conflicts of Interest
References
- Jadhav, S.; Humbe, A.; Padwal, N. Histopathological changes of the intestine of the freshwater fish, Channa gachua (Hamilton, 1822) due to infection of metacercaria of trematode parasites. Int. J. Fish Aquat. Stud. 2019, 7, 279–281. [Google Scholar]
- Skrjabin, K.I.; Guschanskaja, L.K. Family Azygiidae Odhner, 1911. In Trematodes of Animals and Man. Principles of Trematodology; Skrjabin, K.I., Ed.; Akademiya Nauk Press: Moscow, Russia, 1958; Volume 14, pp. 671–788. (In Russian) [Google Scholar]
- Gibson, D.I. Superfamily Azygioidea Lühe, 1909. In Keys to the Trematoda; Gibson, D.I., Jones, A., Bray, R.A., Eds.; CAB International: Wallingford, England, 2002; Volume 1, pp. 19–24. [Google Scholar] [CrossRef]
- Zhang, S.; Peng, J.; Chen, J.; Xu, X.; Zhi, T.; Brown, C.L.; Yang, T. Two Native Trematodes, Azygia sinipercae and Pseudophyllodistomum anguilae, Colonized in Invasive Redbelly Tilapia Tilapia zillii (Cichlidae) in South China and Their Ecology; Life Science Academy, Sun Yat-Sen University: Guangdong, China, 2018; Unpublished. [Google Scholar]
- Calhoun, D.M.; Curran, S.S.; Pulis, E.E.; Provaznik, J.M.; Franks, J.S. Hirudinella ventricosa (Pallas, 1774) Baird, 1853 represents a species complex based on ribosomal DNA. Syst. Parasitol. 2013, 86, 197–208. [Google Scholar] [CrossRef]
- Van Steenkiste, N.; Locke, S.A.; Castelin, M.; Marcogliese, D.J.; Abbott, C.L. New primers for DNA barcoding of digeneans and cestodes (Platyhelminthes). Mol. Ecol. Resour. 2015, 15, 945–952. [Google Scholar] [CrossRef]
- Moszczynska, A.; Locke, S.A.; McLaughlin, J.D.; Marcogliese, D.J.; Crease, T.J. Development of primers for the mitochondrial cytochrome c oxidase I gene in digenetic trematodes (Platyhelminthes) illustrates the challenge of barcoding parasitic helminths. Mol. Ecol. Resour. 2009, 9 (Suppl. S1), 75–82. [Google Scholar] [CrossRef]
- Wu, Y.A.; Gao, J.W.; Cheng, X.F.; Xie, M.; Yuan, X.P.; Liu, D.; Song, R. Characterization and comparative analysis of the complete mitochondrial genome of Azygia hwangtsiyui Tsin, 1933 (Digenea), the first for a member of the family Azygiidae. ZooKeys 2020, 945, 1–16. [Google Scholar] [CrossRef]
- Linstow, O.F.B. Zwei neue Distomum aus Lucioperca sandra der Wolga. Ezegodnik zool. Mus. imp. Akad. Nauk. 1907, 12, 201–202. (In German) [Google Scholar]
- Zmejev, G.J. Flukes and tapeworms of fish of the Amur River. Parazitol. Sb. Zool. Inst. Akad. Nauk SSSR 1936, 6, 408–435. (In Russian) [Google Scholar]
- Shcherbina, T.V.; Frolova, E.N. Distribution of Azygia mirabilis (Trematoda, Azygiidae). Parazitologiia 1980, 14, 112–117. (In Russian) [Google Scholar]
- Zhokhov, A.E.; Tyutin, A.V. Parasitic fauna of fishes in the conditions of acidification of lakes. Structure and functioning of acid lake ecosystems. Tr. Inst. Biol. Vnutr. Vod Ross. Akad. Nauk. 1994, 70, 186–201. (In Russian) [Google Scholar]
- Ermolenko, A.V.; Besprozvannykh, V.V.; Shedko, S.V. The Fauna of Parasites of Salmonids (Salmonidae, Salmoniformes) of Primorsky Krai; Dalnauka: Vladivostok, Russia, 1998; p. 89. [Google Scholar]
- Besprozvannykh, V.V. Life cycles of the trematode species Azygia hwangtsiytii and A. robusta (Azygiidae) in Primorsky Territory. Parazitologiia 2005, 39, 278–284. (In Russian) [Google Scholar]
- Vainutis, K.S.; Voronova, A.N. Study of helminth fauna in the Bolotnaya River and floodplain lake near the Solovey-Klyuch village (Nadezhdinsky district, Primorsky region). Vestn. Far East Branch Russ. Acad. Sci. 2021, 1, 94–101. (In Russian) [Google Scholar] [CrossRef]
- Müller, O.F. Zoologiae Danicae Prodromus: Seu Animalium Daniae et Norvegiae Indigenarum; Characteres, Nomina, et Synonyma Imprimis Popularium; Typis Hallageriis: Copenhagen, Denmark, 1776; p. 322, (In Latin). [Google Scholar] [CrossRef] [Green Version]
- Pavlov, N.N. The problem of parasitology in fish in the River Ob. Trudy Daltnevostoch. Pod. In-ta 1931, 5, 35–46. (In Russian) [Google Scholar]
- Dorovskikh, G.N. Results of the study of fishes’ parasites in river basins of the north-east of the European part of Russia. Trematodes (Trematoda). Parazitologiya 1997, 31, 551–563. (In Russian) [Google Scholar]
- Molodozhnikova, N.M.; Zhokhov, A.E. The taxonomic diversity of the parasites of agnathans and fishes in the Volga basin. III. Aspidogastrea Trematoda. Parazitol. 2007, 41, 28–54. (In Russian) [Google Scholar]
- Romanova, N.N.; Golovina, N.A.; Vishtorskaya, N.A.; Golovin, P.P. Fauna of parasite of cyprinids and percoids in the reservoirs of the Moscow Canal. Russ. J. Parasitol. 2021, 15, 32–47. [Google Scholar] [CrossRef]
- Strelkov, Y.A. Trematodes of fish from Amur River basin. Parasitol. Sb. ZIN AN SSSR 1971, 25, 41–76. (In Russian) [Google Scholar]
- Truett, G.E. Preparation of genomic DNA from animal tissues. In The DNA book: Protocols and Procedures for the Modern Molecular Biology Laboratory; Kieleczawa, J., Ed.; Jones and Bartlett Publisher: Sudbury, ON, Canada, 2006; pp. 33–46. [Google Scholar]
- Lockyer, A.E.; Olson, P.D.; Littlewood, D.T.J. Utility of complete large and small subunit rRNA genes in resolving the phylogeny of the Neodermata (Platyhelminthes): Implications and a review of the cercomer theory. Biol. J. Linn. Soc. 2003, 78, 155–171. [Google Scholar] [CrossRef] [Green Version]
- Bowles, J.; Blair, D.; McManus, D.P. A molecular phylogeny of the human schistosomes. Mol. Phylog. Evol. 1995, 4, 103–109. [Google Scholar] [CrossRef]
- Miura, O.; Kuris, A.M.; Torchin, M.E.; Hechinger, R.F.; Dunham, E.J.; Chiba, S. Molecular-genetic analyses reveal cryptic species of trematodes in the intertidal gastropod, Batillaria cumingi (Crosse). Int. J. Parasitol. 2005, 35, 793–801. [Google Scholar] [CrossRef]
- Kumar, S.; Stecher, G.; Li, M.; Knyaz, C.; Tamura, K. MEGA X: Molecular Evolutionary Genetics Analysis across Computing Platforms. Mol. Biol. Evol. 2018, 35, 1547–1549. [Google Scholar] [CrossRef]
- Huelsenbeck, J.P.; Ronquist, F.; Nielsen, R.; Bollback, J.P. Bayesian inference of phylogeny and its impact on evolutionary biology. Science 2001, 294, 2310–2314. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Darriba, D.; Taboada, G.L.; Doallo, R.; Posada, D. jModelTest 2: More models, new heuristics and parallel computing. Nat. Methods 2012, 9, 772. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Puillandre, N.; Lambert, A.; Brouillet, S.; Achaz, G. ABGD, Automatic Barcode Gap Discovery for primary species delimitation. Mol. Ecol. 2012, 21, 1864–1877. [Google Scholar] [CrossRef]
- Fujisawa, T.; Barraclough, T.G. Delimiting species using single-locus data and the generalized mixed yule coalescent approach: A revised method and evaluation on simulated data sets. Syst. Biol. 2013, 62, 707–724. [Google Scholar] [CrossRef] [Green Version]
- Zhang, J.; Kapli, P.; Pavlidis, P.; Stamatakis, A. A General Species Delimitation Method with Applications to Phylogenetic Placements. Bioinformatics 2013, 29, 2869–2876. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Frolova, E.N.; Shcherbina, T.V. New species in genus Azygia Looss, 1899 (Trematoda, Azygiidae). Parazitologiia 1975, 9, 489–493. (In Russian) [Google Scholar] [PubMed]
- Looss, A. Die Distomen unserer Fische und Frösche. Neue Untersuchungen über Bau und Entwickelung des Distomenkörpers. Bibl. Zool. Stuttg. 1894, 16, 296. (In German) [Google Scholar]
- Markevich, A.P. Parasite Fauna of Freshwater Fishes of the Ukrainian SSR; Publishing House of the Academy of Sciences of the Ukrainian SSR: Kyiv, Ukraine, 1951; p. 375. (In Russian) [Google Scholar]
- Stunkard, H.W. The Morphology and Life-History of the Digenetic Trematode, Azygia sebago Ward, 1910. Biol. Bull. 1956, 111, 248–268. [Google Scholar] [CrossRef]
- Odening, K. Cercaria splendens Szidat, the larva of the trematode Azygia lucii Looss. Parazitologiia 1975, 9, 232–236. (In Russian) [Google Scholar]
- International Commission on Zoological Nomenclature. Article 23. Principle of Priority. In International Code of Zoological Nomenclature., 4th ed.; Adopted by the International Union of Biological Sciences; Publishing House KMK: Moscow, Russia, 2004; p. 64. (In Russian) [Google Scholar]
- Chen, C.L. (Ed.) An Illustrated Guide to the Fish Diseases and Causative Pathogenic Fauna and Flora in the Hubei Province; Science Press: Beijing, China, 1973. (In Chinese) [Google Scholar]
- Vainutis, K.S.; Voronova, A.N.; Urabe, M. Systematics of Crepidostomum species from the Russian Far East and northern Japan, with description of a new species and validation of the genus Stephanophiala. Parasitol. Int. 2021, 84, 102412. [Google Scholar] [CrossRef]
- Vainutis, K.S.; Voronova, A.N.; Duscher, G.G.; Shchelkanov, E.M.; Shchelkanov, M.Y. Origins, phylogenetic relationships and host-parasite interactions of Troglotrematoidea since the cretaceous. Infect. Genet. Evol. 2022, 101, 105274. [Google Scholar] [CrossRef] [PubMed]
- Korlyakov, K.A.; Nohrin, D.Y. Tendencies of occurrence invasion of the corridor Volga-Ob. Vestn. SMUS Chelyabinskoy Obl. 2014, 2, 19–38. (In Russian) [Google Scholar]
- Skog, A.; Vøllestad, L.A.; Stenseth, N.C.; Kasumyan, A.; Jakobsen, K.S. Circumpolar phylogeography of the northern pike (Esox lucius) and its relationship to the Amur pike (E. reichertii). Front. Zool. 2014, 11, 67. [Google Scholar] [CrossRef] [Green Version]
- Takács, P.; Bánó, B.; Czeglédi, I.; Erős, T.; Ferincz, Á.; Gál, B.; Bánó-Kern, B.; Kovács, B.; Nagy, A.A.; Nyeste, K.; et al. The mixed phylogenetic origin of northern pike (Esox lucius Linnaeus 1758) populations in the Middle Danubian drainage. BMC Zool. 2022, 7, 28. [Google Scholar] [CrossRef]
- Blair, P.; Franklin, H.; Kelner, N.; Ke-Lind, P.; Paulmier, M.; Shrestha, M. Targeted rDNA sequence determination from geographically isolated populations of Proterometra macrostoma (Trematoda: Azygiidae). Proc. Indian Acad. Sci. 2018, 127, 82–88. [Google Scholar]
- Olson, P.D.; Cribb, T.H.; Tkach, V.V.; Bray, R.A.; Littlewood, D.T.J. Phylogeny and classification of the Digenea (Platyhelminthes: Trematoda). Int. J. Parasitol. 2003, 33, 733–755. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Parasite Species | Host | Locality | Sequence Data | Reference | |
|---|---|---|---|---|---|
| 28S rRNA | cox1 mtDNA | ||||
| Azygia lucii | Esox lucius | Russia, Yaroslavl region, Rybinsk Reservoir, Volga River basin (near Borok) | OP794033-OP794035 | OP804437-OP804443 | This study |
| Russia, Tyumen region, Ob River (near Surgut) | OP794036 | OP804444 | |||
| Russia, Tyumen region, Ob River (near Priobye) | OP794037-OP794039 | OP804454-OP804468 | |||
| Russia, Tyumen region, Ob River (near Khanty-Mansiysk) | OP794040-OP794042 | OP804445-OP804453 | |||
| Azygia sibirica n. sp. | Esox lucius | Russia, Tyumen region, Ob River (near Surgut) | OP794043-OP794046 | OP804429-OP804435 | |
| Russia, Tyumen region, Ob River (near Priobye) | OP794047 | OP804436 | |||
| Azygia hwangtsiyui | Perccottus glenii | Russia, Primorsky region, Artyomovka River basin, Bolotnaya River | OP794027-OP794029 | OP804412-OP804428 | |
| Russia, Primorsky region, Artyomovka River basin, Ivnyanka River | OP794030-OP794032 | OP804409-OP804411 | |||
| Channa argus | China, Hunan province, Yueyang, Dongting Lake | - | NC_049873, MN844889 | [8] | |
| Azygia sinipercae | Siniperca scherzeri | China, reservoir | MH777021 | - | [4] |
| Tilapia zillii | MG976845 | - | |||
| Azygia longa | Esox niger | USA, Mississippi, Jackson County, Pascagoula River | KC985234 | - | [5] |
| Proterometra sp. | Lepomis macrochirus | USA, Florida, Pitt Spring | KC985237 | - | |
| Proterometra macrostoma | Pleurocera semicarinata | USA, Indiana, Wayne County, Clear Creek | MF927953, MF927954 | - | [22] |
| Otodistomum cestoides | Raja montagui | UK, North Sea | AY222187 | - | [23] |
| Character | EV1 | EV2 | EV3 |
|---|---|---|---|
| Body width/BL * | 0.070 | 0.234 | 0.181 |
| Forebody length/BL | −0.186 | 0.285 | 0.089 |
| Forebody width/maximum body width | 0.077 | 0.048 | −0.329 |
| Pharynx width/length ratio | 0.180 | −0.094 | −0.177 |
| Pharyngeal lumen/pharynx width ratio | 0.196 | −0.147 | −0.371 |
| Ovary length/width ratio | −0.031 | 0.056 | 0.139 |
| Post-testicular space/BL | 0.148 | −0.227 | −0.092 |
| Forebody length/BL | 0.123 | 0.285 | −0.130 |
| Oral sucker length/BL | 0.192 | 0.336 | −0.014 |
| Oral sucker width/BL | 0.180 | 0.327 | 0.125 |
| Pharynx length/BL | 0.185 | 0.252 | −0.150 |
| Pharynx width/BL | 0.270 | 0.116 | −0.240 |
| Pharyngeal lumen/BL | 0.248 | −0.083 | −0.338 |
| Ventral sucker length/BL | 0.220 | 0.331 | −0.006 |
| Ventral sucker width/BL | 0.191 | 0.342 | 0.053 |
| Ovary length/BL | 0.278 | −0.098 | 0.209 |
| Ovary width/BL | 0.320 | −0.142 | 0.138 |
| Anterior testes length/BL | 0.251 | −0.169 | 0.212 |
| Anterior testes width/BL | 0.279 | −0.090 | 0.277 |
| Posterior testes length/BL | 0.253 | −0.220 | 0.268 |
| Posterior testes width/BL | 0.283 | −0.025 | 0.294 |
| Distance of ovary from anterior testis/BL | −0.211 | 0.057 | 0.264 |
| Distance of vitellarium from posterior extremity/BL | 0.117 | −0.214 | −0.107 |
| Eigenvalues | 6.880 | 4.840 | 3.090 |
| Explained variance, % | 29.890 | 21.030 | 13.430 |
| 1 | 2 | 3 | 4 | 5 | 6 | 7 | 8 | 9 | ||
| 1 | Azygia lucii Volga | 0.001 | 0.003 | 0.006 | 0.006 | 0.005 | 0.007 | 0.007 | 0.009 | |
| 2 | Azygia lucii Ob | 0.156 | 0.003 | 0.006 | 0.006 | 0.005 | 0.007 | 0.007 | 0.009 | |
| 3 | Azygia sibirica n. sp. | 1.405 | 1.249 | 0.006 | 0.006 | 0.005 | 0.007 | 0.006 | 0.009 | |
| 4 | Azygia hwangtsiyui | 4.619 | 4.463 | 4.463 | 0.001 | 0.006 | 0.007 | 0.007 | 0.009 | |
| 5 | Azygia sinipercae | 4.528 | 4.372 | 4.372 | 0.091 | 0.006 | 0.007 | 0.007 | 0.009 | |
| 6 | Azygia longa | 4.059 | 3.903 | 3.669 | 4.853 | 4.762 | 0.006 | 0.006 | 0.009 | |
| 7 | Proterometra macrostoma | 6.245 | 6.089 | 6.089 | 6.883 | 6.792 | 4.84 | 0.006 | 0.009 | |
| 8 | Proterometra sp. | 6.328 | 6.172 | 5.938 | 6.419 | 6.328 | 5.078 | 5.078 | 0.009 | |
| 9 | Otodistomum cestoides | 12.588 | 12.432 | 12.353 | 12.366 | 12.353 | 12.432 | 13.370 | 12.989 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Vainutis, K.S.; Voronova, A.N.; Mironovsky, A.N.; Zhigileva, O.N.; Zhokhov, A.E. The Species Diversity Assessment of Azygia Looss, 1899 (Digenea: Azygiidae) from the Volga, Ob, and Artyomovka Rivers Basins (Russia), with Description of A. sibirica n. sp. Diversity 2023, 15, 119. https://doi.org/10.3390/d15010119
Vainutis KS, Voronova AN, Mironovsky AN, Zhigileva ON, Zhokhov AE. The Species Diversity Assessment of Azygia Looss, 1899 (Digenea: Azygiidae) from the Volga, Ob, and Artyomovka Rivers Basins (Russia), with Description of A. sibirica n. sp. Diversity. 2023; 15(1):119. https://doi.org/10.3390/d15010119
Chicago/Turabian StyleVainutis, Konstantin S., Anastasia N. Voronova, Alexander N. Mironovsky, Oksana N. Zhigileva, and Alexander E. Zhokhov. 2023. "The Species Diversity Assessment of Azygia Looss, 1899 (Digenea: Azygiidae) from the Volga, Ob, and Artyomovka Rivers Basins (Russia), with Description of A. sibirica n. sp." Diversity 15, no. 1: 119. https://doi.org/10.3390/d15010119