
DNA Barcoding of Trichobilharzia (Trematoda: Schistosomatidae) Species and Their Detection in eDNA Water Samples
 , , , , ,
, , , , ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Field Work and eDNA Sampling
2.2. Release of Cercariae in the Laboratory
2.3. Cercariae Analysed Genetically
2.4. DNA Extraction
2.5. PCR Amplification of Marker Sequences
2.6. Establishment of the eDNA PCR Primers
2.7. Sequence Analysis
2.8. Detection of Trematode DNA in eDNA Samples from the Field
2.9. eDNA from Aquarium Water
2.10. Sensitivity Tests
3. Results
3.1. Establishment of the eDNA-PCR Primers
3.2. eDNA Experiments
3.2.1. eDNA Samples from the Field
3.2.2. eDNA from Aquarium Water
3.3. Sensitivity Tests
3.4. Cercariae Release Experiments
4. Discussion
4.1. Detection of Trichobilharzia Species—Correspondence between Observations and eDNA Tests
4.2. Stochasticity of eDNA Detection
4.3. DNA Degradation and Sensitivity of PCR Detection
4.4. eDNA Barcoding for the Detection of Trichobilharzia in Practice
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Pawlowski, J.; Apothéloz-Perret-Gentil, L.; Mächler, E.; Altermatt, F. Anwendung von eDNA-Methoden in Biologischen Untersuchungen und bei der Biologischen Bewertung von Aquatischen Ökosystemen: Richtlinien; Bundesamt für Umwelt: Bern, Switzerland, 2020. [Google Scholar]
- Taberlet, P.; Bonin, A.; Zinger, L.; Coissac, E. Environmental DNA: For Biodiversity Research and Monitoring; Online Edi.; Oxford University Press: Oxford, UK, 2018; ISBN 9780198767220. [Google Scholar]
- Harrison, J.B.; Sunday, J.M.; Rogers, S.M. Predicting the fate of eDNA in the environment and implications for studying biodiversity. Proc. R. Soc. B Biol. Sci. 2019, 286, 20191409. [Google Scholar] [CrossRef] [PubMed]
- Jo, T.; Arimoto, M.; Murakami, H.; Masuda, R.; Minamoto, T. Estimating shedding and decay rates of environmental nuclear DNA with relation to water temperature and biomass. Environ. DNA 2020, 2, 140–151. [Google Scholar] [CrossRef] [Green Version]
- Strickler, K.M.; Fremier, A.K.; Goldberg, C.S. Quantifying effects of UV-B, temperature, and pH on eDNA degradation in aquatic microcosms. Biol. Conserv. 2015, 183, 85–92. [Google Scholar] [CrossRef]
- Troth, C.R.; Sweet, M.J.; Nightingale, J.; Burian, A. Seasonality, DNA degradation and spatial heterogeneity as drivers of eDNA detection dynamics. Sci. Total Environ. 2021, 768, 144466. [Google Scholar] [CrossRef]
- Bochove, K.; Bakker, F.T.; Beentjes, K.K.; Hemerik, L.; Vos, R.A.; Gravendeel, B. Organic matter reduces the amount of detectable environmental DNA in freshwater. Ecol. Evol. 2020, 10, 3647–3654. [Google Scholar] [CrossRef]
- Loker, E.S.; DeJong, R.J.; Brant, S.V. Scratching the Itch: Updated Perspectives on the Schistosomes Responsible for Swimmer’s Itch around the World. Pathogens 2022, 11, 587. [Google Scholar] [CrossRef]
- Auer, H.; Aspöck, H. “Vogelbilharzien” als Erreger einer Hautkrankheit: Die Zerkarien-Dermatitis. Denisia 2002, 6, 321–331. [Google Scholar]
- Sattmann, H.; Hörweg, C.; Konecny, R. Zerkariendermatitis in Österreich—Rückblick und Perspektiven. Denisia 2004, 13, 457–461. [Google Scholar]
- Horák, P.; Mikeš, L.; Lichtenbergová, L.; Skála, V.; Soldánová, M.; Brant, S.V. Avian Schistosomes and Outbreaks of Cercarial Dermatitis. Clin. Microbiol. Rev. 2015, 28, 165–190. [Google Scholar] [CrossRef] [Green Version]
- Kolářová, L.; Horák, P.; Skírnisson, K.; Marečková, H.; Doenhoff, M. Cercarial Dermatitis, a Neglected Allergic Disease. Clin. Rev. Allergy Immunol. 2013, 45, 63–74. [Google Scholar] [CrossRef]
- Soldánová, M.; Selbach, C.; Kalbe, M.; Kostadinova, A.; Sures, B. Swimmer’s itch: Etiology, impact, and risk factors in Europe. Trends Parasitol. 2013, 29, 65–74. [Google Scholar] [CrossRef]
- Brant, S.V.; Loker, E.S. Discovery-based studies of schistosome diversity stimulate new hypotheses about parasite biology. Trends Parasitol. 2013, 29, 449–459. [Google Scholar] [CrossRef]
- Jouet, D.; Kolářová, L.; Patrelle, C.; Ferté, H.; Skírnisson, K. Trichobilharzia anseri n. sp. (Schistosomatidae: Digenea), a new visceral species of avian schistosomes isolated from greylag goose (Anser anser L.) in Iceland and France. Infect. Genet. Evol. 2015, 34, 298–306. [Google Scholar] [CrossRef]
- Christiansen, A.Ø.; Olsen, A.; Buchmann, K.; Kania, P.W.; Nejsum, P.; Vennervald, B.J. Molecular diversity of avian schistosomes in Danish freshwater snails. Parasitol. Res. 2016, 115, 1027–1037. [Google Scholar] [CrossRef]
- Neuhaus, W. Biologie und Entwicklung von Trichobilharzia szidati n. sp. (Trematoda, Schistosomatidae), einem Erreger von Dermatitis beim Menschen. Z. Parasitenkd. 1952, 15, 203–266. [Google Scholar] [CrossRef]
- Horák, P.; Kolářová, L.; Dvořák, J. Trichobilharzia regenti n. sp. (Schistosomatidae, Bilharziellinae), a new nasal schistosome from Europe. Parasite 1998, 5, 349–357. [Google Scholar] [CrossRef] [Green Version]
- Müller, V.; Kimmig, P. Trichobilharzia franki n. sp.—Die Ursache für Badedermatitiden in südwestdeutschen Baggerseen [Trichobilharzia franki n. sp.—A causative agent of swimmer’s itch in south-western Germany]. Appl. Parasitol. 1994, 35, 12–31. [Google Scholar]
- Simon-Martin, F.; Simon-Vicente, F. The life cycle of Trichobilharzia salmanticensis n. sp. (Digenea: Schistosomatidae), related to cases of human dermatitis. Res. Rev. Parasitol. 1999, 59, 13–18. [Google Scholar]
- Kolářová, L.; Skírnisson, K.; Ferté, H.; Jouet, D. Trichobilharzia mergi sp. nov. (Trematoda: Digenea: Schistosomatidae), a visceral schistosome of Mergus serrator (L.) (Aves: Anatidae). Parasitol. Int. 2013, 62, 300–308. [Google Scholar] [CrossRef]
- Talbot, S.B. Studies on schistosome dermatitis: II. Morphological and life history studies on three dermatitis-producing schistosome cercariae, C. elvae Miller, 1923, C. stagnicolae n. sp., and C. physellae n. sp. Am. J. Epidemiol. 1936, 23, 372–384. [Google Scholar] [CrossRef]
- Helmer, N.; Blatterer, H.; Hörweg, C.; Reier, S.; Sattmann, H.; Schindelar, J.; Szucsich, N.U.; Haring, E. First Record of Trichobilharzia physellae (Talbot, 1936) in Europe, a Possible Causative Agent of Cercarial Dermatitis. Pathogens 2021, 10, 1473. [Google Scholar] [CrossRef] [PubMed]
- McPhail, B.A.; Froelich, K.; Reimink, R.L.; Hanington, P.C. Simplifying Schistosome Surveillance: Using Molecular Cercariometry to Detect and Quantify Cercariae in Water. Pathogens 2022, 11, 565. [Google Scholar] [CrossRef]
- Sato, M.O.; Rafalimanantsoa, A.; Ramarokoto, C.; Rahetilahy, A.M.; Ravoniarimbinina, P.; Kawai, S.; Minamoto, T.; Sato, M.; Kirinoki, M.; Rasolofo, V.; et al. Usefulness of environmental DNA for detecting Schistosoma mansoni occurrence sites in Madagascar. Int. J. Infect. Dis. 2018, 76, 130–136. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rudko, S.P.; Turnbull, A.; Reimink, R.L.; Froelich, K.; Hanington, P.C. Species-specific qPCR assays allow for high-resolution population assessment of four species avian schistosome that cause swimmer’s itch in recreational lakes. Int. J. Parasitol. Parasites Wildl. 2019, 9, 122–129. [Google Scholar] [CrossRef]
- Alzaylaee, H.; Collins, R.A.; Shechonge, A.; Ngatunga, B.P.; Morgan, E.R.; Genner, M.J. Environmental DNA-based xenomonitoring for determining Schistosoma presence in tropical freshwaters. Parasit. Vectors 2020, 13, 63. [Google Scholar] [CrossRef]
- Sengupta, M.E.; Hellström, M.; Kariuki, H.C.; Olsen, A.; Thomsen, P.F.; Mejer, H.; Willerslev, E.; Mwanje, M.T.; Madsen, H.; Kristensen, T.K.; et al. Environmental DNA for improved detection and environmental surveillance of schistosomiasis. Proc. Natl. Acad. Sci. USA 2019, 116, 8931–8940. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kane, R.A.; Stothard, J.R.; Rollinson, D.; Leclipteux, T.; Evraerts, J.; Standley, C.J.; Allan, F.; Betson, M.; Kaba, R.; Mertens, P.; et al. Detection and quantification of schistosome DNA in freshwater snails using either fluorescent probes in real-time PCR or oligochromatographic dipstick assays targeting the ribosomal intergenic spacer. Acta Trop. 2013, 128, 241–249. [Google Scholar] [CrossRef] [PubMed]
- Jothikumar, N.; Mull, B.J.; Brant, S.V.; Loker, E.S.; Collinson, J.; Secor, W.E.; Hill, V.R. Real-Time PCR and Sequencing Assays for Rapid Detection and Identification of Avian Schistosomes in Environmental Samples. Appl. Environ. Microbiol. 2015, 81, 4207–4215. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rudko, S.P.; Reimink, R.L.; Froelich, K.; Gordy, M.A.; Blankespoor, C.L.; Hanington, P.C. Use of qPCR-Based Cercariometry to Assess Swimmer’s Itch in Recreational Lakes. Ecohealth 2018, 15, 827–839. [Google Scholar] [CrossRef] [Green Version]
- Soldánová, M.; Selbach, C.; Sures, B. The Early Worm Catches the Bird? Productivity and Patterns of Trichobilharzia szidati Cercarial Emission from Lymnaea stagnalis. PLoS ONE 2016, 11, e0149678. [Google Scholar] [CrossRef] [Green Version]
- Al-Jubury, A.; Kania, P.W.; Bygum, A.; Buchmann, K. Temperature and light effects on Trichobilharzia szidati cercariae with implications for a risk analysis. Acta Vet. Scand. 2020, 62, 54. [Google Scholar] [CrossRef] [PubMed]
- Horák, P.; Kolářová, L.; Adema, C.M. Biology of the schistosome genus Trichobilharzia. Adv. Parasitol. 2002, 52, 155–233. [Google Scholar] [CrossRef] [PubMed]
- Reier, S.; Haring, E.; Billinger, F.; Blatterer, H.; Duda, M.; Gorofsky, C.; Grasser, H.-P.; Heinisch, W.; Hörweg, C.; Kruckenhauser, L.; et al. First confirmed record of Trichobilharzia franki Müller & Kimmig, 1994, from Radix auricularia (Linnaeus, 1758) for Austria. Parasitol. Res. 2020, 119, 4135–4141. [Google Scholar] [CrossRef] [PubMed]
- Folmer, O.; Black, M.B.; Hoeh, W.R.; Lutz, R.A.; Vrijenhoek, R.C. DNA primers for amplification of mitochondrial cytochrome c oxidase subunit I from diverse metazoan invertebrates. Mol. Mar. Biol. Biotechnol. 1994, 3, 294–299. [Google Scholar]
- Hohensee, N. Digenean Trematodes in Freshwater Snails in the Surroundings of Vienna with a Focus on Species Involved in Human Infections; University of Vienna: Vienna, Austria, 2016. [Google Scholar]
- Nicholas, K.B.; Nicholas, H.B.J. GeneDoc: A tool for editing and annotating multiple sequence alignments. Embnet. News 1997, 4, 1–4. [Google Scholar]
- Al-Jubury, A.; Duan, Y.; Kania, P.W.; Tracz, E.S.; Bygum, A.; von Jørgensen, L.G.; Horák, P.; Buchmann, K. Avian schistosome species in Danish freshwater lakes: Relation to biotic and abiotic factors. J. Helminthol. 2021, 95, e22. [Google Scholar] [CrossRef]
- Gaub, L.; Sattmann, H.; Hörweg, C.; Walochnik, J. Molecular validation of species determination of larval trematodes from freshwater snail hosts in Austria, with special emphasis on the genus Trichobilharzia Skrjabin & Zakharow, 1920. Arianta 2020, 8, 13–19. [Google Scholar]
- Duan, Y.; Al-Jubury, A.; Kania, P.W.; Buchmann, K. Trematode diversity reflecting the community structure of Danish freshwater systems: Molecular clues. Parasit. Vectors 2021, 14, 43. [Google Scholar] [CrossRef]
- Gordy, M.A.; Hanington, P.C. A fine-scale phylogenetic assessment of digenean trematodes in central Alberta reveals we have yet to uncover their total diversity. Ecol. Evol. 2019, 9, 3153–3238. [Google Scholar] [CrossRef] [Green Version]
- Selbach, C.; Soldánová, M.; Feld, C.K.; Kostadinova, A.; Sures, B. Hidden parasite diversity in a European freshwater system. Sci. Rep. 2020, 10, 2694. [Google Scholar] [CrossRef] [Green Version]
- Mauvisseau, Q.; Halfmaerten, D.; Neyrinck, S.; Burian, A.; Brys, R. Effects of preservation strategies on environmental DNA detection and quantification using ddPCR. Environ. DNA 2021, 3, 815–822. [Google Scholar] [CrossRef]
- Allison, M.J.; Round, J.M.; Bergman, L.C.; Mirabzadeh, A.; Allen, H.; Weir, A.; Helbing, C.C. The effect of silica desiccation under different storage conditions on filter-immobilized environmental DNA. BMC Res. Notes 2021, 14, 106. [Google Scholar] [CrossRef] [PubMed]
- Spens, J.; Evans, A.R.; Halfmaerten, D.; Knudsen, S.W.; Sengupta, M.E.; Mak, S.S.T.; Sigsgaard, E.E.; Hellström, M. Comparison of capture and storage methods for aqueous macrobial eDNA using an optimized extraction protocol: Advantage of enclosed filter. Methods Ecol. Evol. 2017, 8, 635–645. [Google Scholar] [CrossRef] [Green Version]
- Furlan, E.M.; Gleeson, D.; Wisniewski, C.; Yick, J.; Duncan, R.P. eDNA surveys to detect species at very low densities: A case study of European carp eradication in Tasmania, Australia. J. Appl. Ecol. 2019, 56, 2505–2517. [Google Scholar] [CrossRef]
- Sckrabulis, J.P.; Flory, A.R.; Raffel, T.R. Direct onshore wind predicts daily swimmer’s itch (avian schistosome) incidence at a Michigan beach. Parasitology 2020, 147, 431–440. [Google Scholar] [CrossRef] [PubMed]
- Hunter, M.E.; Ferrante, J.A.; Meigs-Friend, G.; Ulmer, A. Improving eDNA yield and inhibitor reduction through increased water volumes and multi-filter isolation techniques. Sci. Rep. 2019, 9, 5259. [Google Scholar] [CrossRef] [Green Version]
- Beentjes, K.K.; Speksnijder, A.G.C.L.; Schilthuizen, M.; Hoogeveen, M.; van der Hoorn, B.B. The effects of spatial and temporal replicate sampling on eDNA metabarcoding. PeerJ 2019, 7, e7335. [Google Scholar] [CrossRef]
- Barnes, M.A.; Turner, C.R.; Jerde, C.L.; Renshaw, M.A.; Chadderton, W.L.; Lodge, D.M. Environmental Conditions Influence eDNA Persistence in Aquatic Systems. Environ. Sci. Technol. 2014, 48, 1819–1827. [Google Scholar] [CrossRef]
- Dejean, T.; Valentini, A.; Duparc, A.; Pellier-Cuit, S.; Pompanon, F.; Taberlet, P.; Miaud, C. Persistence of Environmental DNA in Freshwater Ecosystems. PLoS ONE 2011, 6, e23398. [Google Scholar] [CrossRef] [Green Version]
- Pilliod, D.S.; Goldberg, C.S.; Arkle, R.S.; Waits, L.P. Factors influencing detection of eDNA from a stream-dwelling amphibian. Mol. Ecol. Resour. 2014, 14, 109–116. [Google Scholar] [CrossRef]
- Jo, T.; Murakami, H.; Yamamoto, S.; Masuda, R.; Minamoto, T. Effect of water temperature and fish biomass on environmental DNA shedding, degradation, and size distribution. Ecol. Evol. 2019, 9, 1135–1146. [Google Scholar] [CrossRef] [Green Version]
- Goldberg, C.S.; Strickler, K.M.; Fremier, A.K. Degradation and dispersion limit environmental DNA detection of rare amphibians in wetlands: Increasing efficacy of sampling designs. Sci. Total Environ. 2018, 633, 695–703. [Google Scholar] [CrossRef]
- Seymour, M.; Durance, I.; Cosby, B.J.; Ransom-Jones, E.; Deiner, K.; Ormerod, S.J.; Colbourne, J.K.; Wilgar, G.; Carvalho, G.R.; de Bruyn, M.; et al. Acidity promotes degradation of multi-species environmental DNA in lotic mesocosms. Commun. Biol. 2018, 1, 4. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Balasingham, K.D.; Walter, R.P.; Mandrak, N.E.; Heath, D.D. Environmental DNA detection of rare and invasive fish species in two Great Lakes tributaries. Mol. Ecol. 2018, 27, 112–127. [Google Scholar] [CrossRef] [Green Version]
- Dougherty, M.M.; Larson, E.R.; Renshaw, M.A.; Gantz, C.A.; Egan, S.P.; Erickson, D.M.; Lodge, D.M. Environmental DNA (eDNA) detects the invasive rusty crayfish Orconectes rusticus at low abundances. J. Appl. Ecol. 2016, 53, 722–732. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ushio, M.; Murata, K.; Sado, T.; Nishiumi, I.; Takeshita, M.; Iwasaki, W.; Miya, M. Demonstration of the potential of environmental DNA as a tool for the detection of avian species. Sci. Rep. 2018, 8, 4493. [Google Scholar] [CrossRef] [PubMed]


{kind=link}
| Locality Number | Locality | Coordinates N/E |
|---|---|---|
| 1 | AT, OÖ, Unterer Inn, Reichersberger Au | 48°20′20.71″/13°21′35.47″ |
| 2 | AT, OÖ, Unterer Inn, Hagenauer Bucht | 48°16′31.96″/13°05′33.58″ |
| 3 | AT, OÖ, Eggelsberg, Seeleithensee | 48°03′33.19″/12°58′01.56″ |
| 4 | AT, NÖ, Rekawinkel, pond | 48°11′02.90″/16°01′56.50″ |
| 5 | AT, OÖ, Linz, Lake Pichlinger-See | 48°14′22.62″/14°23′00.05″ |
| 6 | CZ, Prague, Charles University | lab breeding |
| 7 | CZ, Jetřichovice, natural swimming pond | 50°50′38.50″/14°23′43.00″ |
| 8 | AT, NÖ, Niederfellabrunn, Löschteich | 48°27′40.70″/16°18′42.50″ |
| 9 | AT, OÖ, Linz, Lake Pleschinger-See | 48°19′10.00″/14°19′56.80″ |
| Species | Accession Number/ BOLD-ID | Ind.ID | Host | Life Stage | Locality Code |
|---|---|---|---|---|---|
| Trichobilharzia franki | OP347092/NHBP008-21 | Ra2-4-014 | Radix auricularia | cercaria | 1 |
| Trichobilharzia franki | OP347091/NHBP016-21 | Ra13-16-002 | Radix auricularia | cercaria | 7 |
| Trichobilharzia regenti | OP347089/NHBP010-21 | Rlag1-15-002 | Radix lagotis | cercaria | 6 |
| Trichobilharzia szidati | OP347090/NHBP014-21 | Ls17-15-003 | Lymnaea stagnalis | cercaria | 6 |
| Trichobilharzia szidati | OP347093/NHBP018-21 | Ls36-13-002 | Lymnaea stagnalis | cercaria | 4 |
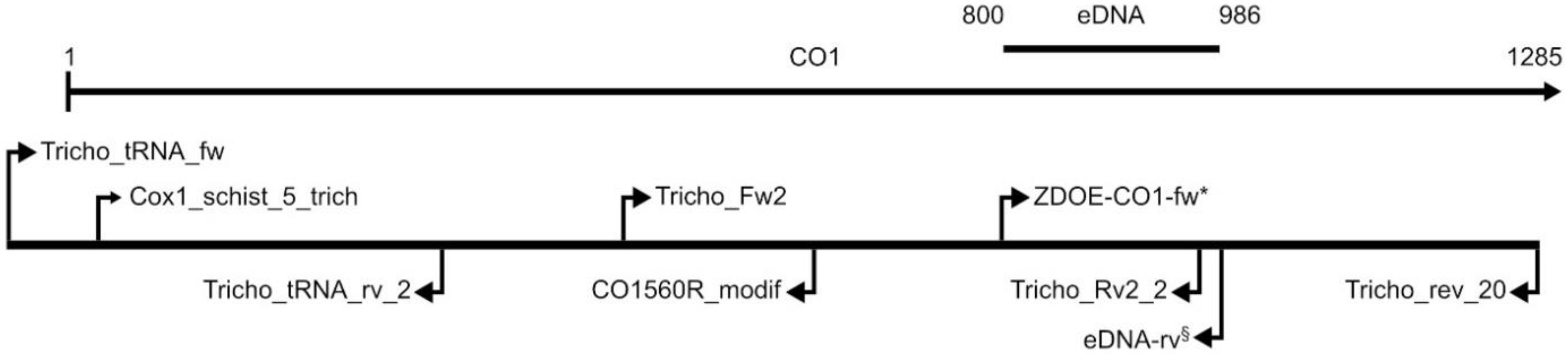
| Gene | Primer Name | Orientation | Sequence 5′-3′ | Source |
|---|---|---|---|---|
| CO1 | Tricho_tRNA_fw | Fwd | GGTTGTCGCTGCTAACGA | [23] |
| CO1 | Cox1_schist_5_trich | Fwd | GTTRGTTTCTTTGGATCATAAGCG | [23] |
| CO1 | Tricho_tRNA_rv_2 | Rev | CCATATAAAACATTGAAGGAACC | [23] |
| CO1 | Tricho_Fw2 | Fwd | GGTTCTGTAAAATTTATAACTAC | [23] |
| CO1 | CO1560R_modif | Rev | GCAGTACCAAATTTTCGATC | [23] |
| CO1 | Tricho_Rv2_2 | Rev | CCTAACATATACAACCAAG | [23] |
| CO1 | Tricho_rev_20 | Rev | GCATTCCTAAATAATGCATAGG | [23] |
| CO1 | ZDOE-CO1-fw | Fwd | TAGTTTGTGCTATGGGTTCTATAGT | [23] |
| CO1 | ZDOE-COI-fw_szidati * | Fwd | TAGTTTGTGCTATGGGTTCTATAGTG | present study |
| CO1 | ZDOE-COI-fw_physellae * | Fwd | TGGTTTGTGCTATGGGTTCTATAGTT | present study |
| CO1 | eDNA-franki-rv * | Rev | CCCCACGCAAATACCTTGTG | present study |
| CO1 | eDNA-regenti-rv * | Rev | CTCCACGTAAATAACTAGTA | present study |
| CO1 | eDNA-szidati-rv * | Rev | CACCCCGCAAGTAGCTAGTC | present study |
| CO1 | eDNA-physellae-rv * | Rev | CTCCTCGCAAATAACTAGTT | present study |
| 18S | Trem-18S-f | Fwd | GGTTCCTTAGATCGTACATGC | [37] |
| 18S | Trem-18S-r | Rev | GTACTCATTCGAATTACGGAGC | [37] |
| Primer Combination | Amplicon Length | Tann/Elongation Time |
|---|---|---|
| Cox1_schist_5_trich/Tricho_rev_20 | 1216 bp | 53 °C/90 s |
| Tricho_tRNA_fw/Tricho_tRNA_rv_2 | 396 bp | 54 °C/60 s |
| Cox1_schist_5_trich/CO1560R_modif | 612 bp | 52 °C/6 0 s |
| Tricho_Fw2/Tricho_Rv2_2 | 491 bp | 49 °C/60 s |
| ZDOE-COI-fw/Tricho_rev_20 | 486 bp | 53 °C/60 s |
| ZDOE-CO1-fw/eDNA-franki-rv | 186 bp | 54 °C/30 s |
| ZDOE-CO1-fw/eDNA-regenti-rv | 186 bp | 50 °C/30 s |
| ZDOE-CO1-fw/eDNA-szidati-rv | 186 bp | 57 °C/30 s |
| ZDOE-CO1-fw/eDNA-physellae-rv | 186 bp | 51 °C/30 s |
| ZDOE-COI-fw_szidati/eDNA-szidati-rv | 186 bp | 57 °C/30 s |
| ZDOE-COI-fw_physellae/eDNA-physellae-rv | 186 bp | 51 °C/30 s |
| Trem-18S-f/Trem-18S-r | 413–428 bp | 57 °C/60 s |
| Ind. ID | Locality Number | Sample Date | Sample Type, Volume | Genbank BLAST Hit | CO1 TestT. szidati |
|---|---|---|---|---|---|
| 1-A-4 | 1 | 04.06.19 | aquarium sample, 600 mL | Echinostoma revolutum (18S) | 0/3 |
| 2-W-13 | 4 | 09.07.19 | water sample from field, 360 mL | 0/3 | |
| 3-W-14 | 5 | 19.07.19 | water sample from field, 600 mL | Lissorchis kritskyi or Auridistomum chelydrae (18S) | 0/3 |
| 4-W-8 | 2 | 19.08.19 | water sample from field, 180 mL | Echinostoma revolutum (18S) | 0/3 |
| eDNA-1 | 2 | 07.07.20 | water sample from field, 200 mL | Sminthurides aquaticus (18S) | 0/3 |
| eDNA-2 | 2 | 07.07.20 | water sample from field, 300 mL | Sminthurides aquaticus (18S) | 0/3 |
| eDNA-3 | 8 | 17.07.20 | water sample from field, 1200 mL | Opisthioglyphe ranae (18S) | 0/3 |
| eDNA-4 | 8 | 17.07.20 | water sample from field 350 mL | Opisthioglyphe ranae (18S) | 0/3 |
| eDNA-5 | (4) | 24.08.20 | aquarium sample(with L. stagnalis), 170 mL | Trichobilharzia szidati (CO1) | 3/3 |
| eDNA-6 | (4) | 24.08.20 | aquarium sample(with L. stagnalis), 200 mL | Trichobilharzia szidati (CO1) | 3/3 |
| eDNA-7 | 4 | 24.08.20 | water sample from field, 120 mL | Trichobilharzia szidati (CO1), Parastrigea robusta (18S) | 1/3 |
| eDNA-8 | 4 | 24.08.20 | water sample from field, 200 mL | Sminthurides aquaticus (18S) | 0/6 |
| eDNA-9 | 4 | 24.08.20 | water sample from field, 170 mL | Trichobilharzia szidati (CO1) | 2/3 |
| eDNA-10 | 4 | 24.08.20 | water sample from field, 100 mL | 0/6 | |
| eDNA-11 | 4 | 24.08.20 | transport water sample(with L. stagnalis), 130 mL | Trichobilharzia szidati (CO1), Trichobilharzia spp.various identical (18S) | 3/3 |
| eDNA-12 | (4) | 25.08.20 | aquarium test sample 1, 450 mL | Trichobilharzia szidati (CO1) | 3/3 |
| eDNA-13 | (4) | 26.08.20 | aquarium test sample 2, 450 mL | Trichobilharzia szidati (CO1) | 3/3 |
| eDNA-14 | (4) | 27.08.20 | aquarium test sample 3, 350 mL | Trichobilharzia szidati (CO1) | 3/3 |
| eDNA-15 | (4) | 28.08.20 | aquarium test sample 4, 350 mL | Trichobilharzia szidati (CO1) | 3/3 |
| eDNA-16 | (4) | 29.08.20 | aquarium test sample 5, 300 mL | Trichobilharzia szidati (CO1) | 3/3 |
| eDNA-17 | (4) | 30.08.20 | aquarium test sample 6, 260 mL | Trichobilharzia szidati (CO1) | 3/3 |
| eDNA-18 | (4) | 31.08.20 | aquarium test sample 7, 250 mL | Trichobilharzia szidati (CO1) | 3/3 |
| eDNA-20 | (4) | 09.09.20 | aquarium test sample 8, 150 mL | 0/6 | |
| eDNA-21 | (4) | 10.09.20 | aquarium test sample 9, 200 mL | 0/6 | |
| eDNA-22 | (4) | 15.09.20 | aquarium test sample 10, 60 mL | 0/6 | |
| eDNA-23 | (4) | 15.09.20 | aquarium test sample 11, 70 mL | 0/6 | |
| eDNA-24 | (4) | 07.10.20 | aquarium test sample 12, 140 mL | Trichobilharzia szidati (CO1) | 1/3 |
| eDNA-25 | 9 | 24.09.20 | water sample from field, 800 mL | Diplostomum pseudospathaceum (18S) | 0/3 |
| eDNA-26 | 9 | 16.10.20 | water sample lab, 850 mL | 0/3 | |
| eDNA-27 | 9 | 16.10.20 | water sample lab, 800 mL | Trichobilharzia physellae (CO1) | 0/3 |
| eDNA-28 | 9 | 16.10.20 | water sample lab, 850 mL | 0/3 | |
| eDNA-29 | 9 | 16.10.20 | water sample lab, 950 mL | 0/3 | |
| eDNA-31 | 3 | 01.08.19 | water sample from field; 780 mL | 0/3 |
| Locality Number | Cercaria Species | Host Snail | Collection Date (dd.mm.yyyy) | Prevalence |
|---|---|---|---|---|
| 2 | Trichobilharzia szidati * | Lymnaea stagnalis | 19.08.2019 | 9/28 (32.1%) |
| 1 | Trichobilharzia franki | Radix auricularia | 27.05.2019 | 1/10 (10.0%) |
| 4 | Trichobilharzia szidati | Lymnaea stagnalis | 20.08.2020 24.08.2020 | 3/9 (33.3%)1/12 (8.3%) |
| 9 | Trichobilharzia physellae | Physella acuta | 16.09.2020 11.11.2020 | 1/6 (16.7%)1/30 (3.3%) |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Helmer, N.; Hörweg, C.; Sattmann, H.; Reier, S.; Szucsich, N.U.; Bulantová, J.; Haring, E. DNA Barcoding of Trichobilharzia (Trematoda: Schistosomatidae) Species and Their Detection in eDNA Water Samples. Diversity 2023, 15, 104. https://doi.org/10.3390/d15010104
Helmer N, Hörweg C, Sattmann H, Reier S, Szucsich NU, Bulantová J, Haring E. DNA Barcoding of Trichobilharzia (Trematoda: Schistosomatidae) Species and Their Detection in eDNA Water Samples. Diversity. 2023; 15(1):104. https://doi.org/10.3390/d15010104
Chicago/Turabian StyleHelmer, Nikolaus, Christoph Hörweg, Helmut Sattmann, Susanne Reier, Nikolaus U. Szucsich, Jana Bulantová, and Elisabeth Haring. 2023. "DNA Barcoding of Trichobilharzia (Trematoda: Schistosomatidae) Species and Their Detection in eDNA Water Samples" Diversity 15, no. 1: 104. https://doi.org/10.3390/d15010104