
The Role of the Environment in Shaping the Genomic Variation in an Insular Wild Boar Population


Abstract
:1. Introduction
2. Materials and Methods
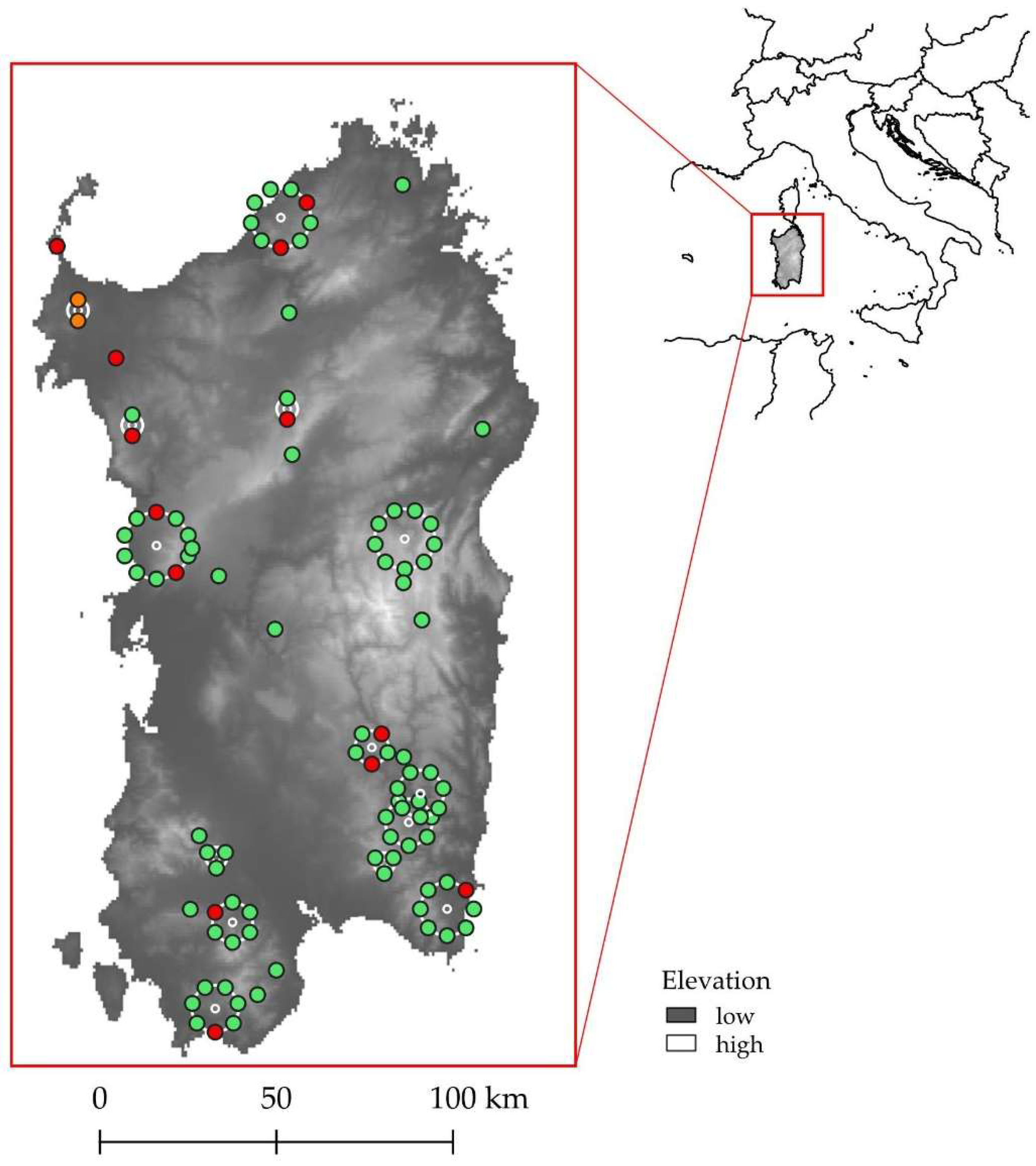
2.1. Study Area
2.2. Dataset Selection and Quality Control
2.3. Population Structure Analysis
2.4. Search for Local Adaptation
2.4.1. Outlier Analysis
2.4.2. Environmental Data Retrieval and Elaboration
2.4.3. Genotype–Environment Association
2.5. Annotation and Gene Ontology Enrichment Analysis
3. Results
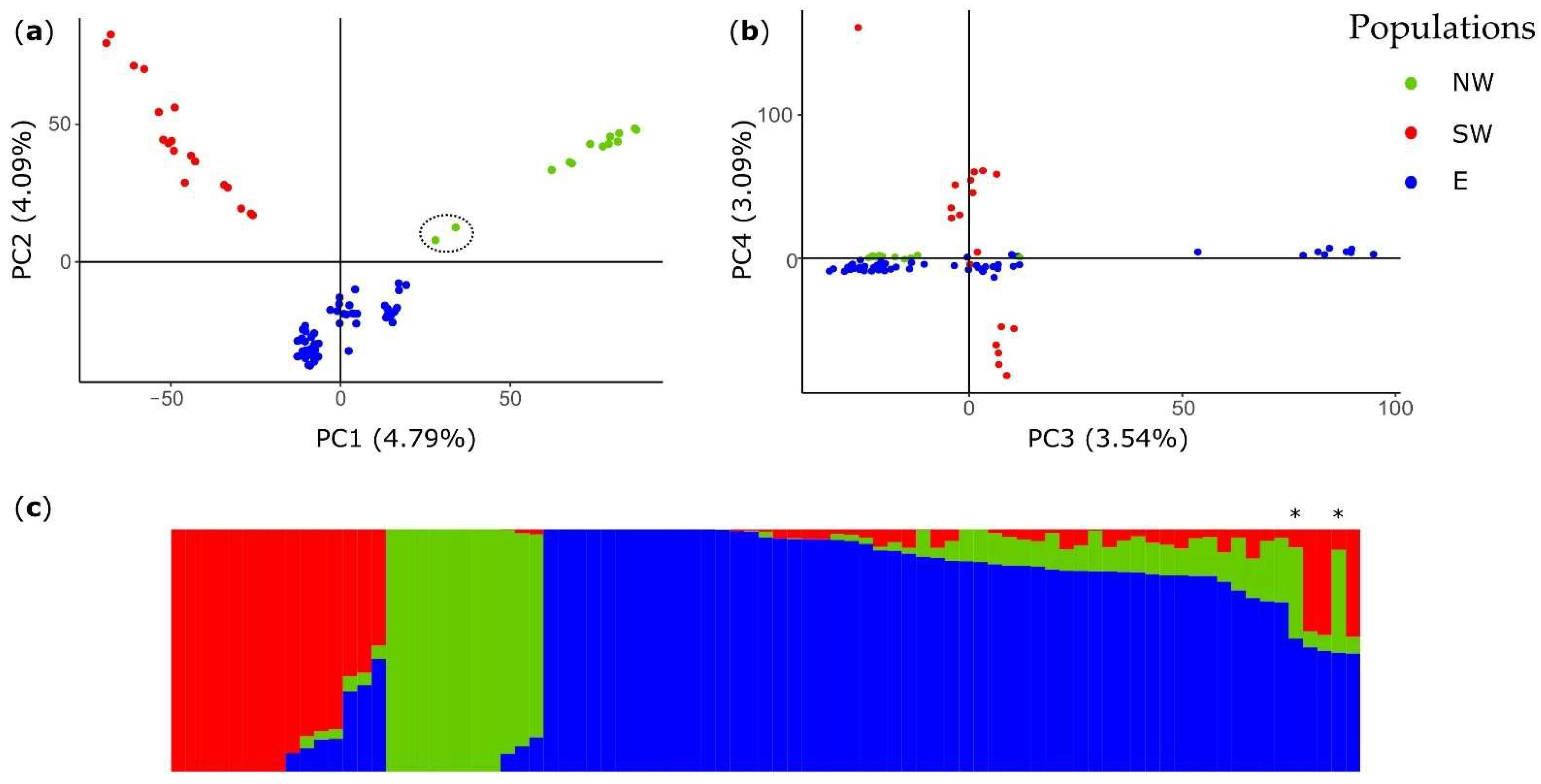
3.1. Population Structure
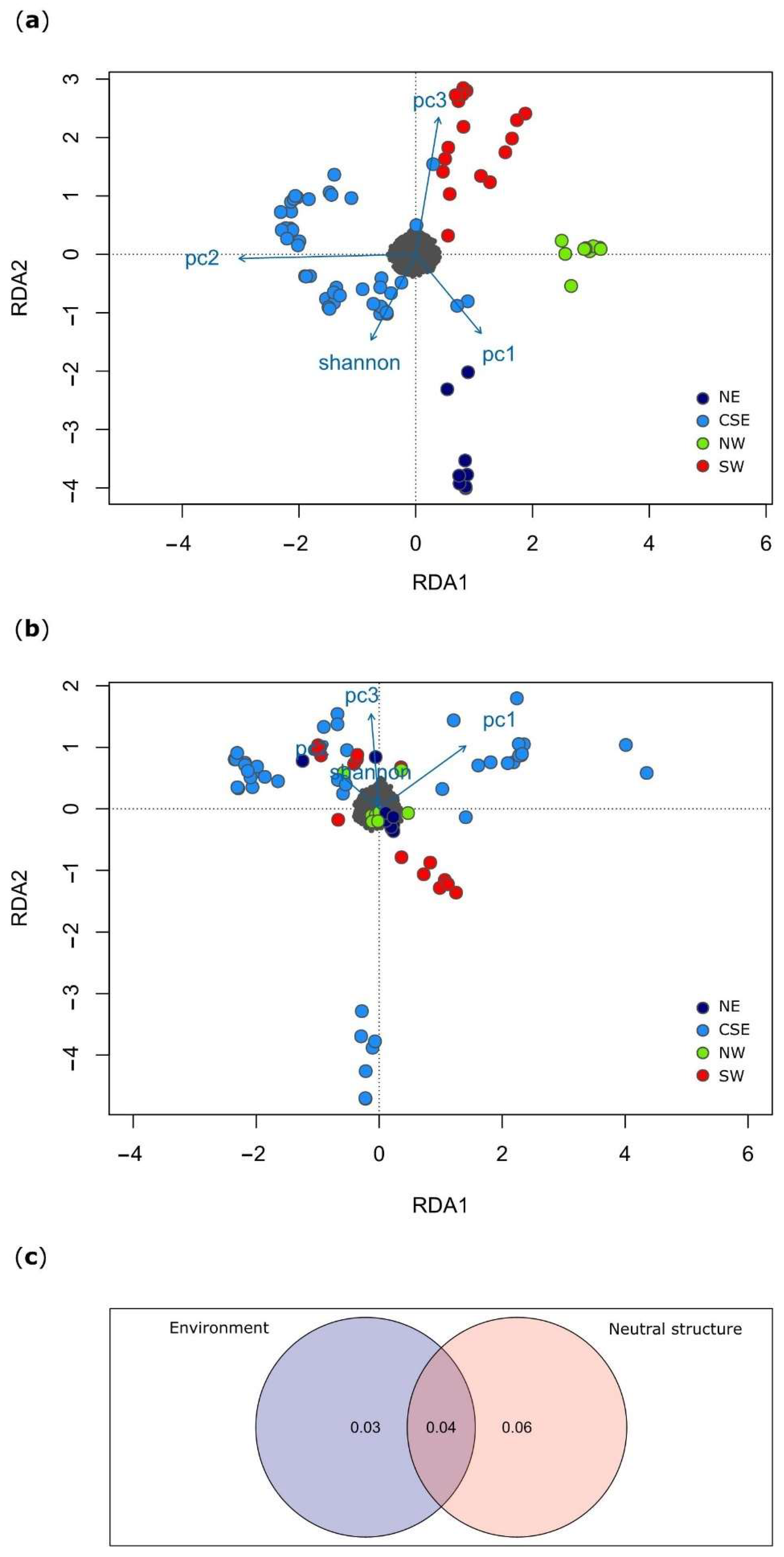
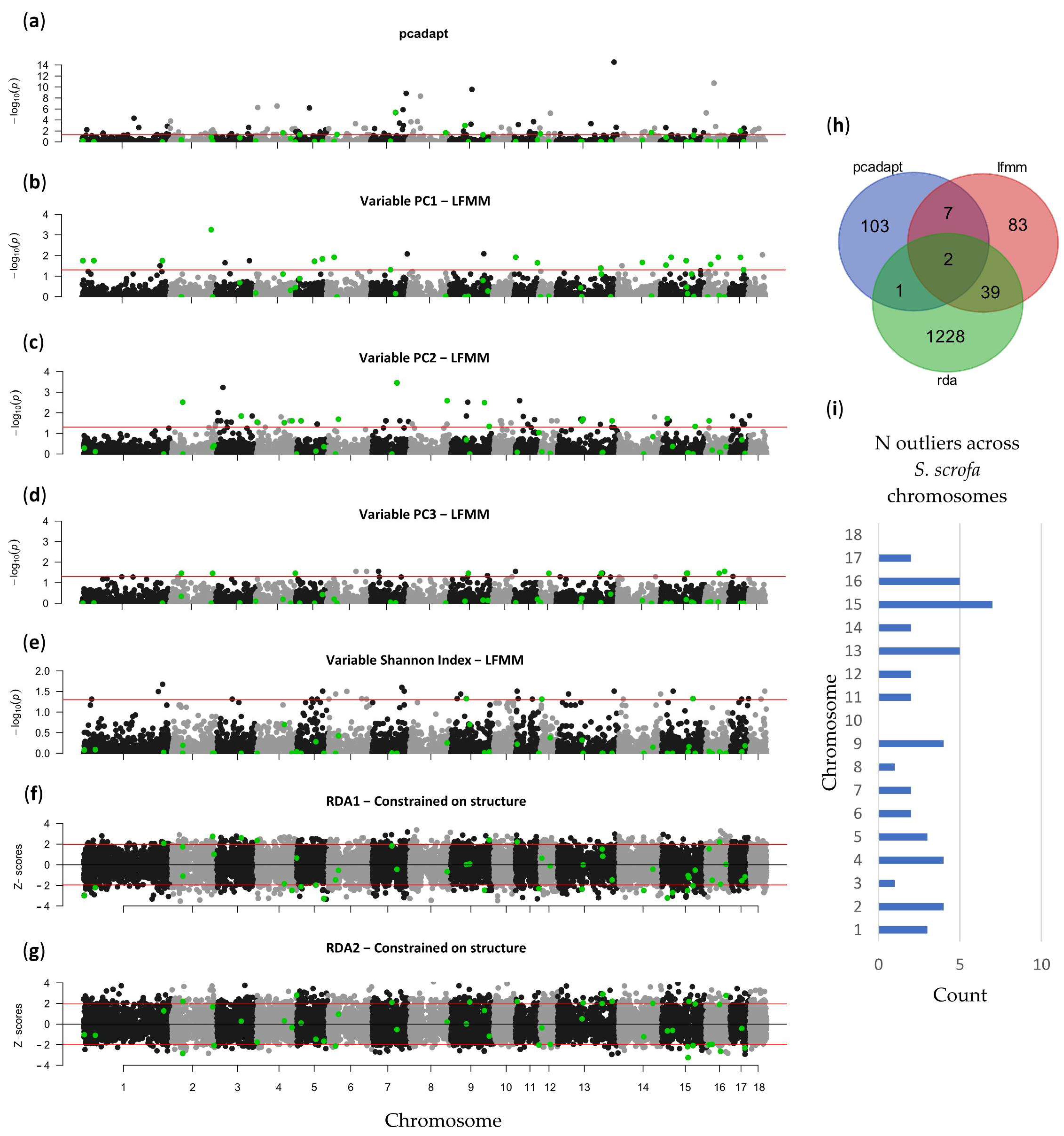
3.2. Local Adaptation
4. Discussion
4.1. Population Structure in the Sardinian Wild Boar
4.2. The Effect of the Environment on the Genetic Diversity of the Sardinian Wild Boar
4.3. Genomic Adaptation to Local Environmental Conditions
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Librado, P.; Orlando, L. Detecting Signatures of Positive Selection along Defined Branches of a Population Tree Using LSD. Mol. Biol. Evol. 2018, 35, 1520–1535. [Google Scholar] [CrossRef] [PubMed]
- Flori, L.; Moazami-Goudarzi, K.; Alary, V.; Araba, A.; Boujenane, I.; Boushaba, N.; Casabianca, F.; Casu, S.; Ciampolini, R.; D’Acier, A.C.; et al. A Genomic Map of Climate Adaptation in Mediterranean Cattle Breeds. Mol. Ecol. 2019, 28, 1009–1029. [Google Scholar] [CrossRef] [PubMed]
- Manthey, J.D.; Moyle, R.G. Isolation by Environment in White-Breasted Nuthatches (Sitta carolinensis) of the Madrean Archipelago Sky Islands: A Landscape Genomics Approach. Mol. Ecol. 2015, 24, 3628–3638. [Google Scholar] [CrossRef] [PubMed]
- Chávez-Galarza, J.; Henriques, D.; Johnston, J.S.; Azevedo, J.C.; Patton, J.C.; Muñoz, I.; De La Rúa, P.; Pinto, M.A. Signatures of Selection in the Iberian Honey Bee (Apis mellifera iberiensis) Revealed by a Genome Scan Analysis of Single Nucleotide Polymorphisms. Mol. Ecol. 2013, 22, 5890–5907. [Google Scholar] [CrossRef] [PubMed]
- Dalongeville, A.; Benestan, L.; Mouillot, D.; Lobreaux, S.; Manel, S. Combining Six Genome Scan Methods to Detect Candidate Genes to Salinity in the Mediterranean Striped Red Mullet (Mullus surmuletus). BMC Genom. 2018, 19, 1–13. [Google Scholar] [CrossRef]
- Friis, G.; Fandos, G.; Zellmer, A.J.; McCormack, J.E.; Faircloth, B.C.; Milá, B. Genome-Wide Signals of Drift and Local Adaptation during Rapid Lineage Divergence in a Songbird. Mol. Ecol. 2018, 27, 5137–5153. [Google Scholar] [CrossRef]
- Wang, I.J.; Bradburd, G.S. Isolation by Environment. Mol. Ecol. 2014, 23, 5649–5662. [Google Scholar] [CrossRef]
- Sexton, J.P.; Hangartner, S.B.; Hoffmann, A.A. Genetic Isolation by Environment or Distance: Which Pattern of Gene Flow Is Most Common? Evolution 2014, 68, 1–15. [Google Scholar] [CrossRef]
- Rellstab, C.; Gugerli, F.; Eckert, A.J.; Hancock, A.M.; Holderegger, R. A Practical Guide to Environmental Association Analysis in Landscape Genomics. Mol. Ecol. 2015, 24, 4348–4370. [Google Scholar] [CrossRef]
- Ahrens, C.W.; Rymer, P.D.; Stow, A.; Bragg, J.; Dillon, S.; Umbers, K.D.L.; Dudaniec, R.Y. The Search for Loci under Selection: Trends, Biases and Progress. Mol. Ecol. 2018, 27, 1342–1356. [Google Scholar] [CrossRef] [Green Version]
- Hohenlohe, P.A.; Bassham, S.; Currey, M.; Cresko, W.A. Extensive Linkage Disequilibrium and Parallel Adaptive Divergence across Threespine Stickleback Genomes. Philos. Trans. R. Soc. B Biol. Sci. 2012, 367, 395–409. [Google Scholar] [CrossRef] [PubMed]
- Felsenstein, J. Skepticism Towards Santa Rosalia, or Why Are There so Few Kinds of Animals? Evolution 1981, 35, 124. [Google Scholar] [CrossRef] [PubMed]
- Li, J.; Li, H.; Jakobsson, M.; Li, S.; Sjödin, P.; Lascoux, M. Joint Analysis of Demography and Selection in Population Genetics: Where Do We Stand and Where Could We Go? Mol. Ecol. 2012, 21, 28–44. [Google Scholar] [CrossRef]
- Privé, F.; Luu, K.; Vilhjálmsson, B.J.; Blum, M.G.B. Performing Highly Efficient Genome Scans for Local Adaptation with R Package Pcadapt Version 4. Mol. Biol. Evol. 2020, 37, 2153–2154. [Google Scholar] [CrossRef] [PubMed]
- Frichot, E.; Schoville, S.D.; Bouchard, G.; François, O. Testing for Associations between Loci and Environmental Gradients Using Latent Factor Mixed Models. Mol. Biol. Evol. 2013, 30, 1687–1699. [Google Scholar] [CrossRef] [PubMed]
- Gautier, M. Genome-Wide Scan for Adaptive Divergence and Association with Population-Specific Covariates. Genetics 2015, 201, 1555–1579. [Google Scholar] [CrossRef]
- Coop, G.; Witonsky, D.; Di Rienzo, A.; Pritchard, J.K. Using Environmental Correlations to Identify Loci Underlying Local Adaptation. Genetics 2010, 185, 1411–1423. [Google Scholar] [CrossRef]
- Stucki, S.; Orozco-TerWengel, P.; Forester, B.R.; Duruz, S.; Colli, L.; Masembe, C.; Negrini, R.; Landguth, E.; Jones, M.R.; Bruford, M.W.; et al. High Performance Computation of Landscape Genomic Models Including Local Indicators of Spatial Association. Mol. Ecol. Resour. 2017, 17, 1072–1089. [Google Scholar] [CrossRef]
- Morin, P.A.; Luikart, G.; Wayne, R.K. SNPs in Ecology, Evolution and Conservation. Trends Ecol. Evol. 2004, 19, 208–216. [Google Scholar] [CrossRef]
- Manel, S.; Perrier, C.; Pratlong, M.; Abi-Rached, L.; Paganini, J.; Pontarotti, P.; Aurelle, D. Genomic Resources and Their Influence on the Detection of the Signal of Positive Selection in Genome Scans. Mol. Ecol. 2016, 25, 170–184. [Google Scholar] [CrossRef] [Green Version]
- Hess, J.E.; Matala, A.P.; Narum, S.R. Comparison of SNPs and Microsatellites for Fine-Scale Application of Genetic Stock Identification of Chinook Salmon in the Columbia River Basin. Mol. Ecol. Resour. 2011, 11, 137–149. [Google Scholar] [CrossRef] [PubMed]
- Defaveri, J.; Viitaniemi, H.; Leder, E.; Merilä, J. Characterizing Genic and Nongenic Molecular Markers: Comparison of Microsatellites and SNPs. Mol. Ecol. Resour. 2013, 13, 377–392. [Google Scholar] [CrossRef] [PubMed]
- Narum, S.R.; Banks, M.; Beacham, T.D.; Bellinger, M.R.; Campbell, M.R.; Dekoning, J.; Elz, A.; Guthrie, C.M.; Kozfkay, C.; Miller, K.M.; et al. Differentiating Salmon Populations at Broad and Fine Geographical Scales with Microsatellites and Single Nucleotide Polymorphisms. Mol. Ecol. 2008, 17, 3464–3477. [Google Scholar] [CrossRef] [PubMed]
- Haasl, R.J.; Payseur, B.A. Multi-Locus Inference of Population Structure: A Comparison between Single Nucleotide Polymorphisms and Microsatellites. Heredity 2011, 106, 158–171. [Google Scholar] [CrossRef] [PubMed]
- Albarella, U.; Manconi, F.; Rowley-Conwy, P.; Vigne, J.-D. Pigs of Corsica and Sardinia: A Biometrical Re-Evaluation of Their Status and History. In Archaeozoological Studies in Honour of Alfredo Riedel; Tecchiati, U., Sala, B., Eds.; Ufficio Beni Archeologici: Bolzano, Italy, 2006; pp. 285–302. [Google Scholar]
- Mur, L.; Atzeni, M.; Martínez-López, B.; Feliziani, F.; Rolesu, S.; Sanchez-Vizcaino, J.M. Thirty-Five-Year Presence of African Swine Fever in Sardinia: History, Evolution and Risk Factors for Disease Maintenance. Transbound. Emerg. Dis. 2016, 63, e165–e177. [Google Scholar] [CrossRef]
- Albarella, U.; Dobney, K.; Rowley-Conwy, P. Size and Shape of the Eurasian Wild Boar (Sus scrofa), with a View to the Reconstruction of Its Holocene History. Environ. Archaeol. 2009, 14, 103–136. [Google Scholar] [CrossRef]
- Iacolina, L.; Scandura, M.; Goedbloed, D.J.; Alexandri, P.; Crooijmans, R.P.M.A.; Larson, G.; Archibald, A.; Apollonio, M.; Schook, L.B.; Groenen, M.A.M.; et al. Genomic Diversity and Differentiation of a Managed Island Wild Boar Population. Heredity 2016, 116, 60–67. [Google Scholar] [CrossRef]
- Scandura, M.; Fabbri, G.; Caniglia, R.; Iacolina, L.; Mattucci, F.; Mengoni, C.; Pante, G.; Apollonio, M.; Mucci, N. Resilience to Historical Human Manipulations in the Genomic Variation of Italian Wild Boar Populations. Front. Ecol. Evol. 2022, 10, 833081. [Google Scholar] [CrossRef]
- Petrelli, S.; Buglione, M.; Maselli, V.; Troiano, C.; Larson, G.; Frantz, L.; Manin, A.; Ricca, E.; Baccigalupi, L.; Wright, D.; et al. Population Genomic, Olfactory, Dietary, and Gut Microbiota Analyses Demonstrate the Unique Evolutionary Trajectory of Feral Pigs. Mol. Ecol. 2022, 31, 220–237. [Google Scholar] [CrossRef]
- Scandura, M.; Iacolina, L.; Cossu, A.; Apollonio, M. Effects of Human Perturbation on the Genetic Make-up of an Island Population: The Case of the Sardinian Wild Boar. Heredity 2011, 106, 1012–1020. [Google Scholar] [CrossRef] [Green Version]
- Lecis, R.; Dondina, O.; Orioli, V.; Biosa, D.; Canu, A.; Fabbri, G.; Iacolina, L.; Cossu, A.; Bani, L.; Apollonio, M.; et al. Main Roads and Land Cover Shaped the Genetic Structure of a Mediterranean Island Wild Boar Population. Ecol. Evol. 2022, 12, e8804. [Google Scholar] [CrossRef] [PubMed]
- Delitala, A.M.S.; Cesari, D.; Chiessa, P.A.; Ward, M.N. Precipitation over Sardinia (Italy) during the 1946–1993 Rainy Seasons and Associated Large-scale Climate Variations. Int. J. Climatol. 2000, 20, 519–541. [Google Scholar] [CrossRef]
- Canu, S.; Rosati, L.; Fiori, M.; Motroni, A.; Filigheddu, R.; Farris, E. Bioclimate Map of Sardinia (Italy). J. Maps 2015, 11, 711–718. [Google Scholar] [CrossRef]
- Muns, R.; Nuntapaitoon, M.; Tummaruk, P. Non-Infectious Causes of Pre-Weaning Mortality in Piglets. Livest. Sci. 2016, 184, 46–57. [Google Scholar] [CrossRef]
- Acevedo, P.; Escudero, M.A.; Muñoz, R.; Gortázar, C. Factors Affecting Wild Boar Abundance across an Environmental Gradient in Spain. Acta Theriol. 2006, 51, 327–336. [Google Scholar] [CrossRef]
- Brogi, R.; Merli, E.; Grignolio, S.; Chirichella, R.; Bottero, E.; Apollonio, M. It Is Time to Mate: Population-Level Plasticity of Wild Boar Reproductive Timing and Synchrony in a Changing Environment. Curr. Zool. 2021, 68, 371–380. [Google Scholar] [CrossRef]
- Gamelon, M.; Douhard, M.; Baubet, E.; Gimenez, O.; Brandt, S.; Gaillard, J.M. Fluctuating Food Resources Influence Developmental Plasticity in Wild Boar. Biol. Lett. 2013, 9, 20130419. [Google Scholar] [CrossRef]
- Fulgione, D.; Buglione, M. The Boar War: Five Hot Factors Unleashing Boar Expansion and Related Emergency. Land 2022, 11, 887. [Google Scholar] [CrossRef]
- Vetter, S.G.; Ruf, T.; Bieber, C.; Arnold, W. What Is a Mild Winter? Regional Differences in within-Species Responses to Climate Change. PLoS ONE 2015, 10, e0132178. [Google Scholar] [CrossRef]
- Cutini, A.; Chianucci, F.; Chirichella, R.; Donaggio, E.; Mattioli, L.; Apollonio, M. Mast Seeding in Deciduous Forests of the Northern Apennines (Italy) and Its Influence on Wild Boar Population Dynamics. Ann. For. Sci. 2013, 70, 493–502. [Google Scholar] [CrossRef] [Green Version]
- Bisi, F.; Chirichella, R.; Chianucci, F.; Von Hardenberg, J.; Cutini, A.; Martinoli, A.; Apollonio, M. Climate, Tree Masting and Spatial Behaviour in Wild Boar (Sus scrofa L.): Insight from a Long-Term Study. Ann. For. Sci. 2018, 75, 1–9. [Google Scholar] [CrossRef]
- Pungetti, G. Anthropological Approach to Agricultural Landscape History in Sardinia. Landsc. Urban Plan. 1995, 31, 47–56. [Google Scholar] [CrossRef]
- Puddu, G.; Falcucci, A.; Maiorano, L. Forest Changes over a Century in Sardinia: Implications for Conservation in a Mediterranean Hotspot. Agrofor. Syst. 2012, 85, 319–330. [Google Scholar] [CrossRef]
- Ramos, A.M.; Crooijmans, R.P.M.A.; Affara, N.A.; Amaral, A.J.; Archibald, A.L.; Beever, J.E.; Bendixen, C.; Churcher, C.; Clark, R.; Dehais, P.; et al. Design of a High Density SNP Genotyping Assay in the Pig Using SNPs Identified and Characterized by next Generation Sequencing Technology. PLoS ONE 2009, 4, e6524. [Google Scholar] [CrossRef]
- Purcell, S.; Neale, B.; Todd-Brown, K.; Thomas, L.; Ferreira, M.A.R.; Bender, D.; Maller, J.; Sklar, P.; De Bakker, P.I.W.; Daly, M.J.; et al. PLINK: A Tool Set for Whole-Genome Association and Population-Based Linkage Analyses. Am. J. Hum. Genet. 2007, 81, 559–575. [Google Scholar] [CrossRef]
- Patterson, N.; Price, A.L.; Reich, D. Population Structure and Eigenanalysis. PLoS Genet. 2006, 2, 2074–2093. [Google Scholar] [CrossRef]
- Alexander, D.H.; Novembre, J.; Lange, K. Fast Model-Based Estimation of Ancestry in Unrelated Individuals. Genome Res. 2009, 19, 1655–1664. [Google Scholar] [CrossRef]
- Jombart, T. Adegenet: A R Package for the Multivariate Analysis of Genetic Markers. Bioinformatics 2008, 24, 1403–1405. [Google Scholar] [CrossRef]
- Frichot, E.; François, O. LEA: An R Package for Landscape and Ecological Association Studies. Methods Ecol. Evol. 2015, 6, 925–929. [Google Scholar] [CrossRef]
- Fick, S.E.; Hijmans, R.J. WorldClim 2: New 1-Km Spatial Resolution Climate Surfaces for Global Land Areas. Int. J. Climatol. 2017, 37, 4302–4315. [Google Scholar] [CrossRef]
- Brogi, R.; Brivio, F.; Grignolio, S.; Cossu, A.; Apollonio, M. Behavior-Specific Resource Selection of the Sardinian Wild Boar. Manuscr. Prep. 2022. [Google Scholar]
- Bian, K.; Zhou, M.; Hu, F.; Lai, W. RF-PCA: A New Solution for Rapid Identification of Breast Cancer Categorical Data Based on Attribute Selection and Feature Extraction. Front. Genet. 2020, 11, 566057. [Google Scholar] [CrossRef] [PubMed]
- Oksanen, J.; Blanchet, F.G.; Friendly, M.; Kindt, R.; Legendre, P.; McGlinn, D.; Minchin, P.R.; O’Hara, B.; Simpson, G.L.; Solymos, P.; et al. Vegan: Community Ecology Package. In Package ‘Vegan’; CRAN: Evanston, IL, USA, 2020. [Google Scholar]
- Revelle, W. Psych: Procedures for Psychological, Psychometric, and Personality Research; R Package Version 1.0–95; CRAN: Evanston, IL, USA, 2021. [Google Scholar]
- Forester, B.R.; Lasky, J.R.; Wagner, H.H.; Urban, D.L. Comparing Methods for Detecting Multilocus Adaptation with Multivariate Genotype—Environment Associations. Mol. Ecol. 2018, 27, 2215–2233. [Google Scholar] [CrossRef] [PubMed]
- Zuur, A.F.; Ieno, E.N.; Elphick, C.S. A Protocol for Data Exploration to Avoid Common Statistical Problems. Methods Ecol. Evol. 2010, 1, 3–14. [Google Scholar] [CrossRef]
- Legendre, P.; Legendre, L. Numerical Ecology; Elsevier: Amsterdam, The Netherlands, 1998; ISBN 978044453868. [Google Scholar]
- Warr, A.; Affara, N.; Aken, B.; Beiki, H.; Bickhart, D.M.; Billis, K.; Chow, W.; Eory, L.; Finlayson, H.A.; Flicek, P.; et al. An Improved Pig Reference Genome Sequence to Enable Pig Genetics and Genomics Research. Gigascience 2020, 9, 1–14. [Google Scholar] [CrossRef]
- Cingolani, P.; Platts, A.; Wang, L.L.; Coon, M.; Nguyen, T.; Wang, L.; Land, S.J.; Lu, X.; Ruden, D.M. A Program for Annotating and Predicting the Effects of Single Nucleotide Polymorphisms, SnpEff: SNPs in the Genome of Drosophila Melanogaster Strain W1118; Iso-2; Iso-3. Fly 2012, 6, 80–92. [Google Scholar] [CrossRef]
- Lv, F.H.; Agha, S.; Kantanen, J.; Colli, L.; Stucki, S.; Kijas, J.W.; Joost, S.; Li, M.H.; Marsan, P.A. Adaptations to Climate-Mediated Selective Pressures in Sheep. Mol. Biol. Evol. 2014, 31, 3324–3343. [Google Scholar] [CrossRef]
- Raudvere, U.; Kolberg, L.; Kuzmin, I.; Arak, T.; Adler, P.; Peterson, H.; Vilo, J. G:Profiler: A Web Server for Functional Enrichment Analysis and Conversions of Gene Lists (2019 Update). Nucleic Acids Res. 2019, 47, 191–198. [Google Scholar] [CrossRef]
- Howe, K.L.; Achuthan, P.; Allen, J.; Allen, J.; Alvarez-Jarreta, J.; Amode, M.R.; Armean, I.M.; Azov, A.G.; Bennett, R.; Bhai, J.; et al. Ensembl 2021. Nucleic Acids Res. 2021, 49, D884–D891. [Google Scholar] [CrossRef]
- Gros, R.; Van Uum, S.; Hutchinson-Jaffe, A.; Ding, Q.; Pickering, J.G.; Hegele, R.A.; Feldman, R.D. Increased Enzyme Activity and β-Adrenergic-Mediated Vasodilation in Subjects Expressing a Single-Nucleotide Variant of Human Adenylyl Cyclase 6. Arterioscler. Thromb. Vasc. Biol. 2007, 27, 2657–2663. [Google Scholar] [CrossRef]
- Singh, U.; Sun, T.; Looman, C.; Heuchel, R.; Elliott, R.; Freichel, M.; Meissner, M.; Flockerzi, V.; Fundele, R. Expression and Function of the Gene Encoding the Voltage-Dependent Calcium Channel Β3-Subunit in the Mouse Placenta. Placenta 2007, 28, 412–420. [Google Scholar] [CrossRef] [PubMed]
- De Chu, Y.; Chen, H.K.; Huang, T.; Chan, S.P. A Novel Function for the DEAD-Box RNA Helicase DDX-23 in Primary MicroRNA Processing in Caenorhabditis elegans. Dev. Biol. 2016, 409, 459–472. [Google Scholar] [CrossRef]
- Nobes, C.D.; Lauritzen, I.; Mattei, M.G.; Paris, S.; Hall, A.; Chardin, P. A New Member of the Rho Family, Rnd1, Promotes Disassembly of Actin Filament Structures and Loss of Cell Adhesion. J. Cell Biol. 1998, 141, 187–197. [Google Scholar] [CrossRef] [PubMed]
- Ren, G.; Vajjhala, P.; Lee, J.S.; Winsor, B.; Munn, A.L. The BAR Domain Proteins: Molding Membranes in Fission, Fusion, and Phagy. Microbiol. Mol. Biol. Rev. 2006, 70, 37–120. [Google Scholar] [CrossRef] [PubMed]
- Bassett, J.H.D.; Gogakos, A.; White, J.K.; Evans, H.; Jacques, R.M.; van der Spek, A.H.; Ramirez-Solis, R.; Ryder, E.; Sunter, D.; Boyde, A.; et al. Rapid-Throughput Skeletal Phenotyping of 100 Knockout Mice Identifies 9 New Genes That Determine Bone Strength. PLoS Genet. 2012, 8, e1002858. [Google Scholar] [CrossRef] [PubMed]
- Wang, H.; Wang, C.; Yang, K.; Liu, J.; Zhang, Y.; Wang, Y.; Xu, X.; Michal, J.J.; Jiang, Z.; Liu, B. Genome Wide Distributions and Functional Characterization of Copy Number Variations between Chinese and Western Pigs. PLoS ONE 2015, 10, e0131522. [Google Scholar] [CrossRef]
- Zhang, X.; Sun, W.; He, L.; Wang, L.; Qiu, K.; Yin, J. Global DNA Methylation Pattern Involved in the Modulation of Differentiation Potential of Adipogenic and Myogenic Precursors in Skeletal Muscle of Pigs. Stem Cell Res. Ther. 2020, 11, 536. [Google Scholar] [CrossRef]
- Tafforeau, L.; Chantier, T.; Pradezynski, F.; Pellet, J.; Mangeot, P.E.; Vidalain, P.-O.; Andre, P.; Rabourdin-Combe, C.; Lotteau, V. Generation and Comprehensive Analysis of an Influenza Virus Polymerase Cellular Interaction Network. J. Virol. 2011, 85, 13010–13018. [Google Scholar] [CrossRef]
- Alonso, I.; Ibáñez-Escriche, N.; Noguera, J.L.; Casellas, J.; de Hijas-Villalba, M.M.; Gracia-Santana, M.J.; Varona, L. Genomic Differentiation among Varieties of Iberian Pig. Span. J. Agric. Res. 2020, 18, e0401. [Google Scholar] [CrossRef]
- Tang, Z.; Yang, Y.; Wang, Z.; Zhao, S.; Mu, Y.; Li, K. Integrated Analysis of MiRNA and MRNA Paired Expression Profiling of Prenatal Skeletal Muscle Development in Three Genotype Pigs. Sci. Rep. 2015, 5, 15544. [Google Scholar] [CrossRef]
- Mochida, G.H.; Ganesh, V.S.; Felie, J.M.; Gleason, D.; Hill, R.S.; Clapham, K.R.; Rakiec, D.; Tan, W.H.; Akawi, N.; Al-Saffar, M.; et al. A Homozygous Mutation in the Tight-Junction Protein JAM3 Causes Hemorrhagic Destruction of the Brain, Subependymal Calcification, and Congenital Cataracts. Am. J. Hum. Genet. 2010, 87, 882–889. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Santoso, S.; Sachs, U.J.H.; Kroll, H.; Linder, M.; Ruf, A.; Preissner, K.T.; Chavakis, T. The Junctional Adhesion Molecule 3 (JAM-3) on Human Platelets Is a Counterreceptor for the Leukocyte Integrin Mac-1. J. Exp. Med. 2002, 196, 679–691. [Google Scholar] [CrossRef] [PubMed]
- Arrate, M.P.; Rodriguez, J.M.; Tran, T.M.; Brock, T.A.; Cunningham, S.A. Cloning of Human Junctional Adhesion Molecule 3 (JAM3) and Its Identification as the JAM2 Counter-Receptor. J. Biol. Chem. 2001, 276, 45826–45832. [Google Scholar] [CrossRef]
- Kulkarni, S.; Sitaru, C.; Jakus, Z.; Anderson, K.E.; Damoulakis, G.; Davidson, K.; Hirose, M.; Juss, J.; Oxley, D.; Chessa, T.A.M.; et al. PI3Kβ Plays a Critical Role in Neutrophil Activation by Immune Complexes. Sci. Signal. 2011, 4, ra23. [Google Scholar] [CrossRef] [PubMed]
- Huo, J.; Xu, S.; Lam, K.P. Faim: An Antagonist of Fas-Killing and Beyond. Cells 2019, 8, 541. [Google Scholar] [CrossRef]
- Takeuchi, A.; Badr, M.E.S.G.; Miyauchi, K.; Ishihara, C.; Onishi, R.; Guo, Z.; Sasaki, Y.; Ike, H.; Takumi, A.; Tsuji, N.M.; et al. CRTAM Determines the CD4+ Cytotoxic T Lymphocyte Lineage. J. Exp. Med. 2016, 213, 123–138. [Google Scholar] [CrossRef]
- Fuller, J.; Ferchaud, A.-L.; Laporte, M.; Le Luyer, J.; Davis, T.B.; Côté, S.D.; Bernatchez, L. Absence of Founder Effect and Evidence for Adaptive Divergence in a Recently Introduced Insular Population of White-Tailed Deer (Odocoileus Virginianus). Mol. Ecol. 2019, 29, 86–104. [Google Scholar] [CrossRef]
- Biello, R.; Zampiglia, M.; Fuselli, S.; Fabbri, G.; Bisconti, R.; Chiocchio, A.; Mazzotti, S.; Trucchi, E.; Canestrelli, D.; Bertorelle, G. From STRs to SNPs via DdRAD-Seq: Geographic Assignment of Confiscated Tortoises at Reduced Costs. bioRxiv 2022. Evolutionary application is the journal. [Google Scholar] [CrossRef]
- Cayuela, H.; Rougemont, Q.; Laporte, M.; Mérot, C.; Normandeau, E.; Dorant, Y.; Tørresen, O.K.; Hoff, S.N.K.; Jentoft, S.; Sirois, P.; et al. Shared Ancestral Polymorphisms and Chromosomal Rearrangements as Potential Drivers of Local Adaptation in a Marine Fish. Mol. Ecol. 2020, 29, 2379–2398. [Google Scholar] [CrossRef]
- Flanagan, S.P.; Forester, B.R.; Latch, E.K.; Aitken, S.N.; Hoban, S. Guidelines for Planning Genomic Assessment and Monitoring of Locally Adaptive Variation to Inform Species Conservation. Evol. Appl. 2018, 11, 1035–1052. [Google Scholar] [CrossRef]
- Gibson, M.J.S.; Moyle, L.C. Regional Differences in the Abiotic Environment Contribute to Genomic Divergence within a Wild Tomato Species. Mol. Ecol. 2020, 29, 2204–2217. [Google Scholar] [CrossRef] [PubMed]
- Massei, G.; Kindberg, J.; Licoppe, A.; Gačić, D.; Šprem, N.; Kamler, J.; Baubet, E.; Hohmann, U.; Monaco, A.; Ozoliņš, J.; et al. Wild Boar Populations up, Numbers of Hunters down? A Review of Trends and Implications for Europe. Pest Manag. Sci. 2015, 71, 492–500. [Google Scholar] [CrossRef] [PubMed]
- Stern, D.B.; Lee, C.E. Evolutionary Origins of Genomic Adaptations in an Invasive Copepod. Nat. Ecol. Evol. 2020, 4, 1084–1094. [Google Scholar] [CrossRef] [PubMed]
- Viscardi, L.H.; Paixão-Côrtes, V.R.; Comas, D.; Salzano, F.M.; Rovaris, D.; Bau, C.D.; Amorim, C.E.G.; Bortolini, M.C. Searching for Ancient Balanced Polymorphisms Shared between Neanderthals and Modern Humans. Genet. Mol. Biol. 2018, 41, 67–81. [Google Scholar] [CrossRef]
- Groenen, M.A.M.; Archibald, A.L.; Uenishi, H.; Tuggle, C.K.; Takeuchi, Y.; Rothschild, M.F.; Rogel-Gaillard, C.; Park, C.; Milan, D.; Megens, H.J.; et al. Analyses of Pig Genomes Provide Insight into Porcine Demography and Evolution. Nature 2012, 491, 393–398. [Google Scholar] [CrossRef]
- Franks, S.J.; Hoffmann, A.A. Genetics of Climate Change Adaptation. Annu. Rev. Genet. 2012, 46, 185–208. [Google Scholar] [CrossRef]
- Peng, W.F.; Xu, S.S.; Ren, X.; Lv, F.H.; Xie, X.L.; Zhao, Y.X.; Zhang, M.; Shen, Z.Q.; Ren, Y.L.; Gao, L.; et al. A Genome-Wide Association Study Reveals Candidate Genes for the Supernumerary Nipple Phenotype in Sheep (Ovis aries). Anim. Genet. 2017, 48, 570–579. [Google Scholar] [CrossRef]
- Arakawa, A.; Okumura, N.; Taniguchi, M.; Hayashi, T.; Hirose, K.; Fukawa, K.; Ito, T.; Matsumoto, T.; Uenishi, H.; Mikawa, S. Genome-Wide Association QTL Mapping for Teat Number in a Purebred Population of Duroc Pigs. Anim. Genet. 2015, 46, 571–575. [Google Scholar] [CrossRef]
- Gardner, J.L.; Peters, A.; Kearney, M.R.; Joseph, L.; Heinsohn, R. Declining Body Size: A Third Universal Response to Warming? Trends Ecol. Evol. 2011, 26, 285–291. [Google Scholar] [CrossRef]
- Zhao, X.; Mo, D.; Li, A.; Gong, W.; Xiao, S.; Zhang, Y.; Qin, L.; Niu, Y.; Guo, Y.; Liu, X.; et al. Comparative Analyses by Sequencing of Transcriptomes during Skeletal Muscle Development between Pig Breeds Differing in Muscle Growth Rate and Fatness. PLoS ONE 2011, 6, e19774. [Google Scholar] [CrossRef]
- Savoldi, I.R.; Ibelli, A.M.G.; Cantão, M.E.; de Peixoto, J.O.; Pires, M.P.; Mores, M.A.Z.; Lagos, E.B.; Lopes, J.S.; Zanella, R.; Ledur, M.C. A Joint Analysis Using Exome and Transcriptome Data Identifies Candidate Polymorphisms and Genes Involved with Umbilical Hernia in Pigs. BMC Genom. 2021, 22, 818. [Google Scholar] [CrossRef] [PubMed]
- Ben-Jemaa, S.; Senczuk, G.; Ciani, E.; Ciampolini, R.; Catillo, G.; Boussaha, M.; Pilla, F.; Portolano, B.; Mastrangelo, S. Genome-Wide Analysis Reveals Selection Signatures Involved in Meat Traits and Local Adaptation in Semi-Feral Maremmana Cattle. Front. Genet. 2021, 12, 675569. [Google Scholar] [CrossRef] [PubMed]
- Scandura, M.; Iacolina, L.; Apollonio, M. Genetic Diversity in the European Wild Boar Sus scrofa: Phylogeography, Population Structure and Wild x Domestic Hybridization. Mamm. Rev. 2011, 41, 125–137. [Google Scholar] [CrossRef]
- Maselli, V.; Rippa, D.; De Luca, A.; Larson, G.; Wilkens, B.; Linderholm, A.; Masseti, M.; Fulgione, D. Southern Italian Wild Boar Population, Hotspot of Genetic Diversity. Hystrix It. J. Mammal. 2016, 27, 137–144. [Google Scholar] [CrossRef]
- Frantz, L.A.F.; Haile, J.; Lin, A.T.; Scheu, A.; Geörg, C.; Benecke, N.; Alexander, M.; Linderholm, A.; Mullin, V.E.; Daly, K.G.; et al. Ancient Pigs Reveal a Near-Complete Genomic Turnover Following Their Introduction to Europe. Proc. Natl. Acad. Sci. USA 2020, 117, 14610–14611. [Google Scholar] [CrossRef]
- Fuller, A.; Mitchell, D.; Maloney, S.K.; Hetem, R.S. Towards a Mechanistic Understanding of the Responses of Large Terrestrial Mammals to Heat and Aridity Associated with Climate Change. Clim. Chang. Responses 2016, 3, 10. [Google Scholar] [CrossRef] [Green Version]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Variable Name | Description | Source |
|---|---|---|
| Bio4 | Temperature Seasonality (standard deviation ×100) | Worldclim2 |
| Bio5 | Max Temperature of Warmest Month | Worldclim2 |
| Bio9 | Mean Temperature of Driest Quarter | Worldclim2 |
| Bio10 | Mean Temperature of Warmest Quarter | Worldclim2 |
| Bio12 | Annual Precipitation | Worldclim2 |
| Bio15 | Precipitation Seasonality (Coefficient of Variation) | Worldclim2 |
| Bio16 | Precipitation of Wettest Quarter | Worldclim2 |
| Bio17 | Precipitation of Driest Quarter | Worldclim2 |
| Bio18 | Precipitation of Warmest Quarter | Worldclim2 |
| Elevation | Elevation derived from the SRTM data | Worldclim2 |
| Shannon Index | Diversity index calculated on land use categories | SardegnaGeoportale |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Fabbri, G.; Iacolina, L.; Apollonio, M.; Scandura, M. The Role of the Environment in Shaping the Genomic Variation in an Insular Wild Boar Population. Diversity 2022, 14, 774. https://doi.org/10.3390/d14090774
Fabbri G, Iacolina L, Apollonio M, Scandura M. The Role of the Environment in Shaping the Genomic Variation in an Insular Wild Boar Population. Diversity. 2022; 14(9):774. https://doi.org/10.3390/d14090774
Chicago/Turabian StyleFabbri, Giulia, Laura Iacolina, Marco Apollonio, and Massimo Scandura. 2022. "The Role of the Environment in Shaping the Genomic Variation in an Insular Wild Boar Population" Diversity 14, no. 9: 774. https://doi.org/10.3390/d14090774