
Trends in Taxonomy of the Rhodniini Tribe (Hemiptera, Triatominae): Reproductive Incompatibility between Rhodnius neglectus Lent, 1954 and Psammolestes spp. Confirms the Generic Status of Psammolestes Bergroth, 1911

 , ,
, ,  ,
,
Abstract
:1. Introduction
2. Materials and Methods
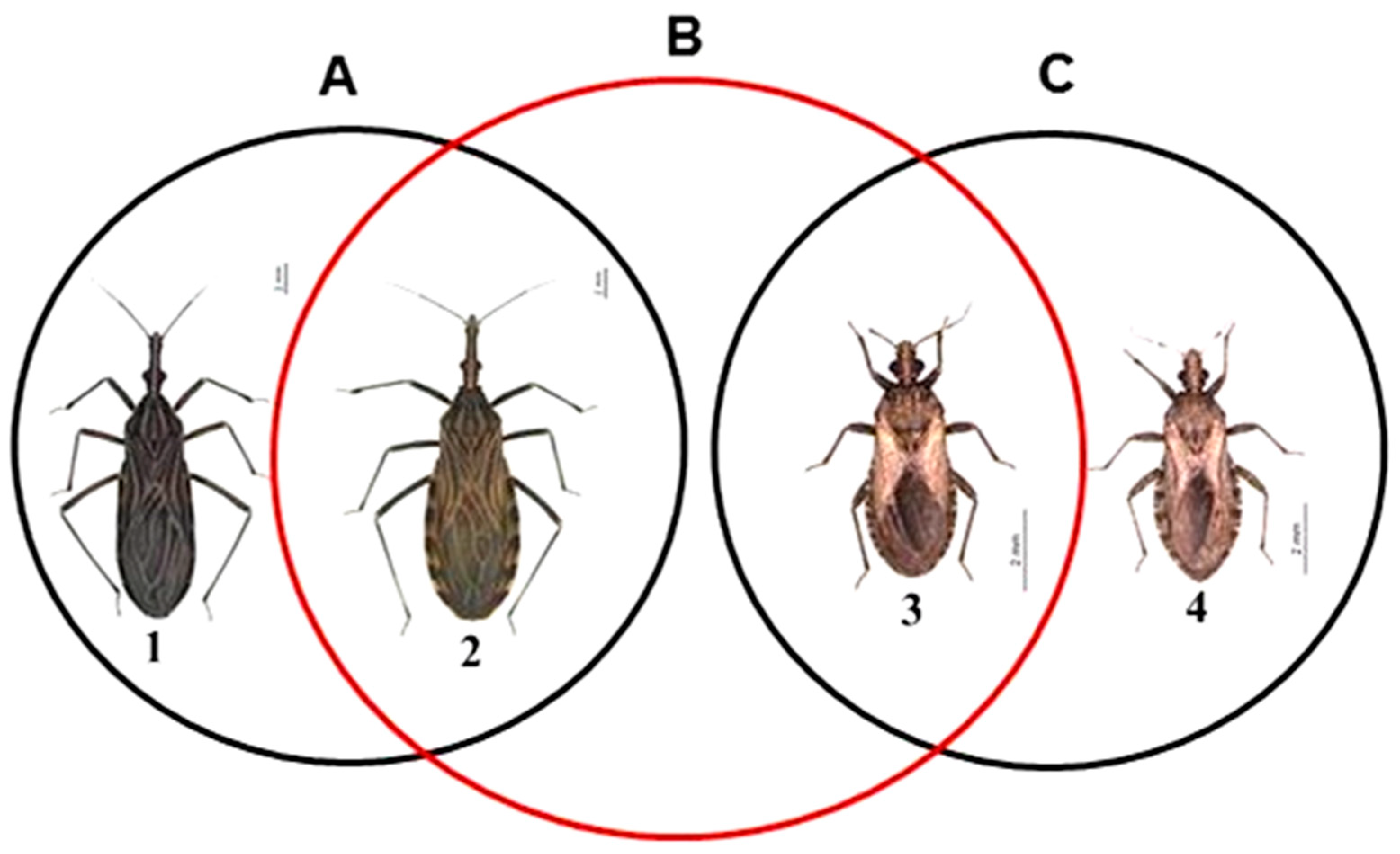
3. Results and Discussion
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- World Health Organization. Available online: https://www.who.int/health-topics/chagas-disease#tab=tab_1 (accessed on 23 March 2022).
- Pan American Health Organization. Available online: https://www.paho.org/en/topics/chagas-disease (accessed on 23 March 2022).
- Alevi, K.C.C.; Oliveira, J.; Rocha, D.S.; Galvão, C. Trends in Taxonomy of Chagas Disease Vectors (Hemiptera, Reduviidae, Triatominae): From Linnaean to Integrative Taxonomy. Pathogens 2021, 10, 1627. [Google Scholar] [CrossRef] [PubMed]
- Oliveira Correia, J.P.S.; Gil-Santana, H.R.; Dale, C.; Galvão, C. Triatoma guazu Lent and Wygodzinsky Is a Junior Synonym of Triatoma williami Galvão, Souza and Lima. Insects 2022, 13, 591. [Google Scholar] [CrossRef] [PubMed]
- Gil-Santana, H.R.; Chavez, T.; Pita, S.; Panzera, F.; Galvão, C. Panstrongylus noireaui, a remarkable new species of Triatominae (Hemiptera, Reduviidae) from Bolivia. ZooKeys 2022, 1104, 203–225. [Google Scholar] [CrossRef]
- Lent, H.; Wygodzinsky, P. Revision of the Triatominae (Hemiptera: Reduviidae) and their significance as vector of Chagas’s disease. Bull. Am. Mus. Nat. Hist. 1979, 163, 123–520. [Google Scholar]
- Gurgel-Gonçalves, R.; Duarte, M.A.; Dias, R.E.; Torre, P.A.R.; Romaña, C.A.; Cuba-Cuba, C.A. Spatial distribution of Triatominae populations (Hemiptera: Reduviidae) in Mauritia flexuosa palm trees in Federal District of Brazil. Rev. Soc. Bras. Med. Trop. 2004, 37, 241–247. [Google Scholar] [CrossRef]
- Cruz-Guzmán, P.; Morocoima, A.; Chique, J.; RamonisQuintero, J.; Uzcátegui, M.; Carrasco, H. Psammolestes arthuri naturally infected with Trypanosoma cruzi found in sympatry with Rhodnius prolixus and Triatoma maculata on bird nests in Anzoátegui State, Venezuela. Saber 2014, 26, 428–440. [Google Scholar]
- Justi, S.; Galvão, C. The evolutionary origin of diversity in Chagas disease vectors. Trends Parasitol. 2017, 33, 42–52. [Google Scholar] [CrossRef]
- Justi, S.A.; Galvão, C.; Schrago, C.G. Geological changes of the Americas and their influence on the diversification of the Neotropical kissing bugs (Hemiptera: Reduviidae: Triatominae). PLoS Negl. Trop. Dis. 2016, 10, e0004527. [Google Scholar] [CrossRef]
- de Paula, A.S.; Barreto, C.; Telmo, M.C.M.; Diotaiuti, L.; Galvão, C. Historical biogeography and the evolution of hematophagy in Rhodniini (Heteroptera: Reduviidae: Triatominae). Front. Ecol. Evol. 2021, 9, 660151. [Google Scholar] [CrossRef]
- Filée, J.; Merle, M.; Bastide, H.; Mougel, F.; Bérenger, J.M.; Folly-Ramos, E.; Almeida, C.E.; Harry, M. Phylogenomics for Chagas Disease Vectors of the Rhodnius Genus (Hemiptera, Triatominae): What We Learn From Mito-Nuclear Conflicts and Recommendations. Front. Ecol. Evol. 2022, 9, 750317. [Google Scholar] [CrossRef]
- Hypsa, V.; Tietz, D.; Zrzavy, J.; Rego, R.O.; Galvão, C.; Jurberg, J. Phylogeny and biogeography of Triatominae (Hemiptera, Reduviidae): Molecular evidence of a New World origin of the asiatic clade. Mol. Phylog. Evol. 2012, 23, 447–457. [Google Scholar] [CrossRef]
- Cracraft, J. Species Concepts and Speciation Analysis. Curr. Ornithol. 1983, 1, 159–187. [Google Scholar]
- de Queiroz, K. Species concepts and species delimitation. Syst. Biol. 2007, 56, 879–886. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mayr, E. Populações, Espécies e Evolução, 1st ed.; Editora Nacional: São Paulo, Brasil, 1963; 485p. [Google Scholar]
- Mayr, E. Populations, Species, and Evolution; Harvard University Press: Cambridge, MA, USA, 1970; 453p. [Google Scholar]
- Arnold, M.L. Natural Hybridization and Evolution; Oxford University Press: Oxford, MS, USA, 1997; 215p. [Google Scholar]
- Pérez, R.; Hernández, M.; Quintero, O.; Scovortzoff, E.; Canale, D.; Méndez, L.; Cohanoff, C.; Martino, M.; Panzera, F. Cytogenetic analysis of experimental hybrids in species of Triatominae (Hemiptera-Reduviidae). Genetica 2005, 125, 261–270. [Google Scholar] [CrossRef]
- Neves, S.J.M.; Sousa, P.S.; Oliveira, J.; Ravazi, A.; Madeira, F.F.; Reis, Y.V. Prezygotic isolation confirms the exclusion of Triatoma melanocephala, T. vitticeps and T. tibiamaculata of the T. brasiliensis subcomplex (Hemiptera, Triatominae). Infect. Genet. Evol. 2020, 79, 104149. [Google Scholar] [CrossRef]
- Ravazi, A.; Oliveira, J.; Campos, F.F.; Madeira, F.F.; Reis, Y.V.; Oliveira, A.B.B.; Azeredo-Oliveira, M.T.V.; Rosa, J.A.; Galvão, C.; Alevi, K.C.C. Trends in evolution of the Rhodniini tribe (Hemiptera, Triatominae): Experimental crosses between Psammolestes tertius Lent & Jurberg, 1965 and P. coreodes Bergroth, 1911 and analysis of the reproductive isolating mechanisms. Parasit. Vec. 2021, 14, 350. [Google Scholar]
- Rosa, J.A.; Barata, J.M.S.; Barelli, N.; Santos, J.L.F.; Belda Neto, F.M. Sexual distinction between 5th instar nymphs of six species (Hemiptera: Reduviidae). Mem. Inst. Oswaldo Cruz 1992, 87, 257–264. [Google Scholar] [CrossRef]
- Martínez-Ibarra, J.A.; Grant-Guillén, Y.; Delgadillo-Aceves, I.N.; Zumaya-Estrada, F.A.; Rocha-Chávez, G.; Salazar-Schettino, P.M.; Alejandre-Aguilar, R.; Villalobos, G.; Nogueda-Torres, B. Biological and genetic aspects of crosses between phylogenetically close species of Mexican Triatomines (Hemiptera: Reduviidae). J. Med. Entomol. 2011, 48, 705–707. [Google Scholar] [CrossRef]
- Carcavallo, R.U.; Girón, I.G.; Jurberg, J.; Galvao, C.; Noireau, F.; Canale, D. Mutations, Hybrids and Teratologies. In Atlas of Chagas Disease Vectors in the Americas; Carcavallo, R.U., Girón, I.G., Jurberg, J., Lent, H., Eds.; Editora FIOCRUZ: Rio de Janeiro, Brazil, 1998; pp. 515–536, Coleção de Triatominae da FCFAR–UNESP Araraquara; Available online: https://www2.fcfar.unesp.br/#!/triatominae (accessed on 9 July 2022).
- Perlowagora-Szumlewics, A.; Correia, M.V. Induction of male sterility manipulation of genetic mechanisms present in vector species of Chagas disease (remarks on integrating sterile-male release with insecticidal control measures against vectors of Chagas disease). Rev. Inst. Med. Trop São Paulo 1972, 14, 360–371. [Google Scholar]
- Cesaretto, N.R.; Oliveira, J.; Ravazi, A.; Madeira, F.F.; Reis, Y.V.; Oliveira, A.B.B.; Vicente, R.D.; Cristal, D.C.; Galvão, C.; Azeredo-Oliveira, M.T.V.; et al. Trends in taxonomy of Triatomini (Hemiptera, Reduviidae, Triatominae): Reproductive compatibility reinforces the synonymization of Meccus Stål, 1859 with Triatoma Laporte, 1832. Parasit Vect. 2021, 14, 340. [Google Scholar] [CrossRef]
- Alevi, K.C.C.; Moreira, F.F.F.; Jurberg, J.; Azeredo-Oliveira, M.T.V. Description of the diploid chromosome set of Triatoma pintodiasi (Hemiptera, Triatominae). Genet. Mol. Res. 2015, 15, gmr.15026343. [Google Scholar] [CrossRef]
- Alevi, K.C.C.; Oliveira, J.; Rosa, J.A.; Azeredo-Oliveira, M.T.V. Karyotype Evolution of Chagas Disease Vectors (Hemiptera, Triatominae). Am. J. Trop Med. Hyg. 2018, 99, 87–89. [Google Scholar] [CrossRef]
- Pita, S.; Panzera, F.; Ferrandis, I.; Galvão, C.; Gómez-Palacio, A.; Panzera, Y. Chromosomal divergence and evolutionary inferences in Rhodniini based on the chromosomal location of ribosomal genes. Mem. Inst. Oswaldo Cruz. 2013, 108, 376–382. [Google Scholar] [CrossRef]
- Pita, S.; Lorite, P.; Cuadrado, A.; Panzera, Y.; Oliveira, J.; Alevi, K.C.C.; Rosa, J.A.; Freitas, S.P.C.; Gómez-Palacio, A.; Solari, A.; et al. High chromosomal mobility of ribosomal clusters in holocentric chromosomes of Triatominae, vectors of Chagas disease (Hemiptera-Reduviidae). Med. Vet. Entomol. 2022, 36, 66–80. [Google Scholar] [CrossRef]


{kind=link}
| Experimental Crosses | Number of Eggs | ||||
|---|---|---|---|---|---|
| C1 | C2 | C3 | Total | Egg Fertility | |
| P. coreodes ♀ x R. neglectus ♂ | 55 | 74 | 52 | 181 | 0% |
| R. neglectus ♀ x P. coreodes ♂ | 60 | 121 | 156 | 337 | 0% |
| P. tertius ♀ x R. neglectus ♂ | 1 | 26 | 28 | 55 | 0% |
| R. neglectus ♀ x P. tertius ♂ | 114 | 130 | 97 | 341 | 0% |
| Control experiments | |||||
| P. coreodes ♀ x P. coreodes ♂ | 61 | 10 | 90 | 161 | 67.7% |
| P. tertius ♀ x P. tertius ♂ | 52 | 47 | 75 | 174 | 72.3% |
| R. neglectus ♀ x R. neglectus ♂ | 337 | 409 | 175 | 901 | 89.8% |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ravazi, A.; Oliveira, J.d.; Madeira, F.F.; Reis, Y.V.d.; Oliveira, A.B.B.d.; Galvão, C.; Azeredo-Oliveira, M.T.V.d.; Rosa, J.A.d.; Alevi, K.C.C. Trends in Taxonomy of the Rhodniini Tribe (Hemiptera, Triatominae): Reproductive Incompatibility between Rhodnius neglectus Lent, 1954 and Psammolestes spp. Confirms the Generic Status of Psammolestes Bergroth, 1911. Diversity 2022, 14, 761. https://doi.org/10.3390/d14090761
Ravazi A, Oliveira Jd, Madeira FF, Reis YVd, Oliveira ABBd, Galvão C, Azeredo-Oliveira MTVd, Rosa JAd, Alevi KCC. Trends in Taxonomy of the Rhodniini Tribe (Hemiptera, Triatominae): Reproductive Incompatibility between Rhodnius neglectus Lent, 1954 and Psammolestes spp. Confirms the Generic Status of Psammolestes Bergroth, 1911. Diversity. 2022; 14(9):761. https://doi.org/10.3390/d14090761
Chicago/Turabian StyleRavazi, Amanda, Jader de Oliveira, Fernanda Fernandez Madeira, Yago Visinho dos Reis, Ana Beatriz Bortolozo de Oliveira, Cleber Galvão, Maria Tercília Vilela de Azeredo-Oliveira, João Aristeu da Rosa, and Kaio Cesar Chaboli Alevi. 2022. "Trends in Taxonomy of the Rhodniini Tribe (Hemiptera, Triatominae): Reproductive Incompatibility between Rhodnius neglectus Lent, 1954 and Psammolestes spp. Confirms the Generic Status of Psammolestes Bergroth, 1911" Diversity 14, no. 9: 761. https://doi.org/10.3390/d14090761