

Protective Effects of Theaflavins and Epigallocatechin Gallate against ZnO-NP-Induced Cell Apoptosis In Vitro

 and
and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Materials
2.2. Preparation of ZnO-NPs Working Solution
2.3. Cell Culture and ZnO-NP Exposure
2.4. Cell Proliferation Inhibition Efficiency
2.5. Detection of Zn in RTE Cells
2.6. Measurement of Intracellular Reactive Oxygen Species (ROS) and Malondialdehyde (MDA) Concentration
2.7. RNA Isolation and Quantitative Real-Time Polymerase Chain Reaction (qPCR)
2.8. Statistical Analysis
3. Results
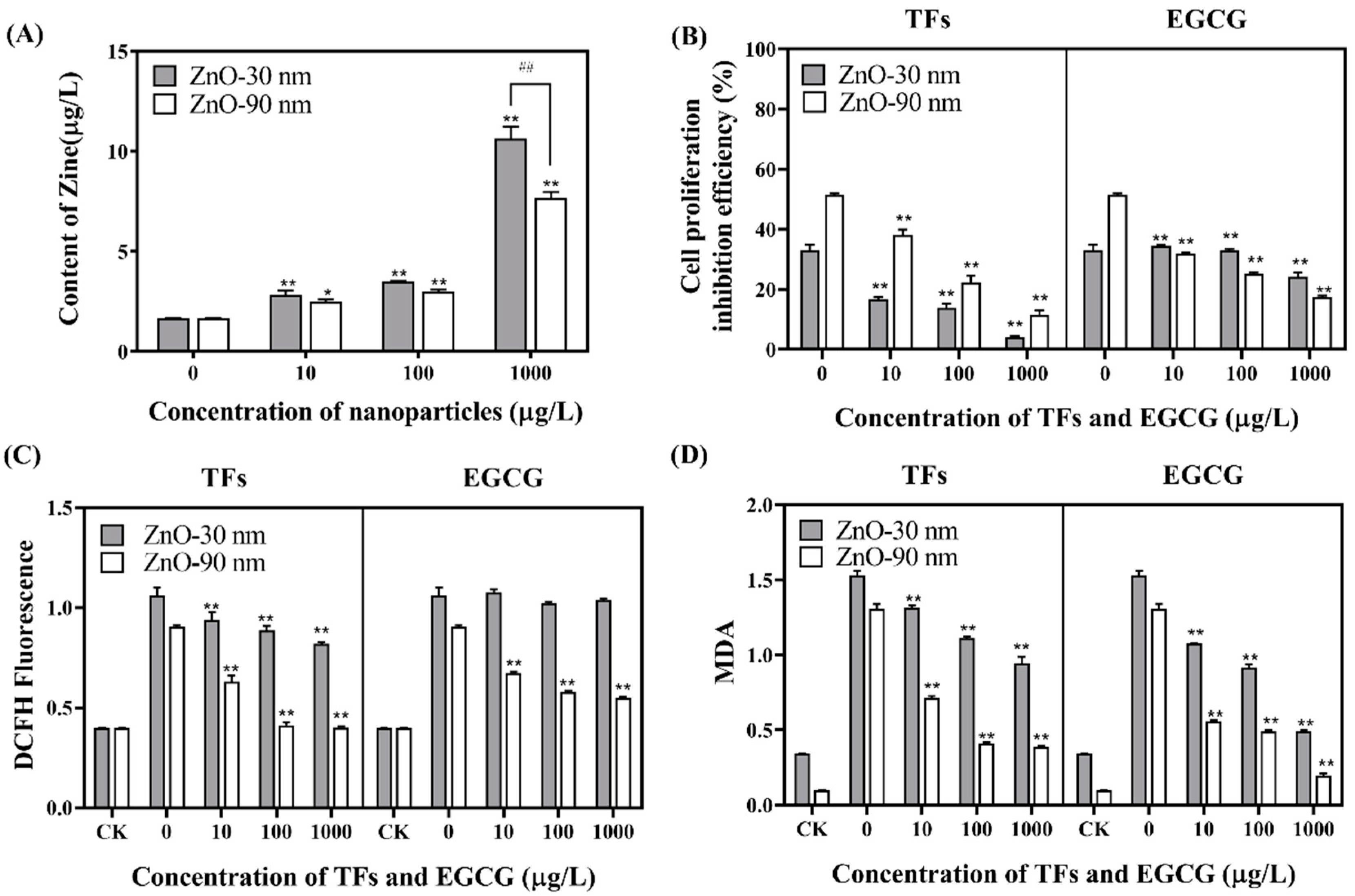
3.1. Uptake of Zinc by RTE Cells and the Proliferation Inhibition Efficiency of RTE Cells
3.2. Regulation Effects of ROS and MDA Generation
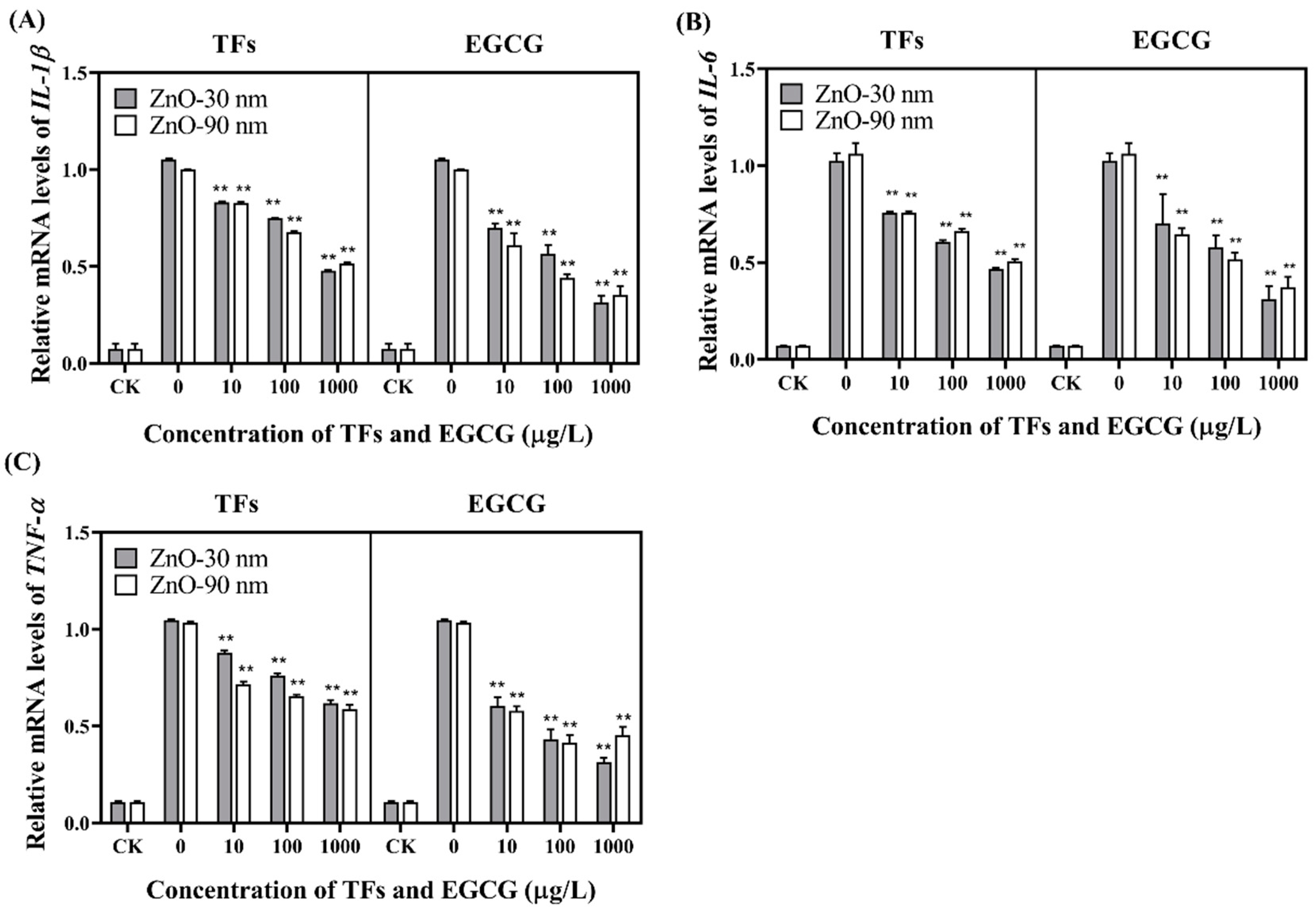
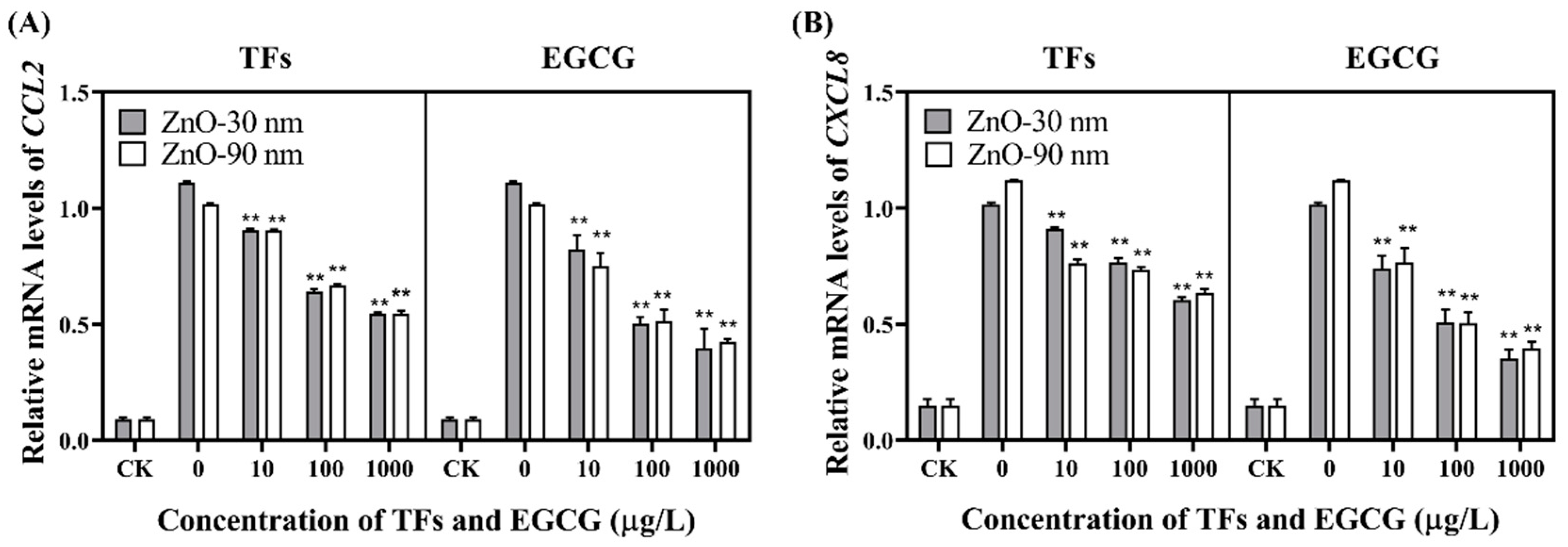
3.3. mRNA Expression Levels of IL-1β, IL-6, TNF-α and Inflammatory Cytokines
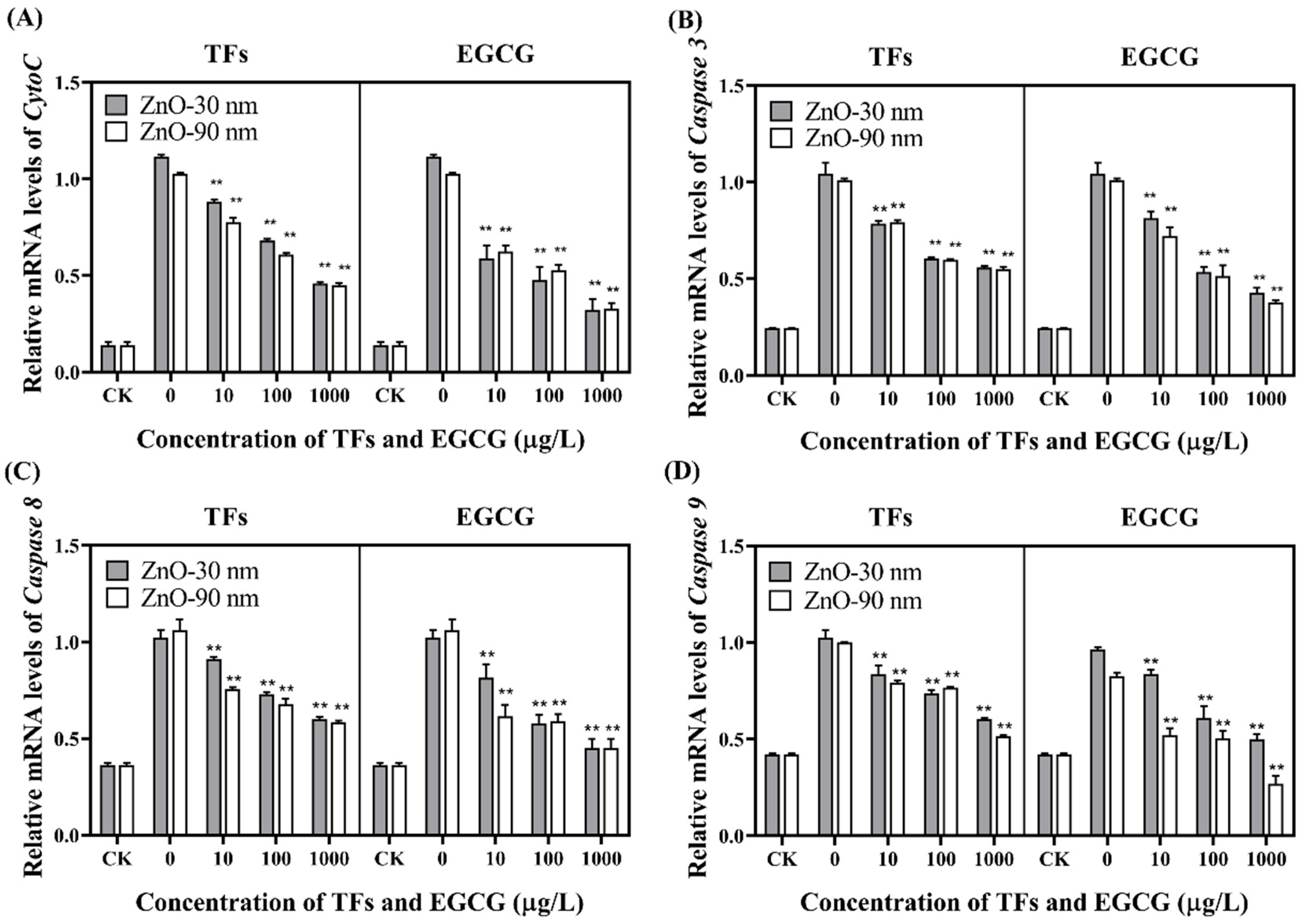
3.4. mRNA Level Expression of Cytochrome C (CytoC) and Caspase 3/8/9
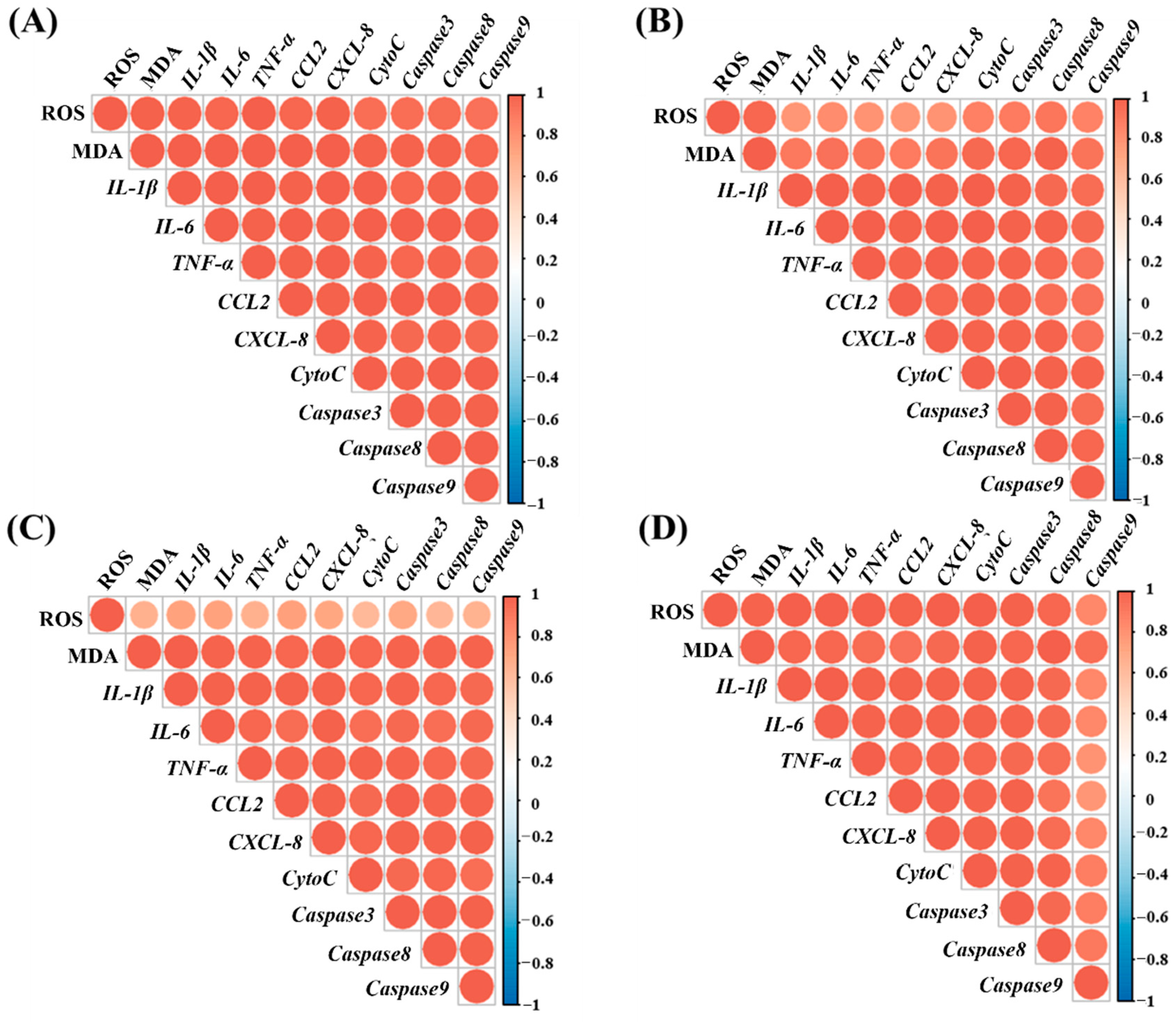
3.5. Correlation Analysis of Oxidative Stress, Inflammation, and Apoptosis
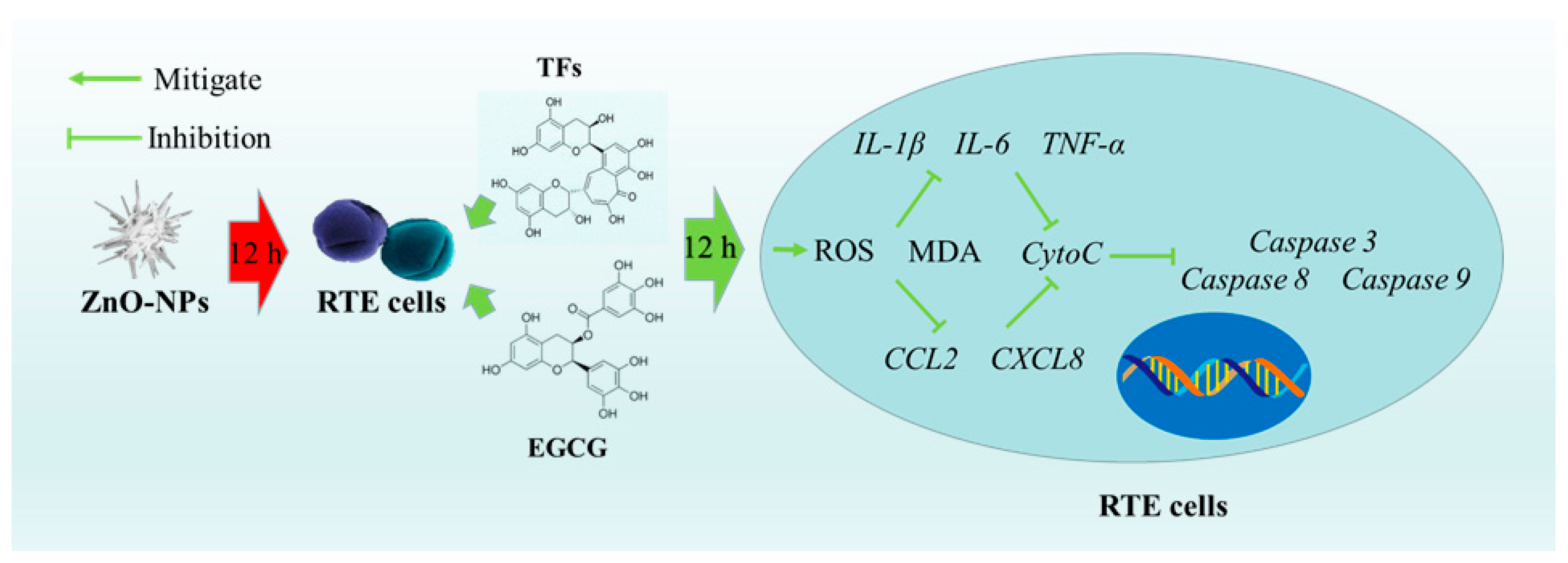
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Hu, D.; Shen, M.; Zhang, Y.; Li, H.; Zeng, G. Microplastics and nanoplastics: Would they affect global biodiversity change? Environ. Sci. Pollut. Res. Int. 2019, 26, 19997–20002. [Google Scholar] [CrossRef] [PubMed]
- Smii, H.; Khazri, A.; Ben Ali, M.; Mezni, A.; Hedfi, A.; Albogami, B.; Almalki, M.; Pacioglu, O.; Beyrem, H.; Boufahja, F.; et al. Titanium Dioxide Nanoparticles Are Toxic for the Freshwater Mussel Unio ravoisieri: Evidence from a Multimarker Approach. Diversity 2021, 13, 679. [Google Scholar] [CrossRef]
- Bringolf, R.B.; Raines, B.K.; Ratajczak, R.E.; Haskins, D.L. Major Ion Toxicity to Glochidia of Common and Imperiled Freshwater Mussel Species. Diversity 2022, 14, 95. [Google Scholar] [CrossRef]
- Yildirimer, L.; Thanh, N.T.; Loizidou, M.; Seifalian, A.M. Toxicology and clinical potential of nanoparticles. Nano Today 2011, 6, 585–607. [Google Scholar] [CrossRef]
- Zhang, Y.; Nayak, T.R.; Hong, H.; Cai, W. Biomedical Applications of Zinc Oxide Nanomaterials. Curr. Mol. Med. 2013, 13, 1633–1645. [Google Scholar] [CrossRef]
- Zuyderduyn, S.; Ninaber, D.K.; Hiemstra, P.S.; Rabe, K.F. The antimicrobial peptide LL-37 enhances IL-8 release by human airway smooth muscle cells. J. Allergy Clin. Immunol. 2006, 117, 1328–1335. [Google Scholar] [CrossRef]
- Goma, A.A.; Tohamy, H.G.; El-Kazaz, S.E.; Soliman, M.M.; Shukry, M.; Elgazzar, A.M.; Rashed, R.R. Insight Study on the Comparison between Zinc Oxide Nanoparticles and Its Bulk Impact on Reproductive Performance, Antioxidant Levels, Gene Expression, and Histopathology of Testes in Male Rats. Antioxidants 2020, 10, 41. [Google Scholar] [CrossRef]
- Rossner, P.; Vrbova, K.; Strapacova, S.; Rossnerova, A.; Ambroz, A.; Brzicova, T.; Libalova, H.; Javorkova, E.; Kulich, P.; Vecera, Z.; et al. Inhalation of ZnO Nanoparticles: Splice Junction Expression and Alternative Splicing in Mice. Toxicol. Sci. 2019, 168, 190–200. [Google Scholar] [CrossRef]
- Arathi, A.; Joseph, X.; Akhil, V.; Mohanan, P.V. L-Cysteine capped zinc oxide nanoparticles induced cellular response on adenocarcinomic human alveolar basal epithelial cells using a conventional and organ-on-a-chip approach. Colloids Surf. B Biointerfaces 2022, 211, 112300. [Google Scholar]
- Jung, A.; Kim, S.H.; Yang, J.Y.; Jeong, J.; Lee, J.K.; Oh, J.H.; Lee, J.H. Effect of Pulmonary Inflammation by Surface Functionalization of Zinc Oxide Nanoparticles. Toxics 2021, 9, 336. [Google Scholar] [CrossRef]
- Gong, F.; Xu, J.C.; Liu, B.; Yang, N.L.; Cheng, L.; Huang, P.; Wang, C.J.; Chen, Q.; Ni, C.F.; Liu, Z. Nanoscale CaH2 materials for synergistic hydrogen-immune cancer therapy. Chem 2022, 8, 268–286. [Google Scholar] [CrossRef]
- Lee, S.R.; Im, K.J.; Suh, S.I.; Jung, J.G. Protective effect of green tea polyphenol (-)-epigallocatechin gallate and other antioxidants on lipid peroxidation in gerbil brain homogenates. Phytother Res. 2003, 17, 206–209. [Google Scholar] [CrossRef] [PubMed]
- Manson, M.M.; Gescher, A.; Hudson, E.A.; Plummer, S.M.; Squires, M.S.; Prigent, S.A. Blocking and suppressing mechanisms of chemoprevention by dietary constituents. Toxicol. Lett. 2000, 112–113, 499–505. [Google Scholar] [CrossRef]
- Wang, Z.Y.; Huang, M.T.; Lou, Y.R.; Xie, J.G.; Reuhl, K.R.; Newmark, H.L.; Ho, C.T.; Yang, C.S.; Conney, A.H. Inhibitory effects of black tea, green tea, decaffeinated black tea, and decaffeinated green tea on ultraviolet B light-induced skin carcinogenesis in 7,12-dimethylbenz[a]anthracene-initiated SKH-1 mice. Cancer Res. 1994, 54, 3428–3435. [Google Scholar]
- Leung, L.K.; Su, Y.; Chen, R.; Zhang, Z.; Huang, Y.; Chen, Z.Y. Theaflavins in Black Tea and Catechins in Green Tea Are Equally Effective Antioxidants. J. Nutr. 2001, 131, 2248–2251. [Google Scholar] [CrossRef]
- Henning, S.M.; Choo, J.J.; Heber, D. Nongallated Compared with Gallated Flavan-3-ols in Green and Black Tea Are More Bioavailable. J. Nutr. 2008, 138, 1529S–1534S. [Google Scholar] [CrossRef]
- Zhou, R.; Li, X.; Li, L.; Zhang, H. Theaflavins alleviate sevoflurane-induced neurocytotoxicity via Nrf2 signaling pathway. Int. J. Neurosci. 2020, 130, 1–8. [Google Scholar] [CrossRef]
- Wang, Z.; Luo, H.; Xia, H. Theaflavins attenuate ethanol-induced oxidative stress and cell apoptosis in gastric mucosa epithelial cells via downregulation of the mitogenactivated protein kinase pathway. Mol. Med. Rep. 2018, 18, 3791–3799. [Google Scholar] [CrossRef]
- Hatasa, Y.; Chikazawa, M.; Furuhashi, M.; Nakashima, F.; Shibata, T.; Kondo, T.; Akagawa, M.; Hamagami, H.; Tanaka, H.; Tachibana, H.; et al. Oxidative Deamination of Serum Albumins by (-)-Epigallocatechin-3-O-Gallate: A Potential Mechanism for the Formation of Innate Antigens by Antioxidants. PLoS ONE 2016, 11, e0153002. [Google Scholar] [CrossRef]
- Ozgur, E.; Guler, G.; Seyhan, N. Mobile phone radiation-induced free radical damage in the liver is inhibited by the antioxidants n-acetyl cysteine and epigallocatechin-gallate. Int. J. Radiat. Biol. 2010, 86, 935–945. [Google Scholar] [CrossRef]
- Nagle, D.G.; Ferreira, D.; Zhou, Y.D. Epigallocatechin-3-gallate (EGCG): Chemical and biomedical perspectives. Phytochemistry 2006, 67, 1849–1855. [Google Scholar] [CrossRef] [PubMed]
- Singh, B.N.; Shankar, S.; Srivastava, R.K. Green tea catechin, epigallocatechin-3-gallate (EGCG): Mechanisms, perspectives and clinical applications. Biochem. Pharmacol. 2011, 82, 1807–1821. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sakagami, H.; Arakawa, H.; Maeda, M.; Satoh, K.; Kadofuku, T.; Fukuchi, K.; Gomi, K. Production of hydrogen peroxide and me-thionine sulfoxide by epigallocatechin gallate and antioxidants. Anticancer Res. 2001, 21, 2633–2641. [Google Scholar]
- Na, H.K.; Surh, Y.J. Modulation of Nrf2-mediated antioxidant and detoxifying enzyme induction by the green tea polyphenol EGCG. Food Chem. Toxicol. 2008, 46, 1271–1278. [Google Scholar] [CrossRef] [PubMed]
- Yi, J.; Chen, C.; Liu, X.; Kang, Q.; Hao, L.; Huang, J.; Lu, J. Radioprotection of EGCG based on immunoregulatory effect and antioxidant activity against (60)Cogamma radiation-induced injury in mice. Food Chem. Toxicol. 2020, 135, 111051. [Google Scholar] [CrossRef]
- Liang, Y.; Ip, M.S.M.; Mak, J.C.W. (-)-Epigallocatechin-3-gallate suppresses cigarette smoke-induced inflammation in human cardiomyocytes via ROS-mediated MAPK and NF-kappaB pathways. Phytomedicine 2019, 58, 152768. [Google Scholar] [CrossRef]
- Shi, J.; Zhang, M.; Zhang, L.; Deng, H. Epigallocatechin-3-gallate attenuates microcystin-LR-induced apoptosis in human umbilical vein endothelial cells through activation of the NRF2/HO-1 pathway. Environ. Pollut. 2018, 239, 466–472. [Google Scholar] [CrossRef]
- Cao, W.; Gu, M.; Wang, S.; Huang, C.; Xie, Y.; Cao, Y. Effects of epigallocatechin gallate on the stability, dissolution and toxicology of ZnO nanoparticles. Food Chem. 2022, 371, 131383. [Google Scholar] [CrossRef]
- Chen, X.; Wang, D.; Guo, X.; Li, X.; Ye, W.; Qi, Y.; Gu, W. Curcumin-Loaded mPEG-PLGA Nanoparticles Attenuates the Apoptosis and Corticosteroid Resistance Induced by Cigarette Smoke Extract. Front. Pharmacol. 2022, 13, 824652. [Google Scholar] [CrossRef]
- Ahamed, M.; Akhtar, M.J.; Alaizeri, Z.M.; Alhadlaq, H.A. TiO2 nanoparticles potentiated the cytotoxicity, oxidative stress and apoptosis response of cadmium in two different human cells. Environ. Sci. Pollut. Res. Int. 2020, 27, 10425–10435. [Google Scholar] [CrossRef]
- Gao, Y.L.; Yoshida, A.; Liu, J.Y.; Shimizu, T.; Shirota, K.; Shiina, Y.; Osatomi, K. Molecular cloning and expression dynamics of UNC-45B upon heat shock in the muscle of yellowtail. Aquaculture 2021, 541, 736827. [Google Scholar] [CrossRef]
- Sedanza, M.G.; Alshaweesh, J.; Gao, Y.-L.; Yoshida, A.; Kim, H.-J.; Yamaguchi, K.; Satuito, C.G. Transcriptome Dynamics of an Oyster Larval Response to a Conspecific Cue-Mediated Settlement Induction in the Pacific Oyster Crassostrea gigas. Diversity 2022, 14, 559. [Google Scholar] [CrossRef]
- Herrmann, F.; Romero, M.R.; Blazquez, A.G.; Kaufmann, D.; Ashour, M.L.; Kahl, S.; Marín, J.J.G.; Efferth, T.; Wink, M.; Mohamed, L.A. Diversity of Pharmacological Properties in Chinese and European Medicinal Plants: Cytotoxicity, Antiviral and Antitrypanosomal Screening of 82 Herbal Drugs. Diversity 2011, 3, 547–580. [Google Scholar] [CrossRef] [Green Version]
- Wang, B.; Zhang, J.; Chen, C.; Xu, G.; Qin, X.; Hong, Y.; Bose, D.D.; Qiu, F.; Zou, Z. The size of zinc oxide nanoparticles controls its toxicity through impairing autophagic flux in A549 lung epithelial cells. Toxicol. Lett. 2018, 285, 51–59. [Google Scholar] [CrossRef] [PubMed]
- Shalini, D.; Senthilkumar, S.; Rajaguru, P. Effect of size and shape on toxicity of zinc oxide (ZnO) nanomaterials in human peripheral blood lymphocytes. Toxicol. Mech. Methods 2018, 28, 87–94. [Google Scholar] [CrossRef]
- Elsaesser, A.; Howard, C.V. Toxicology of nanoparticles. Adv. Drug Deliv. Rev. 2012, 64, 129–137. [Google Scholar] [CrossRef]
- Durnev, A.D. Toxicology of nanoparticles. Bull. Exp. Biol. Med. 2008, 145, 72–74. [Google Scholar] [CrossRef]
- Filippi, C.; Pryde, A.; Cowan, P.; Lee, T.; Hayes, P.; Donaldson, K.; Plevris, J.; Stone, V. Toxicology of ZnO and TiO2nanoparticles on hepatocytes: Impact on metabolism and bioenergetics. Nanotoxicology 2015, 9, 126–134. [Google Scholar] [CrossRef]
- Oberdorster, G.; Oberdorster, E.; Oberdorster, J. Nanotoxicology: An emerging discipline evolving from studies of ultrafine particles. Environ. Health Perspect. 2005, 113, 823–839. [Google Scholar] [CrossRef]
- Xiong, D.W.; Fang, T.; Yu, L.P.; Sima, X.F.; Zhu, W.T. Effects of nano-scale TiO2, ZnO and their bulk counterparts on zebrafish: Acute toxicity, oxidative stress and oxidative damage. Sci. Total Environ. 2011, 409, 1444–1452. [Google Scholar] [CrossRef]
- Kumar, A.; Pandey, A.K.; Singh, S.S.; Shanker, R.; Dhawan, A. Engineered ZnO and TiO2 nanoparticles induce oxidative stress and DNA damage leading to reduced viability of Escherichia coli. Free Radic. Biol. Med. 2011, 51, 1872–1881. [Google Scholar] [CrossRef] [PubMed]
- Yang, Z.; Jie, G.; Dong, F.; Xu, Y.; Watanabe, N.; Tu, Y. Radical-scavenging abilities and antioxidant properties of theaflavins and their gallate esters in H2O2-mediated oxidative damage system in the HPF-1 cells. Toxicol. In Vitro 2008, 22, 1250–1256. [Google Scholar] [CrossRef] [PubMed]
- Ni, J.; Guo, X.; Wang, H.; Zhou, T.; Wang, X. Differences in the Effects of EGCG on Chromosomal Stability and Cell Growth between Normal and Colon Cancer Cells. Molecules 2018, 23, 788. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lorenz, M. Cellular targets for the beneficial actions of tea polyphenols. Am. J. Clin. Nutr. 2013, 98, 1642S–1650S. [Google Scholar] [CrossRef]
- Shen, Y.J.; Li, W.G.; Zhang, H.; Li, B.; Yuan, H.X.; Zhang, J.J. Effects of (-)-epigallocatechin-3-gallate on interleukin-1beta expression in mouse wound healing. Fen Zi Xi Bao Sheng Wu Xue Bao 2009, 42, 179–185. [Google Scholar]
- Yamagata, K.; Xie, Y.; Suzuki, S.; Tagami, M. Epigallocatechin-3-gallate inhibits VCAM-1 expression and apoptosis induction associated with LC3 expressions in TNFalpha-stimulated human endothelial cells. Phytomedicine 2015, 22, 431–437. [Google Scholar] [CrossRef]
- Khazali, A.S.; Clark, A.M.; Wells, A. Inflammatory cytokine IL-8/CXCL8 promotes tumour escape from hepatocyte-induced dormancy. Br. J. Cancer 2018, 118, 566–576. [Google Scholar] [CrossRef]
- Zhang, W.; Shi, H.Y.; Zhang, M. Maspin overexpression modulates tumor cell apoptosis through the regulation of Bcl-2 family proteins. BMC Cancer 2005, 5, 50. [Google Scholar] [CrossRef] [Green Version]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene | Genbank Accession | Primer Sequences (5′ to 3′) |
|---|---|---|
| CytoC | NM_031543.1 | 5′-GACTTTGGCCGACCTGTTCTTT-3′ |
| 5′-CATGAGGATCAGGAGCCCATATCT-3′ | ||
| CCL2 | M57441 | 5′-CACCTGCTGCTACTCATTCACTG-3′ |
| 5′-CTTCTTTGGGACACCTGCTGCT-3′ | ||
| CXCL8 | NM_030845 | 5′-CACCCAAACCGAAGTCATAGCCA-3′ |
| 5′-CTTGGGGACACCCTTTAGCATCT-3′ | ||
| IL-1β | NM_031512 | 5′-CCTAGGAAACAGCAATGGTCGGGAC-3′ |
| 5′-CCTAGGAAACAGCAATGGTCGGGAC-3′ | ||
| IL-6 | NM_012589.2 | 5′-CTTCACAGAGGATACCACCCACA-3′ |
| 5′-CAGTGCATCATCGCTGTTCATACA-3′ | ||
| TNF-α | NM_012675 | 5′-GACCCCTTTATCGTCTACTCCTC-3′ |
| 5′-GCCACTACTTCAGCGTCTCGT-3′ | ||
| Caspase3 | NM_012922.2 | 5′-AGAGTTGGAGCACTGTAGCACACA-3′ |
| 5′-TCATGTCCACCACTGAAGGATGGT- 3 ‘ | ||
| Caspase 8 | NM_022277.1 | 5′-GCTGGGGATGGCTACTGTGAAA-3′ |
| 5′-GGCTCTGGCAAAGTGACTGGATA-3′ | ||
| Caspase 9 | NM_031632.1 | 5′-GGTGAAGAACGACCTGACTGCTAA-3′ |
| 5′-GAGAGGATGACCACCACGAAG-3′ | ||
| Rat GAPDH | NM_017008.4 | 5′-GAAGGTCGGTGTGAACGGATTTG-3′ |
| 5′-CATGTAGACCATGTAGTTGAGGTCA-3′ |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Shan, X.; Chen, F.; Lin, H.; Zhang, H.; Zhong, Y.; Liu, Z.; Li, Y. Protective Effects of Theaflavins and Epigallocatechin Gallate against ZnO-NP-Induced Cell Apoptosis In Vitro. Diversity 2022, 14, 756. https://doi.org/10.3390/d14090756
Shan X, Chen F, Lin H, Zhang H, Zhong Y, Liu Z, Li Y. Protective Effects of Theaflavins and Epigallocatechin Gallate against ZnO-NP-Induced Cell Apoptosis In Vitro. Diversity. 2022; 14(9):756. https://doi.org/10.3390/d14090756
Chicago/Turabian StyleShan, Xiaodong, Feifei Chen, Huikang Lin, Hangjun Zhang, Yuchi Zhong, Zhiquan Liu, and Yan Li. 2022. "Protective Effects of Theaflavins and Epigallocatechin Gallate against ZnO-NP-Induced Cell Apoptosis In Vitro" Diversity 14, no. 9: 756. https://doi.org/10.3390/d14090756