
Genetic Diversity and Relatedness of Group-Living Small Mammals on the Mongolian Plateau

Abstract
:1. Introduction
2. Methods
2.1. Study Areas
2.2. Live-Trapping of Daurian Pikas and Mongolian Gerbils
2.3. Microsatellite Analysis
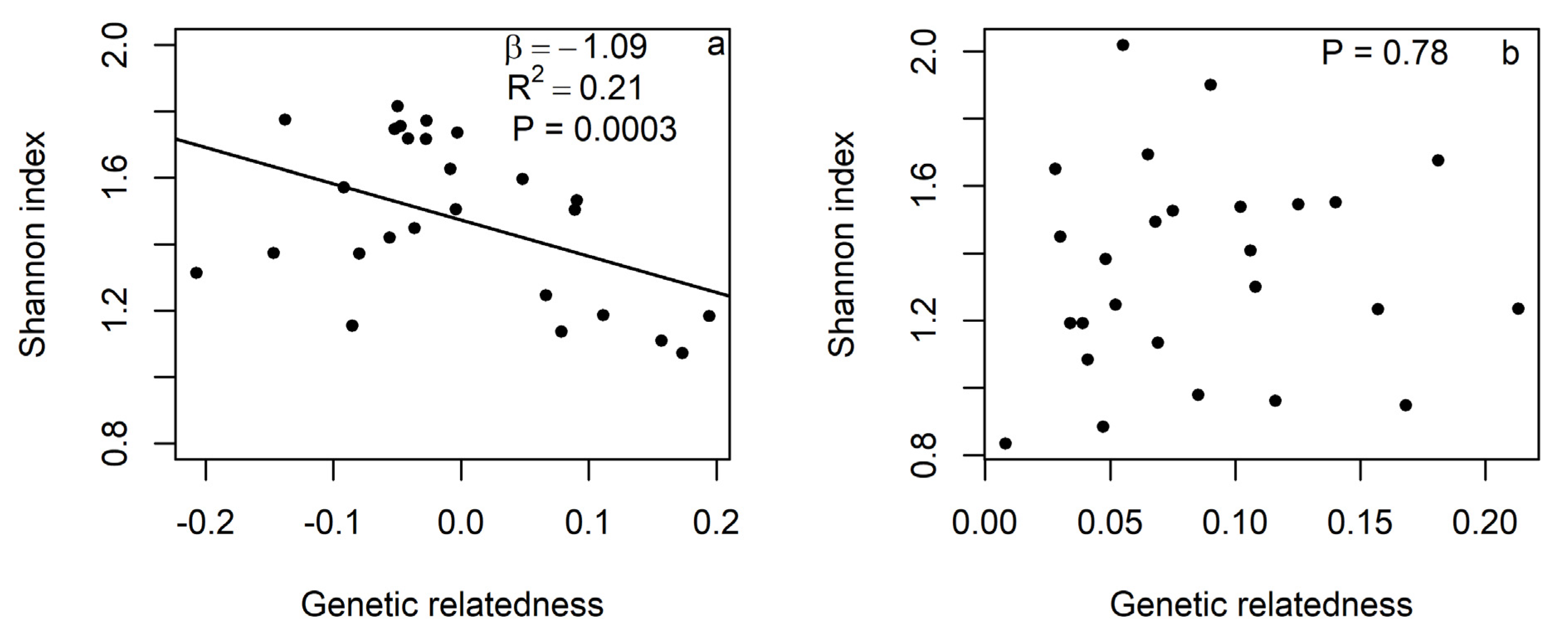
2.4. Statistical Analysis
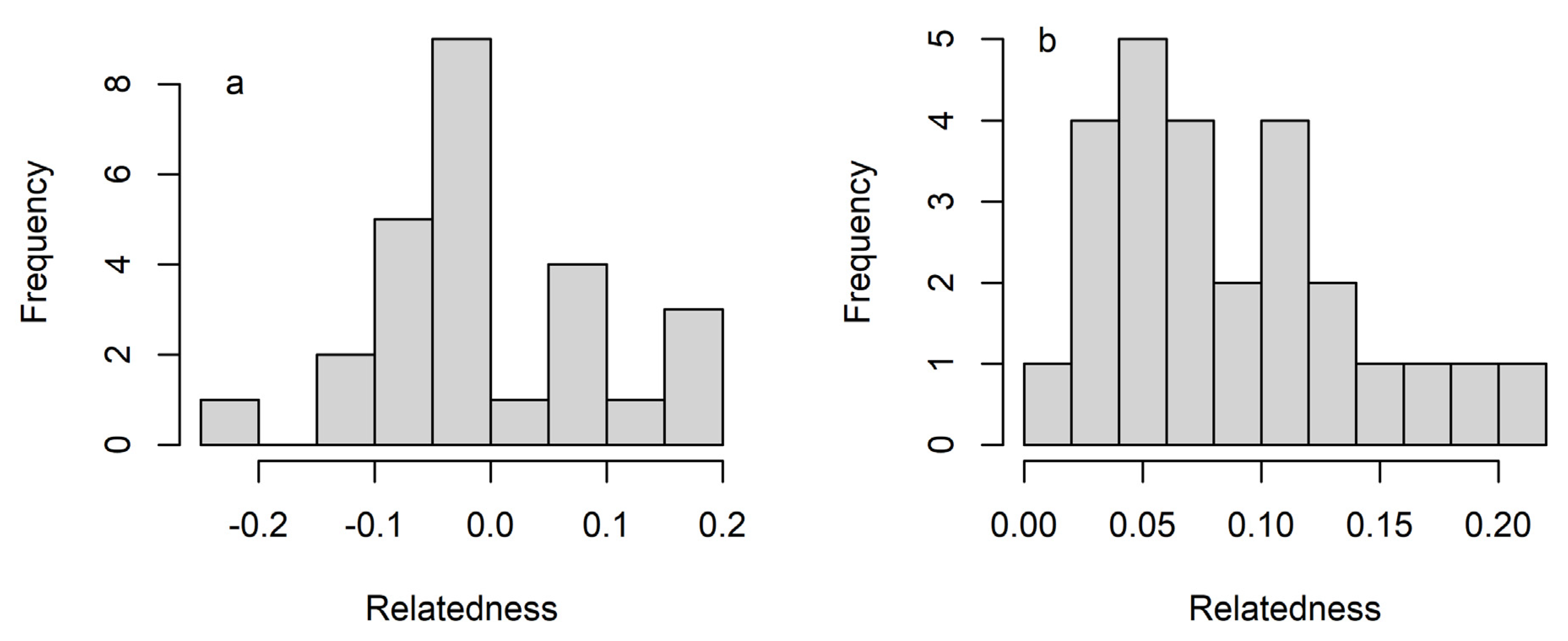
3. Results
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Hughes, A.R.; Inouye, B.D.; Johnson, M.T.; Underwood, N.; Vellend, M. Ecological consequences of genetic diversity. Ecol. Lett. 2008, 11, 609–623. [Google Scholar] [CrossRef] [PubMed]
- Freeland, J.R. Molecular Ecology; Wiley: West Sussex, UK, 2005. [Google Scholar]
- Reed, D.H.; Frankham, R. Correlation between fitness and genetic diversity. Conserv. Biol. 2003, 17, 230–237. [Google Scholar] [CrossRef]
- Frankham, R.; Ballou, S.E.J.D.; Briscoe, D.A.; Ballou, J.D. Introduction to Conservation Genetics; Cambridge University Press: Cambridge, UK, 2002. [Google Scholar]
- Ellegren, H.; Galtier, N. Determinants of genetic diversity. Nat. Rev. Genet. 2016, 17, 422–433. [Google Scholar] [CrossRef]
- Wang, Y.; Liu, W.; Wang, G.; Zhong, W.; Wan, X. Genetic consequences of group living in Mongolian gerbils. J. Hered. 2011, 102, 554–561. [Google Scholar]
- Hamilton, W.D. Genetical evolution of social behaviour I. J. Theor. Biol. 1964, 7, 17–52. [Google Scholar] [CrossRef]
- Agren, G.; Zhou, Q.; Zhong, W. Territoriality, cooperation and resource priority: Hoarding in the Mongolian gerbil, Meriones unguiculatus. Anim. Behav. 1989, 37, 28–32. [Google Scholar] [CrossRef]
- Liu, W.; Wang, G.; Wan, X.; Zhong, W. Effects of supplemental food on the social organization of Mongolian gerbils during the breeding season. J. Zool. 2009, 278, 249–257. [Google Scholar] [CrossRef]
- Armansin, N.C.; Stow, A.J.; Cantor, M.; Leu, S.T.; Klarevas-Irby, J.A.; Chariton, A.A.; Farine, D.R. Social barriers in ecological landscapes: The social resistance hypothesis. Trends Ecol. Evol. 2020, 35, 137–148. [Google Scholar] [CrossRef]
- Wang, G.; Zhou, Q.; Zhong, W.; Wang, Z. Spatial overlap between sympatric Microtus brandti (Rodentia, Microtinae) and Ochotona daurica (Lagomorpha, Ochotonidae) in the steppes of inner Mongolia. Mammalia 2003, 67, 349–354. [Google Scholar] [CrossRef]
- Wang, G.; Zhong, W. Mongolian gerbils and Daurian pikas responded differently to changes in precipitation in the Inner Mongolian grasslands. J. Arid Environ. 2006, 66, 648–656. [Google Scholar] [CrossRef]
- Smith, A.T. Patterns of pikas (Genus Ochotona) life history variation. In Evolution of Life Histories of Mammals; Boyce, M.S., Ed.; Yale University Press: New Haven, CT, USA, 1988; pp. 233–256. [Google Scholar]
- Zhong, W.; Wang, G.; Zhou, Q.; Wan, X. Effects of winter food availability on the abundance of Daurian pikas (Ochotona dauurica) in Inner Mongolian grasslands. J. Arid Environ. 2008, 72, 1383–1387. [Google Scholar] [CrossRef]
- Chen, L.J.; Zhang, W.J.; Zhang, X.Q.; Jia, J.J.; Liu, W.; Wan, X.R. A preliminary study of reproductiive ecology of Daurian pikas in the typica steppe. Chin. J. Zool. 2014, 49, 649–656. [Google Scholar]
- Formozov, A.N. Adaptive modifications of behavior in mammals of the Eurasian steppes. J. Mammal. 1966, 47, 208–223. [Google Scholar] [CrossRef]
- Agren, G.; Zhou, Q.; Zhong, W. Ecology and Social-Behavior of Mongolian Gerbils, Meriones unguiculatus, at Xilinhot, Inner-Mongolia, China. Anim. Behav. 1989, 37, 11–27. [Google Scholar] [CrossRef]
- Wang, Y.; Liu, W.; Wang, G.; Wan, X.; Zhong, W. Home-range sizes of social groups of Mongolian gerbils Meriones unguiculatus. J. Arid Environ. 2011, 75, 132–137. [Google Scholar] [CrossRef]
- Scheibler, E.; Weinandy, R.; Gattermann, R. Social categories in families of Mongolian gerbils. Physiol. Behav. 2004, 81, 455–464. [Google Scholar] [CrossRef]
- Clark, M.M.; Galef, B.G. Socially induced infertility: Familial effects on reproductive development of female Mongolian gerbils. Anim. Behav. 2001, 62, 897–903. [Google Scholar] [CrossRef]
- Chen, L.; Wang, G.; Wan, X.; Liu, W. Complex and nonlinear effects of weather and density on the demography of small herbivorous mammals. Basic Appl. Ecol. 2015, 16, 172–179. [Google Scholar] [CrossRef]
- Liu, W.; Wang, G.; Wang, Y.; Zhong, W.; Wan, X. Population ecology of wild Mongolian gerbils Meriones unguiculatus. J. Mammal. 2009, 90, 832–840. [Google Scholar] [CrossRef]
- Sambrook, J.; Russell, D.W. Molecular Cloning: A Laboratory Manual, 3rd ed.; Cold Spring Harbor Laboratory Press: Cold Spring, NY, USA, 2001. [Google Scholar]
- Peacock, M.M.; Kirchoff, V.S.; Merideth, S.J. Identification and characterization of nine polymorphic microsatellite loci in the North American pika, Ochotona princeps. Mol. Ecol. Notes 2002, 2, 360–362. [Google Scholar] [CrossRef]
- Zgurski, J.M.; Davis, C.S.; Hik, D.S. Isolation and characterization of microsatellite loci for the collared pika (Ochotona collaris) and their cross-amplification in five other Ochotona species. Mol. Ecol. Resour. 2009, 9, 867–871. [Google Scholar] [CrossRef] [PubMed]
- Li, K.X.; Geng, J.N.; Yang, J.; Zhang, Y.M.; Hu, S.N. Isolation and characterization of 13 microsatellite loci in the plateau pika (Ochotona curzoniae). Conserv. Genet. 2009, 10, 785–787. [Google Scholar] [CrossRef]
- Van Oosterhout, C.; Hutchinson, W.F.; Wills, D.P.M.; Shipley, P. MICRO-CHECKER: Software for identifying and correcting genotyping errors in microsatellite data. Mol. Ecol. Notes 2004, 4, 535–538. [Google Scholar] [CrossRef]
- Raymond, M.; Rousset, F. GENEPOP (Version-1.2): Population genetics software for exact tests and ecumenicism. J. Hered. 1995, 86, 248–249. [Google Scholar] [CrossRef]
- Neumann, K.; Maak, S.; Stuermer, I.W.; von Lengerken, G.; Gattermann, R. Low microsatellite variation in laboratory gerbils. J. Hered. 2001, 92, 71–74. [Google Scholar] [CrossRef]
- Queller, D.C.; Goodnight, K.F. Estimating relatedness using genetic markers. Evolution 1989, 43, 258–275. [Google Scholar] [CrossRef]
- Peakall, R.; Smouse, P.E. GENALEX 6: Genetic analysis in Excel. Population genetic software for teaching and research. Mol. Ecol. Notes 2006, 6, 288–295. [Google Scholar] [CrossRef]
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2022; Available online: https://www.R-project.org/ (accessed on 31 July 2022).
- Wang, J. Marker-based estimates of relatedness and inbreeding coefficients: An assessment of current methods. J. Evol. Biol. 2014, 27, 518–530. [Google Scholar] [CrossRef]
- Taylor, H.R. The use and abuse of genetic marker-based estimates of relatedness and inbreeding. Ecol. Evol. 2015, 5, 3140–3150. [Google Scholar] [CrossRef]
- Robson, K.M.; Lamb, C.T.; Russello, M.A. Low genetic diversity, restricted dispersal, and elevation-specific patterns of population decline in American pikas in an atypical environment. J. Mammal. 2016, 97, 464–472. [Google Scholar] [CrossRef]
- Zgurski, J.M.; Hik, D.S. Gene flow and the restoration of genetic diversity in a fluctuating collared pika (Ochotona collaris) population. Conserv. Genet. 2014, 15, 37–48. [Google Scholar] [CrossRef]
- Zhou, L.; Yang, S.; Yu, Z.; Yin, B.; Huai, H.; Li, S.; Wei, W. Genetic diversity and differentiation in four natural populations of plateau pika (Ochotona curzoniae). Acta Theriol. Sin. 2007, 27, 221–228. [Google Scholar]
- Volodin, I.A.; Volodina, E.V.; Frey, R.; Karaseva, K.D.; Kirilyuk, V.E. Daurian pika (Ochotona dauurica) alarm calls: Individual acoustic variation in a lagomorph with audible through ultrasonic vocalizations. J. Mammal. 2021, 102, 947–959. [Google Scholar] [CrossRef]
- Quirici, V.; Faugeron, S.; Hayes, L.D.; Ebensperger, L.A. Absence of kin structure in a population of the group-living rodent Octodon degus. Behav. Ecol. 2011, 22, 248–254. [Google Scholar] [CrossRef]
- Lacey, E.A.; Wieczorek, J.R. Kinship in colonial tuco-tucos: Evidence from group composition and population structure. Behav. Ecol. 2004, 15, 988–996. [Google Scholar] [CrossRef]
- Zhu, X.D.; Li, X.J.; Du, X.Y.; Wang, Y.; Lu, J.; Chen, Z.W. Comparative analysis of population genetic structure for wild and domesticated Mongolian gerbils. Chin. J. Comp. Med. 2008, 18, 39–43. [Google Scholar]
- Liu, W.; Wang, G.; Wan, X.; Zhong, W. Winter food availability limits winter survival of Mongolian gerbils (Meriones unguiculatus). Acta Theriol. 2011, 56, 219–227. [Google Scholar] [CrossRef]
- Wang, G.; Liu, W.; Wang, Y.; Wan, X.; Zhong, W. Restricted dispersal determines fine-scale spatial genetic structure of Mongolian gerbils. Curr. Zool. 2017, 63, 687–691. [Google Scholar] [CrossRef]
- Wimmer, B.; Tautz, D.; Kappeler, P.M. The genetic population structure of the gray mouse lemur (Microcebus murinus), a basal primate from Madagascar. Behav. Ecol. Sociobiol. 2002, 52, 166–175. [Google Scholar] [CrossRef]
- Sumbh, O.; Hof, A.R. Can pikas hold the umbrella? Understanding the current and future umbrella potential of keystone species Pika (Ochotona spp.). Glob. Ecol. Conserv. 2022, 38, e02247. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
| occ2 | ocp1 | ocp2 | ocp3 | ocp5 | p7F | p47F | p124R | p149F | p156F | p172R | |
|---|---|---|---|---|---|---|---|---|---|---|---|
| Number of alleles | 10 | 6 | 7 | 19 | 17 | 26 | 14 | 19 | 31 | 8 | 25 |
| Number of effective alleles | 4.01 | 2.23 | 3.58 | 9.67 | 8.52 | 14.47 | 10.87 | 10.10 | 13.74 | 2.78 | 10.69 |
| Shannon diversity index | 1.66 | 1.00 | 1.50 | 2.49 | 2.35 | 2.86 | 2.49 | 2.48 | 2.91 | 1.43 | 2.68 |
| Observed heterozygosity (Ho) | 0.68 | 0.58 | 0.49 | 0.68 | 0.80 | 0.50 | 0.82 | 0.79 | 0.68 | 0.42 | 0.84 |
| Expected heterozygosity (He) | 0.75 | 0.55 | 0.72 | 0.90 | 0.88 | 0.93 | 0.91 | 0.90 | 0.93 | 0.64 | 0.91 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wang, G.; Wan, X.; Liu, W.; Shan, X. Genetic Diversity and Relatedness of Group-Living Small Mammals on the Mongolian Plateau. Diversity 2022, 14, 732. https://doi.org/10.3390/d14090732
Wang G, Wan X, Liu W, Shan X. Genetic Diversity and Relatedness of Group-Living Small Mammals on the Mongolian Plateau. Diversity. 2022; 14(9):732. https://doi.org/10.3390/d14090732
Chicago/Turabian StyleWang, Guiming, Xinrong Wan, Wei Liu, and Xueyan Shan. 2022. "Genetic Diversity and Relatedness of Group-Living Small Mammals on the Mongolian Plateau" Diversity 14, no. 9: 732. https://doi.org/10.3390/d14090732