
Phylogeography of Mesophotic Coral Ecosystems: Squirrelfish and Soldierfish (Holocentriformes: Holocentridae)

 , , and
, , and
Abstract
:1. Introduction
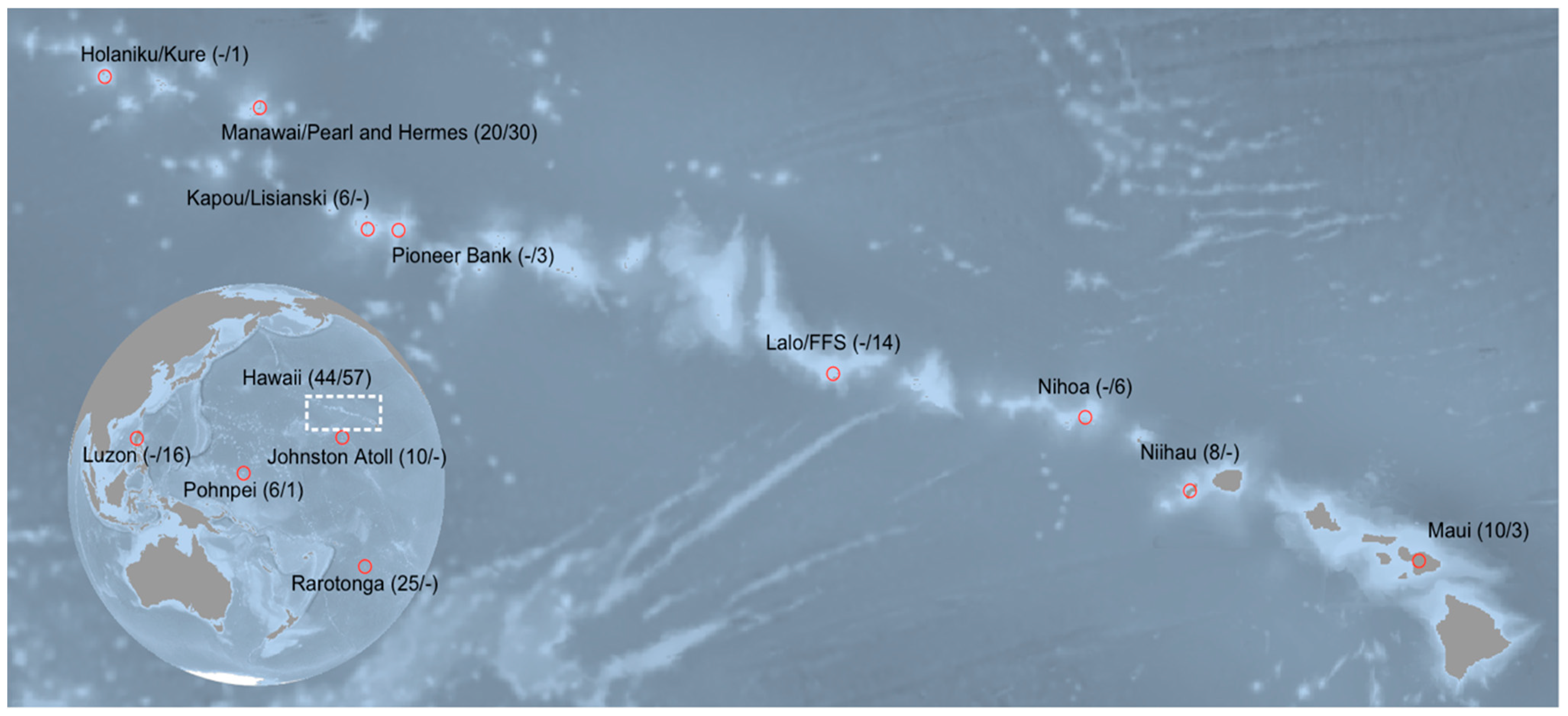
2. Materials and Methods
2.1. Specimen Collection and DNA Extraction
2.2. PCR and Sequencing
2.3. Data Analysis
3. Results
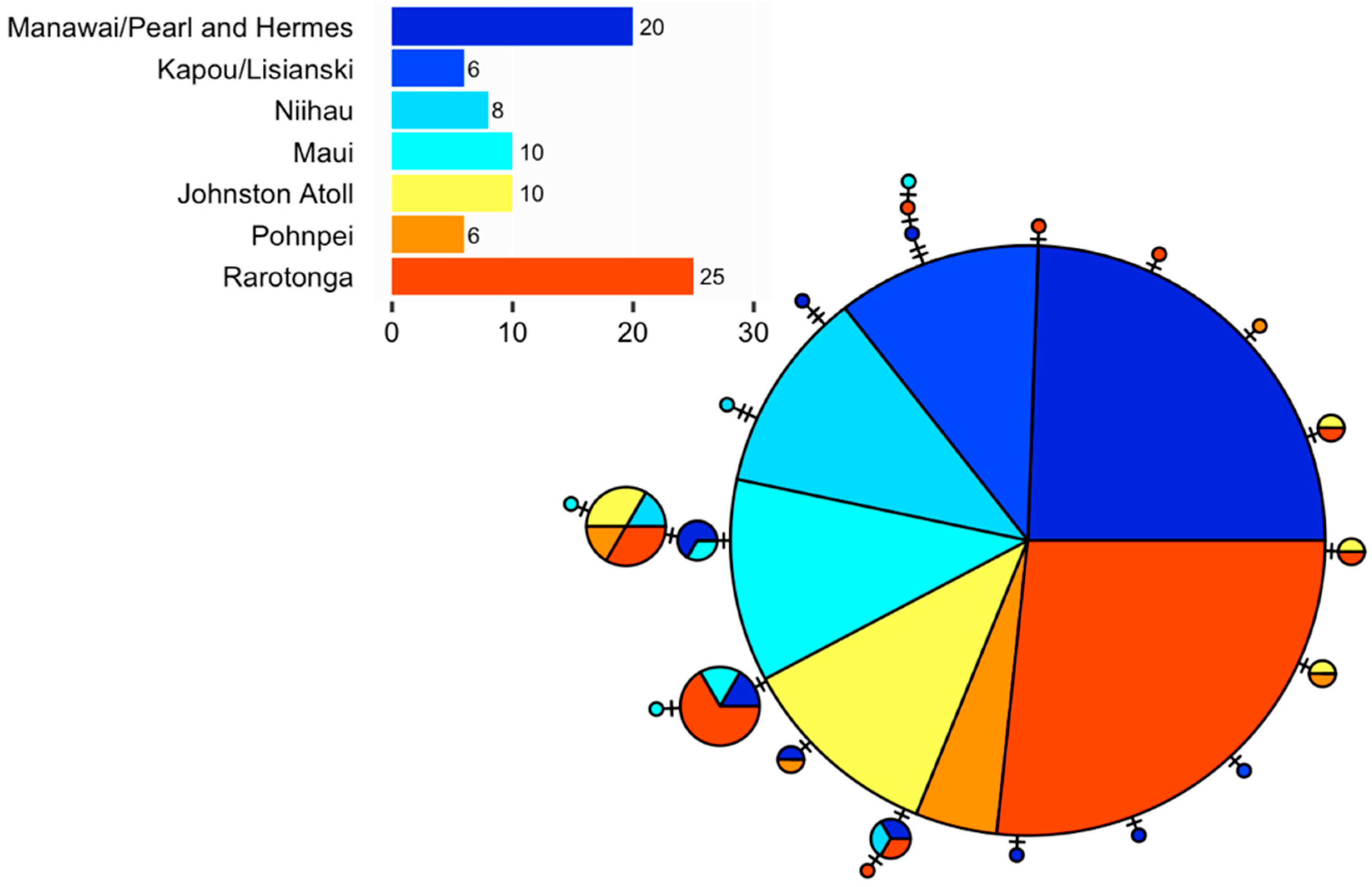
3.1. Myripristis Chryseres
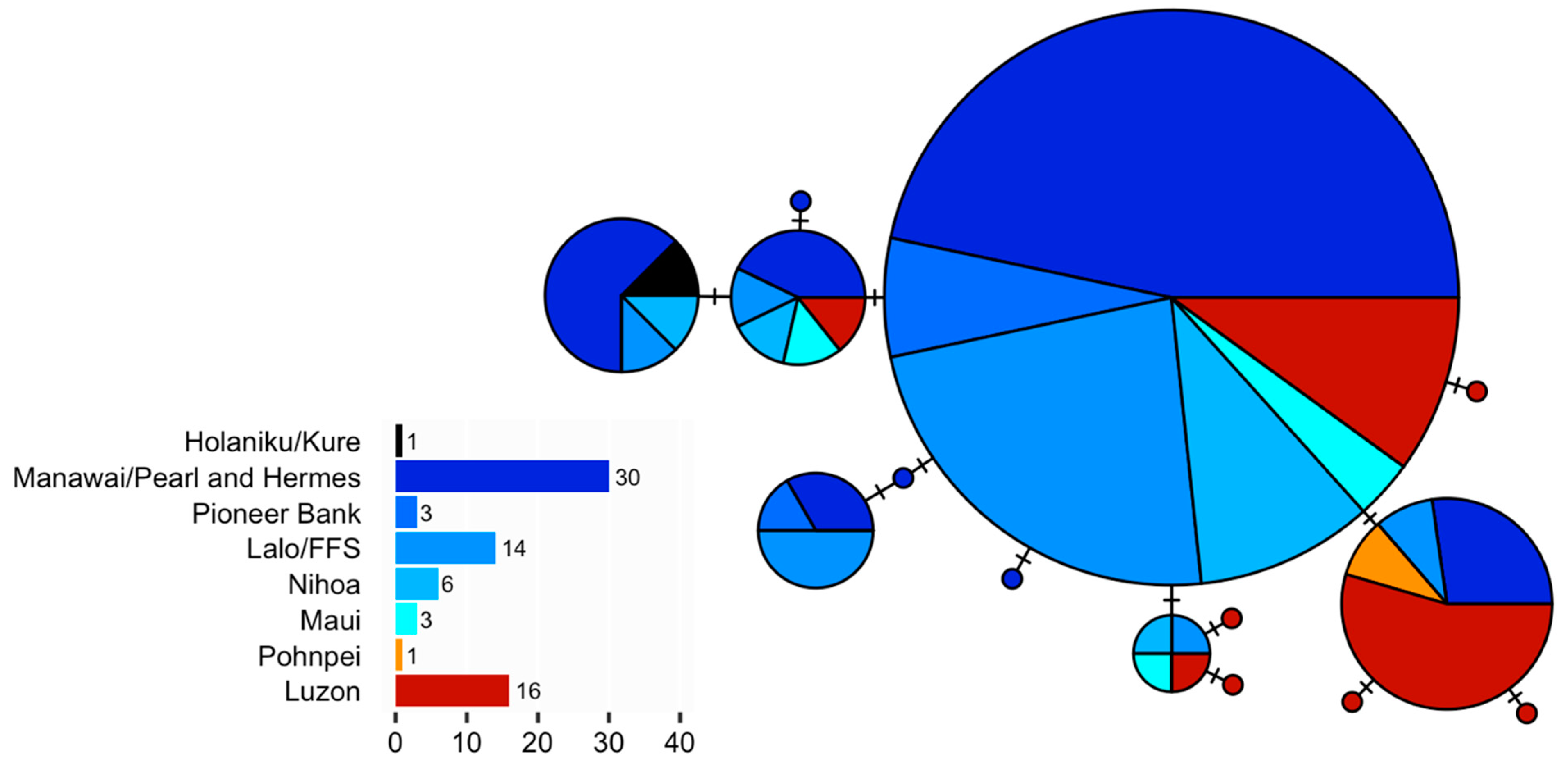
3.2. Neoniphon Aurolineatus
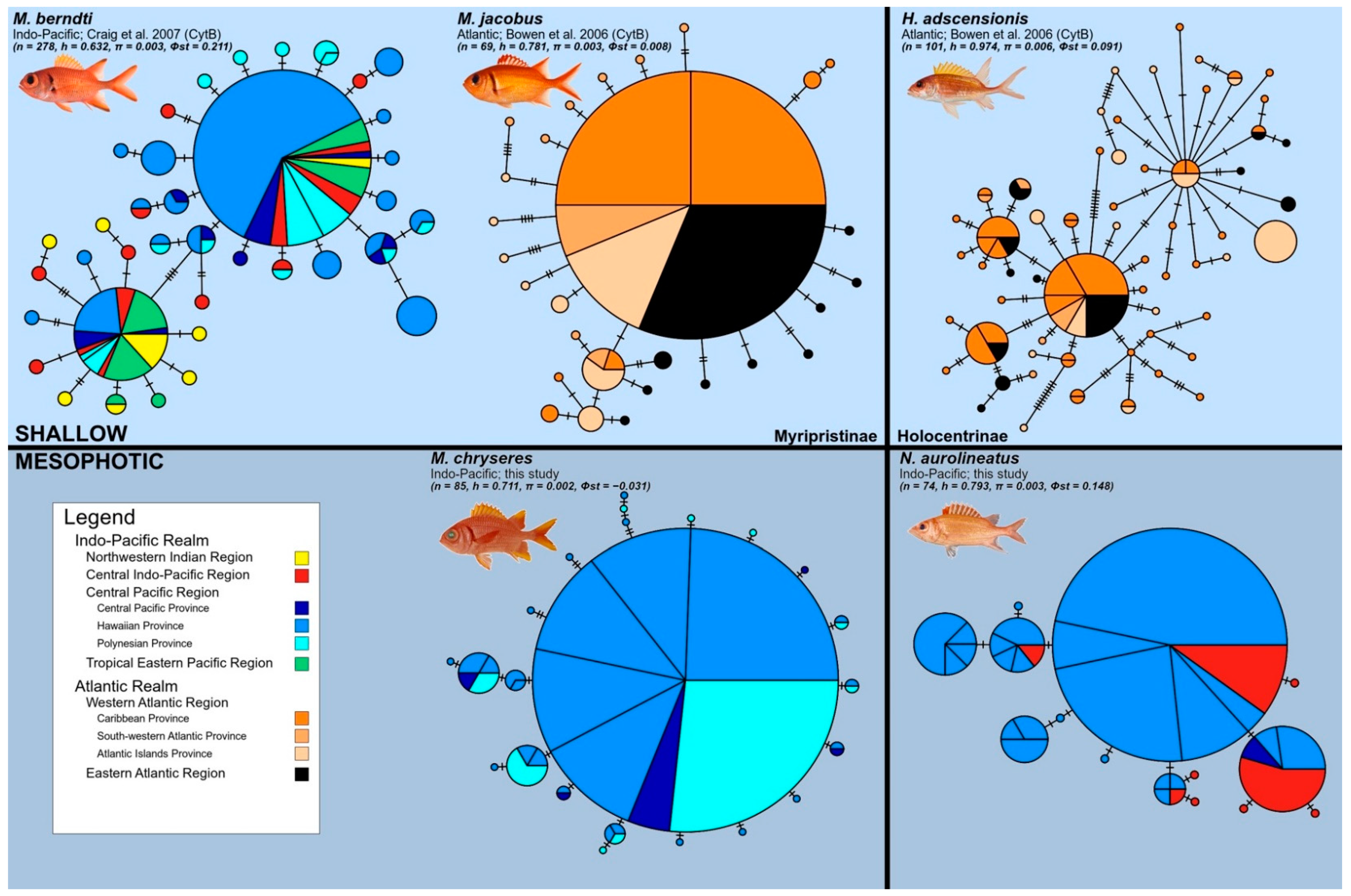
3.3. Multi-Species Haplotype Network Comparison
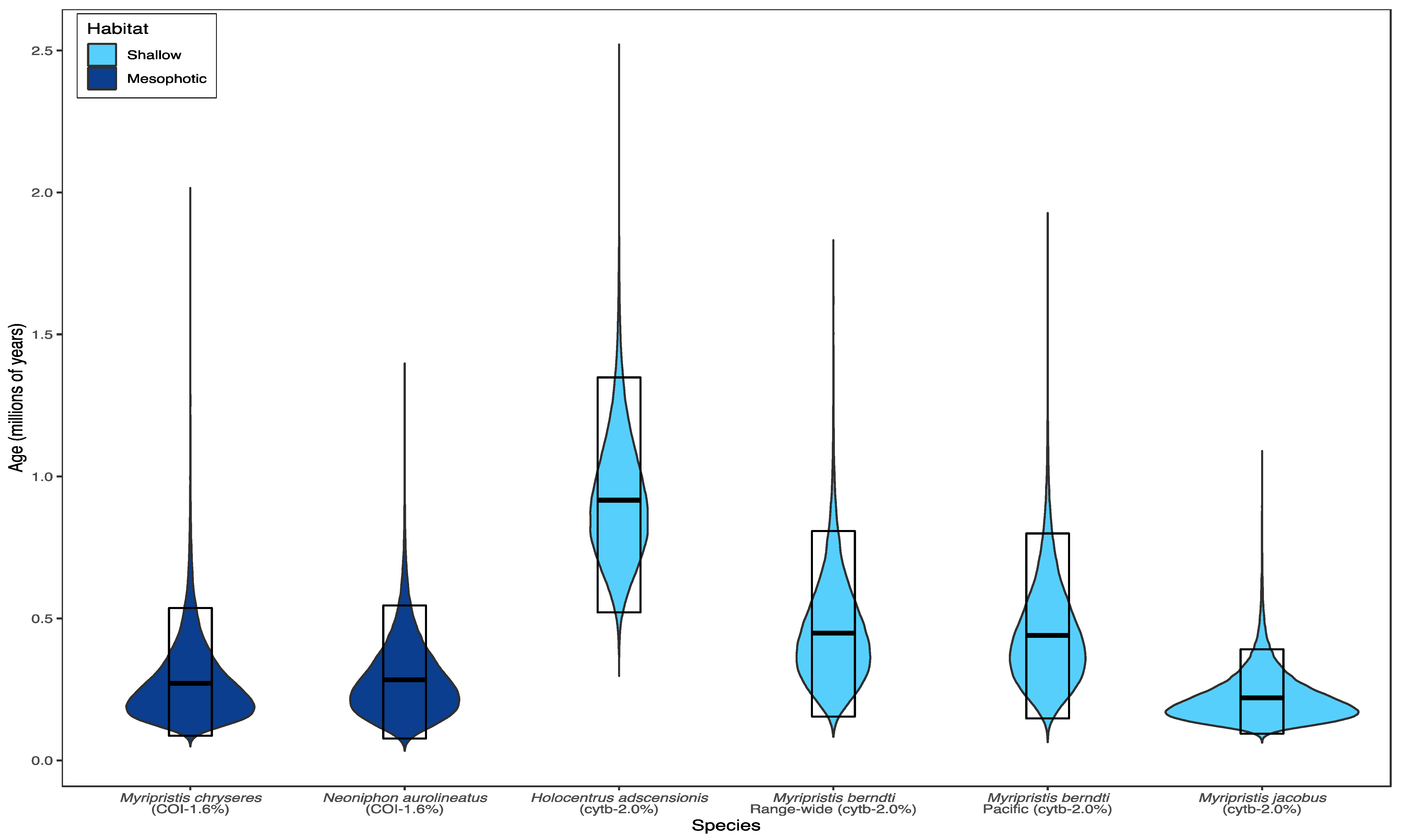
4. Discussion
4.1. Comparisons to Shallow Holocentrids
- M. berndti in the Indo-Pacific has haplotype diversity h = 0.632, nucleotide diversity π = 0.003, a range-wide coalescence at ~448,000 years (95% HPD: 154,000–807,000 years), and a restricted Pacific coalescence of ~441,000 years (95% HPD: 144,000–799,000 years), based on cytb sequences [32].
- M. jacobus in the Atlantic has h = 0.781, π = 0.003, and a coalescence at ~220,000 years (95% HPD: 94,000–391,000 years), based on cytb sequences [33].
- M. chryseres in the mesophotic Pacific has h = 0.712, π = 0.0024, and a coalescence at 272,000 years (95% HPD: 87,000–537,000 years), based on COI sequences (present study)
- H. adscensionis in the Atlantic has h = 0.974, π = 0.006, and a coalescence at ~917,000 years (95% HPD: 521,000–1.3 M years), based on cytb sequences [33].
- N. aurolineatus in the mesophotic Pacific has h = 0.793, π = 0.0028, and a coalescence at 284,000 years (95% HPD: 77,000–546,000 years), based on COI sequences (present study).
4.2. Age and Stability of Mesophotic Populations
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Riginos, C.; Douglas, K.E.; Jin, Y.; Shanahan, D.F.; Treml, E.A. Effects of Geography and Life History Traits on Genetic Differentiation in Benthic Marine Fishes. Ecography 2011, 34, 566–575. [Google Scholar] [CrossRef]
- White, C.; Selkoe, K.A.; Watson, J.; Siegel, D.A.; Zacherl, D.C.; Toonen, R.J. Ocean Currents Help Explain Population Genetic Structure. Proc. R. Soc. B Biol. Sci. 2010, 277, 1685–1694. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Toonen, R.J.; Andrews, K.R.; Baums, I.B.; Bird, C.E.; Concepcion, G.T.; Daly-Engel, T.S.; Eble, J.A.; Faucci, A.; Gaither, M.R.; Iacchei, M.; et al. Defining Boundaries for Ecosystem-Based Management: A Multispecies Case Study of Marine Connectivity across the Hawaiian Archipelago. J. Mar. Biol. 2011, 2011, e460173. [Google Scholar] [CrossRef] [PubMed]
- Selkoe, K.A.; Gaggiotti, O.E.; Laboratory, T.; Bowen, B.W.; Toonen, R.J. Emergent Patterns of Population Genetic Structure for a Coral Reef Community. Mol. Ecol. 2014, 23, 3064–3079. [Google Scholar] [CrossRef] [Green Version]
- Timm, J.; Figiel, M.; Kochzius, M. Contrasting Patterns in Species Boundaries and Evolution of Anemonefishes (Amphiprioninae, Pomacentridae) in the Centre of Marine Biodiversity. Mol. Phylogenetics Evol. 2008, 49, 268–276. [Google Scholar] [CrossRef]
- Barber, P.H.; Cheng, S.H.; Erdmann, M.V.; Tenggardjaja, K. Ambariyanto Evolution and Conservation of Marine Biodiversity in the Coral Triangle: Insights from Stomatopod Crustacea. In Phylogeography and Population Genetics in Crustacea; CRC Press: Boca Raton, FL, USA, 2011; ISBN 9780429064067. [Google Scholar]
- Carpenter, K.E.; Barber, P.H.; Crandall, E.D.; Ablan-Lagman, M.A.; Carmen, A.; Mahardika, G.N.; Manjaji-Matsumoto, B.M.; Juinio-Meñez, M.A.; Santos, M.D.; Starger, C.J.; et al. Comparative Phylogeography of the Coral Triangle and Implications for Marine Management. J. Mar. Biol. 2010, 2011, e396982. [Google Scholar] [CrossRef]
- Gaither, M.R.; Rocha, L.A. Origins of Species Richness in the Indo-Malay-Philippine Biodiversity Hotspot: Evidence for the Centre of Overlap Hypothesis. J. Biogeogr. 2013, 40, 1638–1648. [Google Scholar] [CrossRef]
- Bowen, B.W.; Gaither, M.R.; DiBattista, J.D.; Iacchei, M.; Andrews, K.R.; Grant, W.S.; Toonen, R.J.; Briggs, J.C. Comparative Phylogeography of the Ocean Planet. Proc. Natl. Acad. Sci. USA 2016, 113, 7962–7969. [Google Scholar] [CrossRef] [Green Version]
- Graves, J.E.; McDowell, J.R. Population Structure of Istiophorid Billfishes. Fish. Res. 2015, 166, 21–28. [Google Scholar] [CrossRef]
- Hinderstein, L.M.; Marr, J.C.A.; Martinez, F.A.; Dowgiallo, M.J.; Puglise, K.A.; Pyle, R.L.; Zawada, D.G.; Appeldoorn, R. Theme Section on “Mesophotic Coral Ecosystems: Characterization, Ecology, and Management”. Coral Reefs 2010, 29, 247–251. [Google Scholar] [CrossRef] [Green Version]
- Pochon, X.; Forsman, Z.H.; Spalding, H.L.; Padilla-Gamiño, J.L.; Smith, C.M.; Gates, R.D. Depth Specialization in Mesophotic Corals (Leptoseris spp.) and Associated Algal Symbionts in Hawai’i. R. Soc. Open Sci. 2015, 2, 140351. [Google Scholar] [CrossRef] [Green Version]
- Hurley, K.K.C.; Timmers, M.A.; Godwin, L.S.; Copus, J.M.; Skillings, D.J.; Toonen, R.J. An Assessment of Shallow and Mesophotic Reef Brachyuran Crab Assemblages on the South Shore of O’ahu, Hawai’i. Coral Reefs 2016, 35, 103–112. [Google Scholar] [CrossRef]
- Pinheiro, H.T.; Shepherd, B.; Castillo, C.; Abesamis, R.A.; Copus, J.M.; Pyle, R.L.; Greene, B.D.; Coleman, R.R.; Whitton, R.K.; Thillainath, E.; et al. Deep Reef Fishes in the World’s Epicenter of Marine Biodiversity. Coral Reefs 2019, 38, 985–995. [Google Scholar] [CrossRef]
- Pyle, R.L.; Kosaki, R.K.; Pinheiro, H.T.; Rocha, L.A.; Whitton, R.K.; Copus, J.M. Fishes: Biodiversity. In Mesophotic Coral Ecosystems; Loya, Y., Puglise, K.A., Bridge, T.C.L., Eds.; Springer International Publishing: Cham, Switzerland, 2019; pp. 749–777. ISBN 9783319927350. [Google Scholar]
- Tamir, R.; Eyal, G.; Kramer, N.; Laverick, J.H.; Loya, Y. Light Environment Drives the Shallow-to-Mesophotic Coral Community Transition. Ecosphere 2019, 10, e02839. [Google Scholar] [CrossRef] [Green Version]
- Kahng, S.E.; Copus, J.M.; Wagner, D. Mesophotic Coral Ecosystems. In Marine Animal Forests; Springer International Publishing: Cham, Switzerland, 2017; pp. 1–22. ISBN 9783319170015. [Google Scholar]
- Pyle, R.L. Advanced Technical Diving. In Mesophotic Coral Ecosystems; Loya, Y., Puglise, K.A., Bridge, T.C.L., Eds.; Coral Reefs of the World; Springer International Publishing: Cham, Switzerland, 2019; pp. 959–972. ISBN 9783319927350. [Google Scholar]
- Ovenden, J.R.; Salini, J.; O’connor, S.; Street, R. Pronounced Genetic Population Structure in a Potentially Vagile Fish Species (Pristipomoides Multidens, Teleostei; Perciformes; Lutjanidae) from the East Indies Triangle. Mol. Ecol. 2004, 13, 1991–1999. [Google Scholar] [CrossRef]
- Gaither, M.R.; Jones, S.A.; Kelley, C.; Newman, S.J.; Sorenson, L.; Bowen, B.W. High Connectivity in the Deepwater Snapper Pristipomoides Filamentosus (Lutjanidae) across the Indo-Pacific with Isolation of the Hawaiian Archipelago. PLoS ONE 2011, 6, e28913. [Google Scholar] [CrossRef] [Green Version]
- Andrews, K.R.; Copus, J.M.; Wilcox, C.; Williams, A.J.; Newman, S.J.; Wakefield, C.B.; Bowen, B.W. Range-Wide Population Structure of 3 Deepwater Eteline Snappers across the Indo-Pacific Basin. J. Hered. 2020, 111, 471–485. [Google Scholar] [CrossRef]
- Baker, E.K.; Puglise, K.A.; Harris, P.T. (Eds.) Mesophotic Coral Ecosystem-A Lifeboat for Coral Reefs? The United Nations Environment Programme and GRID-Arendal: Nairobi, Kenya; Arendal, Norway, 2016; ISBN 9788277011509. [Google Scholar]
- Semmler, R.F.; Hoot, W.C.; Reaka, M.L. Are Mesophotic Coral Ecosystems Distinct Communities and Can They Serve as Refugia for Shallow Reefs? Coral Reefs 2017, 36, 433–444. [Google Scholar] [CrossRef]
- Rocha, L.A.; Pinheiro, H.T.; Shepherd, B.; Papastamatiou, Y.P.; Luiz, O.J.; Pyle, R.L.; Bongaerts, P. Mesophotic Coral Ecosystems Are Threatened and Ecologically Distinct from Shallow Water Reefs. Science 2018, 361, 281–284. [Google Scholar] [CrossRef] [Green Version]
- Pyle, R.L.; Copus, J.M. Mesophotic Coral Ecosystems: Introduction and Overview. In Mesophotic Coral Ecosystems; Loya, Y., Puglise, K.A., Bridge, T.C.L., Eds.; Springer International Publishing: Cham, Switzerland, 2019; pp. 3–27. ISBN 9783319927350. [Google Scholar]
- Copus, J.M.; Bowen, B.W.; Kosaki, R.K.; Webster, J.M. The Habitat Persistence Hypothesis: A New Perspective on the Distribution of Coral-Reef Organisms. J. Biogeogr. 2022; in review. [Google Scholar]
- Ludt, W.B.; Rocha, L.A. Shifting Seas: The Impacts of Pleistocene Sea-Level Fluctuations on the Evolution of Tropical Marine Taxa. J. Biogeogr. 2015, 42, 25–38. [Google Scholar] [CrossRef] [Green Version]
- Grant, W.; Bowen, B. Shallow Population Histories in Deep Evolutionary Lineages of Marine Fishes: Insights from Sardines and Anchovies and Lessons for Conservation. J. Hered. 1998, 89, 415–426. [Google Scholar] [CrossRef]
- Randall, J.E. Reef and Shore Fishes of the Hawaiian Islands; Sea Grant College Program; University of Hawai’i: Honolulu, HI, USA, 2007; ISBN 9780824826987. [Google Scholar]
- Dornburg, A.; Moore, J.A.; Webster, R.; Warren, D.L.; Brandley, M.C.; Iglesias, T.L.; Wainwright, P.C.; Near, T.J. Molecular Phylogenetics of Squirrelfishes and Soldierfishes (Teleostei: Beryciformes: Holocentridae): Reconciling More than 100 Years of Taxonomic Confusion. Mol. Phylogenetics Evol. 2012, 65, 727–738. [Google Scholar] [CrossRef]
- Dornburg, A.; Moore, J.; Beaulieu, J.M.; Eytan, R.I.; Near, T.J. The Impact of Shifts in Marine Biodiversity Hotspots on Patterns of Range Evolution: Evidence from the Holocentridae (Squirrelfishes and Soldierfishes). Evolution 2015, 69, 146–161. [Google Scholar] [CrossRef]
- Craig, M.T.; Eble, J.A.; Bowen, B.W.; Robertson, D.R. High Genetic Connectivity across the Indian and Pacific Oceans in the Reef Fish Myripristis Berndti (Holocentridae). Mar. Ecol. Prog. Ser. 2007, 334, 245–254. [Google Scholar] [CrossRef] [Green Version]
- Bowen, B.W.; Bass, A.; Muss, A.; Carlin, J.; Robertson, D. Phylogeography of Two Atlantic Squirrelfishes (Family Holocentridae): Exploring Links between Pelagic Larval Duration and Population Connectivity. Mar. Biol. 2006, 149, 899–913. [Google Scholar] [CrossRef]
- Tyler, J.C.; Johnson, D.G.; Brothers, E.B.; Tyler, D.M.; Smith, L.C. Comparative Early Life Histories of Western Atlantic Squirrelfishes (Holocentridae): Age and Settlement of Rhynchichthys, Meeki, and Juvenile Stages. Bull. Mar. Sci. 1993, 53, 1126–1150. [Google Scholar]
- Lieske, E.; Myers, R. Coral Reef Fishes: Caribbean, Indian Ocean and Pacific Ocean Including the Red Sea; Collins Pocket Guide; HarperCollins: New York, NY, USA, 1994; ISBN 9780002199742. [Google Scholar]
- Heemstra, P.C.; Hissmann, K.; Fricke, H.; Smale, M.J.; Schauer, J. Fishes of the Deep Demersal Habitat at Ngazidja (Grand Comoro) Island, Western Indian Ocean: Coelacanth Research. S. Afr. J. Sci. 2006, 102, 444–460. [Google Scholar] [CrossRef]
- Kuiter, R.H.; Tonozuka, T. Pictorial Guide to Indonesian Reef Fishes; Zoonetics: Bali, Indonesia, 2001; ISBN 9789799818805. [Google Scholar]
- Seutin, G.; White, B.N.; Boag, P.T. Preservation of Avian Blood and Tissue Samples for DNA Analyses. Can. J. Zool. 1991, 69, 82–90. [Google Scholar] [CrossRef]
- Meeker, N.D.; Hutchinson, S.A.; Ho, L.; Trede, N.S. Method for Isolation of PCR-Ready Genomic DNA from Zebrafish Tissues. BioTechniques 2007, 43, 610–614. [Google Scholar] [CrossRef]
- Ward, R.D.; Zemlak, T.S.; Innes, B.H.; Last, P.R.; Hebert, P.D.N. DNA Barcoding Australia’s Fish Species. Philos. Trans. R. Soc. B Biol. Sci. 2005, 360, 1847–1857. [Google Scholar] [CrossRef]
- Baldwin, C.C.; Mounts, J.H.; Smith, D.G.; Weigt, L.A. Genetic Identification and Color Descriptions of Early Life-History Stages of Belizean Phaeoptyx and Astrapogon (Teleostei: Apogonidae) with Comments on Identification of Adult Phaeoptyx. Zootaxa 2009, 2008, 1–22. [Google Scholar] [CrossRef]
- R Core Team. R: A Language and Environment for Statistical Computing; R Core Team: Vienna, Austria, 2021. [Google Scholar]
- Pagès, H.; Aboyoun, P.; Gentleman, R.; DebRoy, S. Biostrings: Efficient Manipulation of Biological Strings; 2021. R package version 2.64.0. Available online: https://bioconductor.org/packages/Biostrings (accessed on 14 July 2022).
- Paradis, E.; Schliep, K. Ape 5.0: An Environment for Modern Phylogenetics and Evolutionary Analyses in R. Bioinformatics 2019, 35, 526–528. [Google Scholar] [CrossRef]
- Paradis, E. Pegas: An R Package for Population Genetics with an Integrated–Modular Approach. Bioinformatics 2010, 26, 419–420. [Google Scholar] [CrossRef] [Green Version]
- Chen, S. Hfufs: Fu’s Fs and Strobeck’s S Calculators for Arbitrary Parameters; 2020. R package version 0.1. Available online: https://github.com/swainechen/hfufs (accessed on 14 July 2022).
- Aktas, C. Haplotypes: Manipulating DNA Sequences and Estimating Unambiguous Haplotype Network with Statistical Parsimony; 2020. R package version 1.1.2. Available online: https://cran.r-project.org/package=haplotypes (accessed on 14 July 2022).
- Benjamini, Y.; Yekutieli, D. The Control of the False Discovery Rate in Multiple Testing under Dependency. Ann. Stat. 2001, 29, 1165–1188. [Google Scholar] [CrossRef]
- Narum, S.R. Beyond Bonferroni: Less Conservative Analyses for Conservation Genetics. Conserv. Genet. 2006, 7, 783–787. [Google Scholar] [CrossRef]
- Bouckaert, R.; Heled, J.; Kühnert, D.; Vaughan, T.; Wu, C.-H.; Xie, D.; Suchard, M.A.; Rambaut, A.; Drummond, A.J. BEAST 2: A Software Platform for Bayesian Evolutionary Analysis. PLoS Comput. Biol. 2014, 10, e1003537. [Google Scholar] [CrossRef] [Green Version]
- Bouckaert, R.R.; Drummond, A.J. BModelTest: Bayesian Phylogenetic Site Model Averaging and Model Comparison. BMC Evol. Biol. 2017, 17, 42. [Google Scholar] [CrossRef] [Green Version]
- Kartavtsev, Y.P.; Lee, J.-S. Analysis of Nucleotide Diversity at the Cytochrome b and Cytochrome Oxidase 1 Genes at the Population, Species, and Genus Levels. Russ. J. Genet. 2006, 42, 341–362. [Google Scholar] [CrossRef]
- Tobe, S.S.; Kitchener, A.C.; Linacre, A.M.T. Reconstructing Mammalian Phylogenies: A Detailed Comparison of the Cytochrome b and Cytochrome Oxidase Subunit I Mitochondrial Genes. PLoS ONE 2010, 5, e14156. [Google Scholar] [CrossRef] [Green Version]
- Lessios, H.A. The Great American Schism: Divergence of Marine Organisms after the Rise of the Central American Isthmus. Annu. Rev. Ecol. Evol. Syst. 2008, 39, 63–91. [Google Scholar] [CrossRef] [Green Version]
- Rambaut, A.; Drummond, A.J.; Xie, D.; Baele, G.; Suchard, M.A. Posterior Summarization in Bayesian Phylogenetics Using Tracer 1.7. Syst. Biol. 2018, 67, 901–904. [Google Scholar] [CrossRef] [Green Version]
- Excoffier, L.; Lischer, H.E.L. Arlequin Suite Ver 3.5: A New Series of Programs to Perform Population Genetics Analyses under Linux and Windows. Mol. Ecol. Resour. 2010, 10, 564–567. [Google Scholar] [CrossRef]
- Lischer, H.E.L.; Excoffier, L. PGDSpider: An Automated Data Conversion Tool for Connecting Population Genetics and Genomics Programs. Bioinformatics 2012, 28, 298–299. [Google Scholar] [CrossRef] [Green Version]
- Wickham, H. Ggplot2: Elegant Graphics for Data Analysis; Springer: New York, NY, USA, 2016; ISBN 9783319242774. [Google Scholar]
- Keitt, T. ColorRamps: Builds Color Tables; 2022. R package version 2.3.1. Available online: https://cran.r-project.org/package=colorRamps (accessed on 14 July 2022).
- Pedersen, T.L. Patchwork: The Composer of Plots; 2020. R package version 1.1.1. Available online: https://cran.r-project.org/package=patchwork (accessed on 14 July 2022).
- Ooms, J. Magick: Advanced Graphics and Image-Processing in R; 2021. R package version 2.7.3. Available online: https://cran.r-project.org/package=magick (accessed on 14 July 2022).
- Kulbicki, M.; Parravicini, V.; Bellwood, D.R.; Arias-Gonzàlez, E.; Chabanet, P.; Floeter, S.R.; Friedlander, A.; McPherson, J.; Myers, R.E.; Vigliola, L.; et al. Global Biogeography of Reef Fishes: A Hierarchical Quantitative Delineation of Regions. PLoS ONE 2013, 8, e81847. [Google Scholar] [CrossRef]
- Lefèvre, A.; Lecomte-Finiger, R. Durée de La Vie Larvaire Pélagique de Deux Espèces Récifales, Stegastes Nigricans (Pomacenridae) et Neoniphon Sammara (Holocentridae) de l’île de Moorea (Polynésie Française). Cybium 1995, 19, 401–409. [Google Scholar]
- Voris, H.K. Maps of Pleistocene Sea Levels in Southeast Asia: Shorelines, River Systems and Time Durations. J. Biogeogr. 2000, 27, 1153–1167. [Google Scholar] [CrossRef] [Green Version]
- Myers, R.F. Micronesian Reef Fishes: A Comprehensive Guide to the Coral Reef Fishes of Micronesia; Coral Graphics: New York, NY, USA, 1999; ISBN 978-0-9621564-5-8. [Google Scholar]
- Kosaki, R.K.; Pyle, R.L.; Randall, J.E.; Irons, D.K. New Records of Fishes from Johnston Atoll, with Notes on Biogeography. Ph.D. Thesis, University of Hawai‘i at Mānoa, Honolulu, HI, USA, 1991. [Google Scholar]
- Fauvelot, C.; Bernardi, G.; Planes, S. Reductions in the Mitochondrial DNA Diversity of Coral Reef Fish Provide Evidence of Population Bottlenecks Resulting from Holocene Sea-Level Change. Evolution 2003, 57, 1571–1583. [Google Scholar] [CrossRef]
- Ludt, W.B.; Bernal, M.A.; Bowen, B.W.; Rocha, L.A. Living in the Past: Phylogeography and Population Histories of Indo-Pacific Wrasses (Genus Halichoeres) in Shallow Lagoons versus Outer Reef Slopes. PLoS ONE 2012, 7, e38042. [Google Scholar] [CrossRef] [Green Version]
- Delrieu-Trottin, E.; Brosseau-Acquaviva, L.; Mona, S.; Neglia, V.; Giles, E.C.; Rapu-Edmunds, C.; Saenz-Agudelo, P. Understanding the Origin of the Most Isolated Endemic Reef Fish Fauna of the Indo-Pacific: Coral Reef Fishes of Rapa Nui. J. Biogeogr. 2019, 46, 723–733. [Google Scholar] [CrossRef]
- Kimura, M. The Neutral Theory of Molecular Evolution; Cambridge University Press: Cambridge, MA, USA, 1983; ISBN 9780521317931. [Google Scholar]
- McDowell, J.R.; Brightman, H.L. High Level of Genetic Connectivity in a Deep-Water Reef Fish, Caulolatilus Microps. J. Fish Biol. 2018, 93, 766–777. [Google Scholar] [CrossRef] [PubMed]
- Baco, A.R.; Etter, R.J.; Ribeiro, P.A.; von der Heyden, S.; Beerli, P.; Kinlan, B.P. A Synthesis of Genetic Connectivity in Deep-Sea Fauna and Implications for Marine Reserve Design. Mol. Ecol. 2016, 25, 3276–3298. [Google Scholar] [CrossRef] [PubMed]
- Tenggardjaja, K.A.; Bowen, B.W.; Bernardi, G. Vertical and Horizontal Genetic Connectivity in Chromis Verater, an Endemic Damselfish Found on Shallow and Mesophotic Reefs in the Hawaiian Archipelago and Adjacent Johnston Atoll. PLoS ONE 2014, 9, e115493. [Google Scholar] [CrossRef] [Green Version]
- Nelson, J.S.; Grande, T.C.; Wilson, M.V.H. Fishes of the World; Wiley: Hoboken, NJ, USA, 2016; ISBN 9781119220824. [Google Scholar]
- Weersing, K.; Toonen, R.J. Population Genetics, Larval Dispersal, and Connectivity in Marine Systems. Mar. Ecol. Prog. Ser. 2009, 393, 1–12. [Google Scholar] [CrossRef] [Green Version]
- Bellwood, D.R.; Fisher, R. Relative Swimming Speeds in Reef Fish Larvae. Mar. Ecol. Prog. Ser. 2001, 211, 299–303. [Google Scholar] [CrossRef]
- Greenfield, D.W. Revision of the Squirrelfish Genus Myripristis Cuvier (Pisces: Holocentridae). Sci. Bull. Nat. Hist. Mus. Los Angeles Co. 1974, 19, 1–54. [Google Scholar]
- Rabosky, D.L.; Chang, J.; Title, P.O.; Cowman, P.F.; Sallan, L.; Friedman, M.; Kaschner, K.; Garilao, C.; Near, T.J.; Coll, M.; et al. An Inverse Latitudinal Gradient in Speciation Rate for Marine Fishes. Nature 2018, 559, 392–395. [Google Scholar] [CrossRef]
- Randall, J.E.; Greenfield, D.W. Revision of the Indo-Pacific Holocentrid Fishes of the Genus Myripristis, with Descriptions of Three New Species. Indo-Pac. Fish. 1996, 25, 1–61. [Google Scholar]
- Chave, E.H.; Mundy, B.C. Deep-Sea Benthic Fish of the Hawaiian Archipelago, Cross Seamount, and Johnston Atoll. Oceanogr. Lit. Rev. 1995, 6, 486. [Google Scholar]
- Briggs, J.C.; Bowen, B.W. A Realignment of Marine Biogeographic Provinces with Particular Reference to Fish Distributions. J. Biogeogr. 2012, 39, 12–30. [Google Scholar] [CrossRef]
- Gasparini, J.L.; Floeter, S.R. The Shore Fishes of Trindade Island, Western South Atlantic. J. Nat. Hist. 2001, 35, 1639–1656. [Google Scholar] [CrossRef]
- Wyatt, J. The Biology, Ecology and Bionomics of the Squirrelfishes, Holocentridae. In Caribbean Coral Reef Fishery Resources; Munro, J.L., Ed.; ICLARM Stud. Rev; International Center for Living Aquatic Resources Management: Pulau Pinang, Malaysia, 1983; Volume 7, pp. 50–58. [Google Scholar]
- Copus, J.M.; Montgomery, W.L.; Forsman, Z.H.; Bowen, B.W.; Toonen, R.J. Geopolitical Species Revisited: Genomic and Morphological Data Indicate That the Roundtail Chub Gila Robusta Species Complex (Teleostei, Cyprinidae) Is a Single Species. PeerJ 2018, 6, e5605. [Google Scholar] [CrossRef] [PubMed]
- Ray, J.M.; Wood, R.M.; Simons, A.M. Phylogeography and Post-Glacial Colonization Patterns of the Rainbow Darter, Etheostoma Caeruleum (Teleostei: Percidae). J. Biogeogr. 2006, 33, 1550–1558. [Google Scholar] [CrossRef]
- White, K.N.; Ohara, T.; Fujii, T.; Kawamura, I.; Mizuyama, M.; Montenegro, J.; Shikiba, H.; Naruse, T.; McClelland, T.Y.; Denis, V.; et al. Typhoon Damage on a Shallow Mesophotic Reef in Okinawa, Japan. PeerJ 2013, 1, e151. [Google Scholar] [CrossRef] [PubMed]
- Millstein, R.L. Natural Selection as a Population-Level Causal Process. Br. J. Philos. Sci. 2006, 57, 627–653. [Google Scholar] [CrossRef] [Green Version]
- Taylor, M.L.; Roterman, C.N. Invertebrate Population Genetics across Earth’s Largest Habitat: The Deep-Sea Floor. Mol. Ecol. 2017, 26, 4872–4896. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wright, S. Evolution in Mendelian Populations. Genetics 1931, 16, 97–159. [Google Scholar] [CrossRef]
- Avise, J.C. Molecular Markers, Natural History and Evolution, 2nd ed.; Sinauer Associates: Sunderland, MA, USA, 2004; ISBN 9781461523819. [Google Scholar]
- Hartl, D.L.; Clark, A.G. Principles of Population Genetics, 4th ed.; Sinauer and Associates: Sunderland, MA, USA, 2006; ISBN 9780878933082. [Google Scholar]
- Hedgecock, D.; Pudovkin, A.I. Sweepstakes Reproductive Success in Highly Fecund Marine Fish and Shellfish: A Review and Commentary. Bull. Mar. Sci. 2011, 87, 971–1002. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Location | n | S | H | h | π | D | F |
|---|---|---|---|---|---|---|---|
| Manawai/Pearl and Hermes | 20 | 9 | 9 | 0.705 | 0.0020 | −1.969 | −6.279 |
| Kapou/Lisianski | 6 | 1 | 2 | 0.333 | 0.0006 | −0.933 | −0.023 |
| Ni‘ihau | 8 | 4 | 4 | 0.643 | 0.0022 | −1.030 | −0.914 |
| Maui | 10 | 8 | 6 | 0.778 | 0.0039 | −1.161 | −1.756 |
| Johnston Atoll | 10 | 5 | 5 | 0.756 | 0.0025 | −1.035 | −1.596 |
| Pohnpei | 6 | 5 | 5 | 0.933 | 0.0031 | −1.337 | −2.526 |
| Rarotonga | 25 | 11 | 10 | 0.757 | 0.0026 | −1.781 | −5.732 |
| Overall | 85 | 20 | 23 | 0.712 | 0.0024 | −2.002 | −22.942 |
| Location | n | S | H | h | π | D | F |
|---|---|---|---|---|---|---|---|
| Hōlanikū/Kure | 1 | - | 1 | - | - | - | - |
| Manawai/Pearl and Hermes | 30 | 7 | 8 | 0.752 | 0.0025 | −0.687 | −2.708 |
| Pioneer Bank | 3 | 2 | 2 | 0.667 | 0.0025 | 0.000 | 1.032 |
| Lalo/French Frigate Shoals | 14 | 6 | 6 | 0.736 | 0.0026 | −0.893 | −1.921 |
| Nihoa | 6 | 3 | 4 | 0.800 | 0.0022 | −0.447 | −1.466 |
| Maui | 3 | 2 | 3 | 1.000 | 0.0025 | 0.000 | −1.216 |
| Pohnpei | 1 | - | 1 | - | - | - | - |
| Luzon | 16 | 7 | 9 | 0.850 | 0.0030 | −0.865 | −5.256 |
| Overall | 74 | 12 | 14 | 0.793 | 0.0028 | −1.088 | −6.681 |
| Species | Habitat | ΦST | h | π | Age (Ma) | Locus | Ref. |
|---|---|---|---|---|---|---|---|
| M. chryseres | deep | −0.031 | 0.712 | 0.0024 | 0.272 | COI | - |
| N. aurolineatus | deep | 0.148 a | 0.793 | 0.0028 | 0.284 | COI | - |
| M. berndti | shallow | −0.007 b | 0.632 c | 0.003 c | 0.448 | cytb | [32] |
| M. jacobus | shallow | 0.008 | 0.781 | 0.003 | 0.220 | cytb | [33] |
| H. adscensionis | shallow | 0.091a | 0.974 | 0.006 | 0.917 | cytb | [33] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Copus, J.M.; Walsh, C.A.J.; Hoban, M.L.; Lee, A.M.; Pyle, R.L.; Kosaki, R.K.; Toonen, R.J.; Bowen, B.W. Phylogeography of Mesophotic Coral Ecosystems: Squirrelfish and Soldierfish (Holocentriformes: Holocentridae). Diversity 2022, 14, 691. https://doi.org/10.3390/d14080691
Copus JM, Walsh CAJ, Hoban ML, Lee AM, Pyle RL, Kosaki RK, Toonen RJ, Bowen BW. Phylogeography of Mesophotic Coral Ecosystems: Squirrelfish and Soldierfish (Holocentriformes: Holocentridae). Diversity. 2022; 14(8):691. https://doi.org/10.3390/d14080691
Chicago/Turabian StyleCopus, Joshua M., Cameron A. J. Walsh, Mykle L. Hoban, Anne M. Lee, Richard L. Pyle, Randall K. Kosaki, Robert J. Toonen, and Brian W. Bowen. 2022. "Phylogeography of Mesophotic Coral Ecosystems: Squirrelfish and Soldierfish (Holocentriformes: Holocentridae)" Diversity 14, no. 8: 691. https://doi.org/10.3390/d14080691