
Cladocera (Crustacea: Branchiopoda) of Man-Made Lakes at the Northeast Part of the United Arab Emirates with a Hypothesis on Their Origin

 ,
, 
 and
and
Abstract
:1. Introduction
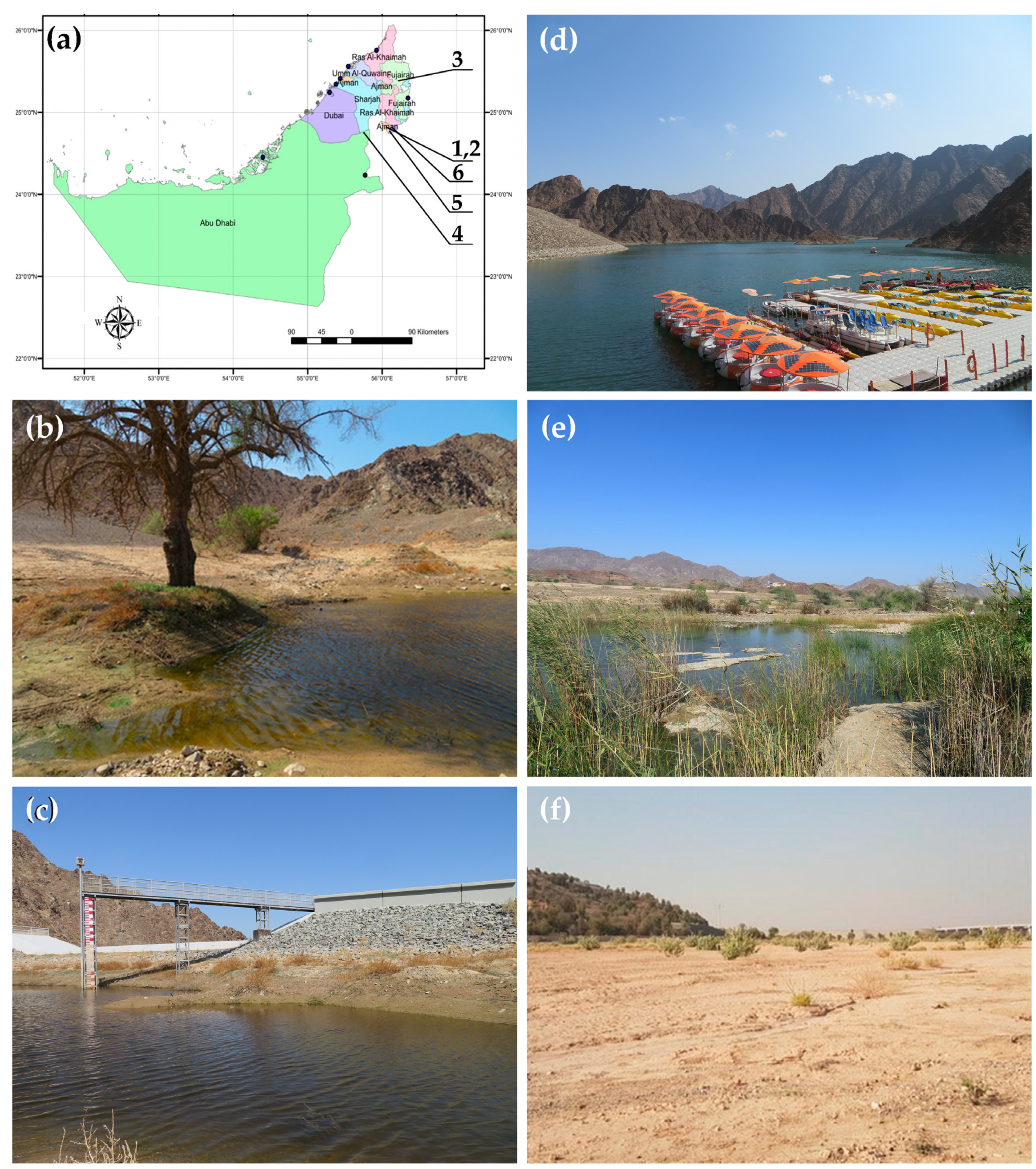
2. Materials and Methods
2.1. Sampling and Morphological Analysis
2.2. Abbreviations
2.3. Genetics
3. Results
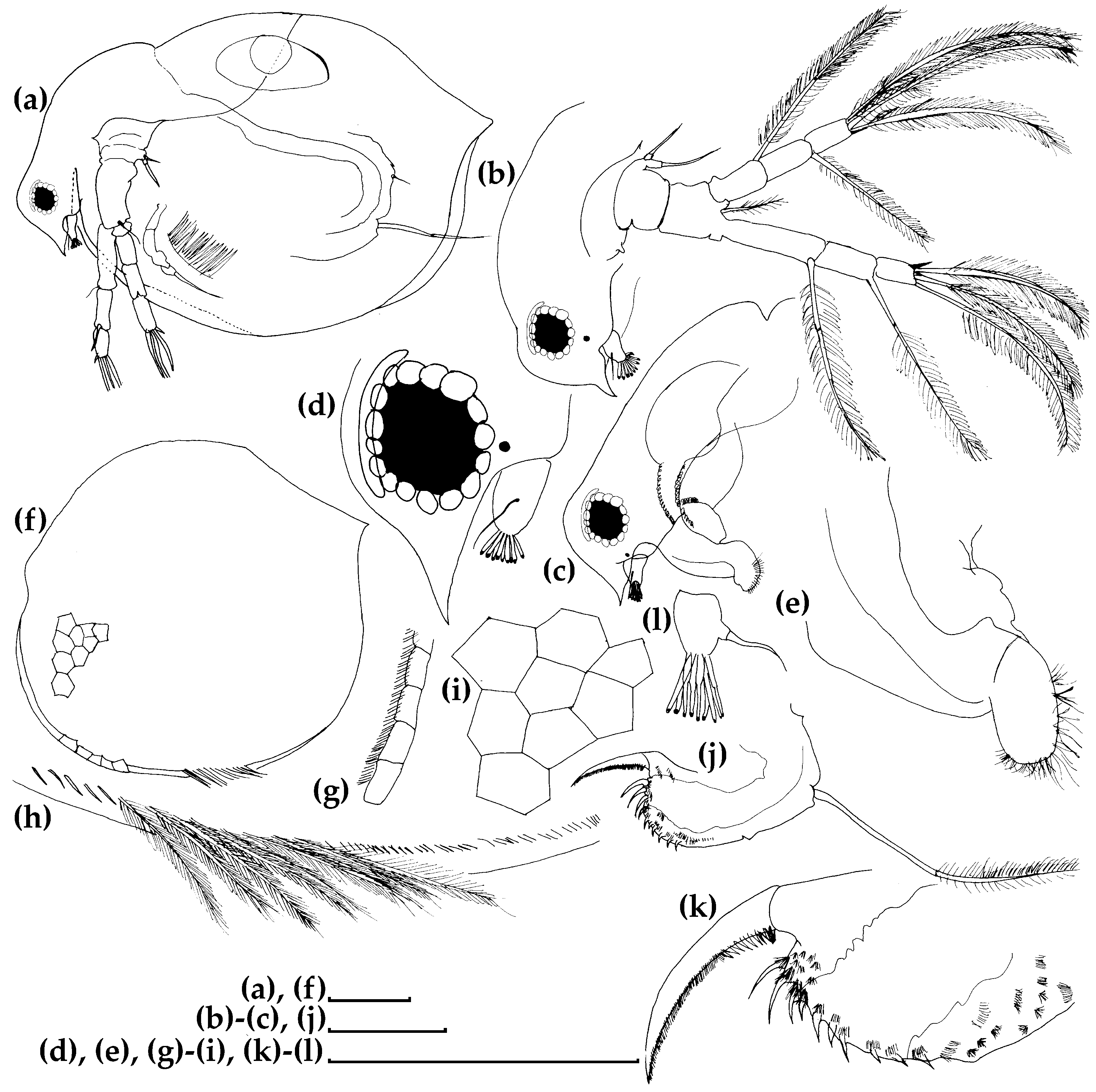
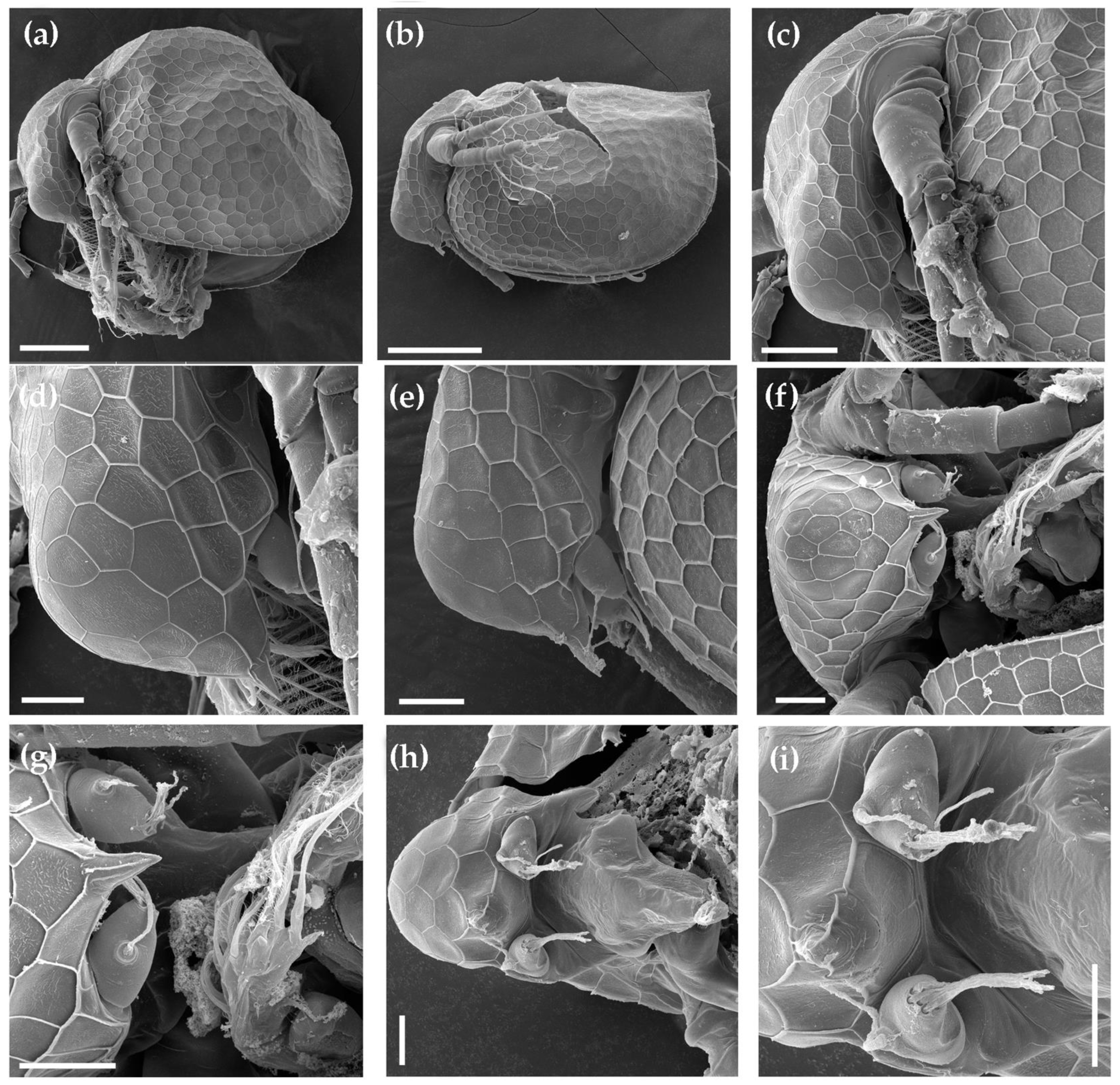
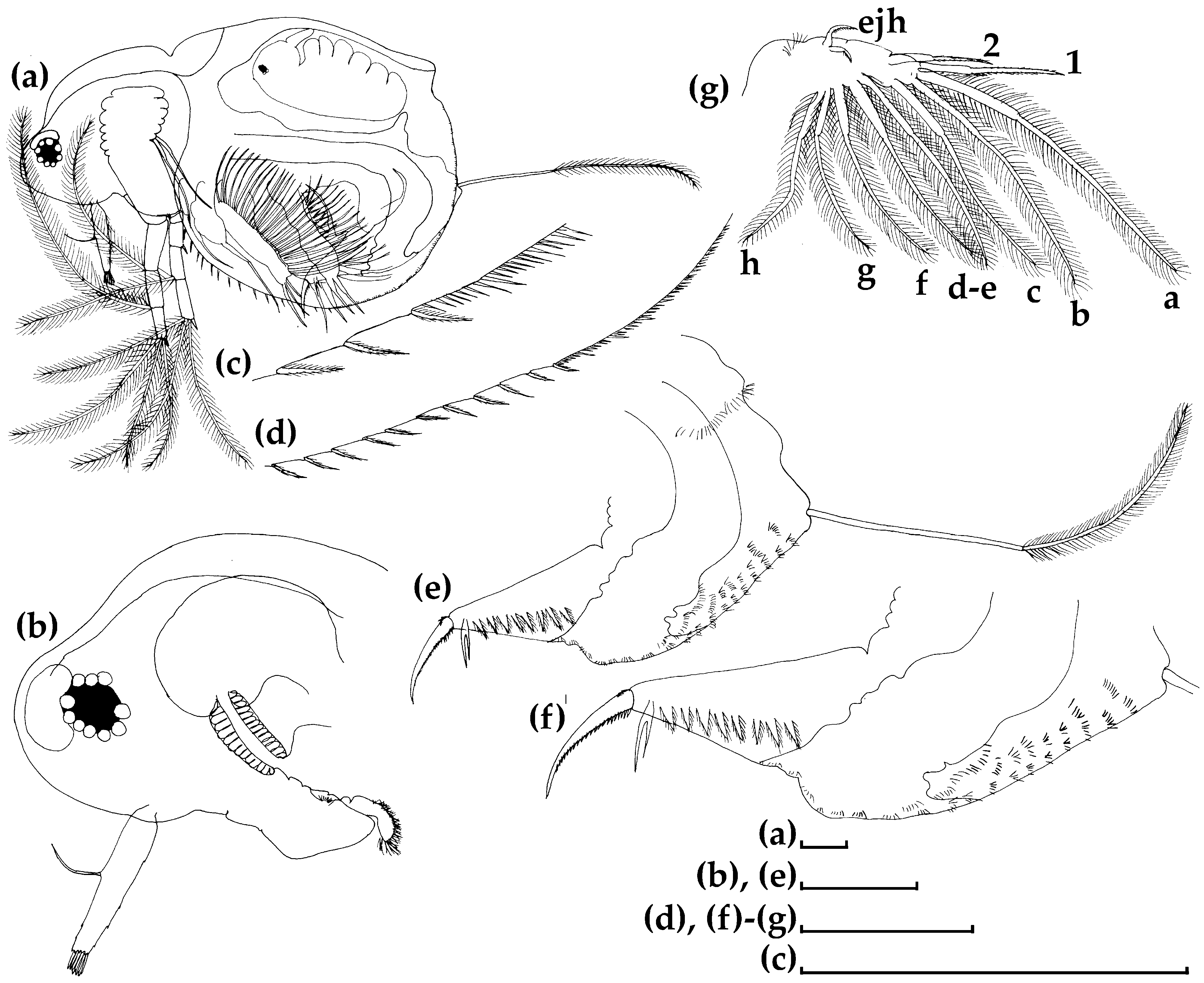
3.1. Morphological Account
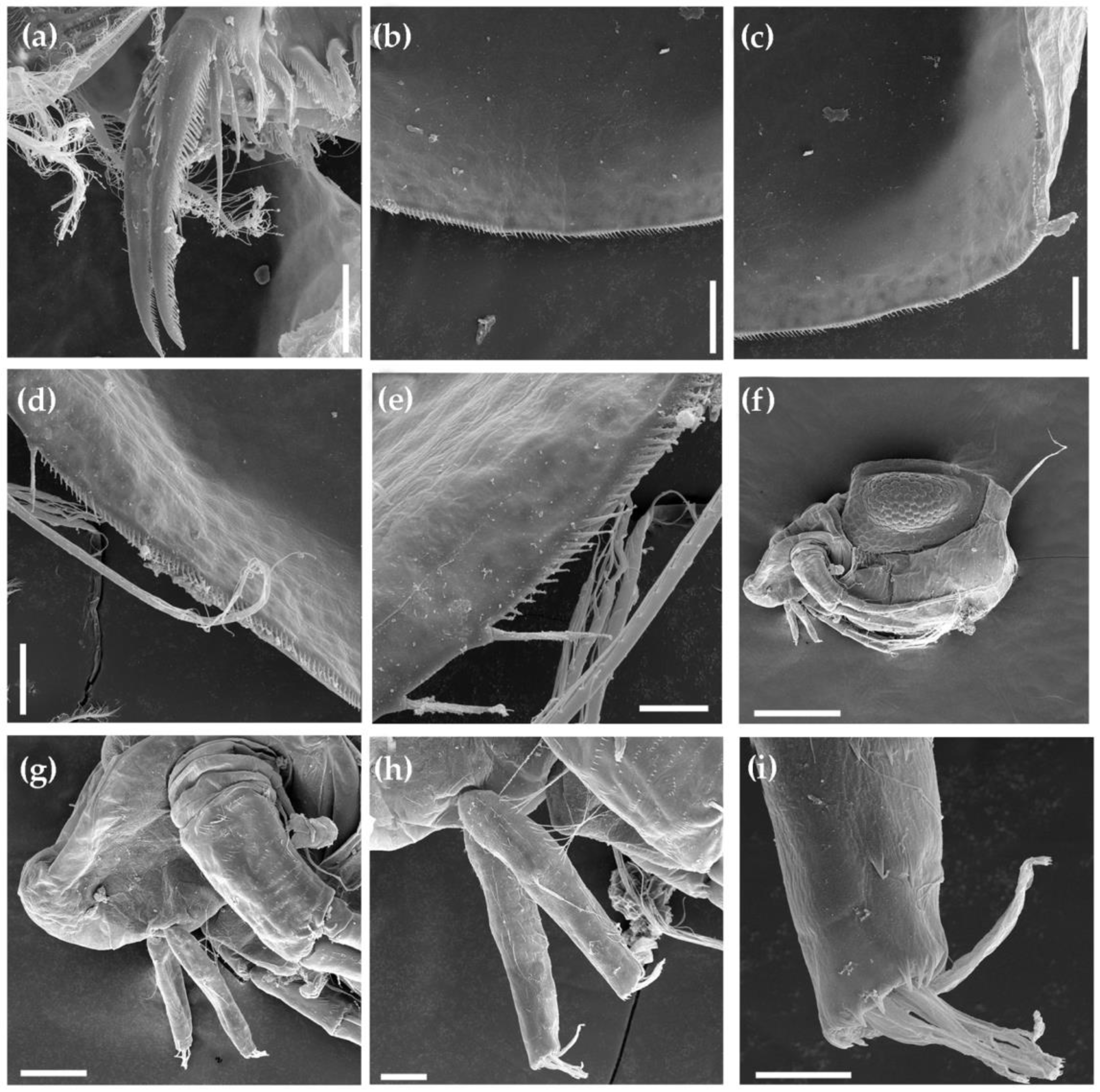
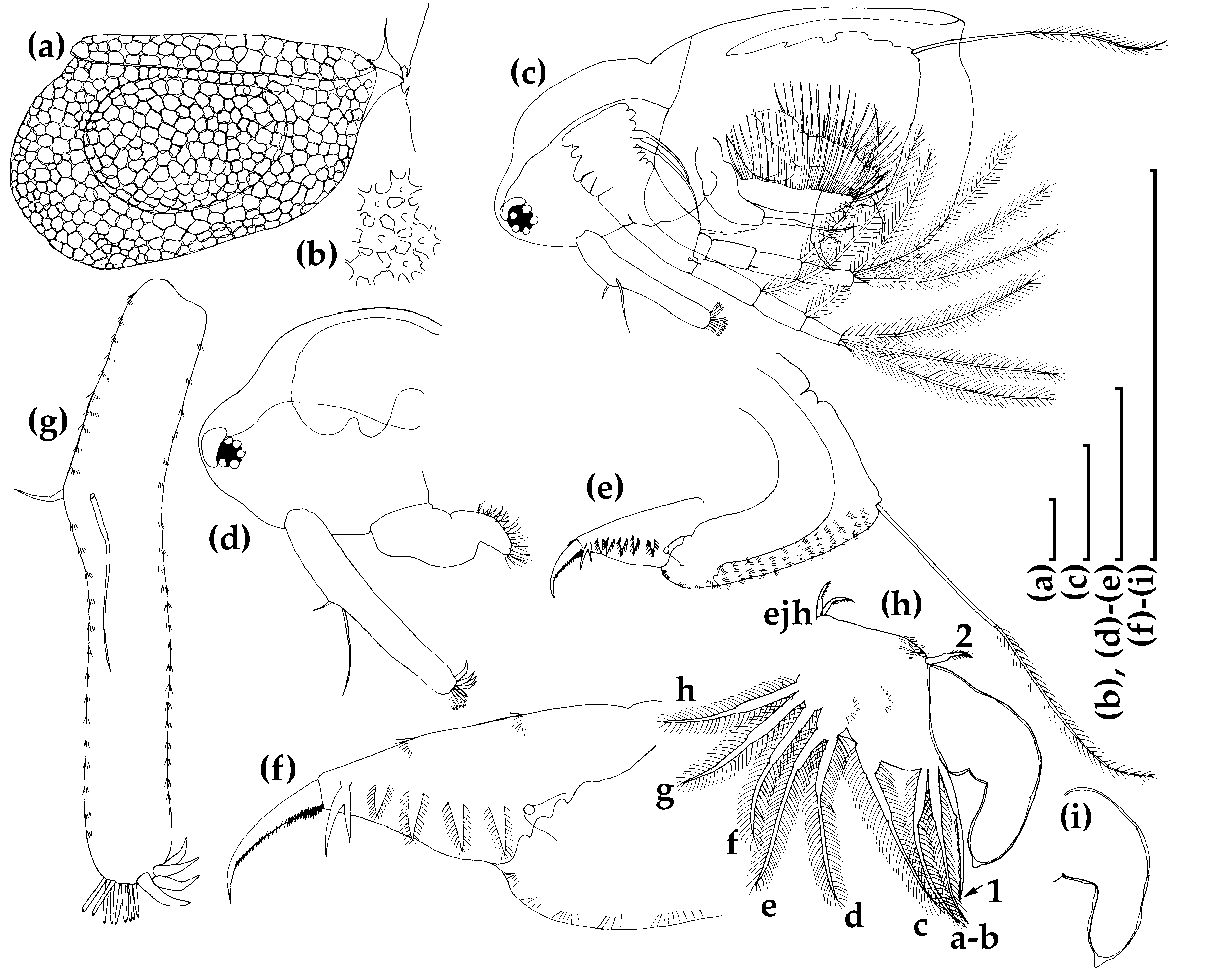
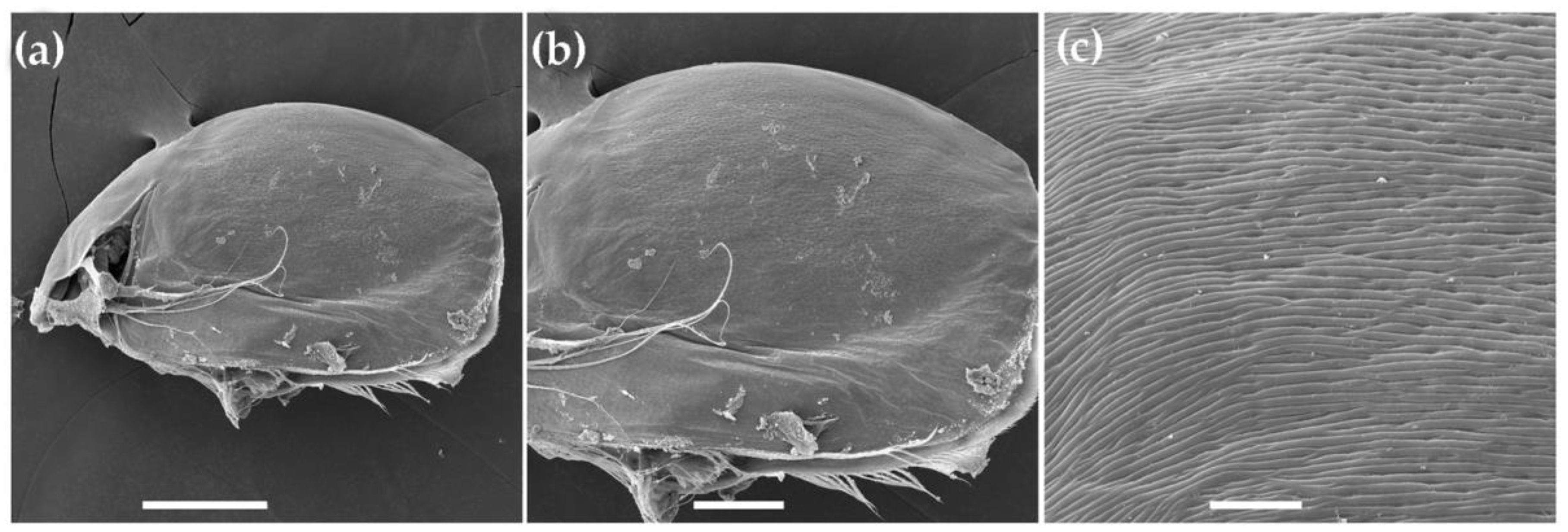
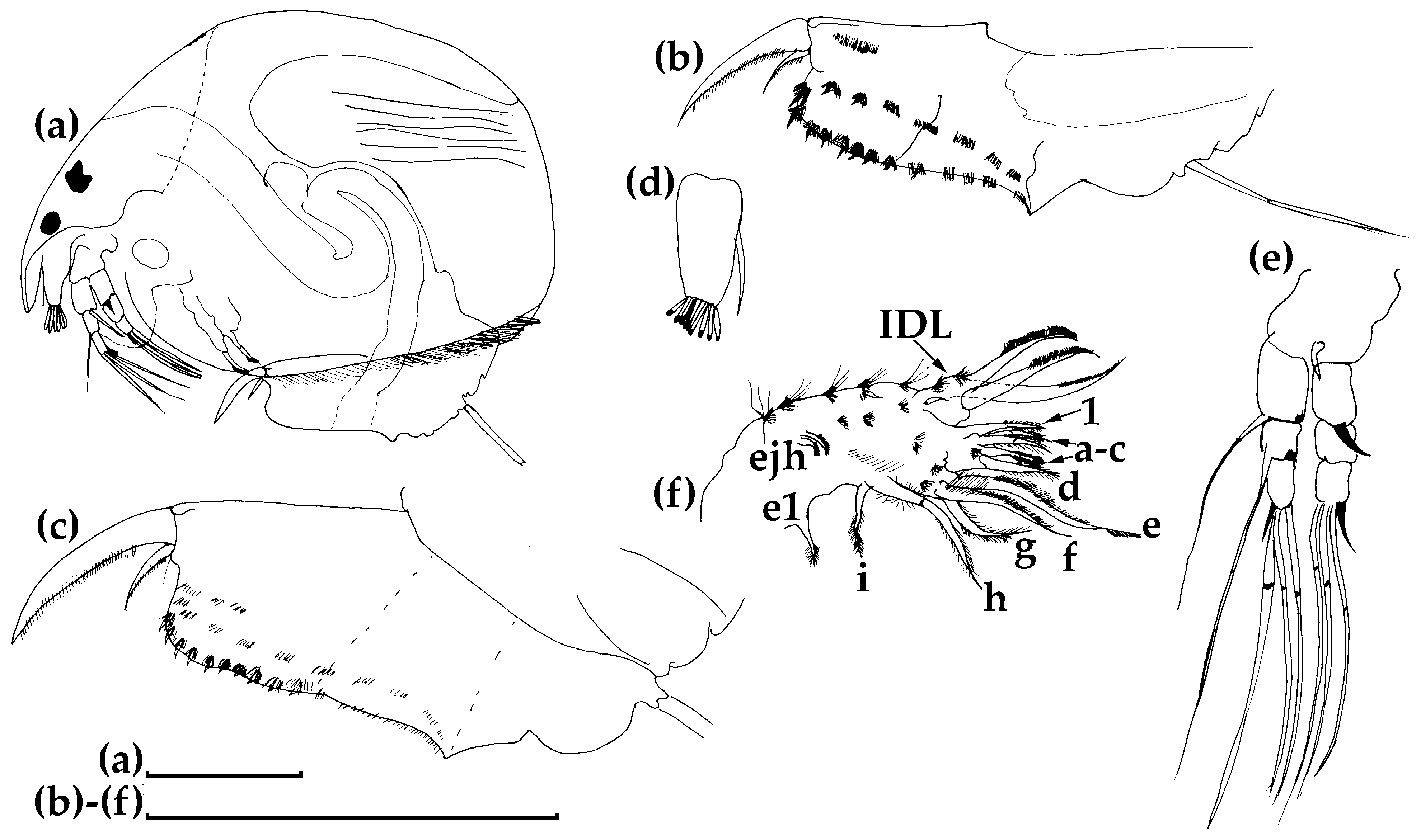
3.1.1. Ceriodaphnia cf. cornuta
3.1.2. Daphnia (Ctenodaphnia) arabica
3.1.3. Moina cf. micrura
3.1.4. Anthalona mediterranea
3.1.5. Coronatella anemae
3.2. Genetic Account
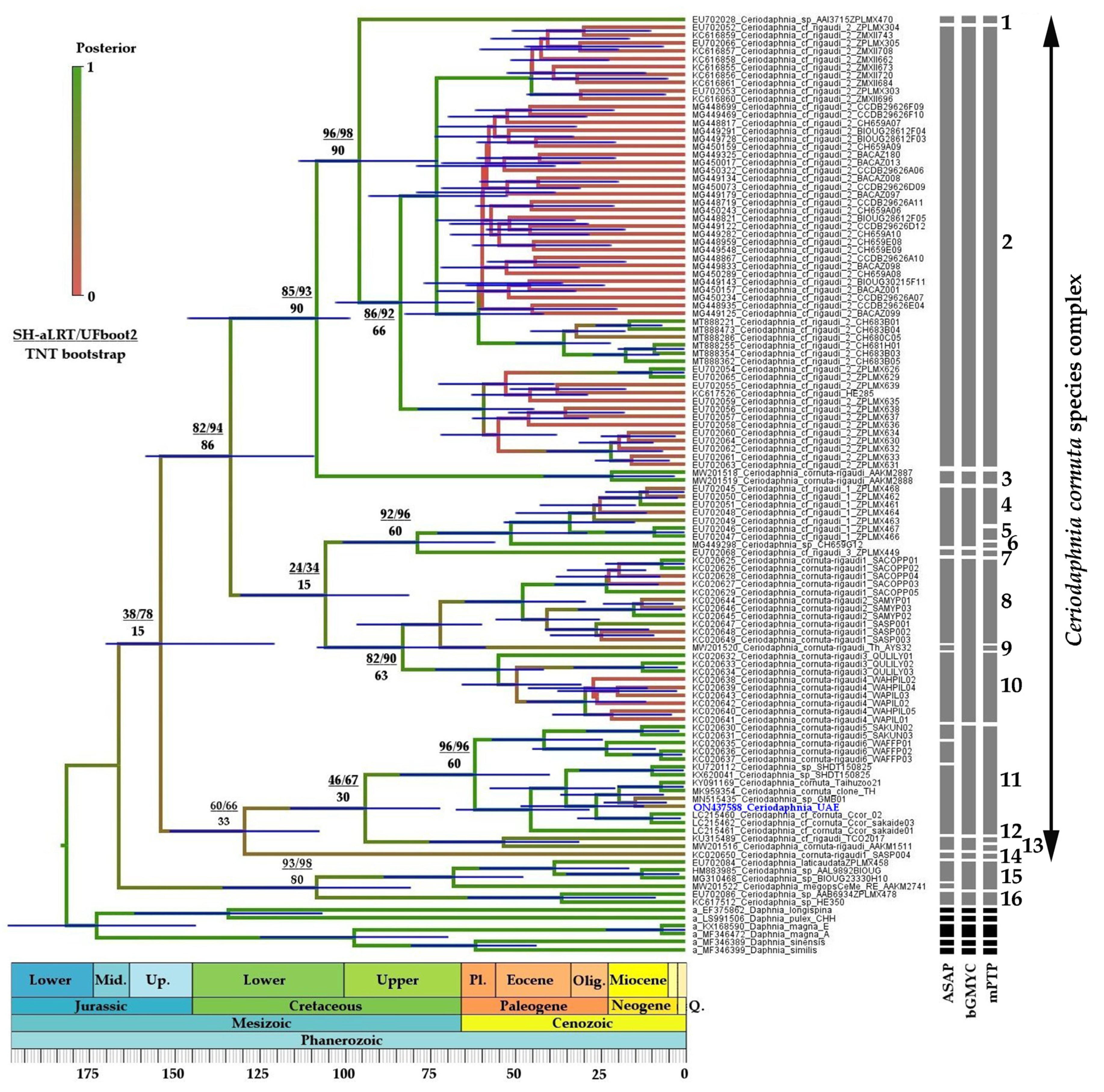
3.2.1. Ceriodaphnia
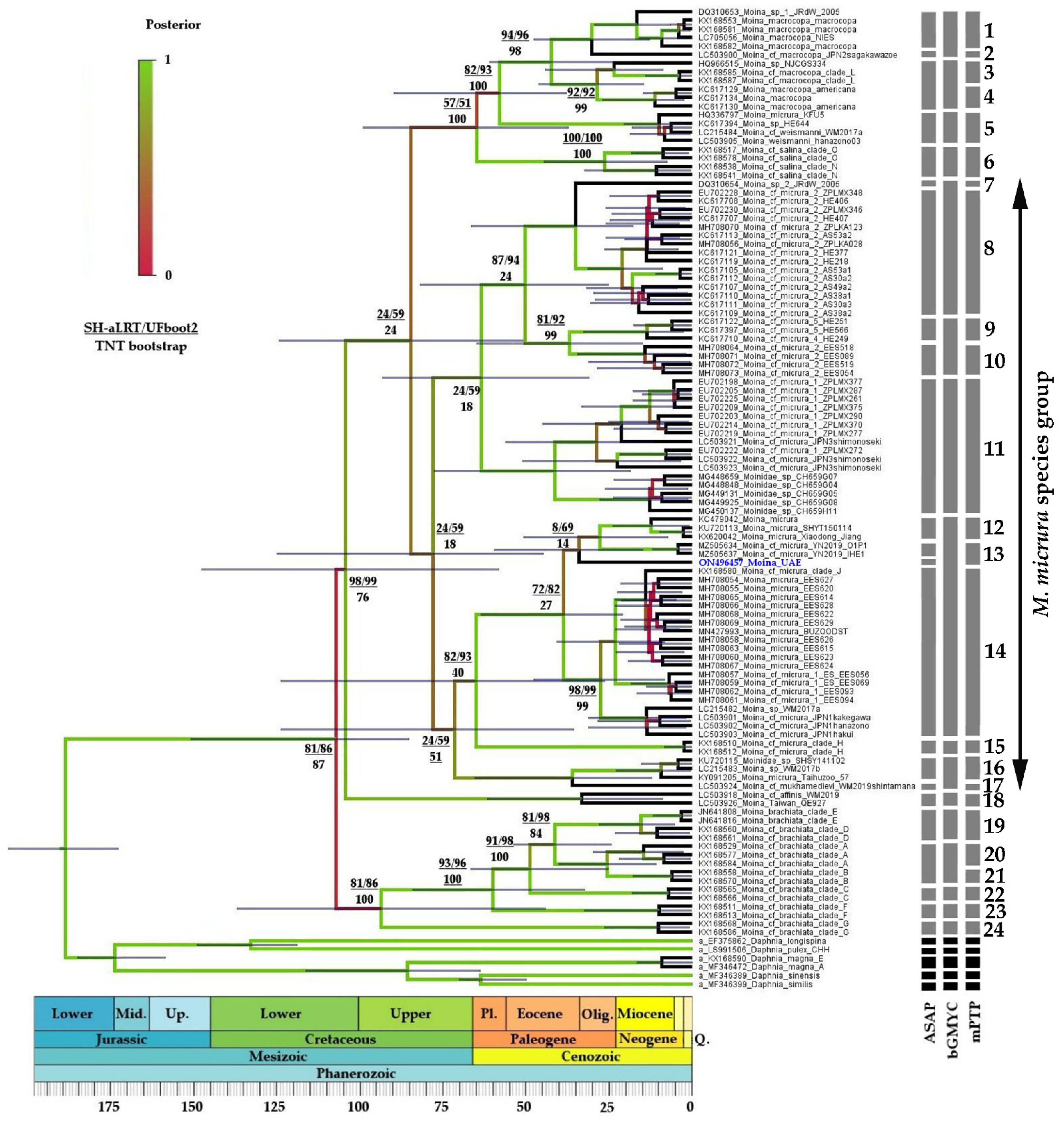
3.2.2. Moina
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- CBD. The Convention on Biological Diversity. Available online: https://www.cbd.int (accessed on 7 July 2022).
- Norman, M. Biodiversity hotspots hevisited. Bioscience 2003, 53, 916. [Google Scholar] [CrossRef] [Green Version]
- Bellard, C.; Leclerc, C.; Leroy, B.; Bakkenes, M.; Veloz, S.; Thuiller, W.; Courchamp, F. Vulnerability of biodiversity hotspots to global change. Glob. Ecol. Biogeogr. 2014, 23, 1376–1386. [Google Scholar] [CrossRef]
- Maciorowski, G.; Jankowiak, Ł.; Sparks, T.H.; Polakowski, M.; Tryjanowski, P. Biodiversity hotspots at a small scale: The importance of eagles’ nests to many other animals. Ecology 2021, 102, e03220. [Google Scholar] [CrossRef] [PubMed]
- Banarescu, P. Zoogeography of Freshwaters: Volume 1. General Distribution and Dispersal of Freshwater Animals; AULA-Verlag: Wiesbaden, Germany, 1990; ISBN 3-89104-480-1. [Google Scholar]
- Avise, J.C. Phylogeography: The History and Formation of Species; Harvard University Press: Cambridge, MA, USA; London, UK, 2000; ISBN 0674666380. [Google Scholar]
- McNeely, J.A. Biodiversity in arid regions: Values and perceptions. J. Arid. Environ. 2003, 54, 61–70. [Google Scholar] [CrossRef]
- Tourenq, C.; Launay, F. Challenges facing biodiversity in the United Arab Emirates. Manag. Environ. Quality 2008, 19, 283–304. [Google Scholar] [CrossRef]
- Garcia, N.; Harrison, I.J.; Cox, N.; Tognelli, M.F. (Eds.) The Status and Distribution of Freshwater Biodiversity in the Arabian Peninsula; IUCN Solprint: Mijas, Spain, 2015; ISBN 978-2-8317-1706-7. [Google Scholar]
- Dumont, H.J.; Negrea, S. Introduction to the class Branchiopoda. In Guides to the Identification of the Microinvertebrates of the Continental Waters of the World; Dumont, H.J., Ed.; Backhuys Publications: Leiden, Germany, 2002; ISBN 9057821125. [Google Scholar]
- Zawisza, E.; Zawiska, I.; Correa-Metrio, A. Cladocera Community Composition as a Function of Physicochemical and Morphological Parameters of Dystrophic Lakes in NE Poland. Wetlands 2016, 36, 1131–1142. [Google Scholar] [CrossRef] [Green Version]
- Ballinger, M.J.; Bruenn, J.A.; Kotov, A.A.; Taylor, D.J. Selectively maintained paleoviruses in Holarctic water fleas reveal an ancient origin for phleboviruses. Virology 2013, 446, 276–282. [Google Scholar] [CrossRef] [Green Version]
- Ebert, D. Genomics. A genome for the environment. Science 2011, 331, 539–540. [Google Scholar] [CrossRef]
- Ebert, D. Daphnia as a versatile model system in ecology and evolution. EvoDevo 2022, 13, 16. [Google Scholar] [CrossRef]
- Figuerola, J.; Green, A.J. Dispersal of aquatic organisms by waterbirds: A review of past research and priorities for future studies. Freshw. Biol. 2002, 47, 483–494. [Google Scholar] [CrossRef] [Green Version]
- Forro, L.; Korovchinsky, N.M.; Kotov, A.A.; Petrusek, A. Global diversity of cladocerans (Cladocera; Crustacea) in freshwater. Hydrobiologia 2008, 595, 177–184. [Google Scholar] [CrossRef]
- Dumont, H.J.; Maas, S.; Martens, K. Cladocera, Copepoda and Ostracoda (Crustacea) from fresh waters in South Yemen. Fauna of Saudi Arabia 1986, 8, 12–19. [Google Scholar]
- Dumont, H.J.; Brancelj, A. Alona alsafadii n. sp. from Yemen, a primitive, groundwater-dwelling member of the A. karua-group. Hydrobiologia 1994, 281, 57–64. [Google Scholar] [CrossRef]
- Schmarda, L.K. Zur Naturgeschichte Agyptens. Denksch. K. Akad. Wiss. Math. Nat. Cl. 1854, VII, 1–28. [Google Scholar]
- Blanchard, R.; Richard, J. Sur les crustaces des Sebkhaset des chotts d’Algerie. Bull. Soc. Zool. Fr. 1890, 15, 136–138. [Google Scholar]
- Ekman, S. Cladoceren und freilebende Copepoden aus Agypten und dem Sudan. In Results of the Swedish Zoological Expedition to Egypt and the White Nile, 1901, under the Direction of L.A. Jagerskiold; Jagerskiold, L.A., Ed.; Library of the Royal University of Uppsala: Uppsala, Sweden, 1904; pp. 1–18. [Google Scholar]
- Gurney, R. X—On the fresh-water Crustacea of Algeria and Tunisia. J. R. Micr. Soc. 1909, 29, 273–305. [Google Scholar] [CrossRef]
- Gauthier, H. Ostracodes et Cladocères de l’Afrique du Nord, deuxieme note. Bull. Soc. Hist. Nat. Afr. Nord. 1928, 19, 69–79. [Google Scholar]
- Harding, J.P. The Armstrong College zoological expedition to Siwa Oasis (Libyan Desert) 1935. Crustacea: Branchiopoda and Ostracoda. Proc. Egypt. Acad. Sci. 1955, 10, 58–68. [Google Scholar]
- Dumont, H.J.; Laureys, P.; Pensaert, J. Anostraca, Conchostraca, Cladocera and Copepoda from Tunisia. Hydrobiologia 1979, 66, 259–274. [Google Scholar] [CrossRef]
- Samraoui, B.; Segers, H.; Maas, S.; Baribwegure, D.; Dumont, H.J. Rotifera, Cladocera, Copepoda, and Ostracoda from coastal wetlands in northeast Algeria. Hydrobiologia 1998, 386, 183–193. [Google Scholar] [CrossRef]
- Marrone, F.; Korn, M.; Stoch, F.; Naselli-Flores, L.; Turki, S. Updated checklist and distribution of large branchiopods (Branchiopoda: Anostraca, Notostraca, Spinicaudata) in Tunisia. Biogeographia 2016, 27–53. [Google Scholar] [CrossRef] [Green Version]
- Ghaouaci, S.; Amarouayache, M.; Sinev, A.Y.; Korovchinsky, N.M.; Kotov, A.A. An annotated checklist of the Algerian Cladocera (Crustacea: Branchiopoda). Zootaxa 2018, 4377, 412–430. [Google Scholar] [CrossRef]
- Van Damme, K.; Dumont, H.J. A new species of Moina Baird, 1950 (Crustacea: Anomopoda) from Socotra Island, Yemen. Zootaxa 2008, 1721, 24. [Google Scholar] [CrossRef]
- Madera, P.; Van Damme, K. Socotra Archipelago (Yemen). In Imperiled: The Encyclopedia of Conservation; DellaSala, D.A., Goldstein, M.I., Eds.; Elsevier: Amsterdam, The Netherlands, 2022; pp. 267–281. ISBN 9780128211397. [Google Scholar]
- Abell, R.; Thieme, M.L.; Revenga, C.; Bryer, M.; Kottelat, M.; Bogutskaya, N.; Coad, B.; Mandrak, N.; Balderas, S.C.; Bussing, W.; et al. Freshwater ecoregions of the World: A new map of biogeographic units for freshwater biodiversity Conservation. Bioscience 2008, 58, 403–414. [Google Scholar] [CrossRef] [Green Version]
- FEOW. Freshwater Ecoregions of the World. Available online: https://www.feow.org/ (accessed on 7 July 2022).
- Dinerstein, E.; Olson, D.; Joshi, A.; Vynne, C.; Burgess, N.D.; Wikramanayake, E.; Hahn, N.; Palminteri, S.; Hedao, P.; Noss, R.; et al. An ecoregion-based approach to protecting half the terrestrial realm. Bioscience 2017, 67, 534–545. [Google Scholar] [CrossRef]
- Feulner, G.R. Wadi fish of the UAE. Tribulus 1998, 8, 16–22. [Google Scholar]
- Feulner, G.R.; Green, S.A. Freshwater snails of the UAE. Tribulus 1999, 9, 5–9. [Google Scholar]
- Saji, A.; Mischke, S.; Soorae, P.S.; Ahmed, S.; Al Dhaheri, S. The alwathba wetland reserve lake in Abu Dhabi, United Arab Emirates and its ostracod (seed shrimp) fauna. Int. J. Aquat. Biol. 2018, 6, 265–273. [Google Scholar] [CrossRef]
- Saji, A.; Al Dhaheri, S.; Shah, J.N.; Soorae, P.S. Influence of chemical parameters on Artemia sp. (Crustacea: Anostraca) population in Al Wathba Lake in the Abu Dhabi Emirate, UAE. Int. J. Aquat. Biol. 2016, 4, 87–95. [Google Scholar] [CrossRef]
- Asem, A.; Schuster, R.; Eimanifar, A.; Lu, H.; Liu, C.; Wu, X.; Yao, L.; Meng, X.; Li, W.; Wang, P. Impact of colonization of an invasive species on genetic differentiation in new environments: A study on American Artemia franciscana (Crustacea: Anostraca) in the United Arab Emirates. J. Ocean Univ. China 2021, 20, 911–920. [Google Scholar] [CrossRef]
- Soorae, P.; Javed, S.; Dhaheri, S.A.; Qassimi, M.A.; Kabshawi, M.; Saji, A.; Khan, S.; Sakkir, S.; Zaabi, R.A.; Ahmed, S.; et al. Alien species recorded in the United Arab Emirates: An initial list of terrestrial and freshwater species. J. Threat. Taxa 2015, 7, 7910–7921. [Google Scholar] [CrossRef]
- Hamza, W.; Ramadan, G.; AlKaabi, M. Morphological and molecular identification of first recorded Cladoceran organisms in the desert of Abu Dhabi, UAE. MOJ Eco. Environ. Sci. 2018, 3, 220–224. [Google Scholar] [CrossRef] [Green Version]
- Soesbergen, M. A preliminary investigation of plankton organisms of fresh and brackish inland waters in the northern United Arab Emirates. Tribulus 2018, 26, 46–58. [Google Scholar]
- Van Damme, K.; Sinev, A.Y.; Dumont, H.J. Separation of Anthalona gen.n. from Alona Baird, 1843 (Branchiopoda: Cladocera: Anomopoda): Morphology and evolution of scraping stenothermic alonines. Zootaxa 2011, 2875, 1–64. [Google Scholar] [CrossRef]
- Hamza, W.; Neretina, A.N.; Al Neyadi, S.E.S.; Amiri, K.; Karabanov, D.P.; Kotov, A.A. Discovery of a New Species of Daphnia (Crustacea: Cladocera) from the Arabian Peninsula Revealed a Southern Origin of a Common Northern Eurasian Species Group. Water 2022, 14, 2350. [Google Scholar] [CrossRef]
- Sayers, E.W.; Cavanaugh, M.; Clark, K.; Ostell, J.; Pruitt, K.D.; Karsch-Mizrachi, I. GenBank. Nucleic Acids Res. 2019, 47, D94–D99. [Google Scholar] [CrossRef] [Green Version]
- Ratnasingham, S.; Hebert, P.D.N. BOLD: The Barcode of Life Data System. Mol. Ecol. Notes 2007, 7, 355–364. [Google Scholar] [CrossRef] [Green Version]
- Van Damme, K.; Dumont, H.J. Further division of Alona Baird, 1843: Separation and position of Coronatella Dybowski et Grochowski and Ovalona gen.n. (Crustacea: Cladocera). Zootaxa 2008, 1960, 1–44. [Google Scholar] [CrossRef]
- Popova, E.V.; Petrusek, A.; Korinek, V.; Mergeay, J.; Bekker, E.I.; Karabanov, D.P.; Galimov, Y.R.; Neretina, T.V.; Taylor, D.J.; Kotov, A.A. Revision of the Old World Daphnia (Ctenodaphnia) similis group (Cladocera: Daphniidae). Zootaxa 2016, 4161, 1–40. [Google Scholar] [CrossRef]
- Korovchinsky, N.M.; Kotov, A.A.; Sinev, A.Y.; Neretina, A.N.; Garibian, P.G. Water Fleas (Crustacea: Cladocera) of Northern Eurasi; KMK Scientific Press Ltd.: Moscow, Russia, 2021; Volume 2, ISBN 978-5-907372-50-4. [Google Scholar]
- Kotov, A.A. Morphology and Phylogeny of the Anomopoda (Crustacea: Cladocera); KMK Scientific Press Ltd.: Moscow, Russia, 2013; ISBN 9785873179237. [Google Scholar]
- Hebert, P.D.N.; Cywinska, A.; Ball, S.L.; deWaard, J.R. Biological identifications through DNA barcodes. Proc. R. Soc. Lond. B Biol. Sci. 2003, 270, 313–321. [Google Scholar] [CrossRef] [Green Version]
- Neretina, A.N.; Karabanov, D.P.; Sacherova, V.; Kotov, A.A. Unexpected mitochondrial lineage diversity within the genus Alonella Sars, 1862 (Crustacea: Cladocera) across the Northern Hemisphere. PeerJ 2021, 9, e10804. [Google Scholar] [CrossRef]
- Okonechnikov, K.; Golosova, O.; Fursov, M. Unipro UGENE: A unified bioinformatics toolkit. Bioinformatics 2012, 28, 1166–1167. [Google Scholar] [CrossRef] [Green Version]
- Sayers, E.W.; Beck, J.; Brister, J.R.; Bolton, E.E.; Canese, K.; Comeau, D.C.; Funk, K.; Ketter, A.; Kim, S.; Kimchi, A.; et al. Database resources of the National Center for Biotechnology Information. Nucleic Acids Res. 2020, 48, D9–D16. [Google Scholar] [CrossRef] [Green Version]
- Katoh, K.; Standley, D.M. MAFFT multiple sequence alignment software version 7: Improvements in performance and usability. Mol. Biol. Evol. 2013, 30, 772–780. [Google Scholar] [CrossRef] [Green Version]
- Kalyaanamoorthy, S.; Minh, B.Q.; Wong, T.K.F.; von Haeseler, A.; Jermiin, L.S. ModelFinder: Fast model selection for accurate phylogenetic estimates. Nat. Methods 2017, 14, 587–589. [Google Scholar] [CrossRef] [Green Version]
- Trifinopoulos, J.; Nguyen, L.-T.; von Haeseler, A.; Minh, B.Q. W-IQ-TREE: A fast online phylogenetic tool for maximum likelihood analysis. Nucleic Acids Res. 2016, 44, W232–W235. [Google Scholar] [CrossRef] [Green Version]
- Posada, D.; Buckley, T.R. Model selection and model averaging in phylogenetics: Advantages of Akaike information criterion and Bayesian approaches over likelihood ratio tests. Syst. Biol. 2004, 53, 793–808. [Google Scholar] [CrossRef] [PubMed]
- Yang, Z.; Rannala, B. Molecular phylogenetics: Principles and practice. Nat. Rev. Genet. 2012, 13, 303–314. [Google Scholar] [CrossRef] [PubMed]
- Vaidya, G.; Lohman, D.J.; Meier, R. SequenceMatrix: Concatenation software for the fast assembly of multi-gene datasets with character set and codon information. Cladistics 2011, 27, 171–180. [Google Scholar] [CrossRef] [PubMed]
- Minh, B.Q.; Schmidt, H.A.; Chernomor, O.; Schrempf, D.; Woodhams, M.D.; von Haeseler, A.; Lanfear, R. IQ-TREE 2: New models and efficient methods for phylogenetic inference in the genomic era. Mol. Biol. Evol. 2020, 37, 1530–1534. [Google Scholar] [CrossRef] [Green Version]
- Hoang, D.T.; Chernomor, O.; von Haeseler, A.; Minh, B.Q.; Le Vinh, S. UFBoot2: Improving the ultrafast bootstrap approximation. Mol. Biol. Evol. 2018, 35, 518–522. [Google Scholar] [CrossRef]
- Guindon, S.; Dufayard, J.-F.; Lefort, V.; Anisimova, M.; Hordijk, W.; Gascuel, O. New algorithms and methods to estimate maximum-likelihood phylogenies: Assessing the performance of PhyML 3.0. Syst. Biol. 2010, 59, 307–321. [Google Scholar] [CrossRef] [Green Version]
- Goloboff, P.A.; Catalano, S.A. TNT version 1.5, including a full implementation of phylogenetic morphometrics. Cladistics 2016, 32, 221–238. [Google Scholar] [CrossRef]
- Heled, J.; Drummond, A.J. Bayesian inference of species trees from multilocus data. Mol. Biol. Evol. 2010, 27, 570–580. [Google Scholar] [CrossRef] [Green Version]
- Bouckaert, R.; Vaughan, T.G.; Barido-Sottani, J.; Duchene, S.; Fourment, M.; Gavryushkina, A.; Heled, J.; Jones, G.; Kuhnert, D.; de Maio, N.; et al. BEAST 2.5: An advanced software platform for Bayesian evolutionary analysis. PLoS Comput. Biol. 2019, 15, e1006650. [Google Scholar] [CrossRef] [Green Version]
- Drummond, A.J.; Suchard, M.A.; Xie, D.; Rambaut, A. Bayesian phylogenetics with BEAUti and the BEAST 1.7. Mol. Biol. Evol. 2012, 29, 1969–1973. [Google Scholar] [CrossRef] [Green Version]
- Cornetti, L.; Fields, P.D.; Van Damme, K.; Ebert, D. A fossil-calibrated phylogenomic analysis of Daphnia and the Daphniidae. Mol. Phylogenet. Evol. 2019, 137, 250–262. [Google Scholar] [CrossRef]
- Rambaut, A.; Drummond, A.J.; Xie, D.; Baele, G.; Suchard, M.A. Posterior summarization in Bayesian phylogenetics using Tracer 1.7. Syst. Biol. 2018, 67, 901–904. [Google Scholar] [CrossRef] [Green Version]
- Drummond, A.J.; Bouckaert, R.R. Bayesian Evolutionary Analysis with BEAST2; Cambridge University Press: Cambridge, UK, 2015; ISBN 978-1-107-01965-2. [Google Scholar]
- Parham, J.F.; Donoghue, P.C.J.; Bell, C.J.; Calway, T.D.; Head, J.J.; Holroyd, P.A.; Inoue, J.G.; Irmis, R.B.; Joyce, W.G.; Ksepka, D.T.; et al. Best practices for justifying fossil calibrations. Syst. Biol. 2012, 61, 346–359. [Google Scholar] [CrossRef]
- Douglas, J.; Zhang, R.; Bouckaert, R. Adaptive dating and fast proposals: Revisiting the phylogenetic relaxed clock model. PLoS Comput. Biol. 2021, 17, e1008322. [Google Scholar] [CrossRef]
- Gernhard, T. The conditioned reconstructed process. J. Theor. Biol. 2008, 253, 769–778. [Google Scholar] [CrossRef] [Green Version]
- Barido-Sottani, J.; Boskova, V.; Du Plessis, L.; Kuhnert, D.; Magnus, C.; Mitov, V.; Muller, N.F.; PecErska, J.; Rasmussen, D.A.; Zhang, C.; et al. Taming the BEAST—A community teaching material resource for BEAST2. Syst. Biol. 2018, 67, 170–174. [Google Scholar] [CrossRef] [PubMed]
- Puillandre, N.; Brouillet, S.; Achaz, G. ASAP: Assemble species by automatic partitioning. Mol. Ecol. Resour. 2021, 21, 609–620. [Google Scholar] [CrossRef] [PubMed]
- Pons, J.; Barraclough, T.G.; Gomez-Zurita, J.; Cardoso, A.; Duran, D.P.; Hazell, S.; Kamoun, S.; Sumlin, W.D.; Vogler, A.P. Sequence-based species delimitation for the DNA taxonomy of undescribed insects. Syst. Biol. 2006, 55, 595–609. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Reid, N.M.; Carstens, B.C. Phylogenetic estimation error can decrease the accuracy of species delimitation: A Bayesian implementation of the general mixed Yule-coalescent model. BMC Evol. Biol. 2012, 12, 196. [Google Scholar] [CrossRef] [Green Version]
- Microsoft R Open Application Network; Microsoft, R. Core Team: Redmond, DC, USA, 2014.
- Carstens, B.C.; Pelletier, T.A.; Reid, N.M.; Satler, J.D. How to fail at species delimitation. Mol. Ecol. 2013, 22, 4369–4383. [Google Scholar] [CrossRef]
- Kapli, P.; Lutteropp, S.; Zhang, J.; Kobert, K.; Pavlidis, P.; Stamatakis, A.; Flouri, T. Multi-rate Poisson tree processes for single-locus species delimitation under maximum likelihood and Markov chain Monte Carlo. Bioinformatics 2017, 33, 1630–1638. [Google Scholar] [CrossRef] [Green Version]
- Alonso, M.; Neretina, A.N.; Ventura, M. Ceriodaphnia smirnovi (Crustacea: Cladocera), a new species from the Mediterranean region, and a phylogenetic analysis of the commonest species. Zootaxa 2021, 4974, 146. [Google Scholar] [CrossRef]
- Smirnov, N.N. Macrothricidae i Moinidae fauni mira. Fauna SSSR. Rakoobraznye. [Macrothricidae and Moinidae of the World’s fauna. Fauna of the USSR. Crustacea]; Nauka: Leningrad, Russia, 1976. [Google Scholar]
- Sars, G.O. Contributions to the knowledge of the fresh-water Entomostraca of South America, as shown by artificial hatching from dried material. Arch. Math. Naturvid. B 1901, XXIII, 1–102. [Google Scholar]
- Van Damme, K.; Dumont, H.J. Cladocera of the Lencois Maranhenses (NE—Brazil): Faunal composition and a reappraisal of Sars’ Method. Braz. J. Biol. 2010, 70, 755–779. [Google Scholar] [CrossRef]
- Diniz, L.P.; Melo-Junior, M.D. Can nearby eutrophic reservoirs sustain a differentiated biodiversity of planktonic microcrustaceans in a tropical semiarid basin? An. Acad. Bras. Cienc. 2017, 89, 2771–2783. [Google Scholar] [CrossRef] [Green Version]
- Mallin, M.A.; Partin, W.E. Thermal tolerances of common Cladocera. J. Freshw. Ecol. 1989, 5, 45–51. [Google Scholar] [CrossRef]
- Smirnov, N.N. Physiology of the Cladocera, 2nd ed.; Academic Press: Amsterdam, The Netherlands, 2017; ISBN 9780128051948. [Google Scholar]
- Maksimova, L.P. Instructions on cultivation of Moina macrocopa Straus; GosNIORKh: Leningrad, Russia, 1969. [Google Scholar]
- Korovchinsky, N.M. Cladocerans of the Order Ctenopoda of the World Fauna (Morphology, Systematics, Ecology, Biogeography); KMK Press: Moscow, Russia, 2004. [Google Scholar]
- Brown, L.A. The natural history of Cladocerans in relation to temperature. I. Distribution and the temperature limits for vital activities. Am. Nat. 1929, 63, 248–264. [Google Scholar] [CrossRef]
- Korovchinsky, N.M.; Kotov, A.A.; Boikova, O.S.; Smirnov, N.N. Water Fleas (Crustacea: Cladocera) of Northern Eurasia; KMK Scientific Press Ltd.: Moscow, Russia, 2021; Volume 1, ISBN 978-5-907372-27-6. [Google Scholar]
- Berner, D.B. Morphological differentiation among species in the Ceriodaphnia cornuta complex (Crustacea, Cladocera). Verh. Internat. Verein. Limnol. 1985, 22, 3099–3103. [Google Scholar] [CrossRef]
- Adamowicz, S.J.; Petrusek, A.; Colbourne, J.K.; Hebert, P.D.N.; Witt, J.D.S. The scale of divergence: A phylogenetic appraisal of intercontinental allopatric speciation in a passively dispersed freshwater zooplankton genus. Mol. Phylogenet. Evol. 2009, 50, 423–436. [Google Scholar] [CrossRef] [PubMed]
- Bekker, E.I.; Karabanov, D.P.; Galimov, Y.R.; Haag, C.R.; Neretina, T.V.; Kotov, A.A. Phylogeography of Daphnia magna Straus (Crustacea: Cladocera) in Northern Eurasia: Evidence for a deep longitudinal split between mitochondrial lineages. PLoS ONE 2018, 13, e0194045. [Google Scholar] [CrossRef]
- Juracka, P.J.; Laforsch, C.; Petrusek, A. Neckteeth formation in two species of the Daphnia curvirostris complex (Crustacea: Cladocera). J. Limnol. 2011, 70, 359–368. [Google Scholar] [CrossRef]
- Zuykova, E.I.; Simonov, E.P.; Bochkarev, N.A.; Taylor, D.J.; Kotov, A.A. Resolution of the Daphnia umbra problem (Crustacea: Cladocera) using an integrated taxonomic approach. Zool. J. Linn. Soc. 2018, 50, 969–998. [Google Scholar] [CrossRef]
- Kotov, A.A.; Garibian, P.G.; Bekker, E.I.; Taylor, D.J.; Karabanov, D.P. A new species group from the Daphnia curvirostris species complex (Cladocera: Anomopoda) from the eastern Palaearctic: Taxonomy, phylogeny and phylogeography. Zool. J. Linn. Soc. 2021, 191, 772–822. [Google Scholar] [CrossRef]
- Gu, Y.-L.; Sun, C.-H.; Liu, P.; Zhang, X.; Sinev, A.Y.; Dumont, H.J.; Han, B.-P. Complete mitochondrial genome of Ovalona pulchella (Branchiopoda, Cladocera) as the first representative in the family Chydoridae: Gene rearrangements and phylogenetic analysis of Cladocera. Gene 2022, 818, 146230. [Google Scholar] [CrossRef]
- Sinev, A.Y.; Karabanov, D.P.; Kotov, A.A. A new North Eurasian species of the Alona affinis complex (Cladocera: Chydoridae). Zootaxa 2020, 4767, 115–137. [Google Scholar] [CrossRef]
- Elias-Gutierrez, M.; Valdez–Moreno, M. A new cryptic species of Leberis Smirnov, 1989 (Crustacea, Cladocera, Chydoridae) from the Mexican semi-desert region, highlighted by DNA barcoding. Hidrobiologica 2008, 18, 63–74. [Google Scholar]
- Bekker, E.I.; Karabanov, D.P.; Galimov, Y.R.; Kotov, A.A. DNA barcoding reveals high cryptic diversity in the North Eurasian Moina species (Crustacea: Cladocera). PLoS ONE 2016, 11, e0161737. [Google Scholar] [CrossRef] [Green Version]
- Sousa, F.D.R.; Elmoor-Loureiro, L.M.A.; Santos, S. Redescription of Coronatella poppei (Richard, 1897) (Crustacea, Branchiopoda, Chydoridae) and a revision of the genus in Brazil, with descriptions of new taxa. Zootaxa 2015, 3955, 211–244. [Google Scholar] [CrossRef]
- Sousa, F.D.R.; Elmoor-Loureiro, L.M.A.; Santos, S. Position of the dentifera-group in the Coronatella-branch and its relocation to a new genus: Magnospina gen. n. (Crustacea, Chydoridae, Aloninae). Zookeys 2016, 586, 95–119. [Google Scholar] [CrossRef] [Green Version]
- Sinev, A.Y. Revision of the elegans-group of Alona s. lato and its status as a subgenus of Coronatella Dybowski et Grochowski, 1894 (Cladocera: Anomopoda: Chydoridae). Zootaxa 2020, 4732, 501–526. [Google Scholar] [CrossRef]
- Neretina, A.N.; Kirdyasheva, A.G. A redescription of Moina australiensis Sars, 1896 (Cladocera: Moinidae) with short notes on Australian moinids. Zootaxa 2019, 4577, zootaxa.4577.1.10. [Google Scholar] [CrossRef]
- Alonso, M.; Neretina, A.N.; Sanoamuang, L.-O.; Saengphan, N.; Kotov, A.A. A new species of Moina Baird, 1850 (Cladocera: Moinidae) from Thailand. Zootaxa 2019, 4554, 199–218. [Google Scholar] [CrossRef]
- Sharma, P.; Kotov, A.A. Molecular approach to identify sibling species of the Ceriodaphnia cornuta complex (Cladocera: Daphniidae) from Australia with notes on the continental endemism of this group. Zootaxa 2013, 3702, 79–89. [Google Scholar] [CrossRef] [Green Version]
- Elias-Gutierrez, M.; Juracka, P.J.; Montoliu-Elena, L.; Miracle, M.R.; Petrusek, A.; Korinek, V. Who is Moina micrura? Redescription of one of the most confusing cladocerans from terra typica, based on integrative taxonomy. Limnetica 2019, 38, 227–252. [Google Scholar] [CrossRef]
- Montoliu-Elena, L.; Elias-Gutierrez, M.; Silva-Briano, M. Moina macrocopa (Straus, 1820): A species complex of a common Cladocera, highlighted by morphology and DNA barcodes. Limnetica 2019, 1, 253–277. [Google Scholar] [CrossRef]
- Ni, Y.; Ma, X.; Hu, W.; Blair, D.; Yin, M. New lineages and old species: Lineage diversity and regional distribution of Moina (Crustacea: Cladocera) in China. Mol. Phylogenet. Evol. 2019, 134, 87–98. [Google Scholar] [CrossRef] [PubMed]
- Luo, A.; Ling, C.; Ho, S.Y.W.; Zhu, C.-D. Comparison of methods for molecular species delimitation across a range of speciation scenarios. Syst. Biol. 2018, 67, 830–846. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sacherova, V.; Hebert, P.D.N. The evolutionary history of the Chydoridae (Crustacea: Cladocera). Biol. J. Linn. Soc. 2003, 79, 629–643. [Google Scholar] [CrossRef]
- Van Damme, K.; Kotov, A.A. The fossil record of the Cladocera (Crustacea: Branchiopoda): Evidence and hypotheses. Earth. Sci. Rev. 2016, 163, 162–189. [Google Scholar] [CrossRef]
- Hegna, T.A.; Kotov, A.A. Ephippia belonging to Ceriodaphnia Dana, 1853 (Cladocera: Anomopoda: Daphniidae) from the Lower Cretaceous of Australia. Palaeontol. Electron. 2016, 19, 1–9. [Google Scholar] [CrossRef]
- Kotov, A.A.; Karabanov, D.P.; Bekker, E.I.; Neretina, T.V.; Taylor, D.J. Phylogeography of the Chydorus sphaericus Group (Cladocera: Chydoridae) in the Northern Palearctic. PLoS ONE 2016, 11, e0168711. [Google Scholar] [CrossRef] [Green Version]
- Zuykova, E.I.; Simonov, E.P.; Bochkarev, N.A.; Abramov, S.A.; Sheveleva, N.G.; Kotov, A.A. Contrasting phylogeographic patterns and demographic history in closely related species of Daphnia longispina group (Crustacea: Cladocera) with focus on North-Eastern Eurasia. PLoS ONE 2018, 13, e0207347. [Google Scholar] [CrossRef] [Green Version]
- Grandcolas, P.; Nattier, R.; Trewick, S. Relict species: A relict concept? Trends Ecol. Evol. 2014, 29, 655–663. [Google Scholar] [CrossRef]
- Garibian, P.G.; Karabanov, D.P.; Neretina, A.N.; Taylor, D.J.; Kotov, A.A. Bosminopsis deitersi (Crustacea: Cladocera) as an ancient species group: A revision. PeerJ 2021, 9, e11310. [Google Scholar] [CrossRef]
- Zanazzi, A.; Kohn, M.J.; MacFadden, B.J.; Terry, D.O. Large temperature drop across the Eocene-Oligocene transition in central North America. Nature 2007, 445, 639–642. [Google Scholar] [CrossRef]
- Pekar, S.F.; Christie-Blick, N. Resolving apparent conflicts between oceanographic and Antarctic climate records and evidence for a decrease in pCO2 during the Oligocene through early Miocene (34–16 Ma). Palaeogeogr. Palaeoclimatol. Palaeoecol. 2008, 260, 41–49. [Google Scholar] [CrossRef] [Green Version]
- Li, S.; Xing, Y.; Valdes, P.J.; Huang, Y.; Su, T.; Farnsworth, A.; Lunt, D.J.; Tang, H.; Kennedy, A.T.; Zhou, Z. Oligocene climate signals and forcings in Eurasia revealed by plant macrofossil and modelling results. Gondwana Res. 2018, 61, 115–127. [Google Scholar] [CrossRef] [Green Version]
- Korovchinsky, N.M. The Cladocera (Crustacea: Branchiopoda) as a relict group. Zool. J. Linn. Soc. 2006, 147, 109–124. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Z.; Ramstein, G.; Schuster, M.; Li, C.; Contoux, C.; Yan, Q. Aridification of the Sahara desert caused by Tethys Sea shrinkage during the Late Miocene. Nature 2014, 513, 401–404. [Google Scholar] [CrossRef]
- Hoelzmann, P.; Gasse, F.; Dupont, L.M.; Salzmann, U.; Staubwasser, M.; Leuschner, D.C.; Sirocko, F. Palaeoenvironmental changes in the arid and sub arid belt (Sahara-Sahel-Arabian Peninsula) from 150 kyr to present. In Past Climate Variability through Europe and Africa; Smol, J.P., Last, W.M., Battarbee, R.W., Gasse, F., Stickley, C.E., Eds.; Springer Netherlands & Dordrecht: Dordrecht, The Netherlands, 2004; pp. 219–256. ISBN 978-1-4020-2120-6. [Google Scholar]
- Groucutt, H.S.; Petraglia, M.D. The prehistory of the Arabian peninsula: Deserts, dispersals, and demography. Evol. Anthropol. 2012, 21, 113–125. [Google Scholar] [CrossRef]
- Keiser, N. Uber die Cladoceren und Copepoden der Wuste Kara-Kum. Int. Revue ges. Hydrobiol. Hydrogr. 1931, 25, 355–372. [Google Scholar] [CrossRef]
- Gauthier, H. Euphyllopodes et Cladoceres continentaux recoltes par M. Monod au Sahara occidental et en Mauritanie. Bull. Soc. Sci. Nat. Phys. Maroc 1937, 17, 75–98. [Google Scholar]
- Zharov, A.A.; Neretina, A.N.; Rogers, D.C.; Reshetova, S.A.; Sinitsa, S.M.; Kotov, A.A. Pleistocene Branchiopods (Cladocera, Anostraca) from Transbaikalian Siberia demonstrate morphological and ecological stasis. Water 2020, 12, 3063. [Google Scholar] [CrossRef]
- Neretina, A.N.; Gololobova, M.A.; Neplyukhina, A.A.; Zharov, A.A.; Rogers, C.D.; Horne, D.J.; Protopopov, A.V.; Kotov, A.A. Crustacean remains from the Yuka mammoth raise questions about non-analogue freshwater communities in the Beringian region during the Pleistocene. Sci. Rep. 2020, 10, 859. [Google Scholar] [CrossRef] [Green Version]
- Dumont, H.J. Relict distribution patterns of aquatic animals: Another tool in evaluating Late Pleistocene climate changes in the Sahara and Sahel. In Palaeoecology of Africa and the surrounding islands, 1st ed.; Routledge: Milton Park, UK, 1982; Volume 14, ISBN 9780203744529. [Google Scholar]























{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| № | Emirate | Locality | N | E | C. c. | D. a. | M. m. | A. m. | C. a. |
|---|---|---|---|---|---|---|---|---|---|
| 1 | Ajman | Kholaiban Dam Masfut 1 | 24.81019 | 56.09278 | 1 | 0 | 1 | 1 | 1 |
| 2 | Ajman | Kholaiban Dam Masfut 2 | 24.80982 | 56.09306 | 0 | 0 | 1 | 0 | 0 |
| 3 | Fujairah | Wadi Maidaq—Blue Pool | 25.35205 | 56.08864 | 0 | 0 | 1 | 1 | 1 |
| 4 | Abu Dhabi | Al Shuwaib Dam, near Al Ain city | 24.77174 | 55.80146 | 0 | 1 | 0 | 0 | 0 |
| 5 | Dubai | Hatta Dam | 24.7835 | 56.11186 | 0 | 0 | 0 | 0 | 0 |
| 6 | Dubai | Sheikh Maktoum Bin Rashid Al Maktoum Dam | 24.81213 | 56.1469 | 0 | 0 | 0 | 0 | 0 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kotov, A.A.; Neretina, A.N.; Al Neyadi, S.E.S.; Karabanov, D.P.; Hamza, W. Cladocera (Crustacea: Branchiopoda) of Man-Made Lakes at the Northeast Part of the United Arab Emirates with a Hypothesis on Their Origin. Diversity 2022, 14, 688. https://doi.org/10.3390/d14080688
Kotov AA, Neretina AN, Al Neyadi SES, Karabanov DP, Hamza W. Cladocera (Crustacea: Branchiopoda) of Man-Made Lakes at the Northeast Part of the United Arab Emirates with a Hypothesis on Their Origin. Diversity. 2022; 14(8):688. https://doi.org/10.3390/d14080688
Chicago/Turabian StyleKotov, Alexey A., Anna N. Neretina, Shamma Eisa Salem Al Neyadi, Dmitry P. Karabanov, and Waleed Hamza. 2022. "Cladocera (Crustacea: Branchiopoda) of Man-Made Lakes at the Northeast Part of the United Arab Emirates with a Hypothesis on Their Origin" Diversity 14, no. 8: 688. https://doi.org/10.3390/d14080688