
Applicable Life-History and Molecular Traits for Studying the Effects of Anhydrobiosis on Aging in Tardigrades



Abstract
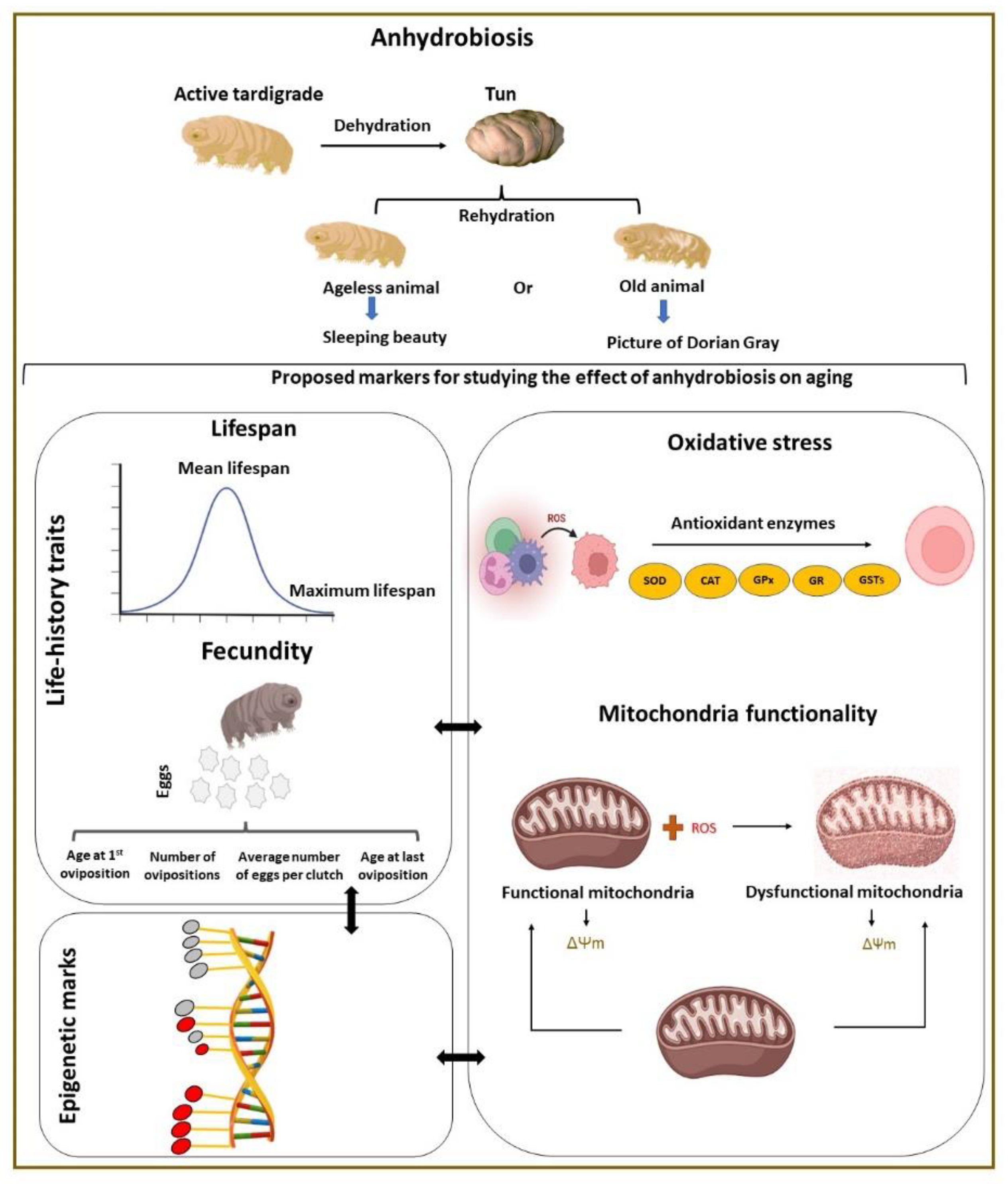
:1. Introduction
2. Anhydrobiosis
3. Aging
3.1. Levels of Research
3.2. Proposed Mechanism
4. Markers in Research on the Effect of Anhydrobiosis on Aging
4.1. Life-History Traits
4.1.1. Lifespan
4.1.2. Fecundity
4.2. Possible Cellular Markers
4.2.1. Oxidative Stress
ROS Generation and Oxidative Modifications of Different Molecules
ROS-Scavenging Enzymes
Mitochondria Functioning
4.2.2. Epigenetic Modifications
Epigenetics and Stress Conditions
Possible Epigenetic Markers
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Kaczmarek, Ł. Tardigrada: An emerging animal model to study the Endoplasmic Reticulum stress response to environmental extremes. In Cellular Biology of the Endoplasmic Reticulum. Progress in Molecular and Subcellular Biology; Agellon, L.B., Michalak, M., Eds.; Springer: Cham, Switzerland, 2021; Volume 59, pp. 305–327. [Google Scholar] [CrossRef]
- Guidetti, R.; McInnes, S.J.; Kristensen, R.M. World List of Tardigrada; World Register of Marine Species: Ostend, Belgium, 2022; Available online: https://www.marinespecies.org/tardigrada (accessed on 15 August 2022). [CrossRef]
- Degma, P.; Bertolani, R.; Guidetti, R. Actual Checklist of Tardigrada Species, 41th ed; University of Modena and Reggio Emilia: Modena, Italy, 2022; Available online: https://iris.unimore.it/retrieve/e31e1250-6907-987f-e053-3705fe0a095a/Actual%20checklist%20of%20Tardigrada%2041th%20Edition%2016-05-22.pdf (accessed on 21 July 2022). [CrossRef]
- Fleming, J.F.; Arakawa, K. Systematics of Tardigrada: A reanalysis of tardigrade taxonomy with specific reference to Guil et al. (2019). Zool. Scr. 2021, 50, 376–382. [Google Scholar] [CrossRef]
- Bertolani, R. Evolution of the reproductive mechanisms in tardigrades—A review. Zool. Anz. 2001, 240, 247–252. [Google Scholar] [CrossRef]
- Lemloh, M.; Brümmer, F.; Schill, R.O. Life-history traits of the bisexual tardigrades Paramacrobiotus tonollii and Macrobiotus sapiens. J. Zoolog. Syst. Evol. Res. 2011, 49, 58–61. [Google Scholar] [CrossRef]
- Nelson, D.R.; Guidetti, R.; Rebecchi, L. Phylum Tardigrada. In Thorp and Covich’s Freshwater Invertebrates; James, H., Thorp, D., Rogers, C., Eds.; Academic Press: London, UK, 2015; pp. 347–380. [Google Scholar] [CrossRef]
- Schill, R.O.; Jönsson, K.I.; Pfannkuchen, M.; Brümmer, F. Food of tardigrades: A case study to understand food choice, intake and digestion. J. Zoolog. Syst. Evol. Res. 2011, 49, 66–70. [Google Scholar] [CrossRef]
- Roszkowska, M.; Wojciechowska, D.; Kmita, H.; Cerbin, S.; Dziuba, M.K.; Fiałkowska, E.; Sobkowiak, R.; Szydło, W.; Kaczmarek, Ł. Tips and tricks how to culture water bears: Simple protocols for culturing eutardigrades (Tardigrada) under laboratory conditions. Eur. Zool. J. 2021, 88, 449–465. [Google Scholar] [CrossRef]
- Roszkowska, M.; Bartels, P.J.; Gołdyn, B.; Ciobanu, D.A.; Fontoura, P.; Michalczyk, Ł.; Nelson, D.R.; Ostrowska, M.; Moreno-Talamantes, A.; Kaczmarek, Ł. Is the gut content of Milnesium (Eutardigrada) related to buccal tube size? Zool. J. Linn. Soc. 2016, 178, 794–803. [Google Scholar] [CrossRef]
- Jönsson, K.I. Tardigrades—Evolutionary explorers in extreme environments. In Extremophiles as Astrobiological Models; Seckbach, J., Stan-Lotter, H., Eds.; Wiley: Beverly, MA, USA, 2020; pp. 255–274. [Google Scholar] [CrossRef]
- Keilin, D. The problem of anabiosis or latent life: History and current concept. Proc. R. Soc. Lond. B Biol. Sci. 1959, 150, 149–191. [Google Scholar] [CrossRef]
- Schill, R.O.; Hengherr, S. Environmental adaptations: Desiccation tolerance. In Water Bears: The Biology of Tardigrades; Schill, R.O., Ed.; Springer: Cham, Switzerland, 2018; Volume 2, pp. 273–293. [Google Scholar] [CrossRef]
- Hengherr, S.; Schill, R.O. Environmental adaptations: Cryobiosis. In Water Bears: The Biology of Tardigrades; Schill, R.O., Ed.; Springer: Cham, Switzerland, 2018; Volume 2, pp. 295–310. [Google Scholar] [CrossRef]
- Clegg, J.S. Cryptobiosis—A peculiar state of biological organization. Comp. Biochem. Physiol. B Biochem. 2001, 128, 613–624. [Google Scholar] [CrossRef]
- Møbjerg, N.; Neves, R.C. New insights into survival strategies of tardigrades. Comp. Biochem. Physiol. A Mol. Integr. Physiol. 2021, 254, 110890. [Google Scholar] [CrossRef]
- Hengherr, S.; Brümmer, F.; Schill, R.O. Anhydrobiosis in tardigrades and its effects on longevity traits. J. Zool. 2008, 275, 216–220. [Google Scholar] [CrossRef]
- Tsujimoto, M.; Imura, S.; Kanda, H. Recovery and reproduction of an Antarctic tardigrade retrieved from a moss sample frozen for over 30 years. Cryobiology 2016, 72, 78–81. [Google Scholar] [CrossRef]
- Kaczmarek, Ł.; Roszkowska, M.; Fontaneto, D.; Jezierska, M.; Pietrzak, B.; Wieczorek, R.; Poprawa, I.; Kosicki, J.Z.; Karachitos, A.; Kmita, H. Staying young and fit? Ontogenetic and phylogenetic consequences of animal anhydrobiosis. J. Zool. 2019, 309, 1–11. [Google Scholar] [CrossRef]
- Ricci, C.; Pagani, M. Desiccation of Panagrolaimus rigidus (Nematoda): Survival, reproduction and the influence on the internal clock. Hydrobiologia 1997, 347, 1–13. [Google Scholar] [CrossRef]
- Ricci, C.; Covino, C. Anhydrobiosis of Adineta ricciae: Costs and benefits. In Developments in Hydrobiology; Herzig, A., Gulati, R.D., Jersabek, C.D., May, L., Eds.; Springer: Dordrecht, The Netherlands, 2005; Volume 181, pp. 307–314. [Google Scholar] [CrossRef]
- Ricci, C.; Vaghi, L.; Manzini, M.L. Desiccation of rotifers (Macrotrachela Quadricornifera): Survival and reproduction. Ecology 1987, 68, 1488–1494. [Google Scholar] [CrossRef]
- Clegg, J.S. Do dried cryptobiotes have a metabolism? In Anhydrobiosis; Crowe, J.H., Clegg, J.S., Eds.; Dowden Hutchinson and Ross: Stroudsburg, PA, USA, 1973; pp. 141–147. [Google Scholar]
- Pigoń, A.; Weglarska, B. Rate of metabolism in tardigrades during active life and anabiosis. Nature 1955, 176, 121–122. [Google Scholar] [CrossRef]
- Barrett, J. Metabolic responses to anabiosis in the fourth stage juveniles of Ditylenchus dipsaci (Nematoda). Proc. R. Soc. Lond. B Biol. Sci. 1982, 216, 159–177. [Google Scholar] [CrossRef]
- Wharton, D.A.; Barrett, J.; Perry, R.N. Water uptake and morphological changes during recovery from anabiosis in the plant-parasitic nematode, Ditylenchus dipsaci. J. Zool. 1985, 206, 391–402. [Google Scholar] [CrossRef]
- Wojciechowska, D.; Karachitos, A.; Roszkowska, M.; Rzeźniczak, W.; Sobkowiak, R.; Kaczmarek, Ł.; Kosicki, J.Z.; Kmita, H. Mitochondrial alternative oxidase contributes to successful tardigrade anhydrobiosis. Front. Zool. 2021, 18, 15. [Google Scholar] [CrossRef]
- Murai, Y.; Yagi-Utsumi, M.; Fujiwara, M.; Tanaka, S.; Tomita, M.; Kato, K.; Arakawa, K. Multiomics study of a heterotardigrade, Echinisicus testudo, suggests the possibility of convergent evolution of abundant heat-soluble proteins in Tardigrada. BMC Genom. 2021, 22, 813. [Google Scholar] [CrossRef]
- Yoshida, Y.; Koutsovoulos, G.; Laetsch, D.R.; Stevens, L.; Kumar, S.; Horikawa, D.D.; Ishino, K.; Komine, S.; Kunieda, T.; Tomita, M.; et al. Comparative genomics of the tardigrades Hypsibius dujardini and Ramazzottius varieornatus. PLoS Biol. 2017, 15, e2002266. [Google Scholar] [CrossRef]
- Kamilari, M.; Jørgensen, A.; Schiøtt, M.; Møbjerg, N. Comparative transcriptomics suggest unique molecular adaptations within tardigrade lineages. BMC Genom. 2019, 20, 607. [Google Scholar] [CrossRef]
- Hengherr, S.; Heyer, A.G.; Köhler, H.R.; Schill, R.O. Trehalose and anhydrobiosis in tardigrades—Evidence for divergence in responses to dehydration. FEBS J. 2008, 275, 281–288. [Google Scholar] [CrossRef]
- Greven, H. From Johann August Ephraim Goeze to Ernst Marcus: A ramble through the history of early tardigrade research (1773 Until 1929). In Water Bears: The Biology of Tardigrades; Schill, R.O., Ed.; Springer: Cham, Switzerland, 2018; Volume 2, pp. 1–55. [Google Scholar] [CrossRef]
- Pigoń, A.; Weglarska, B. The respiration of Tardigrada: A study in animal anabiosis. Bull. Pol. Acad. Sci. 1953, 1, 69–72. [Google Scholar]
- Crowe, J. The physiology of cryptobiosis in tardigrades. Mem. Ist. Ital. Idrobiol. 1975, 32, 37–59. [Google Scholar]
- Hygum, T.L.; Clausen, L.K.B.; Halberg, K.A.; Jørgensen, A.; Møbjerg, N. Tun formation is not a prerequisite for desiccation tolerance in the marine tidal tardigrade Echiniscoides sigismundi. Zool. J. Linn. Soc. 2016, 178, 907–911. [Google Scholar] [CrossRef]
- Wright, J.C. Structural correlates of permeability and tun formation in tardigrade cuticle: An image analysis study. J. Ultrastruct. Res. 1988, 101, 23–39. [Google Scholar] [CrossRef]
- Wełnicz, W.; Grohme, M.A.; Kaczmarek, Ł.; Schill, R.O.; Frohme, M. Anhydrobiosis in tardigrades—The last decade. J. Insect Physiol. 2011, 57, 577–583. [Google Scholar] [CrossRef]
- Wright, J.C. The tardigrade cuticle. I. Fine structure and the distribution of lipids. Tissue Cell 1988, 20, 745–758. [Google Scholar] [CrossRef]
- Sørensen-Hygum, T.L.; Stuart, R.M.; Jørgensen, A.; Møbjerg, N. Modelling extreme desiccation tolerance in a marine tardigrade. Sci. Rep. 2018, 8, 11495. [Google Scholar] [CrossRef]
- Hibshman, J.D.; Clegg, J.S.; Goldstein, B. Mechanisms of desiccation tolerance: Themes and variations in brine shrimp, roundworms, and tardigrades. Front. Physiol. 2020, 11, 592016. [Google Scholar] [CrossRef]
- Giovannini, I.; Boothby, T.C.; Cesari, M.; Goldstein, B.; Guidetti, R.; Rebecchi, L. Production of reactive oxygen species and involvement of bioprotectants during anhydrobiosis in the tardigrade Paramacrobiotus spatialis. Sci. Rep. 2022, 12, 1938. [Google Scholar] [CrossRef]
- Westh, P.; Ramløv, H. Trehalose accumulation in the tardigrade Adorybiotus coronifer during anhydrobiosis. J. Exp. Biol. 1991, 258, 303–311. [Google Scholar] [CrossRef]
- Crowe, L.M. Lessons from nature: The role of sugars in anhydrobiosis. Comp. Biochem. Physiol. Part A Mol. Integr. Physiol. 2002, 131, 505–513. [Google Scholar] [CrossRef]
- Yamaguchi, A.; Tanaka, S.; Yamaguchi, S.; Kuwahara, H.; Takamura, C.; Imajoh-Ohmi, S.; Horikawa, D.D.; Toyoda, A.; Katayama, T.; Arakawa, K.; et al. Two Novel heat-soluble protein families abundantly expressed in an anhydrobiotic tardigrade. PLoS ONE 2012, 7, e44209. [Google Scholar] [CrossRef]
- Boothby, T.C.; Tapia, H.; Brozena, A.H.; Piszkiewicz, S.; Smith, A.E.; Giovannini, I.; Rebecchi, L.; Pielak, G.J.; Koshland, D.; Goldstein, B. Tardigrades use intrinsically disordered proteins to survive desiccation. Mol. Cell 2017, 65, 975–984. [Google Scholar] [CrossRef]
- Schill, R.O.; Steinbrück, G.H.B.; Köhler, H.-R. Stress gene (hsp70) sequences and quantitative expression in Milnesium tardigradum (Tardigrada) during active and cryptobiotic stages. J. Exp. Biol. 2004, 207, 1607–1613. [Google Scholar] [CrossRef]
- Schokraie, E.; Hotz-Wagenblatt, A.; Warnken, U.; Frohme, M.; Dandekar, T.; Schill, R.O.; Schnölzer, M. Investigating heat shock proteins of tardigrades in active versus anhydrobiotic state using shotgun proteomics. J. Zoolog. Syst. Evol. Res. 2011, 49, 111–119. [Google Scholar] [CrossRef]
- Wang, C.; Grohme, M.A.; Mali, B.; Schill, R.O.; Frohme, M. Towards decrypting cryptobiosis—Analyzing anhydrobiosis in the tardigrade Milnesium tardigradum using transcriptome sequencing. PLoS ONE 2014, 9, e92663. [Google Scholar] [CrossRef]
- Wright, J.C. Cryptobiosis 300 years on from van Leuwenhoek: What have we learned about tardigrades? Zool. Anz. 2001, 240, 563–582. [Google Scholar] [CrossRef]
- França, M.B.; Panek, A.D.; Eleutherio, E.C.A. Oxidative stress and its effects during dehydration. Comp. Biochem. Physiol. Part A Mol. Integr. Physiol. 2007, 146, 621–631. [Google Scholar] [CrossRef]
- Hekimi, S.; Lapointe, J.; Wen, Y. Taking a “good” look at free radicals in the aging process. Trends Cell Biol. 2011, 21, 569–576. [Google Scholar] [CrossRef]
- Zhang, J.; Wang, X.; Vikash, V.; Ye, Q.; Wu, D.; Liu, Y.; Dong, W. ROS and ROS-mediated cellular signaling. Oxid. Med. Cell. Longev. 2016, 2016, 4350965. [Google Scholar] [CrossRef]
- Schokraie, E.; Warnken, U.; Hotz-Wagenblatt, A.; Grohme, M.A.; Hengherr, S.; Förster, F.; Schill, R.O.; Frohme, M.; Dandekar, T.; Schnölzer, M. Comparative proteome analysis of Milnesium tardigradum in early embryonic state versus adults in active and anhydrobiotic state. PLoS ONE 2012, 7, e45682. [Google Scholar] [CrossRef]
- Rizzo, A.M.; Negroni, M.; Altiero, T.; Montorfano, G.; Corsetto, P.; Berselli, P.; Berra, B.; Guidetti, R.; Rebecchi, L. Antioxidant defences in hydrated and desiccated states of the tardigrade Paramacrobiotus richtersi. Comp. Biochem. Physiol. B Biochem. 2010, 156, 115–121. [Google Scholar] [CrossRef]
- Förster, F.; Beisser, D.; Grohme, M.A.; Liang, C.; Mali, B.; Siegl, A.M.; Engelmann, J.C.; Shkumatov, A.V.; Schokraie, E.; Müller, T.; et al. Transcriptome analysis in tardigrade species reveals specific molecular pathways for stress adaptations. Bioinform. Biol. Insights 2012, 6, BBI-S9150. [Google Scholar] [CrossRef]
- Rebecchi, L. Dry up and survive: The role of antioxidant defences in anhydrobiotic organisms. J. Limnol. 2013, 72, 62–72. [Google Scholar] [CrossRef]
- Mousseau, T.A.; Roff, D.A. Natural selection and the heritability of fitness components. Heredity 1987, 59, 181–197. [Google Scholar] [CrossRef]
- Tissenbaum, H.A. Using C. elegans for aging research. Invertebr. Reprod. Dev. 2015, 59, 59–63. [Google Scholar] [CrossRef]
- Colman, R.J. Non-human primates as a model for aging. Biochim. Biophys. Acta Mol. Basis Dis. 2018, 1864, 2733–2741. [Google Scholar] [CrossRef]
- Promislow, D.E.L.; Flatt, T.; Bonduriansky, R. The biology of aging in insects: From Drosophila to other insects and back. Annu. Rev. Entomol. 2022, 67, 83–103. [Google Scholar] [CrossRef]
- Austad, S.N. Diverse aging rates in metazoans: Targets for functional genomics. Mech. Ageing Dev. 2005, 126, 43–49. [Google Scholar] [CrossRef]
- Bloom, D.E.; Canning, D.; Lubet, A. Global population aging: Facts, challenges, solutions & perspectives. Daedalus 2015, 144, 80–92. [Google Scholar] [CrossRef]
- Bilinski, T.; Bylak, A.; Zadrag-Tecza, R. The budding yeast Saccharomyces cerevisiae as a model organism: Possible implications for gerontological studies. Biogerontology 2017, 18, 631–640. [Google Scholar] [CrossRef]
- Deweerdt, S. Comparative biology: Looking for a master switch. Nature 2012, 492, S10–S11. [Google Scholar] [CrossRef]
- Hayflick, L. The limited in vitro lifetime of human diploid cell strains. Exp. Cell Res. 1965, 37, 614–636. [Google Scholar] [CrossRef]
- Beckman, K.B.; Ames, B.N. The free radical theory of aging matures. Physiol. Rev. 1998, 78, 547–581. [Google Scholar] [CrossRef]
- Blackburn, E.H. Telomere states and cell fates. Nature 2000, 408, 53–56. [Google Scholar] [CrossRef]
- Murthy, M.; Ram, J.L. Invertebrates as model organisms for research on aging biology. Invertebr. Reprod. Dev. 2015, 59, 1–4. [Google Scholar] [CrossRef]
- Labuschagne, C.F.; Brenkman, A.B. Current methods in quantifying ROS and oxidative damage in Caenorhabditis elegans and other model organism of aging. Ageing Res. Rev. 2013, 12, 918–930. [Google Scholar] [CrossRef]
- Yuan, R.; Peters, L.L.; Paigen, B. Mice as a mammalian model for research on the genetics of aging. ILAR J. 2011, 52, 4–15. [Google Scholar] [CrossRef]
- Brooks-Wilson, A.R. Genetics of healthy aging and longevity. Hum. Genet. 2013, 132, 1323–1338. [Google Scholar] [CrossRef]
- He, C.; Zhou, C.; Kennedy, B.K. The yeast replicative aging model. Biochim. Biophys. Acta Mol. Basis Dis. 2018, 1864, 2690–2696. [Google Scholar] [CrossRef]
- Kirkwood, T.B.L. Understanding the odd science of aging. Cell J. 2005, 120, 437–447. [Google Scholar] [CrossRef]
- Bronikowski, A.M.; Meisel, R.P.; Biga, P.R.; Walters, J.R.; Mank, J.E.; Larschan, E.; Wilkinson, G.S.; Valenzuela, N.; Conard, A.M.; de Magalhães, J.P.; et al. Sex-specific aging in animals: Perspective and future directions. Aging Cell 2022, 21, e13542. [Google Scholar] [CrossRef]
- Hill, K. Life history theory and evolutionary anthropology. Evol. Anthropol. 1993, 2, 78–88. [Google Scholar] [CrossRef]
- Brown, J.L.; Choe, J.C. Behavioral ecology and sociobiology. In Encyclopedia of Animal Behavior; Breed, M.D., Moore, J., Eds.; Academic Press: Oxford, UK, 2019; Volume 1, pp. 103–108. [Google Scholar] [CrossRef]
- Darlington, C.D. New paths in genetics. Nature 1942, 149, 317. [Google Scholar] [CrossRef]
- Williams, G.C. Pleiotropy, natural selection and the evolution of senescence. Evolution 1957, 11, 398–411. [Google Scholar] [CrossRef]
- Stearns, S.C. The Evolution of Life Histories; Oxford University Press: Oxford, UK, 1992; p. 249. [Google Scholar]
- Snell, T.W.; King, C.E. Lifespan and fecundity patterns in rotifers: The cost of reproduction. Evolution 1977, 31, 882–890. [Google Scholar] [CrossRef]
- Schrempf, A.; Cremer, S.; Heinze, J. Social influence on age and reproduction: Reduced lifespan and fecundity in multi-queen ant colonies. J. Evol. Biol. 2011, 24, 1455–1461. [Google Scholar] [CrossRef]
- Monaghan, P.; Maklakov, A.A.; Metcalfe, N.B. Intergenerational transfer of ageing: Parental age and offspring lifespan. Trends Ecol. Evol. 2020, 35, 927–937. [Google Scholar] [CrossRef]
- Kirkwood, T.B.L.; Holliday, R. Ageing as a consequence of natural selection. In The Biology of Human Ageing; Collins, A., Bittles, A., Eds.; Cambridge University Press: Cambridge, UK, 1986; Volume 1, pp. 1–16. [Google Scholar] [CrossRef]
- Hayflick, L. Theories of biological aging. Exp. Gerontol. 1985, 20, 145–159. [Google Scholar] [CrossRef]
- Campisi, J. Cellular senescence and apoptosis: How cellular responses might influence aging phenotypes. Exp. Gerontol. 2003, 38, 5–11. [Google Scholar] [CrossRef]
- Shringarpure, R.; Davies, K.J.A. Protein turnover by the proteasome in aging and disease. Free Radic. Biol. Med. 2002, 32, 1084–1089. [Google Scholar] [CrossRef]
- Deelen, J.; Evans, D.S.; Arking, D.E.; Tesi, N.; Nygaard, M.; Liu, X.; Wojczynski, M.K.; Biggs, M.L.; van der Spek, A.; Atzmon, G.; et al. A meta-analysis of genome-wide association studies identifies multiple longevity genes. Nat. Commun. 2019, 10, 3669. [Google Scholar] [CrossRef]
- López-Otín, C.; Blasco, M.A.; Partridge, L.; Serrano, M.; Kroemer, G. The hallmarks of aging. Cell J. 2013, 153, 1194–1217. [Google Scholar] [CrossRef]
- Gems, D.; Partridge, L. Genetics of longevity in model organisms: Debates and paradigm shifts. Annu. Rev. Physiol. 2013, 75, 621–644. [Google Scholar] [CrossRef]
- Rollins, J.A.; Shaffer, D.; Snow, S.S.; Kapahi, P.; Rogers, A.N. Dietary restriction induces posttranscriptional regulation of longevity genes. Life Sci. Alliance 2019, 2, e201800281. [Google Scholar] [CrossRef]
- Nylin, S.; Gotthard, K. Plasticity in life-history traits. Annu. Rev. Entomol. 1998, 43, 63–83. [Google Scholar] [CrossRef]
- Schill, R.O. Life-history traits in the tardigrade species Paramacrobiotus kenianus and Paramacrobiotus palaui. J. Limnol. 2013, 72, 160–165. [Google Scholar] [CrossRef]
- Ito, M.; Saigo, T.; Abe, W.; Kubo, T.; Kunieda, T. Establishment of an isogenic strain of the desiccation-sensitive tardigrade Isohypsibius myrops (Parachela, Eutardigrada) and its life history traits. Zool. J. Linn. Soc. 2016, 178, 863–870. [Google Scholar] [CrossRef]
- Perry, E.; Miller, W.R.; Kaczmarek, Ł. Recommended abbreviations for the names of genera of the phylum Tardigrada. Zootaxa 2019, 4608, 145–154. [Google Scholar] [CrossRef] [PubMed]
- Perry, E.; Miller, W.R.; Kaczmarek, Ł. Additional recommended abbreviations for the names of genera of the phylum Tardigrada. Zootaxa 2021, 4981, 398–400. [Google Scholar] [CrossRef] [PubMed]
- Altiero, T.; Giovannini, I.; Guidetti, R.; Rebecchi, L. Life history traits and reproductive mode of the tardigrade Acutuncus antarcticus under laboratory conditions: Strategies to colonize the Antarctic environment. Hydrobiologia 2015, 761, 277–291. [Google Scholar] [CrossRef]
- Tsujimoto, M.; Komori, O.; Imura, S. Effect of lifespan and age on reproductive performance of the tardigrade Acutuncus antarcticus: Minimal reproductive senescence. Hydrobiologia 2016, 772, 93–102. [Google Scholar] [CrossRef]
- Altiero, T.; Rebecchi, L. Rearing tardigrades: Results and problems. Zool. Anz. 2001, 240, 217–221. [Google Scholar] [CrossRef]
- Stone, J.; Vasanthan, T. Life history traits for the freshwater tardigrade species Hypsibius exemplaris reared under laboratory conditions. JWB J. Wildl. Biodivers. 2020, 4, 65–72. [Google Scholar] [CrossRef]
- Bingemer, J.; Hohberg, K.; Schill, R.O. First detailed observations on tardigrade mating behaviour and some aspects of the life history of Isohypsibius dastychi Pilato, Bertolani & Binda 1982 (Tardigrada, Isohypsibiidae). Zool. J. Linn. Soc. 2016, 178, 856–862. [Google Scholar] [CrossRef]
- Baumann, H. Lebenslauf und Lebensweise von Macrobiotus hufelandi Schultze (Tardigrada). Veröff. Überseemus. Bremen 1970, 4, 29–43. [Google Scholar]
- Guidetti, R.; Gneuß, E.; Cesari, M.; Altiero, T.; Schill, R.O. Life-history traits and description of the new gonochoric amphimictic Mesobiotus joenssoni (Eutardigrada: Macrobiotidae) from the island of Elba, Italy. Zool. J. Linn. Soc. 2020, 188, 848–859. [Google Scholar] [CrossRef]
- Suzuki, A.C. Life history of Milnesium tardigradum Doyère (Tardigrada) under a rearing environment. Zool. Sci. 2003, 20, 49–57. [Google Scholar] [CrossRef]
- Neumann, S.; Reuner, A.; Brümmer, F.; Schill, R.O. DNA damage in storage cells of anhydrobiotic tardigrades. Comp. Biochem. Physiol. Part A Mol. Integr. Physiol. 2009, 153, 425–429. [Google Scholar] [CrossRef] [PubMed]
- Altiero, T.; Rebecchi, L.; Bertolani, R. Phenotypic variations in the life history of two clones of Macrobiotus richtersi (Eutardigrada, Macrobiotidae). Hydrobiologia 2006, 558, 33–40. [Google Scholar] [CrossRef]
- Hohberg, K. Tardigrade species composition in young soils and some aspects on life history of Macrobiotus richtersi J. Murray, 1911. Pedobiologia 2006, 50, 267–274. [Google Scholar] [CrossRef]
- Guidetti, R.; Jönsson, K.I. Long-term anhydrobiotic survival in semi-terrestrial micrometazoans. J. Zool. 2002, 257, 181–187. [Google Scholar] [CrossRef]
- Rebecchi, L.; Guidetti, R.; Borsari, S.; Altiero, T.; Bertolani, R. Dynamics of long-term anhydrobiotic survival of lichen-dwelling tardigrades. Hydrobiologia 2006, 558, 23–30. [Google Scholar] [CrossRef]
- Horikawa, D.D.; Kunieda, T.; Abe, W.; Watanabe, M.; Nakahara, Y.; Yukuhiro, F.; Sakashita, T.; Hamada, N.; Wada, S.; Funayama, T.; et al. Establishment of a rearing system of the extremotolerant tardigrade Ramazzottius varieornatus: A new model animal for astrobiology. Astrobiology 2008, 8, 549–556. [Google Scholar] [CrossRef] [PubMed]
- Olshansky, S.J.; Gavrilov, L.A.; Gavrilova, N.S.; Skulachev, V.P. The Biology of life span: A quantitative approach. Popul. Dev. Rev. 1992, 18, 555–558. [Google Scholar] [CrossRef]
- Sutphin, G.L.; Kaeberlein, M. Dietary restriction by bacterial deprivation increases life span in wild-derived nematodes. Exp. Gerontol. 2008, 43, 130–135. [Google Scholar] [CrossRef]
- Brüssow, H. What is health? Microb. Biotechnol. 2013, 6, 341–348. [Google Scholar] [CrossRef]
- Kirkwood, T.B.L.; Holliday, R. The evolution of ageing and longevity. Proc. R. Soc. Lond. B Biol. Sci. 1979, 205, 531–546. [Google Scholar] [CrossRef]
- Richaud, M.; Galas, S. Defining the viability of tardigrades with a molecular sensor related to death. PLoS ONE 2018, 13, e0206444. [Google Scholar] [CrossRef] [PubMed]
- Nelson, D.R.; Guidetti, R.; Rebecchi, L. Tardigrada. In Ecology and Classification of North American Freshwater Invertebrates; Thorp, J.H., Covich, A.P., Eds.; Elsevier: Amsterdam, The Netherlands, 2010; Volume 1, pp. 455–484. [Google Scholar] [CrossRef]
- Kristensen, R.M. The first record of cyclomorphosis in Tardigrada based on a new genus and species from Arctic meiobenthos. J. Zool. Syst. Evol. Res. 1983, 20, 249–270. [Google Scholar] [CrossRef]
- Ellison, P.T. Energetics and reproductive effort. Am. J. Hum. Biol. 2003, 15, 342–351. [Google Scholar] [CrossRef] [PubMed]
- Llodra, E.R. Fecundity and life-history strategies in marine invertebrates. Adv. Mar. Biol. 2002, 43, 87–170. [Google Scholar] [CrossRef]
- Ricci, C.; Caprioli, M.; Fontaneto, D. Stress and fitness in parthenogens: Is dormancy a key feature for bdelloid rotifers? BMC Evol. Biol. 2007, 7, S9. [Google Scholar] [CrossRef]
- Kinchin, I.M. The Biology of Tardigrades; Portland Press: London, UK, 1994; Volume 75, pp. 1–186. [Google Scholar] [CrossRef]
- Poprawa, I.; Janelt, K. Reproduction, gonad structure, and oogenesis in tardigrades. In Evo-Devo: Non-Model Species in Cell and Developmental Biology; Tworzydlo, W., Bilinski, S.M., Eds.; Springer: Cham, Switzerland, 2019; Volume 68, pp. 495–513. [Google Scholar] [CrossRef]
- Baker, G.T.; Sprott, R.L. Special issue on biomarkers of aging. Exp. Gerontol. 1988, 23, 223–438. [Google Scholar] [CrossRef]
- Sander, M.; Avlund, K.; Lauritzen, M.; Gottlieb, T.; Halliwell, B.; Stevnsner, T.; Wewer, U.; Bohr, V.A. Aging-from molecules to populations. Mech. Ageing Dev. 2008, 129, 614–623. [Google Scholar] [CrossRef]
- Gladyshev, V.N. On the cause of aging and control of lifespan: Heterogeneity leads to inevitable damage accumulation, causing aging; control of damage composition and rate of accumulation define lifespan. Bioessays 2012, 34, 925–929. [Google Scholar] [CrossRef] [PubMed]
- Pearl, R. Experiments on longevity. Q. Rev. Biol. 1928, 3, 391–407. [Google Scholar] [CrossRef]
- Harman, D. Aging: A theory based on free radical and radiation chemistry. J. Gerontol. 1955, 11, 2–5. [Google Scholar] [CrossRef]
- Arking, R.; Buck, S.; Wells, R.A.; Pretzlaff, R. Metabolic rates in genetically based long lived strains of Drosophila. Exp. Gerontol. 1988, 23, 59–76. [Google Scholar] [CrossRef]
- Le Bourg, E. The rate of living theory. Spontaneous locomotor activity, aging and longevity in Drosophila melanogaster. Exp. Gerontol. 1987, 22, 359–369. [Google Scholar] [CrossRef]
- Lints, F.A.; Le Bourg, E.; Lints, C.V. Spontaneous locomotor activity and life span: A test of the rate of living theory in Drosophila melanogaster. Gerontology 1984, 30, 376–387. [Google Scholar] [CrossRef] [PubMed]
- Golden, T.R.; Hinerfeld, D.A.; Melov, S. Oxidative stress and aging: Beyond correlation. Aging Cell 2002, 1, 117–123. [Google Scholar] [CrossRef] [PubMed]
- Rahman, T.; Hosen, I.; Islam, M.M.T.; Shekhar, H.U. Oxidative stress and human health. Adv. Biosci. Biotechnol. 2012, 03, 997–1019. [Google Scholar] [CrossRef]
- Giovannini, I.; Corsetto, P.A.; Altiero, T.; Montorfano, G.; Guidetti, R.; Rizzo, A.M.; Rebecchi, L. Antioxidant response during the kinetics of anhydrobiosis in two eutardigrade species. Life 2022, 12, 817. [Google Scholar] [CrossRef]
- Levine, R.L.; Stadtman, E.R. Oxidative modification of proteins during aging. Exp. Gerontol. 2001, 36, 1495–1502. [Google Scholar] [CrossRef]
- Turrens, J.F. Mitochondrial formation of reactive oxygen species. Physiol. J. 2003, 552, 335–344. [Google Scholar] [CrossRef]
- Monaghan, P.; Metcalfe, N.B.; Torres, R. Oxidative stress as a mediator of life history trade-offs: Mechanisms, measurements and interpretation. Ecol. Lett. 2009, 12, 75–92. [Google Scholar] [CrossRef]
- Winston, G.W.; Di Giulio, R.T. Prooxidant and antioxidant mechanisms in aquatic organisms. Aquat. Toxicol. 1991, 19, 137–161. [Google Scholar] [CrossRef]
- Zhao, R.-Z.; Jiang, S.; Zhang, L.; Yu, Z.-B. Mitochondrial electron transport chain, ROS generation and uncoupling (Review). Int. J. Mol. Med. 2019, 44, 3–15. [Google Scholar] [CrossRef] [PubMed]
- Rebecchi, L.; Cesari, M.; Altiero, T.; Frigieri, A.; Guidetti, R. Survival and DNA degradation in anhydrobiotic tardigrades. J. Exp. Biol. 2009, 212, 4033–4039. [Google Scholar] [CrossRef] [PubMed]
- Rebecchi, L.; Altiero, T.; Guidetti, R. Anhydrobiosis: The extreme limit of desiccation tolerance. Invertebr. Surviv. J. 2007, 4, 65–81. [Google Scholar]
- Jones, D.P. Radical-free biology of oxidative stress. Am. J. Physiol. Cell Physiol. 2008, 295, C849–C868. [Google Scholar] [CrossRef] [PubMed]
- Møller, I.M.; Jensen, P.E.; Hansson, A. Oxidative modifications to cellular components in plants. Annu. Rev. Plant Biol. 2007, 58, 459–481. [Google Scholar] [CrossRef]
- Kuzmic, M.; Richaud, M.; Cuq, P.; Frelon, S.; Galas, S. Carbonylation accumulation of the Hypsibius exemplaris anhydrobiote reveals age-associated marks. PLoS ONE 2018, 13, e0208617. [Google Scholar] [CrossRef]
- Schokraie, E.; Hotz-Wagenblatt, A.; Warnken, U.; Mali, B.; Frohme, M.; Forster, F.; Dandekar, T.; Hengherr, S.; Schill, R.O.; Schnolzer, M. Proteomic analysis of tardigrades: Towards a better understanding of molecular mechanisms by anhydrobiotic organisms. PLoS ONE 2010, 5, e9502. [Google Scholar] [CrossRef]
- Jové, M.; Mota-Martorell, N.; Pradas, I.; Galo-Licona, J.D.; Martín-Gari, M.; Obis, È.; Sol, J.; Pamplona, R. The lipidome fingerprint of longevity. Molecules 2020, 25, 4343. [Google Scholar] [CrossRef]
- Yavuzer, H.; Yavuzer, S.; Cengiz, M.; Erman, H.; Doventas, A.; Balci, H.; Erdincler, D.S.; Uzun, H. Biomarkers of lipid peroxidation related to hypertension in aging. Hypertens. Res. 2016, 39, 342–348. [Google Scholar] [CrossRef]
- Ziada, A.S.; Smith, M.-S.R.; Côté, H.C.F. Updating the free radical theory of aging. Front. Cell Dev. Biol. 2020, 8, 575645. [Google Scholar] [CrossRef]
- Davies, K.J.A.; Pomatto, L.C.D. Adaptive homeostasis and the free radical theory of ageing. Free Radic. Biol. Med. 2018, 124, 420–430. [Google Scholar] [CrossRef]
- Fernández, C.; Ferreira, E.; Miguel, E.S.; Fernández-Briera, A. Erratum to: Superoxide dismutase and catalase: Tissue activities and relation with age in the long-lived species Margaritifera margaritifera. Biol. Res. 2016, 49, 35. [Google Scholar] [CrossRef]
- Yoshida, Y.; Satoh, T.; Ota, C.; Tanaka, S.; Horikawa, D.D.; Tomita, M.; Kato, K.; Arakawa, K. Time-series transcriptomic screening of factors contributing to the cross-tolerance to UV radiation and anhydrobiosis in tardigrades. BMC Genom. 2022, 23, 405. [Google Scholar] [CrossRef] [PubMed]
- Kondo, K.; Mori, M.; Tomita, M.; Arakawa, K. Pre-treatment with D942, a furancarboxylic acid derivative, increases desiccation tolerance in an anhydrobiotic tardigrade Hypsibius exemplaris. FEBS Open Bio. 2020, 10, 1774–1781. [Google Scholar] [CrossRef]
- Amorim, J.A.; Coppotelli, G.; Rolo, A.P.; Palmeira, C.M.; Ross, J.M.; Sinclair, D.A. Mitochondrial and metabolic dysfunction in ageing and age-related diseases. Nat. Rev. Endocrinol. 2022, 18, 243–258. [Google Scholar] [CrossRef]
- Giorgi, C.; Marchi, S.; Simoes, I.C.M.; Ren, Z.; Morciano, G.; Perrone, M.; Patalas-Krawczyk, P.; Borchard, S.; Jędrak, P.; Pierzynowska, K.; et al. Mitochondria and reactive oxygen species in aging and age-related diseases. Int. Rev. Cell Mol. Biol. 2018, 340, 209–344. [Google Scholar] [CrossRef]
- Finaud, J.; Lac, G.; Filaire, E. Oxidative stress. Sports Med. 2006, 36, 327–358. [Google Scholar] [CrossRef]
- Barja, G. The flux of free radical attack through mitochondrial DNA is related to aging rate. Aging Clin. Exp. Res. 2000, 12, 342–355. [Google Scholar] [CrossRef]
- Kazachkova, N.; Ramos, A.; Santos, C.; Lima, M. Mitochondrial DNA damage patterns and aging: Revising the evidences for humans and mice. Aging Dis. 2013, 4, 337–350. [Google Scholar] [CrossRef]
- Klaus, S.; Ost, M. Mitochondrial uncoupling and longevity—A role for mitokines? Exp. Gerontol. 2020, 130, 110796. [Google Scholar] [CrossRef]
- Son, J.M.; Lee, C. Mitochondria: Multifaceted regulators of aging. BMB Rep. 2019, 52, 13. [Google Scholar] [CrossRef] [PubMed]
- Rurek, M. Plant mitochondria under a variety of temperature stress conditions. Mitochondrion 2014, 19, 289–294. [Google Scholar] [CrossRef] [PubMed]
- Manoli, I.; Alesci, S.; Blackman, M.R.; Su, Y.A.; Rennert, O.M.; Chrousos, G.P. Mitochondria as key components of the stress response. Trends Endocrinol. Metab. 2007, 18, 190–198. [Google Scholar] [CrossRef] [PubMed]
- Halberg, K.A.; Jørgensen, A.; Møbjerg, N. Desiccation tolerance in the tardigrade Richtersius coronifer relies on muscle mediated structural reorganization. PLoS ONE 2013, 8, e85091. [Google Scholar] [CrossRef]
- Poprawa, I.; Bartylak, T.; Kulpla, A.; Erdmann, W.; Roszkowska, M.; Chajec, Ł.; Kaczmarek, Ł.; Karachitos, A.; Kmita, H. Verification of Hypsibius exemplaris Gasiorek et al., 2018 (Eutardigrada; Hypsibiidae) application in anhydrobiosis research. PLoS ONE 2022, 17, e0261485. [Google Scholar] [CrossRef]
- Tanaka, S.; Tanaka, J.; Miwa, Y.; Horikawa, D.D.; Katayama, T.; Arakawa, K.; Toyoda, A.; Kubo, T.; Kunieda, T. Novel mitochondria-targeted heat-soluble proteins identified in the anhydrobiotic tardigrade improve osmotic tolerance of human cells. PLoS ONE 2015, 10, e0118272. [Google Scholar] [CrossRef]
- Tammen, S.A.; Friso, S.; Choi, S.W. Epigenetics: The link between nature and nurture. Mol. Asp. Med. 2013, 34, 753–764. [Google Scholar] [CrossRef]
- Peterson, C.L.; Laniel, M.A. Histones and histone modifications. Curr. Biol. 2004, 14, R546–R551. [Google Scholar] [CrossRef]
- Harshman, S.W.; Young, N.L.; Parthun, M.R.; Freitas, M.A. H1 histones: Current perspectives and challenges. Nucleic Acids Res. 2013, 41, 9593–9609. [Google Scholar] [CrossRef]
- Saul, D.; Kosinsky, R.L. Epigenetics of aging and aging-associated diseases. Int. J. Mol. Sci. 2021, 22, 401. [Google Scholar] [CrossRef]
- Sen, P.; Shah, P.P.; Nativio, R.; Berger, S.L. Epigenetic mechanisms regulating longevity and aging. Cell J. 2016, 166, 822–839. [Google Scholar] [CrossRef] [PubMed]
- D’Aquila, P.; Bellizzi, D.; Passarino, G. Mitochondria in health, aging and diseases: The epigenetic perspective. Biogerontology 2015, 16, 569–585. [Google Scholar] [CrossRef] [PubMed]
- Averbeck, D.; Rodriguez-Lafrasse, C. Role of mitochondria in radiation responses: Epigenetic, metabolic, and signaling impacts. Int. J. Mol. Sci. 2021, 22, 11047. [Google Scholar] [CrossRef] [PubMed]
- Shaughnessy, D.T.; McAllister, K.; Worth, L.; Haugen, A.C.; Meyer, J.N.; Domann, F.E.; van Houten, B.; Mostoslavsky, R.; Bultman, S.J.; Baccarelli, A.A.; et al. Mitochondria, energetics, epigenetics, and cellular responses to stress. Environ. Health Perspect. 2014, 122, 1271–1278. [Google Scholar] [CrossRef]
- Sharma, N.; Pasala, M.S.; Prakash, A. Mitochondrial DNA: Epigenetics and environment. Environ. Mol. Mutagen. 2019, 60, 668–682. [Google Scholar] [CrossRef]
- Storey, K.B. Regulation of hypometabolism: Insights into epigenetic controls. J. Exp. Biol. 2015, 218, 150–159. [Google Scholar] [CrossRef]
- Zhu, X.; Chen, Z.; Shen, W.; Huang, G.; Sedivy, J.M.; Wang, H.; Ju, Z. Inflammation, epigenetics, and metabolism converge to cell senescence and ageing: The regulation and intervention. Signal Transduct. Target. Ther. 2021, 6, 245. [Google Scholar] [CrossRef]
- Cui, D.; Xu, X. Dna methyltransferases, DNA methylation, and age-associated cognitive function. Int. J. Mol. Sci. 2018, 19, 1315. [Google Scholar] [CrossRef]
- Roberts, S.B.; Gavery, M.R. Is there a relationship between DNA methylation and phenotypic plasticity in invertebrates? Front. Physiol. 2012, 2, 116. [Google Scholar] [CrossRef]
- Akhter, Z.; Bi, Z.; Ali, K.; Sun, C.; Fiaz, S.; Haider, F.U.; Bai, J. In response to abiotic stress, DNA methylation confers epigenetic changes in plants. Plants 2021, 10, 1096. [Google Scholar] [CrossRef]
- Dowen, R.H.; Pelizzola, M.; Schmitz, R.J.; Lister, R.; Dowen, J.M.; Nery, J.R.; Dixon, J.E.; Ecker, J.R. Widespread dynamic DNA methylation in response to biotic stress. Proc. Natl. Acad. Sci. USA 2012, 109, E2183–E2191. [Google Scholar] [CrossRef] [PubMed]
- Engelhardt, J.; Scheer, O.; Stadler, P.F.; Prohaska, S.J. Evolution of DNA methylation across Ecdysozoa. J. Mol. Evol. 2022, 90, 56–72. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Yuan, Q.; Xie, L. Histone modifications in aging: The underlying mechanisms and implications. Curr. Stem Cell Res. Ther. 2018, 13, 125–135. [Google Scholar] [CrossRef] [PubMed]
- Henikoff, S.; Shilatifard, A. Histone modification: Cause or cog? Trends Genet. 2011, 27, 389–396. [Google Scholar] [CrossRef] [PubMed]
- Yuan, L.; Liu, X.; Luo, M.; Yang, S.; Wu, K. Involvement of histone modifications in plant abiotic stress responses. J. Integr. Plant Biol. 2013, 55, 892–901. [Google Scholar] [CrossRef]
- Chavez, C.; Cruz-Becerra, G.; Fei, J.; Kassavetis, G.A.; Kadonaga, J.T. The tardigrade damage suppressor protein binds to nucleosomes and protects DNA from hydroxyl radicals. eLife 2019, 8, e47682. [Google Scholar] [CrossRef]
- Gonzalo, S. Epigenetic alterations in aging. J. Appl. Physiol. 2010, 109, 586–597. [Google Scholar] [CrossRef]


{kind=link}
| Species and Reproduction Mode | Lifespan | Fecundity | SSLH | Anhydrobiosis Capability | References | ||||
|---|---|---|---|---|---|---|---|---|---|
| ALS (Days) (Mean + SD) | ML (Days) | AFO (Days) (Mean + SD) | NO (Mean + SD) | ANEC (Mean + SD) | ALO (Mean ± SD) | (High/Medium/Low) | |||
| Acu. antarcticusgeneration F1; parthenogenesis | 88.8 ± 20.0 | ND | 17.1 ± 3.6 | 6.4 ± 1.0 | 1.8 ± 0.8 | 22 * | 22 | ND | [96] |
| Acu. antarcticusgeneration F2; parthenogenesis | 49.5 ± 26.4 | ND | 16.9 ± 3.5 | 3.0 ± 1.9 | 1.7 ± 0.8 | 30 * | 43 | ND | [96] |
| Acu. antarcticus; parthenogenesis | 69.2 ± 36.4 | ND | 9.3 ± 1.1 | 7.5 ± 4.3 | 3.4 ± 2.6 | 66 * | 68 | High | [97] |
| Dip.cf. scoticum; parthenogenesis | ND | 263 | ND | 15 | 1.3 ± 4.7 | ND | 1 | ND | [98] |
| Gre. myrons; parthenogenesis | 18.8 ± 7.0 | 30 | ND | ND | ND | ND | 29 | ND | [93] |
| Hys. exemplaris; parthenogenesis | 61.9 ± 9.9 | 75 | 8.0 ± 3.1 | 10.5 ± 2.2 | 8 * | ND | ≈100 | Low | [99] |
| Iso. dastychi; dioecious | ND | 29 | ND | ND | ND | ND | 15 | ND | [100] |
| Mac. hefelandi; parthenogenesis | ND | 84 | 31 | ND | ND | ND | ND | High | [101] |
| Mac. joannae; dioecious | ND | 266 | ND | ND | 1.7 ± 5.3 | ND | ND | ND | [98] |
| Mac. sapiens; dioecious | 83.0 ± 33.5 | 145 | 16.5 ± 3.8 | ND | Max no 16 * | ND | 66 | ND | [6] |
| Meb. joenssoni; dioecious | 86.5 ± 18.0 | 150 | 19.8 ± 1.7 | ND | ND | ND | 224 | ND | [102] |
| Mil. tardigradum; parthenogenesis | 82.7 ± 2.7 | 107 | ND | ND | ND | ND | 16 | High | [103] |
| Mil. tardigradum; parthenogenesis | 42.7 ± 11.8 | 58 | 15.3 ± 1.6 | 12 * | 1.8 ± 11.2 | ND | ND | High | [16,104] |
| Pam. fairbanski clone 1 parthenogenesis | 194.9 ± 164.4 | ND | 76.9 ± 16.4 | ND | ND | ND | 16 | ND | [105] |
| Pam. fairbanski clone 2 parthenogenesis | 137.3 ± 136.4 | ND | 70.7 ± 19.4 | ND | ND | ND | 15 | ND | [105] |
| Pam. kenianus, Population (I); parthenogenesis | 125 ± 35 | 204 | 10 * | ND | ND | ND | 22 | ND | [92] |
| Pam. kenianus, population (II); parthenogenesis | 141 ± 54 | 212 | 10 * | ND | ND | ND | 22 | ND | [92] |
| Pam. palaui; parthenogenesis | 97 ± 31 | 187 | 10 * | ND | ND | ND | 22 | ND | [92] |
| Pam. richtersi; parthenogenesis | ND | ND | 64.2 ± 1.7 | ND | ND | ND | 80 | High | [106] |
| Pam. tonollii; dioecious | 69.0 ± 45.1 | 237 | 24.4 ± 4.4 | ND | Max no 19 * | ND | 104 | ND | [6] |
| Ram. oberhaeuseri; parthenogenesis | ND | 70 | ND | ND | ND | ND | ND | High | [107,108] |
| Ram. varieornatus; parthenogenesis | 13-87 | 87 | ND | ND | ND | ND | 10 | High | [109] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Nagwani, A.K.; Kaczmarek, Ł.; Kmita, H. Applicable Life-History and Molecular Traits for Studying the Effects of Anhydrobiosis on Aging in Tardigrades. Diversity 2022, 14, 664. https://doi.org/10.3390/d14080664
Nagwani AK, Kaczmarek Ł, Kmita H. Applicable Life-History and Molecular Traits for Studying the Effects of Anhydrobiosis on Aging in Tardigrades. Diversity. 2022; 14(8):664. https://doi.org/10.3390/d14080664
Chicago/Turabian StyleNagwani, Amit Kumar, Łukasz Kaczmarek, and Hanna Kmita. 2022. "Applicable Life-History and Molecular Traits for Studying the Effects of Anhydrobiosis on Aging in Tardigrades" Diversity 14, no. 8: 664. https://doi.org/10.3390/d14080664