
Activity Patterns of Stenocercus iridescens in an Ecuadorian Coastal Agroecosystem: Is Temperature Important?

 , ,
, ,
Abstract
:1. Introduction
2. Materials and Methods
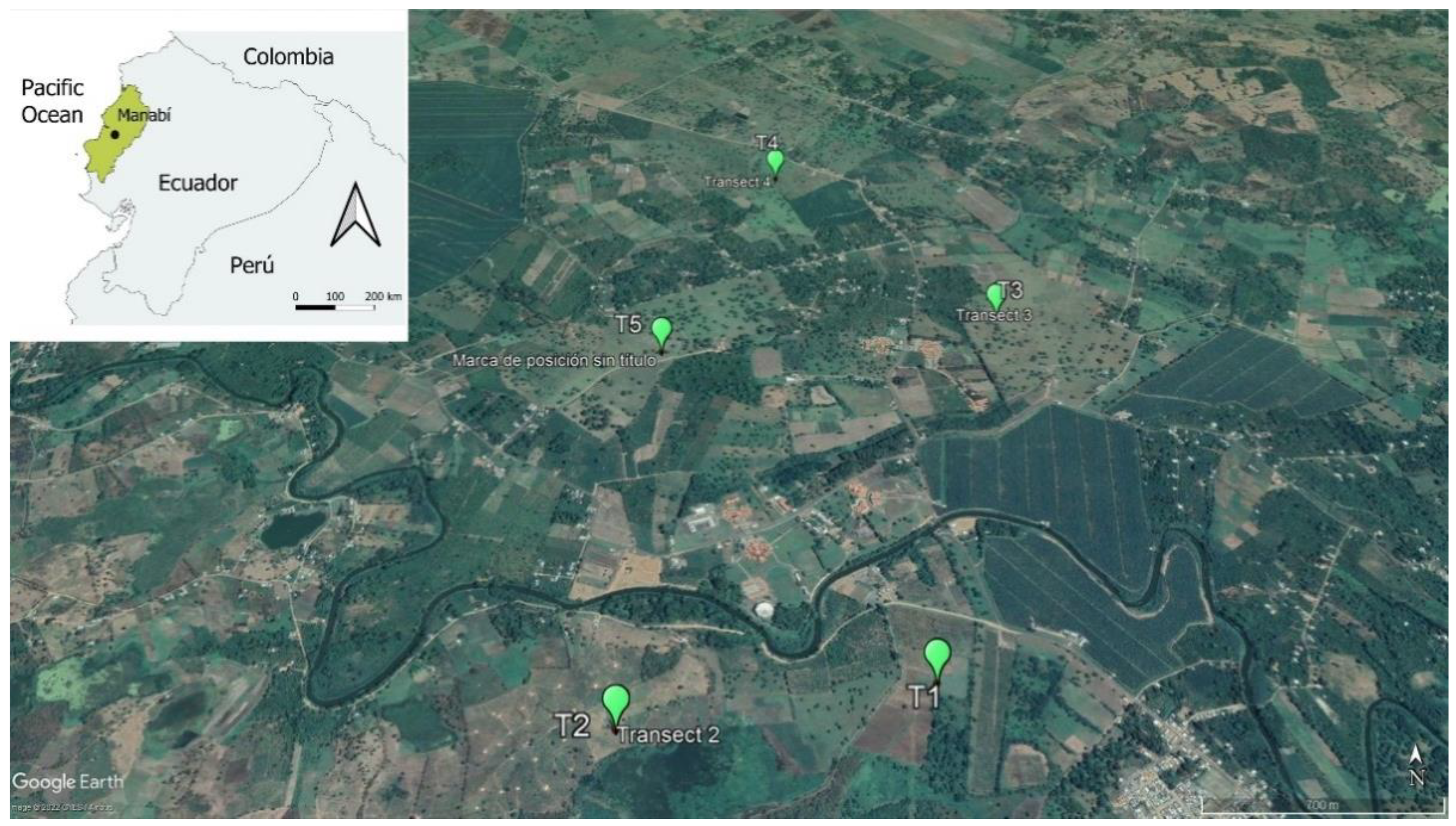
2.1. Study Area
2.2. Lizard Sampling
2.3. Statistical Analysis
3. Results
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Foley, J.A.; DeFries, R.; Asner, G.P.; Barford, C.; Bonan, G.; Carpenter, S.R.; Chapin, F.S.; Coe, M.T.; Daily, G.C.; Gibbs, H.K.; et al. Global consequences of land use. Science 2005, 309, 570–574. [Google Scholar] [CrossRef] [PubMed]
- Stuart, S.N.; Chanson, J.S.; Cox, N.A.; Young, B.E.; Rodrigues, A.S.L.; Fischman, D.L.; Waller, R.W. Status and trends of amphibian declines and extinctions worldwide. Science 2004, 306, 1783–1786. [Google Scholar] [CrossRef] [PubMed]
- Doherty, T.S.; Balouch, S.; Bell, K.; Burns, T.J.; Feldman, A.; Fist, C.; Garvey, T.F.; Jessop, T.S.; Meiri, S.; Driscoll, D.A. Reptile responses to anthropogenic habitat modification: A global meta-analysis. Glob. Ecol. Biogeogr. 2020, 29, 1265–1279. [Google Scholar] [CrossRef]
- Stoate, C.; Baldi, A.; Beja, P.; Boatman, N.D.; Herzon, I.; van Doorn, A.; de Snoo, G.R.; Rakosy, L.; Ramwell, C. Ecological impacts of early 21st century agricultural change in Europe—A review. J. Environ. Manag. 2009, 91, 22–46. [Google Scholar] [CrossRef] [PubMed]
- Myers, N.; Mittermeier, R.A.; Mittermeier, C.G.; da Fonseca, G.A.B.; Kent, J. Biodiversity otspots for conservation priorities. Nature 2000, 403, 853–858. [Google Scholar] [CrossRef] [PubMed]
- Espinosa, C.I.; de la Cruz, M.; Luzuriaga, A.; Escudero, A. Bosques tropicales secos de la región Pacífico Ecuatorial: Diversidad, estructura, funcionamiento e implicaciones para la conservación. Rev. Ecosistemas 2012, 21, 167–179. [Google Scholar]
- Reyes-Puig, C.; Almendáriz, A.; Torres-Carvajal, O. Diversity, threat, and conservation of reptiles from continental Ecuador. Amphib. Reptile Conserv. 2017, 11, 51–58. [Google Scholar]
- Ferrer-Paris, J.R.; Zager, I.; Keith, D.A.; Oliveira-Miranda, M.A.; Rodríguez, J.P.; Barrow, E. An ecosystem risk assessment of temperate and tropical forests of the Americas with an outlook on future conservation strategies. Conserv. Lett. 2019, 12, e12623. [Google Scholar] [CrossRef]
- Rivas, C.A.; Navarro-Cerillo, R.M.; Johnston, J.C.; Guerrero-Casado, J. Dry forest is more threatened but less protected than evergreen forest in Ecuador’s coastal region. Environ. Conserv. 2020, 47, 79–83. [Google Scholar] [CrossRef]
- Rivas, C.A.; Guerrero-Casado, J.; Navarro-Cerillo, R.M. Deforestation and fragmentation trends of seasonal dry tropical forest in Ecuador: Impact on conservation. For. Ecosyst. 2021, 8, 46. [Google Scholar] [CrossRef]
- Ghosh, D.; Basu, P. Factors influencing herpetofauna abundance and diversity in a tropical agricultural landscape mosaic. Biotropica 2020, 52, 927–937. [Google Scholar] [CrossRef]
- Rodriguez, A.C.Z.; Gamez, M.R.; Faure, L.G. Design, construction, and energy of sustainable solar dryers in Jipijapa Canton. Int. J. Phys. Sci. Eng. 2018, 2, 88–100. [Google Scholar] [CrossRef]
- Žagar, A.; Carretero, M.A.; Osojnik, N.; Sillero, N.; Vrezec, A. A place in the sun: Interspecific interference affects thermoregulation in coexisting lizards. Behav. Ecol. Sociobiol. 2015, 69, 1127–1137. [Google Scholar] [CrossRef]
- Méndez-Galeano, M.A.; Calderón-Espinosa, M.L. Thermoregulation in the Andean lizard Anolis heterodermus (Squamata: Dactyloidae) at high elevation in the Eastern Cordillera of Colombia. Iheringia. Série Zool. 2017, 107, e2017018. [Google Scholar] [CrossRef]
- Templeton, J.R.; Whittow, G.C. (Eds.) Comparative Physiology of Thermoregulation: Invertebrates and Nonmammalian Vertebrates; Academic Press, Inc.: London, UK, 1970; Volume 1, pp. 167–218. [Google Scholar]
- Gunderson, A.R.; Stillman, J.H. Plasticity in thermal tolerance has limited potential to buffer ectotherms from global warming. Proc. R. Soc. B Biol. Sci. 2015, 282, 20150401. [Google Scholar] [CrossRef]
- Abram, P.K.; Boivin, G.; Moiroux, J.; Brodeur, J. Behavioural effects of temperature on ectothermic animals: Unifying thermal physiology and behavioural plasticity. Biol. Rev. 2017, 92, 1859–1876. [Google Scholar] [CrossRef]
- Huey, R.; Stevenson, R. Integrating Thermal Physiology and Ecology of Ectotherms: A Discussion of Approaches. Am. Zool. 1979, 19, 357–366. [Google Scholar] [CrossRef]
- Kearney, M.; Shine, R.; Porter, W.P. The potential for behavioral thermoregulation to buffer “cold-blooded” animals against climate warming. Proc. Natl. Acad. Sci. USA 2009, 106, 3835–3840. [Google Scholar] [CrossRef]
- Huey, R.B.; Deutsch, C.A.; Tewksbury, J.J.; Vitt, L.J.; Hertz, P.E.; Alvarez Pérez, H.J.; Garland, T., Jr. Why tropical forest lizards are vulnerable to climate warming. Proc. R. Soc. B Biol. Sci. 2009, 276, 1939–1948. [Google Scholar] [CrossRef]
- Ibargüengoytía, N. Field, selected body temperature and thermal tolerance of the syntopic lizards Phymaturus patagonicus and Liolaemus elongatus (Iguania: Liolaemidae). J. Arid Environ. 2005, 62, 435–448. [Google Scholar] [CrossRef]
- Rozen-Rechels, D.; Dupoué, A.; Lourdais, O.; Chamaillé-Jammes, S.; Meylan, S.; Clobert, J.; Le Galliard, J.F. When water interacts with temperature: Ecological and evolutionary implications of thermo-hydroregulation in terrestrial ectotherms. Ecol. Evol. 2019, 9, 10029–10043. [Google Scholar] [CrossRef]
- Pawson, S.; Gass, J. Modern Era Retrospective Analysis Research and Applications, Version 2 (MERA-2); NASA: Washington, DC, USA, 2019. Available online: https://gmao.gsfc.nasa.gov/reanalysis/MERRA-2/ (accessed on 2 January 2019).
- Carpio, A.J.; Cabrera, M.; Tortosa, F.S. Evaluation of Methods for Estimating Species Richness and Abundance of Reptiles in Olive Groves. Herpetol. Conserv. Biol. 2015, 10, 54–63. [Google Scholar]
- Hutchens, S.J.; DePerno, C.S. Efficacy of sampling techniques for determining species richness estimates of reptiles and amphibians. Wild. Biol. 2009, 15, 113–122. [Google Scholar] [CrossRef]
- Sung, Y.H.; Karraker, N.E.; Hau, B.C. Evaluation of the effectiveness of three survey methods for sampling terrestrial herpetofauna in South China. Herpetol. Conserv. Biol. 2011, 6, 479–489. [Google Scholar]
- Burnham, K.P.; Anderson, D.R.; Huyvaert, K.P. AIC model selection and multimodel inference in behavioral ecology: Some background, observations, and comparisons. Behav. Ecol. Sociobiol. 2011, 65, 23–35. [Google Scholar] [CrossRef]
- Di Rienzo, J.A.; Casanoves, F.; Balzarini, M.G.; González, L.; Tablada, M.; Robledo, C.W. InfoStat Version; Universidad Nacional de Córdoba: Córdoba, Argentina, 2020. [Google Scholar]
- Radder, R.S.; Saidapur, S.K.; Shanbhag, B.A. Population density, microhabitat use and activity pattern of the Indian rock lizard, Psammophilus dorsalis (Agamidae). Curr. Sci. 2005, 89, 560–565. [Google Scholar]
- Mukherjee, R.K.; Parida, P. Behavioral ecology, breeding period, sexual dimorphism and ovipositional behavior of Psammophilus blanfordanus (Family: Agamidae): Case study. Indian J. Appl. Res. Biol. 2014, 4, 28–32. [Google Scholar] [CrossRef]
- Bauwens, D.; Castilla, A.M.; Mounton, F.N. Field body temperatures, activity levels and opportunities for thermoregulation in an extreme microhabitat specialist, the girdled lizard (Cordylus macropholis). J. Zool. 1999, 249, 11–18. [Google Scholar] [CrossRef]
- Winne, C.T.; Keck, M.B. Daily activity patterns of whiptail lizards (Squamata: Teiidae: Aspidoscelis): A proximate response to environmental conditions or an endogenous rhythm? Funct. Ecol. 2004, 18, 314–321. [Google Scholar] [CrossRef]
- Atencia, P.L.; Castillo, C.J.; Montes, L.F. Use of microhabitat and activity patterns of two lizard species from a seasonal dry forest in northern Colombia. Neotrop. Biol. Conserv. 2020, 15, 153. [Google Scholar] [CrossRef]
- Rozen-Rechels, D.; Rutschmann, A.; Dupoué, A.; Blaimont, P.; Chauveau, V.; Miles, D.B.; Guillon, M.; Richard, M.; Badiane, A.; Meylan, S.; et al. Interaction of hydric and thermal conditions drive geographic variation in thermoregulation in a widespread lizard. Ecol. Monogr. 2021, 91, e01440. [Google Scholar] [CrossRef]
- Dupoué, A.; Rutschmann, A.; Le Galliard, J.F.; Miles, D.B.; Clobert, J.; DeNardo, D.F.; Meylan, S. Water availability and environmental temperature correlate with geographic variation in water balance in common lizards. Oecologia 2017, 185, 561–571. [Google Scholar] [CrossRef] [PubMed]
- Kobayashi, D.; Mautz, W.J.; Nagy, K.A. Evaporative Water Loss: Humidity Acclimation in Anolis carolinensis Lizards. Copeia 1983, 1983, 701–704. [Google Scholar] [CrossRef]
- Sannolo, M.; Carretero, M.A. Dehydration constrains thermoregulation and space use in lizards. PLoS ONE 2019, 14, e0220384. [Google Scholar]
- Wei, X.; Yan, L.; Zhao, C.; Zhang, Y.; Xu, Y.; Cai, B.; Jiang, N.; Huang, Y. Geographic variation in body size and its relationship with environmental gradients in the Oriental Garden Lizard, Calotes versicolor. Ecol. Evol. 2018, 8, 4443–4454. [Google Scholar] [CrossRef]
- Tewksbury, J.J.; Huey, R.B.; Deutsch, C.A. Putting the heat on tropical animals. Science 2008, 320, 1296–1297. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| Candidate Models | k | AICc | ∆AICc |
|---|---|---|---|
| Dry Season | |||
| Null model | 0 | 578.81 | |
| Time range × Heliophany | 2 | 493.63 | 0.00 |
| Time range × Heliophany × Temperature | 3 | 494.46 | 0.83 |
| Time range × Heliophany × Precipitation | 3 | 495.50 | 1.77 |
| Time range | 1 | 495.79 | 2.16 |
| Wet Season | |||
| Null model | 0 | 536 | |
| Time range | 1 | 486.67 | 0.00 |
| Time range × Heliophany | 2 | 486.83 | 0.16 |
| Time range × Heliophany × Precipitation | 3 | 488.62 | 1.92 |
| Time range × Temperature | 2 | 488.62 | 1.95 |
| Time range × Heliophany × Temperature | 3 | 488.72 | 2.05 |
| Variables | Estimate (±SE) | Z | p-Value |
|---|---|---|---|
| Dry Season | |||
| Intercept | 0.13 (±0.29) | 0.46 | 0.646 |
| Time range 10–13 h | 0.78 (±0.27) | 2.88 | 0.004 |
| Time range 13–16 h | 1.50 (±0.27) | 5.63 | <0.001 |
| Time range 16–18 h | 1.47 (±0.27) | 5.52 | <0.001 |
| Heliophany | −0.17 (±0.07) | −2.31 | 0.021 |
| Wet Season | |||
| Intercept | 0.31 (±0.18) | 1.69 | 0.091 |
| Time range 10–13 h | 0.52 (±0.21) | 2.46 | 0.014 |
| Time range 13–16 h | 1.03 (±0.20) | 5.09 | <0.001 |
| Time range 16–18 h | 1.16 (±0.20) | 5.83 | <0.001 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zambrano, R.H.; Guerrero-Casado, J.; Centeno, V.A.; Tortosa, F.S. Activity Patterns of Stenocercus iridescens in an Ecuadorian Coastal Agroecosystem: Is Temperature Important? Diversity 2022, 14, 662. https://doi.org/10.3390/d14080662
Zambrano RH, Guerrero-Casado J, Centeno VA, Tortosa FS. Activity Patterns of Stenocercus iridescens in an Ecuadorian Coastal Agroecosystem: Is Temperature Important? Diversity. 2022; 14(8):662. https://doi.org/10.3390/d14080662
Chicago/Turabian StyleZambrano, Ramón H., José Guerrero-Casado, Víctor A. Centeno, and Francisco S. Tortosa. 2022. "Activity Patterns of Stenocercus iridescens in an Ecuadorian Coastal Agroecosystem: Is Temperature Important?" Diversity 14, no. 8: 662. https://doi.org/10.3390/d14080662