
Diversity and Functional Relevance of Canopy Arthropods in Central Europe

Abstract
:1. Introduction
2. Materials and Methods
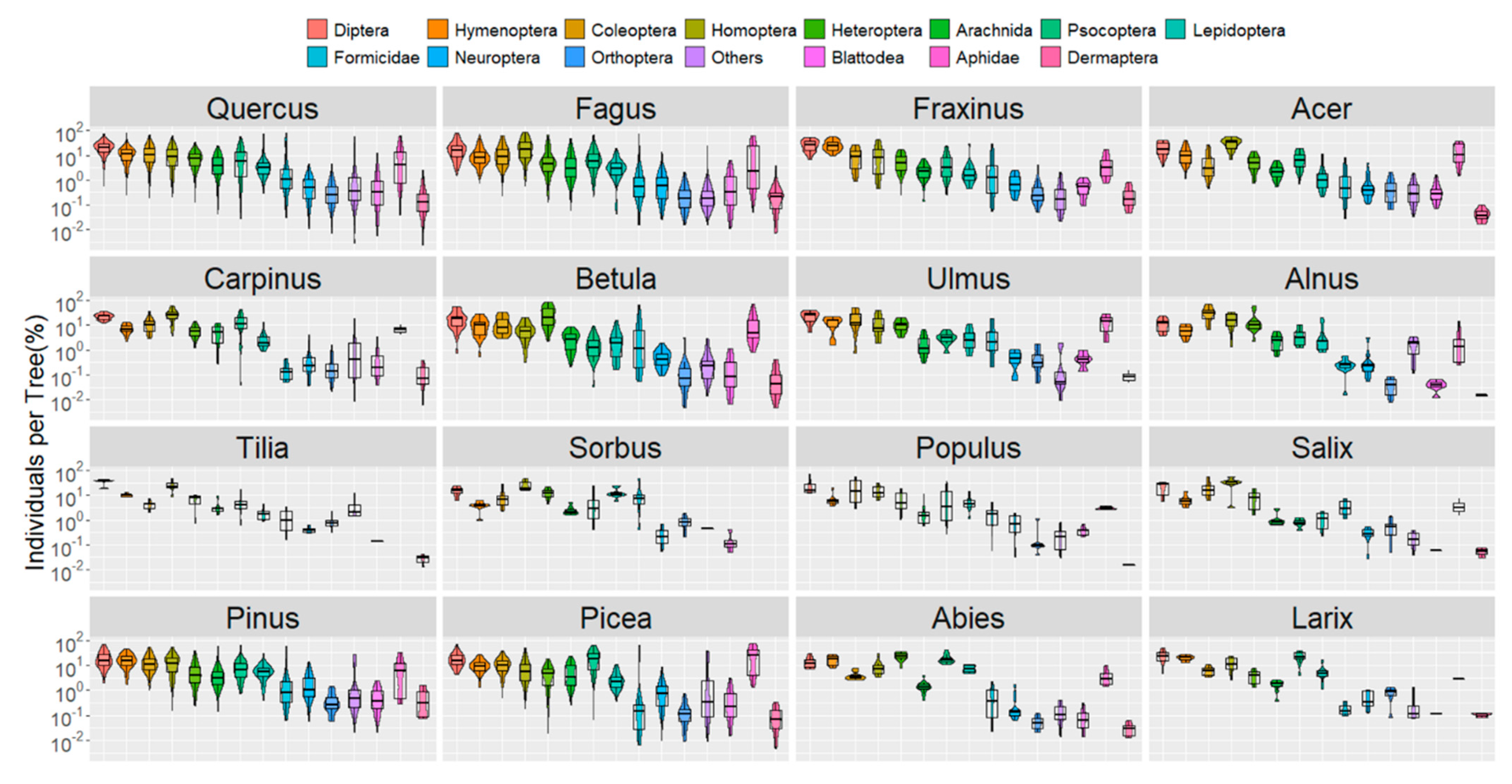
- Comparison between tree genera (violin plots, bar plots of taxa ranks);
- Comparison between deciduous and coniferous trees (boxplots of individuals per tree genera).
- Comparison of guild composition between tree genera (level-plot of individual percentage per guild, boxplot of deciduous vs. coniferous tree genera). To verify the significance of the results, the analyses were then performed on a more homogeneous, smaller, nested data set. For this purpose, we chose the oaks in Poland and, in particular, in the Bialowieza Forest, which also enabled comparison between primary and commercial forests located in the same forest matrix. Selected data level 3: Quercus trees in Poland (N = 103):
- Guild composition (PERMANOVA; Permutational Multivariate Analysis of Variance using Distance Matrices); adonis2 function, as implemented in vegan R package [28].
- Guild composition in primary and managed forest sites (logistic regression).
Bialowieza Forest
3. Results
3.1. Arthropods in Trees
3.2. Functional Classification of Canopy Communities
4. Discussion
4.1. Composition of Canopy Communities
4.2. Guild Composition
4.3. Functional Importance of Canopy Arthropods
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Erwin, T.L. Tropical Forests: Their richness in Coleoptera and other arthropod species. Coleopt. Bull. 1982, 361, 74–75. [Google Scholar]
- Floren, A.; Schmidl, J. Canopy Arthropod Research in Central Europe—Basic and Applied Studies from the High Frontier; Bioform: Nürnberg, Germany, 2008; p. 576. [Google Scholar]
- Volf, M.; Klimes, P.; Lamarre, G.P.A.; Redmond, C.M.; Seifert, C.L.; Abe, T.; Auga, J.; Anderson-Teixeira, K.; Basset, Y.; Beckett, S.; et al. Quantitative assessment of plant-arthropod interactions in forest canopies: A plot-based approach. PLoS ONE 2019, 14, e0222119. [Google Scholar] [CrossRef] [PubMed]
- Swart, R.C.; Samways, M.J.; Roets, F. Tree canopy arthropods have idiosyncratic responses to plant ecophysiological traits in a warm temperate forest complex. Sci. Rep. 2020, 10, 19905. [Google Scholar] [CrossRef]
- Floren, A.; Sprick, P.; Horchler, P.J.; Müller, T. Baumkronen als Habitat gefährdeter Käfer am Beispiel von Hartholzauwäldern in Sachsen-Anhalt, Region Mittelelbe. Nat. Lanschaft 2021, 11, 509–516. [Google Scholar]
- Floren, A.; Horchler, P.J.; Müller, T. The Impact of the Neophyte Tree Fraxinus pennsylvanica [Marshall] on Beetle Diversity under Climate Change. Sustainability 2022, 14, 1914. [Google Scholar] [CrossRef]
- Brockerhoff, E.G.; Barbaro, L.; Castagneyrol, B.; Forrester, D.I.; Gardiner, B.; González-Olabarria, J.R.; Lyver, P.O.; Meurisse, N.; Oxbrough, A.; Taki, H.; et al. Forest biodiversity, ecosystem functioning and the provision of ecosystem services. Biodivers. Conserv. 2017, 26, 3005–3035. [Google Scholar] [CrossRef]
- Floren, A. Sampling arthropods from the canopy by insecticidal knockdown. In Manual on Field Recording Techniques and Protocols for All Taxa Biodiversity Inventories and Monitoring; Eymann, J., Degreff, J., Häuser, C., Eds.; Abc Taxa: Brussels, Belgium, 2010; Volume 1, pp. 158–172. [Google Scholar]
- Vaca-Sánchez, M.S.; Maldonado-López, Y.; González-Rodríguez, A.; Oyama, K.; Fernandes, G.W.; Fagundes, M.; López-Barbosa, E.C.; Aguilar-Peralta, J.S.; Cuevas-Reyes, P. Canopy arthropod diversity associated with Quercus laurina: Importance of an oak species diversity gradient on abundance, species richness and guild composition. J. Insect Conserv. 2021, 25, 859–874. [Google Scholar] [CrossRef]
- Southwood, T.R.E. The Number of Species of Insect Associated with Various Trees. J. Anim. Ecol. 1961, 30, 1–8. [Google Scholar] [CrossRef]
- Moran, V.C.; Southwood, T.R.E. The Guild Composition of Arthropod Communities in Trees. J. Anim. Ecol. 1982, 51, 289. [Google Scholar] [CrossRef]
- Southwood, T.E.R.; Wint, G.W.; Kennedy, C.E.; Greenwood, S.R. The composition of the arthropod fauna of the canopies of some species of oak (Quercus). Eur. J. Entomol. 2005, 102, 65–72. [Google Scholar] [CrossRef]
- Ozanne, C.M.P. A comparison of the canopy arthropod communities of coniferous and broad-leaved trees in the United Kingdom. Selbyana 1999, 20, 290–298. [Google Scholar]
- Thunes, K.H.; Skarveit, J.; Gjerde, I. The canopy arthropods of old and mature pine Pinus sylvestris in Norway. Ecography 2003, 26, 490–502. [Google Scholar] [CrossRef]
- Mottl, O.; Fibich, P.; Klimes, P.; Volf, M.; Tropek, R.; Anderson-Teixeira, K.; Auga, J.; Blair, T.; Butterill, P.; Carscallen, G.; et al. Spatial covariance of herbivorous and predatory guilds of forest canopy arthropods along a latitudinal gradient. Ecol. Lett. 2020, 23, 1499–1510. [Google Scholar] [CrossRef]
- Leidinger, J.; Seibold, S.; Weisser, W.W.; Lange, M.; Schall, P.; Türke, M.; Gossner, M.M. Effects of forest management on herbivorous insects in temperate Europe. For. Ecol. Manag. 2019, 437, 232–245. [Google Scholar] [CrossRef]
- Brändle, M.; Brandl, R. Species richness of insects and mites on trees: Expanding Soutwood. J. Anim. Ecol. 2001, 70, 491–504. [Google Scholar] [CrossRef]
- Thunes, K.H.; Søli, G.E.; Thuróczy, C.; Fjellberg, A.; Olberg, S.; Roth, S.; Coulianos, C.-C.; Disney, R.H.L.; Starý, J.; Vierbergen, G.B.; et al. The Arthropod Fauna of Oak Quercus spp., Fagaceae Canopies in Norway. Diversity 2021, 13, 332. [Google Scholar] [CrossRef]
- Kovács, B.; Tinya, F.; Németh, C.; Ódor, P. Unfolding the effects of different forestry treatments on microclimate in oak forests: Results of a 4-yr experiment. Ecol. Appl. 2020, 30, e02043. [Google Scholar] [CrossRef]
- Schall, P.; Heinrichs, S.; Ammer, C.; Ayasse, M.; Boch, S.; Buscot, F.; Fischer, M.; Goldmann, K.; Overmann, J.; Schulze, E.-D.; et al. Can multi-taxa diversity in European beech forest landscapes be increased by combining different management systems? J. Appl. Ecol. 2020, 57, 1363–1375. [Google Scholar] [CrossRef]
- Blicharska, M.; Angelstam, P.; Giessen, L.; Hilszczański, J.; Hermanowicz, E.; Holeksa, J.; Jacobsen, J.; Jaroszewicz, B.; Konczal, A.; Konieczny, A.; et al. Between biodiversity conservation and sustainable forest management—A multidisciplinary assessment of the emblematic Białowieża Forest case. Biol. Conserv. 2020, 248, 108614. [Google Scholar] [CrossRef]
- Faliński, J.B. Vegetation Dynamics in Temperate Lowland Primeval Forests: Ecological Studies in Białowieża Forest; Geobotany 8; Springer: Dordrecht, The Netherlands, 1986; 537p. [Google Scholar]
- Jedrzejewska, B.; Jedrzejewski, W. Predation in Vertebrate Communities: The Bialowieza Primeval Forest as a Case Study; Ecological studies; Springer: Berlin/Heidelberg, Germany, 1998; Volume 135, p. 450. [Google Scholar]
- Oguh, C.E.; Okpaka, C.O.; Ubani, C.S.; Okekeaji, U.; Joseph, P.S.; Amadi, E.U. Natural Pesticides Biopesticides and Uses in Pest Management—A Critical Review. Asian J. Biotechnol. Genet. Eng. 2019, 2, 1–18. [Google Scholar]
- Soderlund, D.M.; Clark, J.M.; Sheets, L.P.; Mullin, L.S.; Piccirillo, V.J.; Sargent, D.; Stevens, J.T.; Weiner, M.L. Mechanisms of pyrethroid neurotoxicity: Implications for cumulative risk assessment. Toxicology 2002, 171, 3–59. [Google Scholar] [CrossRef]
- Floren, A.; Sprick, P.; Horchler, P.J.; Müller, T. Response to the letter to the editor by Raimund Rodewald: Beetle census by means of pesticide fogging—An ethical no-go! Nat. Landsch. 2022, 4, 217–218. [Google Scholar]
- Floren, A. Abundance and ordinal composition of arboreal arthropod communities of various trees in old primary and managed forests. In Canopy Arthropod Research in Europe; Floren, A., Schmidl, J., Eds.; Bioform: Nürnberg, Germany, 2008; pp. 279–298. [Google Scholar]
- Oksanen, J.; Blanchet, F.G.; Kindt, R.; Legendre, P.; Minchin, P.R.; O’Hara, R.B.; Simpson, G.V.; Solymnos, P.; Henry, M.; Stevens, H. Vegan: Community Ecology; R Package Version 1.18-28/r1569; 2013. Available online: http://R-Forge.R-project.org/projects/vegan (accessed on 13 July 2022).
- Huber, W.; Carey, V.J.; Gentleman, R.; Anders, S.; Carlson, M.; Carvalho, B.S.; Bravo, H.C.; Davis, S.; Gatto, L.; Girke, T.; et al. Orchestrating high-throughput genomic analysis with Bioconductor. Nat. Methods 2015, 12, 115–121. [Google Scholar] [CrossRef] [PubMed]
- Dunnington, D. Rosm: Plot Rater Map Tiles from Open Street Map and Other Sources; R-Package Version 0.2.5; 2019. Available online: http://docplayer.net/206163828-Package-rosm-july-22-2019.html (accessed on 13 July 2022).
- Dunnington, D. Prettymapr: Scale Bar, North Arrow, and Pretty Margins in R; R Package Version 0.2.2; 2017. Available online: https://pkgsrc.se/geography/R-prettymapr (accessed on 13 July 2022).
- Benjamini, Y.; Hochberg, Y. Controlling the False Discovery Rate: A Practical and Powerful Approach to Multiple Testing. J. R. Stat. Soc. Ser. B 1995, 57, 289–300. [Google Scholar] [CrossRef]
- Wickham, H. ggplot2: Elegant Graphics for Data Analysis; Springer: New York, NY, USA, 2016. [Google Scholar]
- McArdle, B.H.; Anderson, M.J. Fitting multivariate models to community data: A comment on distance-baseed redundancy analysis. Ecology 2001, 82, 290–297. [Google Scholar] [CrossRef]
- Leroy, B.M.L.; Seibold, S.; Morinière, J.; Bozicevic, V.; Jaworek, J.; Roth, N.; Vogel, S.; Zytynska, S.; Petercord, R.; Eichel, P.; et al. Metabarcoding of canopy arthropods reveals negative impacts of forestry insecticides on community structure across multiple taxa. J. Appl. Ecol. 2022, 59, 997–1012. [Google Scholar] [CrossRef]
- Mupepele, A.-C.; Müller, T.; Dittrich, M.; Floren, A. Are Temperate Canopy Spiders Tree-Species Specific? PLoS ONE 2014, 9, e86571. [Google Scholar]
- Nyffeler, M.; Birkhofer, K. An estimated 400–800 million tons of prey are annually killed by the global spider community. Sci. Nat. 2017, 104, 30. [Google Scholar] [CrossRef]
- Huryn, A.D.; Wallace, J.B. Life History and Production of Stream Insects. Annu. Rev. Entomol. 2000, 45, 83–110. [Google Scholar] [CrossRef]
- Chimeno, C.; Hausmann, A.; Schmidt, S.; Raupach, M.J.; Doczkal, D.; Baranov, V.; Hübner, J.; Höcherl, A.; Albrecht, R.; Jaschhof, M.; et al. Peering into the Darkness: DNA Barcoding Reveals Surprisingly High Diversity of Unknown Species of Diptera (Insecta) in Germany. Insects 2022, 13, 82. [Google Scholar] [CrossRef]
- Huang, J.; Miao, X.; Wang, Q.; Menzel, F.; Tang, P.; Yang, D.; Wu, H.; Vogler, A.P. Metabarcoding reveals massive species diversity of Diptera in a subtropical ecosystem. Ecol. Evol. 2022, 12, e8535. [Google Scholar] [CrossRef] [PubMed]
- Cornell, H.V.; Kahn, D.M. Guild Structure in the British Arboreal Arthropods: Is it Stable and Predictable? J. Anim. Ecol. 1989, 58, 1003–1020. [Google Scholar] [CrossRef]
- Offenberg, J.; Nielsen, J.S.; Damgaard, C. Wood Ant (Formica polyctena) Services and Disservices in a Danish Apple Plantation. Sociobiology 2019, 66, 247–256. [Google Scholar] [CrossRef]
- Bauhaus, J.; Seeling, U.; Dieter, M.; Farwig, N.; Hafner, A.; Kätzel, R.; Kleinschmit, B.; Lang, F.; Lindner, M.; Möhring, B.; et al. Wissenschaftlicher Beirat für Waldpolitik: Die Anpassung von Wäldern und Waldwirtschaft an den Klimawandel; Federal Ministry of Food and Agriculture: Berlin, Germany, 2021; p. 192. [Google Scholar]
- Nielsen, U.N. Soil Fauna Assemblages: Global to Local Scales; Cambridge University Press: Cambidge, MA, USA, 2019; p. 365. [Google Scholar]
- Bardgett, R.D.; Wardle, D.A. Aboveground-Belowground Linkages: Biotic Interactions, Ecosystem Processes, and Global Change; Oxford Series in Ecology and Evolution; Harvey, P.H., May, R.M., Godfray, H.C., Dunne, J.A., Eds.; Oxford University Press: Oxford, UK, 2012; p. 301. [Google Scholar]
- Castaño, C.; Camarero, J.J.; Zas, R.; Sampedro, L.; Bonet, J.A.; Alday, J.G.; Oliva, J. Insect defoliation is linked to a decrease in soil ectomycorrhizal biomass and shifts in needle endophytic communities. Tree Physiol. 2020, 40, 1712–1725. [Google Scholar] [CrossRef]
- Price, P.W.; Denno, R.F.; Eubanks, M.D.; Finke, D.L.; Kaplan, I. Insect Ecology: Behavior, Populations and Communities; Cambridge University Press: Cambridge, MA, USA, 2011. [Google Scholar]
- Balla, A.; Silini, A.; Cherif-Silini, H.; Bouket, A.C.; Moser, W.K.; Nowakowska, J.A.; Oszako, T.; Benia, F.; Belbahri, L. The Threat of Pests and Pathogens and the Potential for Biological Control in Forest Ecosystems. Forests 2021, 12, 1579. [Google Scholar] [CrossRef]
- Freeman, B.E. Ecological and Econoic Entomology: A Global Synthesis; CABI: Boston, MA, USA, 2021; p. 695. [Google Scholar]
- Losey, J.E.; Vaughan, M. The Economic Value of Ecological Services Provided by Insects. BioScience 2006, 56, 311–323. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Grazer | Parasitoids | Plant-Suckers | Xylophages | Mycetophages | Plant-Chewers | Saprophages | Zoophages |
|---|---|---|---|---|---|---|---|
| 0.15 *** | 1.38 ** | 0.57 *** | 2.21 *** | 1.69 *** | 1.97 *** | 3.04 *** | 1.63 *** |
| 2.75 *** | 1.07 ns | 0.91 ns | 1.07 ns | 0.71 ns | 0.96 ns | 0.66 ns | 0.60 *** |
| 0.99 * | 1.03 ** | 1.01 * |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Floren, A.; Linsenmair, K.E.; Müller, T. Diversity and Functional Relevance of Canopy Arthropods in Central Europe. Diversity 2022, 14, 660. https://doi.org/10.3390/d14080660
Floren A, Linsenmair KE, Müller T. Diversity and Functional Relevance of Canopy Arthropods in Central Europe. Diversity. 2022; 14(8):660. https://doi.org/10.3390/d14080660
Chicago/Turabian StyleFloren, Andreas, Karl Eduard Linsenmair, and Tobias Müller. 2022. "Diversity and Functional Relevance of Canopy Arthropods in Central Europe" Diversity 14, no. 8: 660. https://doi.org/10.3390/d14080660