
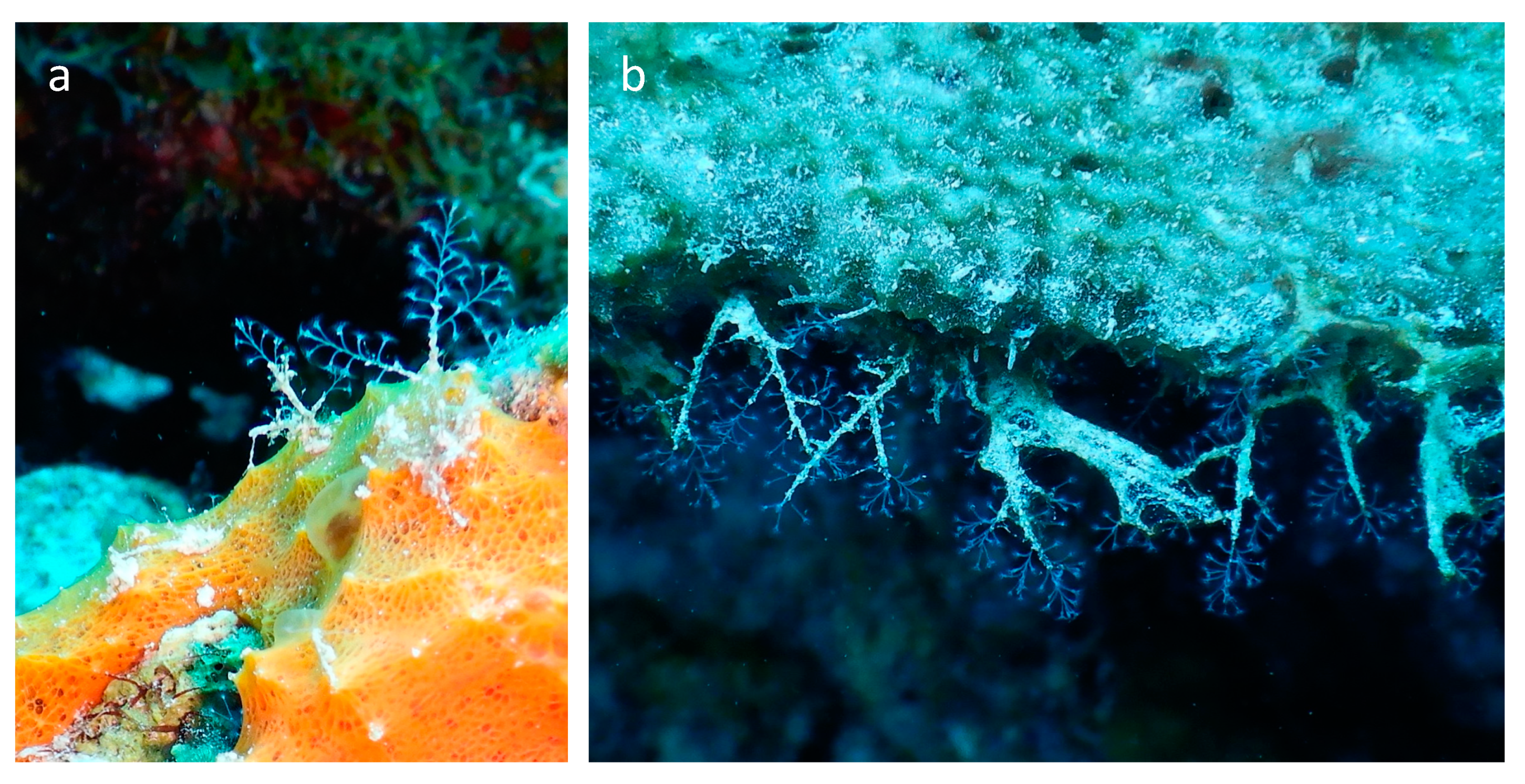
Spatial Ecology of the Association between Demosponges and Nemalecium lighti at Bonaire, Dutch Caribbean

 ,
,  ,
,  and
and
Abstract
:1. Introduction
2. Materials and Methods
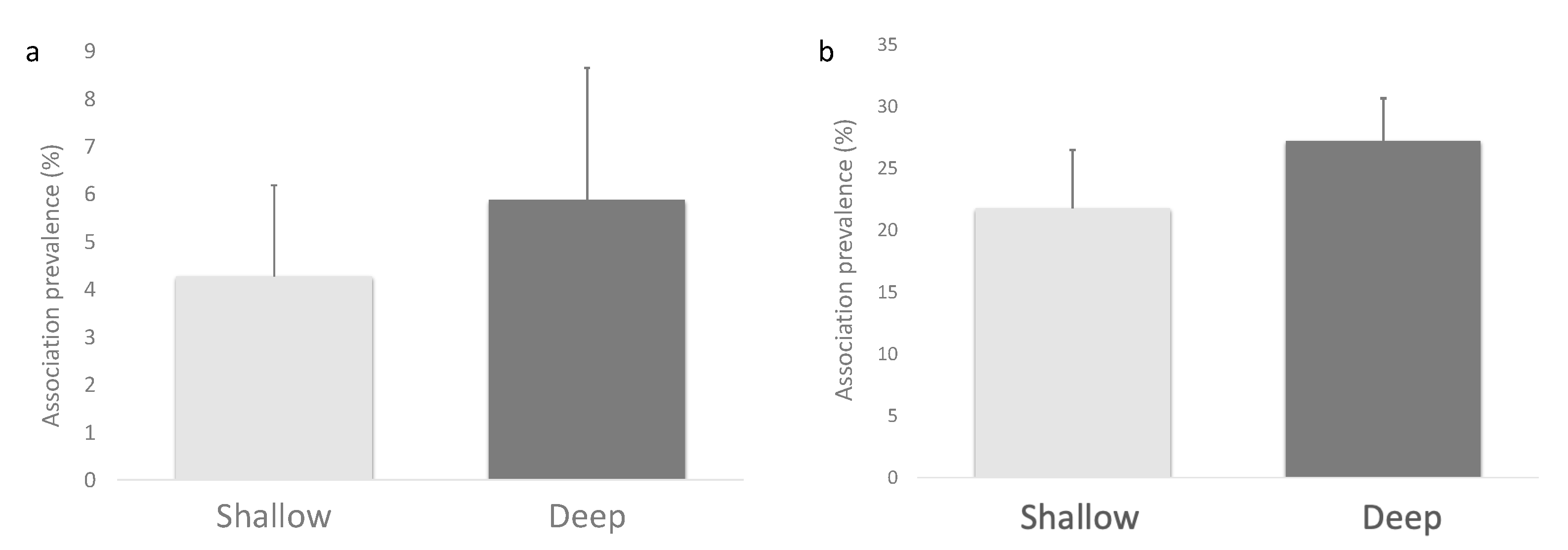
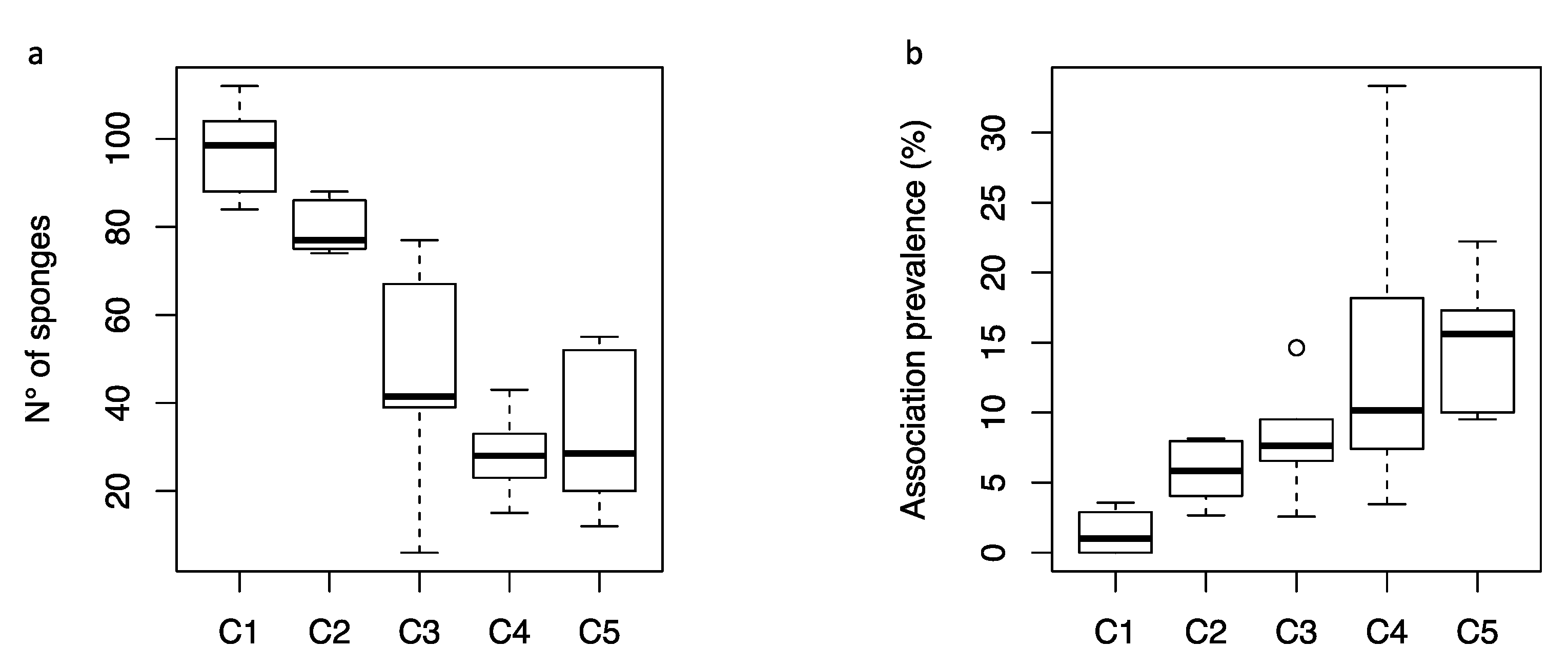
3. Results
4. Discussion
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Fisher, R.; O’Leary, R.A.; Low-Choy, S.; Mengersen, K.; Knowlton, N.; Brainard, R.E.; Caley, M.J. Species richness on coral reefs and the pursuit of convergent global estimates. Curr. Biol. 2015, 25, 500–505. [Google Scholar] [CrossRef] [Green Version]
- Stella, J.S.; Jones, G.P.; Pratchett, M.S. Variation in the structure of epifaunal invertebrate assemblages among coral hosts. Coral Reefs 2010, 29, 957–973. [Google Scholar] [CrossRef]
- Caley, M.J.; Buckley, K.A.; Jones, G.P. Separating ecological effects of habitat fragmentation, degradation, and loss on coral commensals. Ecology 2001, 82, 3435–3448. [Google Scholar] [CrossRef]
- Montano, S. The Extraordinary Importance of Coral-Associated Fauna. Diversity 2020, 12, 357. [Google Scholar] [CrossRef]
- Spotte, S. Supply of regenerated nitrogen to sea anemones by their symbiotic shrimp. J. Exp. Mar. Bio. Ecol. 1996, 198, 27–36. [Google Scholar] [CrossRef]
- Rouzé, H.; Lecellier, G.; Mills, S.C.; Planes, S.; Berteaux-Lecellier, V.; Stewart, H. Juvenile Trapezia spp. crabs can increase juvenile host coral survival by protection from predation. Mar. Ecol. Prog. Ser. 2014, 515, 151–159. [Google Scholar] [CrossRef]
- Pollock, F.J.; Katz, S.M.; Bourne, D.G.; Willis, B.L. Cymo melanodactylus crabs slow progression of white syndrome lesions on corals. Coral Reefs 2013, 32, 43–48. [Google Scholar] [CrossRef]
- Montano, S.; Fattorini, S.; Parravicini, V.; Berumen, M.L.; Galli, P.; Maggioni, D.; Arrigoni, R.; Seveso, D.; Strona, G. Corals hosting symbiotic hydrozoans are less susceptible to predation and disease. Proc. R. Soc. Biol. Sci. 2017, 284, 20172405. [Google Scholar] [CrossRef] [Green Version]
- Stella, J.S.; Pratchett, M.S.; Hutchings, P.A.; Jones, G.P. Coral-associated invertebrates: Diversity, ecological importance, and vulnerability to disturbance. Oceanogr. Mar. Biol. Annu. Rev. 2011, 49, 43–104. [Google Scholar]
- Bo, M.; Di Camillo, C.G.; Puce, S.; Canese, S.; Giusti, M.; Angiolillo, M.; Bavestrello, G. A tubulariid hydroid associated with anthozoan corals in the Mediterranean Sea. Ital. J. Zool. 2011, 78, 487–496. [Google Scholar] [CrossRef]
- Reimer, J.D.; Wee, H.B.; García-Hernández, J.E.; Hoeksema, B.W. Zoantharia (Anthozoa: Hexacorallia) abundance and associations with Porifera and Hydrozoa across a depth gradient on the west coast of Curaçao. Syst. Biodivers. 2018, 16, 820–830. [Google Scholar] [CrossRef]
- Montano, S.; Arrigoni, R.; Pica, D.; Maggioni, D.; Puce, S. New insights into the symbiosis between Zanclea (cnidaria, hydrozoa) and scleractinians. Zool. Scr. 2015, 44, 92–105. [Google Scholar] [CrossRef]
- Montano, S.; Seveso, D.; Galli, P.; Puce, S.; Hoeksema, B.W. Mushroom corals as newly recorded hosts of the hydrozoan symbiont Zanclea sp. Mar. Biol. Res. 2015, 11, 773–779. [Google Scholar] [CrossRef]
- Montano, S.; Maggioni, D.; Arrigoni, R.; Seveso, D.; Puce, S.; Galli, P. The hidden diversity of Zanclea associated with scleractinians revealed by molecular data. PLoS ONE 2015, 10, e0133084. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Puce, S.; Cerrano, C.; Di Camillo, C.G.; Bavestrello, G. Hydroidomedusae (Cnidaria: Hydrozoa) symbiotic radiation. J. Mar. Biol. Assoc. UK 2008, 88, 1715–1721. [Google Scholar] [CrossRef]
- Puce, S.; Calcinai, B.; Bavestrello, G.; Cerrano, C.; Gravili, C.; Boero, F. Hydrozoa (Cnidaria) symbiotic with Porifera: A review. Mar. Ecol. 2005, 26, 73–81. [Google Scholar] [CrossRef]
- Villamizar, E.; Laughlin, R.A. Fauna Associated with the Sponges Aplysina archeri and Aplysina lacunosa in a Coral Reef of the Archipelago de Los Roques, National Park, Venezuela. In Fossil and Recent Sponges, 1st ed.; Reitner, J., Keupp, H., Eds.; Springer: Berlin/Heidelberg, Germany, 1991; pp. 522–542. [Google Scholar] [CrossRef]
- IUCN. Coral Reef Resilience Assessment of the Bonaire National Marine Park. Netherlands Antilles. 2011. Available online: https://data.iucn.org/dbtw-wpd/edocs/2011-008.pdf (accessed on 23 May 2022).
- Frade, P.R.; Bongaerts, P.; Baldwin, C.C.; Trembanis, A.C.; Bak, R.P.M.; Vermeij, M.J.A. Bonaire and Curaçao. In Mesophotic Coral Ecosystems, 1st ed.; Loya, Y., Puglise, K., Bridge, T., Eds.; Series Coral Reef of the World; Springer: Cham, Switzerland, 2019; pp. 149–162. [Google Scholar] [CrossRef]
- Montano, S.; Allevi, V.; Seveso, D.; Maggioni, D.; Galli, P. Habitat preferences of the Pteroclava krempfi-alcyonaceans symbiosis: Inner vs outer coral reefs. Symbiosis 2017, 72, 225–231. [Google Scholar] [CrossRef]
- García-Hernández, J.E.; van Moorsel, G.W.N.M.; Hoeksema, B.W. Lettuce corals overgrowing tube sponges at St. Eustatius, Dutch Caribbean. Mar. Biodivers. 2017, 47, 55–56. [Google Scholar] [CrossRef] [Green Version]
- García-Hernández, J.E.; de Gier, W.; van Moorsel, G.W.N.M.; Hoeksema, B.W. The scleractinian Agaricia undata as a new host for the coral-gall crab Opecarcinus hypostegus at Bonaire, southern Caribbean. Symbiosis 2020, 81, 303–311. [Google Scholar] [CrossRef]
- Garcia-Hernandez, J.E.; Reimer, J.D.; Hoeksema, B.W. Sponges hosting the zoantharia-associated crab Platypodiella spectabilis at St. Eustatius, Dutch Caribbean. Coral Reefs 2016, 35, 209–210. [Google Scholar] [CrossRef] [Green Version]
- García-Hernández, J.E.; Hoeksema, B.W. Sponges as secondary hosts for Christmas tree worms at Curaçao. Coral Reefs 2017, 36, 1243–1244. [Google Scholar] [CrossRef]
- Hoeksema, B.W.; García-Hernández, J.E.; van Moorsel, G.W.N.M.; Olthof, G.; ten Hove, H.A. Extension of the recorded host range of Caribbean Christmas tree worms (Spirobranchus spp.) with two scleractinians, a zoantharian, and an ascidian. Diversity 2020, 12, 115. [Google Scholar] [CrossRef] [Green Version]
- Montano, S.; Reimer, J.D.; Ivanenko, V.N.; García-Hernández, J.E.; van Moorsel, G.W.N.M.; Galli, P.; Hoeksema, B.W. Widespread occurrence of a rarely known association between the hydrocorals Stylaster roseus and Millepora alcicornis at Bonaire, Southern Caribbean. Diversity 2020, 12, 218. [Google Scholar] [CrossRef]
- Gravier-Bonnet, N.; Bourmaud, C.A.F. Hydroids (Cnidaria, Hydrozoa) of Coral Reefs: Preliminary Results on Community Structure, Species Distribution and Reproductive Biology in Juan de Nova Island (Southwest Indian Ocean). West. Indian Ocean J. Mar. Sci. 2006, 5, 123–132. [Google Scholar] [CrossRef] [Green Version]
- Coma, R.; Ribes, M.; Orejas, C.; Gili, J.M. Prey capture by a benthic coral reef hydrozoan. Coral Reefs 1999, 18, 141–145. [Google Scholar] [CrossRef]
- Di Camillo, C.G.; Bavestrello, G.; Valisano, L.; Puce, S. Spatial and temporal distribution in a tropical hydroid assemblage. J. Mar. Biol. Assoc. UK 2008, 88, 1589–1599. [Google Scholar] [CrossRef]
- Bouillon, J.; Gravili, C.; Pagès, F.; Gili, J.M.; Boero, F. An Introduction to Hydrozoa, 1st ed.; Betsch, J.M., Bouchet, P., Érard, C., Eds.; Mémoires du Muséum national d’histoire naturelle: Paris, France, 2006. [Google Scholar]
- Lewis, T.B.; Finelli, C.M. Epizoic zoanthids reduce pumping in two Caribbean vase sponges. Coral Reefs 2015, 34, 291–300. [Google Scholar] [CrossRef]
- Puyana, M.; Fenical, W.; Pawlik, J.R. Are there activated chemical defenses in sponges of the genus Aplysina from the Caribbean? Mar. Ecol. Prog. Ser. 2003, 246, 127–135. [Google Scholar] [CrossRef] [Green Version]
- Wulff, J.L. Ecological interactions of marine sponges. Can. J. Zool. 2006, 84, 146–166. [Google Scholar] [CrossRef]
- Padua, A.; Lanna, E.; Klautau, M. Macrofauna inhabiting the sponge Paraleucilla magna (Porifera: Calcarea) in Rio de Janeiro, Brazil. J. Mar. Biol. Assoc. UK 2013, 93, 889–898. [Google Scholar] [CrossRef] [Green Version]
- Zea, S.; Henkel, T.P.; Pawlik, J.R. The Sponge Guide: A Picture Guide to Caribbean Sponges. Available online: https://www.spongeguide.org (accessed on 21 January 2022).
- Calder, D.R. Local distribution and biogeography of the hydroids (Cnidaria) of Bermuda. Caribb. J. Sci. 1993, 29, 61–74. [Google Scholar]
- Calder, D.R. Associations between hydroid species assemblages and substrate types in the mangal at Twin Cays, Belize. Can. J. Zool. 1991, 69, 2067–2074. [Google Scholar] [CrossRef]
- Calder, D.R. Abundance and distribution of hydroids in a mangrove ecosystem at Twin Cays, Belize, Central America. Hydrobiologia 1991, 216, 221–228. [Google Scholar] [CrossRef]
- Diaz, M.C.; Rützler, K. Sponges: An essential component of Caribbean coral reefs. Bull. Mar. Sci. 2001, 69, 535–546. [Google Scholar]
- Hargitt, C.M. Hydroids of the Philippine Islands. Philipp. J. Sci. 1924, 24, 467–507. Available online: https://www.marinespecies.org/aphia.php?p=sourcedetails&id=24462 (accessed on 14 July 2022).
- Amelia, M.; Suaberon, F.A.; Vad, J.; Fahmi, A.D.M.; Saludes, J.P.; Bhubalan, K. Recent Advances of Marine Sponge—Associated Microorganisms as a Source of Commercially Viable Natural Products. Mar. Biotechnol. 2022, 24, 492–512. [Google Scholar] [CrossRef]
- Hentschel, U.; Schmid, M.; Wagner, M.; Fieseler, L.; Gernert, C. Isolation and phylogenetic analysis of bacteria with antimicrobial activities from the Mediterranean sponges Aplysina aerophoba and Aplysina cavernicola. FEMS Microbiol. Ecol. 2001, 35, 305–312. [Google Scholar] [CrossRef]
- Thoms, C.; Ebel, R.; Proksch, P. Activated chemical defense in Aplysina sponges revisited. J. Chem. Ecol. 2006, 32, 97–123. [Google Scholar] [CrossRef]
- Assmann, M.; Lichte, E.; Pawlik, J.R.; Köck, M. Chemical defenses of the Caribbean sponges Agelas wiedenmayeri and Agelas conifera. Mar. Ecol. Prog. Ser. 2000, 207, 255–262. [Google Scholar] [CrossRef] [Green Version]
- Ebel, R.; Brenzinger, M.; Kunze, A.; Gross, H.J.; Proksch, P. Wound activation of protoxins in marine sponge Aplysina aerophoba. Can. Field-Nat. 1997, 111, 1451–1462. [Google Scholar] [CrossRef]
- Weiss, B.; Ebel, R.; Elbrächter, M.; Kirchner, M.; Proksch, P. Defense metabolites from the marine sponge Verongia aerophoba. Biochem. Syst. Ecol. 1996, 24, 1–7. [Google Scholar] [CrossRef]
- Teeypant, R.; Woerdenbang, H.J.; Gross, H.J.; Proksch, P. Biotransformation of the Brominated Compounds in the Marine Sponge Verongia aerophoba: Evidence for an Induced Chemical Defense? Planta Med. 1993, 59, A641–A642. [Google Scholar] [CrossRef]
- Thoms, C.; Wolff, M.; Padmakumar, K.; Ebel, R.; Proksch, P. Chemical Defense of Mediterranean Sponges Aplysina cavernicola and Aplysina aerophoba. Z. Fur Naturforsch. 2004, 59, 113–122. [Google Scholar] [CrossRef] [PubMed]
- Chanas, B.; Pawlik, J.R.; Lindel, T.; Fenical, W. Chemical defense of the Caribbean sponge Agelas clathrodes (Schmidt). J. Exp. Mar. Bio. Ecol. 1997, 208, 185–196. [Google Scholar] [CrossRef]
- Wulff, J.L. Resistance vs recovery: Morphological strategies of coral reef sponges. Funct. Ecol. 2006, 20, 699–708. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
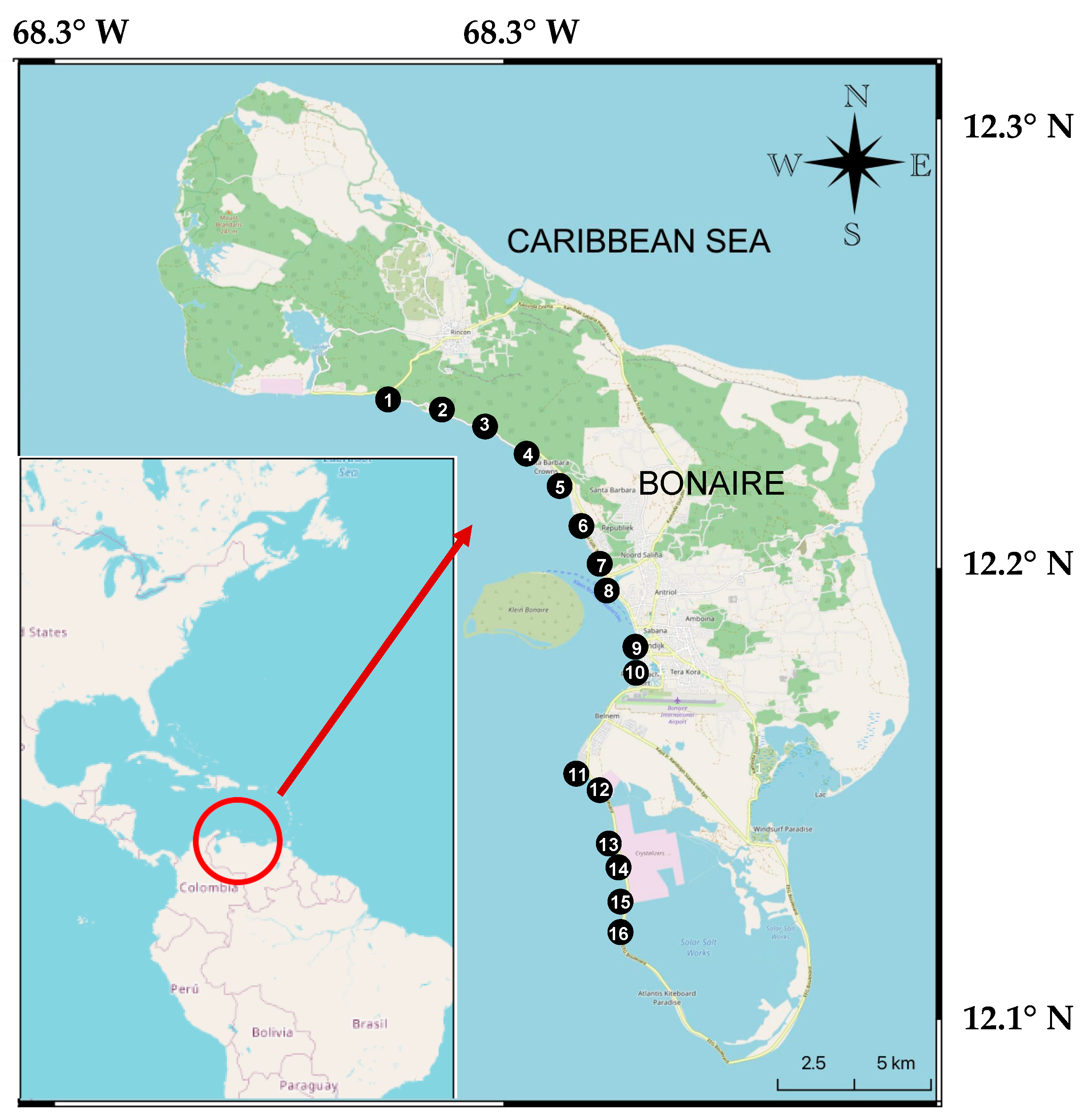
| N° | Dive Sites | Coordinates | Maximum Prevalence (%) | Mean Prevalence (% ± SE) |
|---|---|---|---|---|
| 1 | Tolo Reef (Tol) | 12°12′92″ N; 068°20′22″ W | 15.38 | 4.89 ± 2.68 |
| 2 | Jeff Davies Memorial (JDM) | 12°12′18″ N; 068°18′50″ W | 8.33 | 2.67 ± 1.69 |
| 3 | Oil Slick Leap (OSL) | 12°12′03″ N; 068°18′51″ W | 5.55 | 0.93 ± 0.93 |
| 4 | Andrea I (AI) | 12°11′29″ N; 068°17′80″ W | 12.50 | 2.08 ± 2.08 |
| 5 | Andrea II (AII) | 12°11′18″ N; 068°17′48″ W | 13.33 | 2.22 ± 2.22 |
| 6 | La Machaca (LM) | 12°10′20″ N; 068°17′22″ W | 17.86 | 8.67 ± 2.73 |
| 7 | Buddy’s Reef (BM) | 12°10′14″ N; 068°17′18″ W | 16.67 | 4.01 ± 2.81 |
| 8 | Bari Reef (BF) | 12°10′04″ N; 068°17′10″ W | 33.33 | 20.82 ± 3.56 |
| 9 | Something Special (SS) | 12°09′70″ N; 068°17′02″ W | 3.70 | 2.12 ± 0.70 |
| 10 | Town Pier (TP) | 12°08′57″ N; 068°16′40″ W | 2.63 | 0.44 ± 0.44 |
| 11 | Punt Viekant (PV) | 12°06′91″ N; 068°17′66″ W | 0.00 | 0.00 ± 0.00 |
| 12 | Alice in Wonderland (AiW) | 12°05′99″ N; 068°17′12″ W | 22.22 | 17.59 ± 2.31 |
| 13 | Salt Pier (SP) | 12°05′01″ N; 068°16′91″ W | 9.52 | 4.37 ± 2.01 |
| 14 | Invisibles (I) | 12°04′65″ N; 068°16′80″ W | 17.65 | 2.94 ± 2.94 |
| 15 | Tori Reef (Tor) | 12°04′25″ N; 068°16′84″ W | 10.00 | 1.67 ± 1.67 |
| 16 | Pink Beach (PB) | 12°03′85″ N; 068°16′90″ W | 11.11 | 5.56 ± 1.87 |
| Genus | Relative Abundance (%) | Maximum Prevalence (%) | Mean Prevalence (% ± SE) |
|---|---|---|---|
| Agelas | 13.16 | 7.27 | 2.70 ± 1.12 |
| Aiolochroia | 9.46 | 8.62 | 4.09 ± 1.69 |
| Aplysina | 14.30 | 36.17 | 26.06 ± 4.76 |
| Callyspongia | 3.13 | 0.00 | 0.00 ± 0.00 |
| Clathria | 0.23 | 0.00 | 0.00 ± 0.00 |
| Cliona | 0.40 | 25.00 | 5.00 ± 4.56 |
| Desmapsamma | 4.05 | 8.69 | 2.74 ± 1.62 |
| Ectyoplasia | 3.82 | 0.00 | 0.00 ± 0.00 |
| Halisarca | 0.17 | 0.00 | 0.00 ± 0.00 |
| Iotrochota | 4.73 | 4.00 | 0.80 ± 0.73 |
| Ircinia | 6.38 | 5.88 | 2.81 ± 1.01 |
| Monanchora | 0.46 | 0.00 | 0.00 ± 0.00 |
| Niphates | 6.84 | 0.00 | 0.00 ± 0.00 |
| Phorbas | 0.11 | 0.00 | 0.00 ± 0.00 |
| Scopalina | 26.67 | 4.11 | 1.78 ± 0.76 |
| Verongula | 6.09 | 15.38 | 6.98 ± 2.34 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gobbato, J.; Magrini, A.; García-Hernández, J.E.; Virdis, F.; Galli, P.; Seveso, D.; Montano, S. Spatial Ecology of the Association between Demosponges and Nemalecium lighti at Bonaire, Dutch Caribbean. Diversity 2022, 14, 607. https://doi.org/10.3390/d14080607
Gobbato J, Magrini A, García-Hernández JE, Virdis F, Galli P, Seveso D, Montano S. Spatial Ecology of the Association between Demosponges and Nemalecium lighti at Bonaire, Dutch Caribbean. Diversity. 2022; 14(8):607. https://doi.org/10.3390/d14080607
Chicago/Turabian StyleGobbato, Jacopo, Andrea Magrini, Jaaziel E. García-Hernández, Francesca Virdis, Paolo Galli, Davide Seveso, and Simone Montano. 2022. "Spatial Ecology of the Association between Demosponges and Nemalecium lighti at Bonaire, Dutch Caribbean" Diversity 14, no. 8: 607. https://doi.org/10.3390/d14080607