
Investigation of the Genetic Diversity of Dagestan Mountain Cattle Using STR-Markers

 ,
,  , , and
, , and 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Sample Collection
2.2. DNA Extraction and STR-Genotyping
2.3. Data Analysis
3. Results
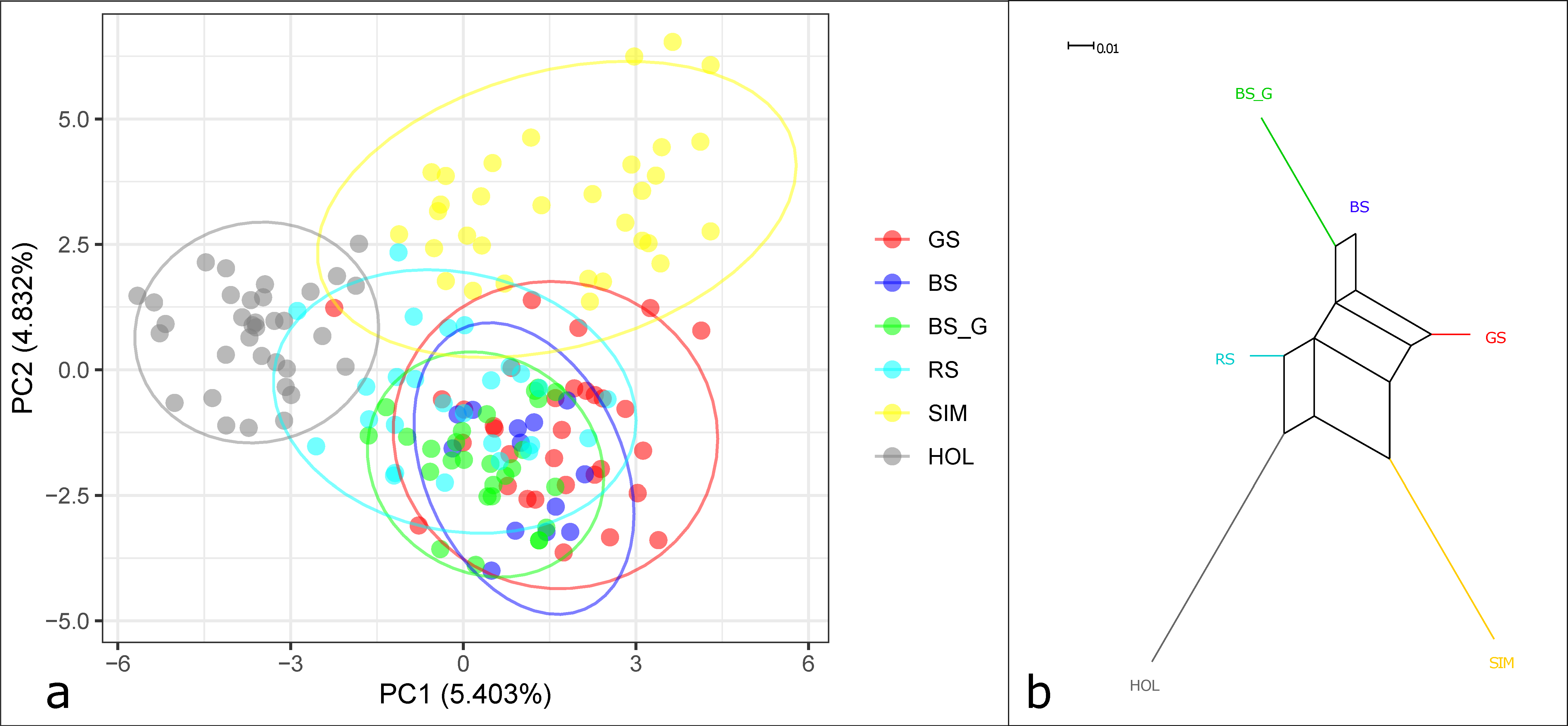
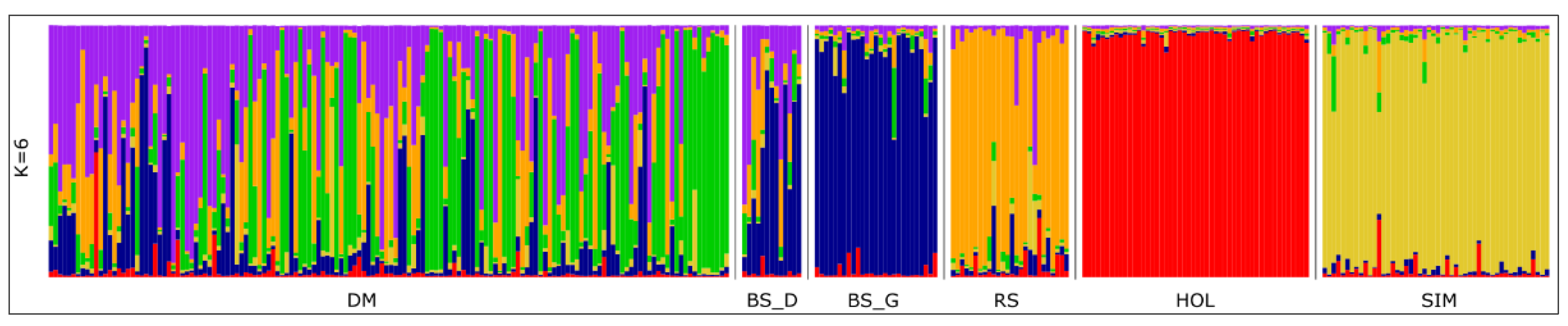
3.1. Genetic Variation among and within Breeds
3.2. Genetic Differentiation
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Ibragimov, R.E.; Dzhalalov, A.P.; Alilov, M.M. Outlook for the development to mountain beef cattle breeding in Dagestan. Collect. Sci. Pap. North Cauc. Res. Inst. Anim. Husb. 2013, 2, 68–74. [Google Scholar]
- Nuratinov, D.A. Traditional Forms of Management in Dagestan. Issues Struct. Econ. 2008, 4, 22–29. [Google Scholar]
- Dunin, I.M.; Dankvert, A.G. Directory of Breeds and Types of Farm Animals Bred in the Russian Federation; VNIIPLEM: Moscow, Russia, 2013; 560p. [Google Scholar]
- Petherbridge, G.; Ismailov, M.M.; Ismailov, S.M.; Rabadanov, M.K.; Gadzhiev, A.A.; Teymurov, A.A.; Rabadanov, M.R.; Daudova, M.G.; Abdulaev, A.; Verkhnee Gakvari, G.M. The contribution of adat, religious beliefs and public education to collective environmental management in an agro-pastoral community in the Dagestan high Caucasus. South Russ. Ecol. Dev. 2021, 16, 142–179. [Google Scholar] [CrossRef]
- Population United Nations. Available online: https://www.un.org/en/global-issues/population (accessed on 6 June 2022).
- Aligazieva, P.A.; Aligaziev, A.M.; Alidibirov, A.T. Influence of Factors on Milk Productivity of First-Heifer Heifers of Mountain Cattle. In The Current State and Main Directions of Development of Seed Production in the Republic of Dagestan: Materials of the All-Russian Scientific and Practical Conference; Publisher Federal State Budgetary Educational Institution of Higher Education “Dagestan State Agrarian University named after M.M. Dzhambulatov”: Makhachkala, Russia, 2019; pp. 119–125. [Google Scholar]
- Sadykov, M.M. Ways to increase the meat productivity of mountain cattle. Mt. Agric. 2016, 3, 167–170. [Google Scholar]
- UNU. Available online: https://www.vij.ru/infrastruktura/unu (accessed on 20 February 2020).
- FAO. Molecular Genetic Characterization of Animal Genetic Resources; FAO: Rome, Italy, 2011; pp. 68–69. Available online: http://www.fao.org/3/i2413e/i2413e00.pdf (accessed on 6 June 2022).
- Bishop, M.D.; Kappes, S.M.; Keele, J.W.; Stone, R.T.; Sunden, S.L.; Hawkins, G.A.; Toldo, S.S.; Fries, R.; Grosz, M.D.; Yoo, J.; et al. A genetic linkage map for cattle. Genetics 1994, 136, 619–639. [Google Scholar] [CrossRef]
- Toldo, S.S.; Fries, R.; Steffen, P.; Neiberg, H.L.; Barendse, W.; Womack, J.E.; Hetzel, D.J.S.; Stranzinger, G. Physically mapped, cosmid-derived microsatellite markers as anchor loci on bovine chromosomes. Mamm. Genome 1993, 4, 720–727. [Google Scholar] [CrossRef]
- Steffen, P.; Eggen, A.; Stranzinger, G.; Fries, R.; Dietz, A.B.; Womack, J.E. Isolation and mapping of polymorphic microsatellites in cattle. Anim. Genet. 2009, 24, 121–124. [Google Scholar] [CrossRef]
- Vaiman, D.; Mercier, D.; Moazami-Goudarzi, K.; Eggen, A.; Ciampolini, R.; Lepingle, A.; Velmala, R.; Kaukinen, J.; Varvio, S.L.; Martin, P.; et al. A set of 99 cattle microsatellites: Characterization, synteny mapping, and polymorphism. Mamm. Genome 1994, 5, 288–297. [Google Scholar] [CrossRef]
- Moore, S.S.; Byrne, K.; Berger, K.T.; Barendse, W.; McCarthy, F.; Womack, J.E.; Hetzel, D.J.S. Characterization of 65 bovine microsatellites. Mamm. Genome 1994, 5, 84–90. [Google Scholar] [CrossRef]
- Barendse, W.; Armitage, S.M.; Kossarek, L.M.; Shalom, A.; Kirkpatrick, B.W.; Ryan, A.M.; Clayton, D.; Li, L.; Neibergs, H.L.; Zhang, N.; et al. A genetic linkage map of the bovine genome. Nat. Genet. 1994, 6, 227–235. [Google Scholar] [CrossRef]
- Zinovieva, N.A.; Popov, A.N.; Ernst, L.K. Guidelines for the Use of the Polymerase Chain Reaction Method in Animal Husbandry; VIJ: Dubrovitsy, Russia, 1998; 47p. [Google Scholar]
- Peakall, R.; Smouse, P.E. GenAlEx 6.5: Genetic analysis in Excel. Population genetic software for teaching and research-an update. Bioinformatics 2012, 28, 2537–2539. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Keenan, K.; McGinnity, P.; Cross, T.F.; Crozier, W.W.; Prodöhl, P.A. diveRsity: An R package for the estimation and exploration of population genetics parameters and their associated errors. Methods Ecol. Evol. 2013, 4, 782–788. [Google Scholar] [CrossRef] [Green Version]
- Weir, B.S.; Cockerham, C.C. Estimating F-Statistics for the Analysis of Population Structure. Evolution 1984, 38, 1358–1370. [Google Scholar] [CrossRef] [PubMed]
- Jost, L. GST and its relatives do not measure differentiation. Mol. Ecol. 2008, 17, 4015. [Google Scholar] [CrossRef]
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2018; Available online: https://www.R-project.org/ (accessed on 23 July 2020).
- Huson, D.H.; Bryant, D. Application of Phylogenetic Networks in Evolutionary Studies. Mol. Biol. Evol. 2006, 23, 254–267. [Google Scholar] [CrossRef]
- Jombart, T. Adegenet: A R package for the multivariate analysis of genetic markers. Bioinformatics 2008, 24, 1403–1405. [Google Scholar] [CrossRef] [Green Version]
- Wickham, H. Ggplot2: Elegant Graphics for Data Analysis (Use R!); Springer: New York, NY, USA, 2009; ISBN 978-0-387-98140-6. [Google Scholar]
- Becker, R.A.; Wilks, A.R.; Brownrigg, R.; Minka, T.P.; Deckmyn, A. Maps: Draw Geographical Maps. R Package Version 3.3.0. 2018. Available online: https://CRAN.R-project.org/package=maps (accessed on 22 December 2020).
- Pritchard, J.K.; Stephens, M.; Donnelly, P. Inference of population structure using multilocus genotype data. Genetics 2000, 155, 945–959. [Google Scholar] [CrossRef]
- Kopelman, N.M.; Mayzel, J.; Jakobsson, M.; Rosenberg, N.A.; Mayrose, I. CLUMPAK: A program for identifying clustering modes and packaging population structure inferences across K. Mol. Ecol. Resour. 2015, 15, 1179–1191. [Google Scholar] [CrossRef] [Green Version]
- CLUMPAK—Clustering Markov Packager across K. Available online: http://clumpak.tau.ac.il (accessed on 23 May 2022).
- Evanno, G.; Regnaut, S.; Goudet, J. Detecting the number of clusters of individuals using the software STRUCTURE: A simulation study. Mol. Ecol. 2005, 14, 2611–2620. [Google Scholar] [CrossRef] [Green Version]
- Hartl, D.L.; Clark, A.G. Principles of Population Genetics; Sinauer Associates: Sunderland, UK, 1997. [Google Scholar]
- Dmitriev, N.G.; Ernst, L.K. Animal Genetic Resources of the USSR; FAO: Rome, Italy, 1989; Available online: http://www.fao.org/3/ah759e/AH759E07.htm (accessed on 25 May 2022).
- Wang, C.; Reinhold, S.; Kalmykov, A.; Wissgott, A.; Brandt, G.; Jeong, C.; Cheronet, O.; Ferry, M.; Harney, E.; Keating, D.; et al. Ancient human genome-wide data from a 3000-year interval in the Caucasus corresponds with eco-geographic regions. Nat. Commun. 2019, 10, 590. [Google Scholar] [CrossRef] [Green Version]
- Yunusbayev, B.; Metspalu, M.; Järve, M.; Kutuev, I.; Rootsi, S.; Metspalu, E.; Behar, D.M.; Varendi, K.; Sahakyan, H.; Khusainova, R.; et al. The Caucasus as an asymmetric semipermeable barrier to ancient human migrations. Mol. Biol. Evol. 2012, 29, 359–365. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Caucasian Cattle. (n.d.) The Great Soviet Encyclopedia. 3rd Edition. (1970–1979). Available online: https://encyclopedia2.thefreedictionary.com/Caucasian+Cattle (accessed on 23 July 2020).
- Porter, V.; Alderson, L.; Hall, S.J.; Sponenberg, D.P. Mason’s World Encyclopedia of Livestock Breeds and Breeding; CABI: Wallingford, UK, 2016; Volume 1–2. [Google Scholar]
- Abdelmanova, A.S.; Kharzinova, V.R.; Volkova, V.V.; Mishina, A.I.; Dotsev, A.V.; Sermyagin, A.A.; Boronetskaya, O.I.; Petrikeeva, L.V.; Chinarov, R.Y.; Brem, G.; et al. Genetic Diversity of Historical and Modern Populations of Russian Cattle Breeds Revealed by Microsatellite Analysis. Genes 2020, 11, 940. [Google Scholar] [CrossRef] [PubMed]
- Abdelmanova, A.S.; Kharzinova, V.R.; Volkova, V.V.; Dotsev, A.V.; Sermyagin, A.A.; Boronetskaya, O.I.; Chinarov, R.Y.; Lutshikhina, E.M.; Sölkner, J.; Brem, G.; et al. Comparative study of the genetic diversity of local steppe cattle breeds from Russia, Kazakhstan and Kyrgyzstan by microsatellite analysis of museum and modern samples. Diversity 2021, 13, 351. [Google Scholar] [CrossRef]
- Sermyagin, A.A.; Dotsev, A.V.; Gladyr, E.A.; Traspov, A.A.; Deniskova, T.E.; Kostyunina, O.V.; Reyer, H.; Wimmers, K.; Barbato, M.; Paronyan, I.A.; et al. Whole-genome SNP analysis elucidates the genetic structure of Russian cattle and its relationship with eurasian taurinebreeds. Genet. Sel. Evol. 2018, 50, 1. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yurchenko, A.; Yudin, N.; Aitnazarov, R.; Plyusnina, A.; Brukhin, V.; Soloshenko, V.; Lhasaranov, B.; Popov, R.; Paronyan, I.A.; Plemyashov, K.V.; et al. Genome-wide genotyping uncovers genetic profiles and history of the Russian cattle breeds. Heredity 2018, 120, 125–137. [Google Scholar] [CrossRef]
- Zinovieva, N.A.; Dotsev, A.V.; Sermyagin, A.A.; Deniskova, T.E.; Abdelmanova, A.S.; Kharzinova, V.R.; Sölkner, J.; Reyer, H.; Wimmers, K.; Brem, G. Selection signatures in two oldest Russian native cattle breeds revealed using high-density single nucleotide polymorphism analysis. PLoS ONE 2020, 15, e0242200. [Google Scholar] [CrossRef]
- Zinovieva, N.A.; Sheiko, I.P.; Dotsev, A.V.; Sheiko, R.I.; Mikhailova, M.E.; Sermyagin, A.A.; Abdelmanova, A.S.; Kharzinova, V.R.; Reyer, H.; Wimmers, K.; et al. Genome-wide SNP analysis clearly distinguished the Belarusian Red cattle from other European cattle breeds. Anim. Genet. 2021, 52, 720–724. [Google Scholar] [CrossRef]
- Sharma, R.; Maitra, A.; Singh, P.K.; Tantia, M.S. Genetic diversity and relationship of cattle populations of East India: Distinguishing lesser known cattle populations and established breeds based on STR markers. SpringerPlus 2013, 2, 359. [Google Scholar] [CrossRef] [Green Version]
- Ginja, C.; Gama, L.T.; Cortes, Ó.; Delgado, J.V.; Dunner, S.; García, D.; Landi, V.; Martín-Burriel, I.; Martínez-Martínez, A.; Penedo, M.C.T.; et al. Analysis of conservation priorities of Iberoamerican cattle based on autosomal microsatellite markers. Genet. Sel. Evol. 2013, 45, 35. [Google Scholar] [CrossRef] [Green Version]
- Tu, P.A.; Lin, D.Y.; Li, G.F.; Huang, J.C.; Wang, D.C.; Wang, P.H. Characterization of the genetic diversity and population structure for the yellow cattle in Taiwan based on microsatellite markers. Anim. Biotechnol. 2014, 25, 234–249. [Google Scholar] [CrossRef]
- Ginja, C.; Gama, L.T.; Penedo, M.C. Analysis of STR markers reveals high genetic structure in Portuguese native cattle. J. Hered. 2010, 101, 201–210. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Laoun, A.; Harkat, S.; Lafri, M.; Bechir, S.; Gaouar, S.; Belabdi, I.; Ciani, E.; De Groot, M.; Blanquet, V.; Leroy, G.; et al. Inference of Breed Structure in Farm Animals: Empirical Comparison between SNP and Microsatellite Performance. Genes 2020, 11, 57. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Martín-Burriel, I.; Rodellar, C.; Cañón, J.; Cortés, O.; Dunner, S.; Landi, V.; Martínez-Martínez, A.; Gama, L.T.; Ginja, C.; Penedo, M.C.; et al. Genetic diversity, structure, and breed relationships in Iberian cattle. J. Anim. Sci. 2011, 89, 893–906. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ndiaye, N.P.; Sow, A.; Dayo, G.K.; Ndiaye, S.; Sawadogo, G.J.; Sembène, M. Genetic diversity and phylogenetic relationships in local cattle breeds of Senegal based on autosomal microsatellite markers. Vet. World 2015, 8, 994–1005. [Google Scholar] [CrossRef] [PubMed] [Green Version]




{kind=link}
{kind=link}
{kind=link}
| Breed | Code | n | Breeding Region |
|---|---|---|---|
| Dagestan Mountain | DM | 150 | The Republic of Dagestan |
| Brown Swiss, Caucasian branch | BS_D | 13 | The Republic of Dagestan |
| Brown Swiss, Germany | BS_G | 27 | Germany |
| Red Steppe | RS | 26 | The Republic of Dagestan |
| Simmental | SIM | 50 | Oryol region |
| Holstein 1 | HOL | 50 | Holstein Association USA, Inc. |
| Locus | Population | In Total 1 | |||||
|---|---|---|---|---|---|---|---|
| DM | BS_D | BS_G | RS | HOL | SIM | ||
| TGLA227 | 12/1 | 7 | 6 | 9 | 7/1 | 8 | 49/2 |
| BM2113 | 11/4 | 6 | 7 | 6 | 5 | 5 | 40/4 |
| TGLA53 | 17/2 | 9 | 9 | 8 | 7 | 11 | 61/2 |
| ETH10 | 8/1 | 5/1 | 4 | 5 | 6 | 5 | 33/2 |
| SPS115 | 9/2 | 4 | 5 | 6 | 4 | 6 | 34/2 |
| TGLA122 | 17/6 | 6 | 8/2 | 7/1 | 7/1 | 9 | 54/10 |
| INRA23 | 11/1 | 7 | 5 | 8 | 4 | 8/1 | 43/2 |
| TGLA126 | 8/3 | 4 | 3 | 4 | 4 | 6/1 | 29/4 |
| BM1818 | 8 | 5 | 6 | 3 | 4 | 6 | 32/0 |
| ETH225 | 11/4 | 7/1 | 5 | 7 | 5 | 6 | 41/5 |
| BM1824 | 6/1 | 4 | 4 | 4 | 3 | 5 | 26/1 |
| Total | 118/25 | 64/2 | 62/2 | 67/1 | 56/2 | 75/2 | 442/34 |
| Population | n 1 | Ar 2 (M ± SE) 7 | Ho 3 (M ± SE) | uHe 4 (M ± SE) | uFis 5 [CI] 6 |
|---|---|---|---|---|---|
| DM | 150 | 6.827 ± 0.654 | 0.723 ± 0.032 | 0.764 ± 0.031 | 0.052 [0.005; 0.099] |
| BS_D | 13 | 5.818 ± 0.483 | 0.692 ± 0.069 | 0.713 ± 0.055 | 0.023 [−0.093; 0.139] |
| BS_G | 27 | 5.146 ± 0.444 | 0.741 ± 0.045 | 0.701 ± 0.039 | −0.058 [−0.111; −0.005] |
| RS | 26 | 5.278 ± 0.450 | 0.692 ± 0.054 | 0.718 ± 0.026 | 0.046 [−0.059; 0.151] |
| HOL | 50 | 4.376 ± 0.396 | 0.669 ± 0.044 | 0.645 ± 0.039 | −0.038 [−0.110; 0.034] |
| SIM | 50 | 5.307 ± 0.373 | 0.729 ± 0.033 | 0.682 ± 0.031 | −0.073 [−0.115; −0.031] |
| Population | DM | BS_D | BS_G | RS | HOL | SIM |
|---|---|---|---|---|---|---|
| DM | 0 | 0.023 1 | 0.053 1 | 0.036 1 | 0.113 1 | 0.058 1 |
| BS_D | 0.049 2 | 0 | 0.047 1 | 0.049 1 | 0.135 1 | 0.093 1 |
| BS_G | 0.130 2 | 0.054 2 | 0 | 0.081 1 | 0.134 1 | 0.116 1 |
| RS | 0.066 2 | 0.043 2 | 0.150 2 | 0 | 0.105 1 | 0.084 1 |
| HOL | 0.271 2 | 0.261 2 | 0.250 2 | 0.189 2 | 0 | 0.138 1 |
| SIM | 0.114 2 | 0.195 2 | 0.210 2 | 0.132 2 | 0.234 2 | 0 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Volkova, V.V.; Abdelmanova, A.S.; Deniskova, T.E.; Romanenkova, O.S.; Khozhokov, A.A.; Ozdemirov, A.A.; Sermyagin, A.A.; Zinovieva, N.A. Investigation of the Genetic Diversity of Dagestan Mountain Cattle Using STR-Markers. Diversity 2022, 14, 569. https://doi.org/10.3390/d14070569
Volkova VV, Abdelmanova AS, Deniskova TE, Romanenkova OS, Khozhokov AA, Ozdemirov AA, Sermyagin AA, Zinovieva NA. Investigation of the Genetic Diversity of Dagestan Mountain Cattle Using STR-Markers. Diversity. 2022; 14(7):569. https://doi.org/10.3390/d14070569
Chicago/Turabian StyleVolkova, Valeria V., Alexandra S. Abdelmanova, Tatiana E. Deniskova, Olga S. Romanenkova, Abdusalam A. Khozhokov, Alimsoltan A. Ozdemirov, Alexander A. Sermyagin, and Natalia A. Zinovieva. 2022. "Investigation of the Genetic Diversity of Dagestan Mountain Cattle Using STR-Markers" Diversity 14, no. 7: 569. https://doi.org/10.3390/d14070569