
Differential Effects of Tree Species on Soil Microbiota 45 Years after Afforestation of Former Pastures


Abstract
:1. Introduction
2. Materials and Methods
2.1. Study Sites and Soil Sampling
2.2. Laboratory Analyses
2.3. Statistical Analysis
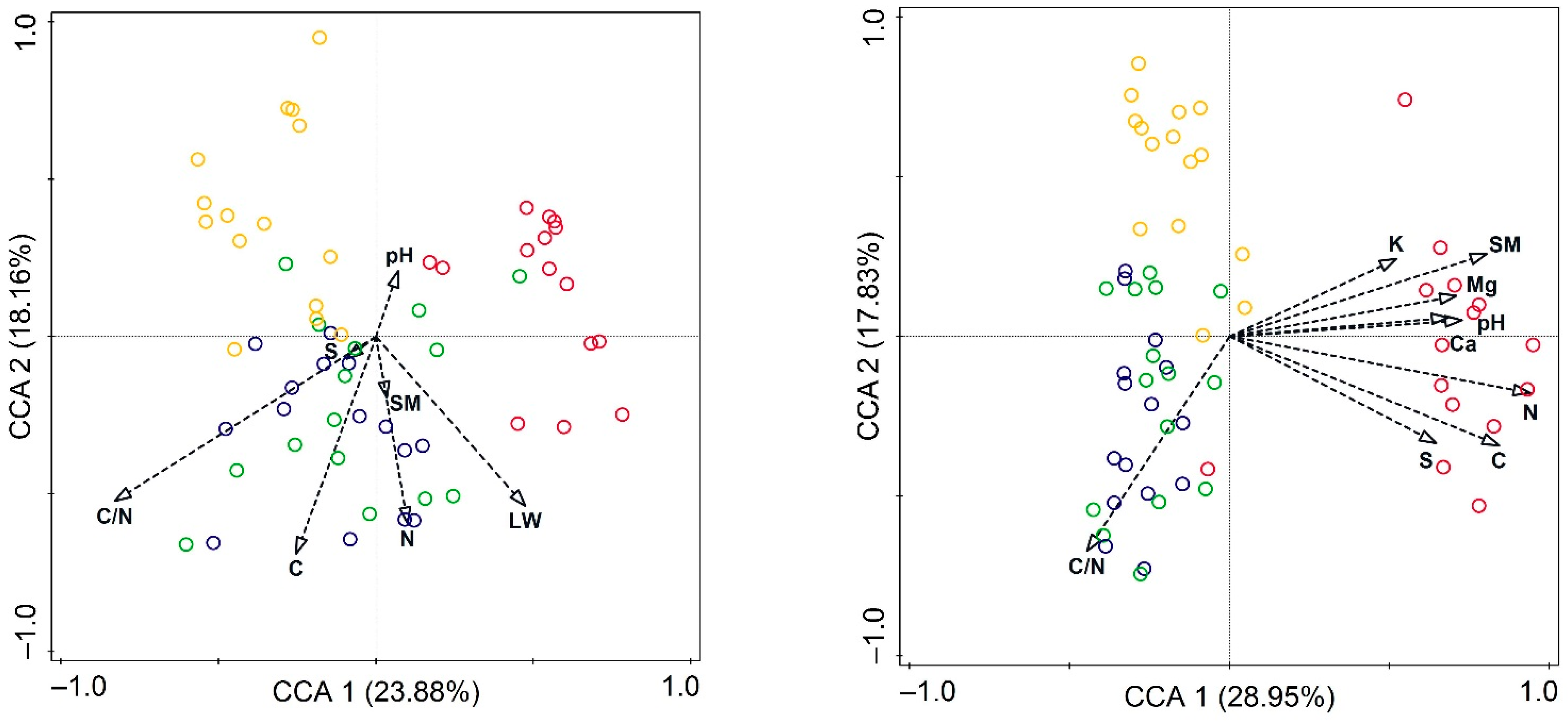
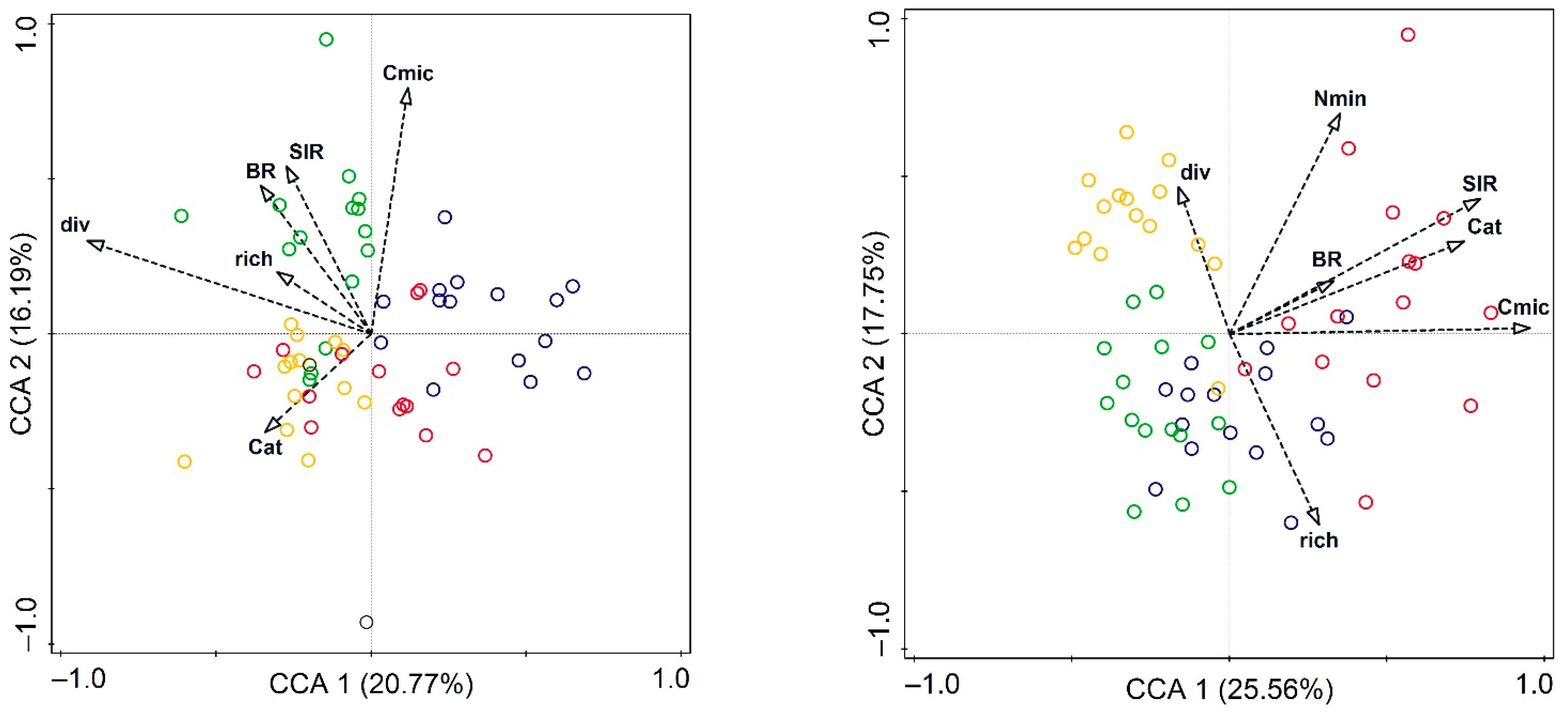
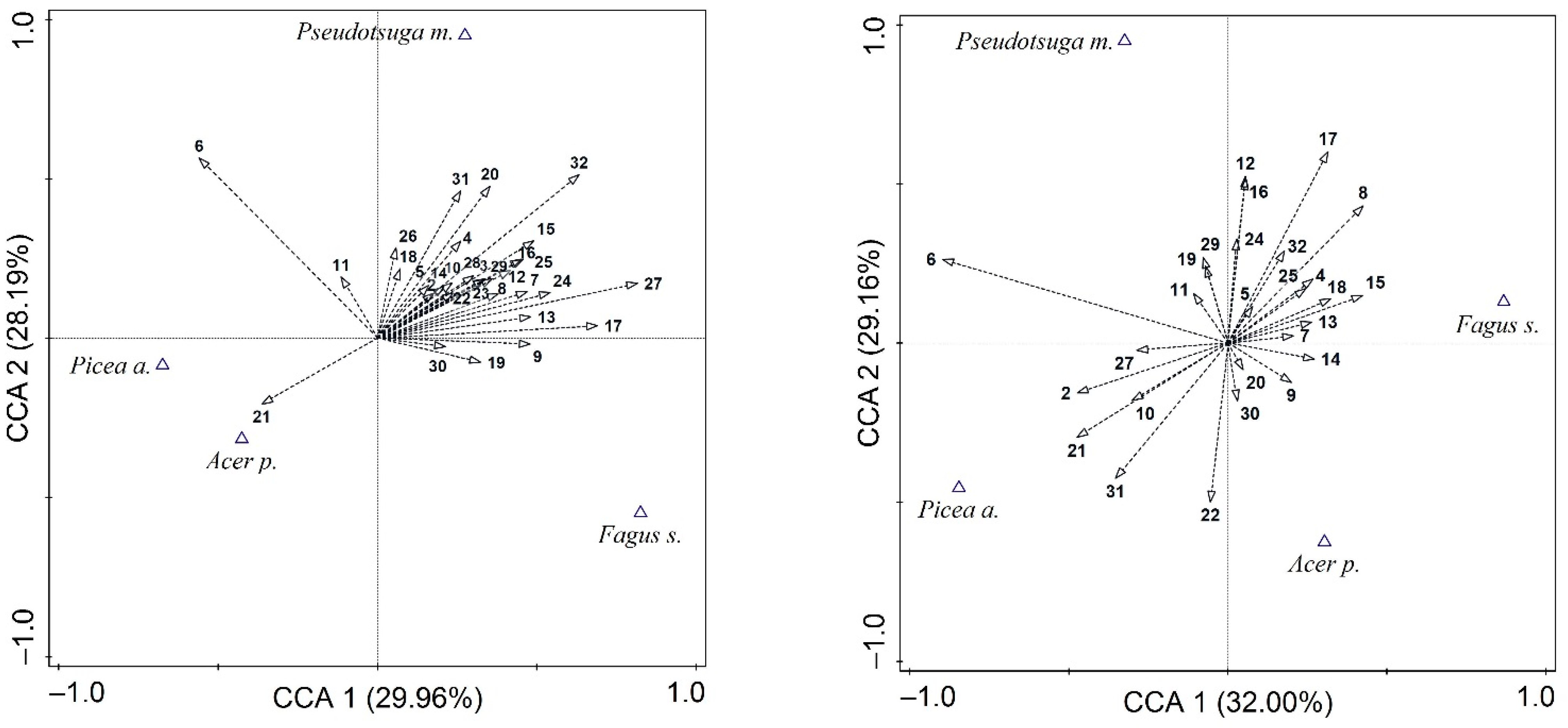
3. Results
3.1. Soil Chemical Properties
3.2. Soil Microbial Characteristics
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Augusto, L.; Ranger, J.; Binkley, D.; Rothe, A. Impact of several common tree species of European temperate forests on soil fertility. Ann. For. Sci. 2002, 59, 233–253. [Google Scholar] [CrossRef] [Green Version]
- Bonan, G.B. Forests and climate change: Forcings, feedbacks, and the climate benefits of forests. Science 2008, 320, 1444–1449. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gömöryová, E.; Hrivnák, R.; Janišová, M.; Ujházy, K.; Gömöry, D. Changes of the functional diversity of soil microbial community during the colonization of abandoned grassland by a forest. Appl. Soil. Ecol. 2009, 43, 191–199. [Google Scholar] [CrossRef]
- Augusto, L.; De Schrijver, L.A.; Vesterdal, L.; Smolander, A.; Prescott, C.; Ranger, J. Influences of evergreen gymnosperm and deciduous angiosperm tree species on the functioning of temperate and boreal forests. Biol. Rev. 2015, 90, 444–466. [Google Scholar] [CrossRef] [PubMed]
- Haesen, S.; Lembrects, J.J.; De Frenne, P.; Lenoir, J.; Aalto, J.; Ashcroft, M.B.; Kopecký, M.; Luoto, M.; Maclean, I.; Nijs, I.; et al. Forest Temp–Sub-canopy microclimate temperatures of European forests. Glob. Chang. Biol. 2021, 27, 6307–6319. [Google Scholar] [CrossRef] [PubMed]
- Finzi, A.C.; Canham, C.D. Non-additive effects of litter mixtures on net N mineralization in a southern New England forest. For. Ecol. Manag. 1998, 105, 129–136. [Google Scholar] [CrossRef]
- Berg, B. Litter decomposition and organic matter turnover on northern forest soils. For. Ecol. Man. 2000, 133, 13–22. [Google Scholar] [CrossRef]
- Smolander, A.; Kitunen, V. Soil microbial activities and characteristics of dissolved organic C and N in relation to tree species. Soil Biol. Biochem. 2002, 34, 651–660. [Google Scholar] [CrossRef]
- Saetre, P.; Bååth, E. Spatial variation and patterns of soil microbial community structure in a mixed spruce-birch stand. Soil Biol. Biochem. 2000, 32, 909–917. [Google Scholar] [CrossRef]
- Vesterdal, L.; Schmidt, I.K.; Callesen, I.; Nilsson, L.O.; Gundersen, P. Carbon and nitrogen in forest floor and mineral soil under six common European tree species. For. Ecol. Manag. 2008, 255, 35–48. [Google Scholar] [CrossRef]
- Brand, D.G.; Kehoe, P.; Connors, M. Coniferous afforestation leads to soil acidification in central Ontario. Can. J. For. Res. 1986, 16, 1389–1391. [Google Scholar] [CrossRef]
- Zwanzig, L.; Zwanzig, M.; Sauer, M. Outcomes of a quantitative analysis of 48 soil chronosequence studies in humid mid and high latitudes: Importance of vegetation in driving podsolization. Catena 2021, 196, 104821. [Google Scholar] [CrossRef]
- Mareschal, L.; Bonnaud, P.; Turpault, M.P.; Ranger, J. Impact of common European tree species on the chemical and physicochemical properties of fine earth: An unusual pattern. Soil Sci. 2010, 61, 14–23. [Google Scholar] [CrossRef]
- Achilles, F.; Tischer, A.; Bernhardt-Römermann, M.; Heinze, M.; Reinhardt, F.; Makeschin, F.; Michalzik, B. European beech leads to more bioactive humus forms but stronger mineral soil acidification as Norway spruce and Scots pine—Results of a repeated site assessment after 63 and 82 years of forest conversion in Central Germany. For. Ecol. Manag. 2021, 483, 118769. [Google Scholar] [CrossRef]
- Šály, R.; Capuliak, J.; Pavlenda, P. Vrchdobroč—Sľubný objekt k štúdiu pôdotvorného procesu pod vplyvom zalesnenia. In Diagnostika, Klasifikácia a Mapovanie Pôd; Sobocká, J., Ed.; VÚPOP: Bratislava, Slovakia, 2011; pp. 25–31. (In Slovak) [Google Scholar]
- Alef, K.; Nannipieri, P. Methodenhandbuch Bodenmikrobiologie; Academic Press: London, UK, 1995; 576p. [Google Scholar]
- Khazijev, F.C. Fermentativnaja Aktivnost’ Počv; Metodičeskoje Posobje: Moskva, Russia, 1976; p. 180. (In Russian) [Google Scholar]
- Islam, K.R.; Weil, R.R. Microwave irradiation of soil for routine measurement of microbial biomass carbon. Biol. Fertil. Soils 1998, 27, 408–416. [Google Scholar] [CrossRef]
- Kandeler, E. Bestimmung der N-mineralisation im anaeroben Brutversuch. In Bodenbiologische Arbeitsmethoden; Schinner, F., Ohlinger, R., Kandeler, E., Margesin, E.R., Eds.; Springer: Heidelberg, Germany, 1993; pp. 160–161. (In German) [Google Scholar]
- Insam, H. A new set of substrates proposed for community characterization in environmental samples. In Microbial Communities: Functional Versus Structural Approaches; Insam, H., Rangger, A., Eds.; Springer: Berlin, Germany, 1997; pp. 260–261. [Google Scholar]
- Hill, M.O. Diversity and evenness: A unifying notation and its consequences. Ecology 1973, 54, 427–432. [Google Scholar] [CrossRef] [Green Version]
- StatSoft, Inc. STATISTICA Data Analysis Software System, Version 12.0; StatSoft, Inc.: Tulsa, OH, USA, 2013. [Google Scholar]
- Ter Braak, C.J.F.; Smilauer, P. CANOCO Reference Manual and User’s Guide to Canoco for Windows: Software for Canonical Community Ordination (Version 4); Centre of Biometry Wageningen: Winnipeg, MA, Canada, 2002; p. 353. [Google Scholar]
- Lal, R. Forest soils and carbon sequestration. For. Ecol. Manag. 2005, 220, 242–258. [Google Scholar] [CrossRef]
- Armolaitis, K.; Aleinikoviene, J.; Baniuniene, A.; Lubyte, J.; Zekaite, V. 2007 Carbon sequestration and nitrogen status in Arenosols following afforestation or following abandonment of arable land. Balt. For. 2007, 13, 169–178. [Google Scholar]
- Cerli, C.; Celi, L.; Kaiser, K.; Guggenberger, G.; Johansson, M.B.; Cignetti, A.; Zanini, E. Changes in humic substances along an age sequence of Norway spruce stands planted on former agricultural land. Org. Geochem. 2008, 39, 1269–1280. [Google Scholar] [CrossRef]
- Kuzyakov, Y. Sources of CO2 efflux from soil and review of partitioning methods. Soil Biol. Biochem. 2006, 38, 425–448. [Google Scholar] [CrossRef]
- Hiltbrunner, D.; Zimmermann, S.; Hagedorn, F. Afforestation with Norway spruce on a subalpine pasture alters carbon dynamics but only moderately affects soil carbon storage. Biogeochemistry 2013, 115, 251–266. [Google Scholar] [CrossRef] [Green Version]
- Guidi, C.; Magid, J.; Rodeghiero, M.; Gianelle, D.; Vesterdal, L. Effects of forest expansion on mountain grassland: Changes within soil organic carbon fractions. Plant Soil 2014, 385, 373–387. [Google Scholar] [CrossRef]
- Saiz, G.; Bird, M.; Wurster, C.; Quesada, C.A.; Ascough, P.L.; Domingues, T.; Schrodt, F.; Schwarz, M.; Feldpausch, T.R.; Veenendaal, E.M.; et al. The influence of C3 and C4 vegetation on soil organic matter dynamics in contrasting semi-natural tropical ecosystems. Biogeosciences 2015, 12, 5041–5059. [Google Scholar] [CrossRef] [Green Version]
- Ortiz, C.; Fernandez-Alonsó, M.J.; Kitzler, B.; Díaz-Pines, E.; Saiz, G.; Rubio, A.; Benito, M. Variations in soil aggregation, microbial community structure and soil organic matter cycling associated to long-term afforestation and woody encroachment in a Mediterranean alpine ecotone. Geoderma 2022, 405, 115450. [Google Scholar] [CrossRef]
- Hübblová, L.; Frouz, J. Contrasting effect of coniferous and broadleaf trees on soil carbon storage during reforestation of forest soils and afforestation of agricultural and post-mining soils. J. Environ. Manag. 2021, 290, 112567. [Google Scholar] [CrossRef] [PubMed]
- Peng, Y.; Holmstrup, M.; Kappel Schmidt, I.; De Schrijver, A.; Schelfhout, S.; Heděnec, P.; Zheng, H.; Ruggiero Bachega, L.; Yue, K.; Vesterdal, L. Litter quality, mycorrhizal association, and soil properties regulate effects of tree species on the soil fauna community. Geoderma 2022, 407, 115570. [Google Scholar] [CrossRef]
- Frouz, L.; Livečková, M.; Albrechtová, J.; Chroňáková, A.; Cajthaml, T.; Pizl, V.; Háněl, L.; Starý, J.; Baldrian, P.; Lhotáková, Z.; et al. Is the effect of trees on soil properties mediated by soil fauna? A case study from post-mining sites. For. Ecol. Manag. 2013, 309, 87–95. [Google Scholar] [CrossRef]
- Tedersoo, L.; Bahram, M.; Cajthaml, T.; Polme, S.; Hiiesalu, I.; Anslan, S.; Harend, H.; Buegger, F.; Pritsch, K.; Koricheva, J.; et al. Tree diversity and species identity effects on soil fungi, protists and animals are context dependent. ISME J. 2016, 10, 346–362. [Google Scholar] [CrossRef] [Green Version]
- Curiel Yuste, J.; Barba, J.; Fernández-Gonzalez, J.A.; Fernández-López, M.; Mattana, S.; Nolis, P.; Lloret, F. Changes in soil bacterial community triggered by drought-induced gap succession preceded changes in soil C stocks and quality. Ecol. Evol. 2012, 2, 3016–3031. [Google Scholar] [CrossRef] [Green Version]
- Wang, K.; Zhang, Y.W.; Tang, Z.S.; Shangguan, Z.P.; Chang, F.; Jia, F.; Chen, Y.P.; He, X.H.; Shi, W.Y.; Deng, L. Effects of grassland afforestation on structure and function of soil bacterial and fungal communities. Sci. Total Environ. 2019, 676, 396–406. [Google Scholar] [CrossRef]
- Ding, L.L.; Wang, P.C. Afforestation suppresses soil nitrogen availability and soil multifunctionality on a subtropical grassland. Sci. Total Environ. 2021, 761, 143663. [Google Scholar] [CrossRef] [PubMed]
- Hornung, M. Acidification of soils by trees and forests. Soil Use Manag. 1985, 1, 24–27. [Google Scholar] [CrossRef]
- Côté, B.; Fyles, J.W. Nutrient concentration and acid–base status of leaf litter of tree species characteristic of the hardwood forest of southern Quebec. Can. J. For. Res. 1994, 24, 192–196. [Google Scholar] [CrossRef]
- Guckland, A.; Jacob, M.; Flessa, H.; Thomas, F.M.; Leuschner, C. Acidity, nutrient stocks, and organic-matter content in soils of a temperate deciduous forest with different abundance of European beech (Fagus sylvatica L.). J. Plant Nutr. Soil Sci. 2009, 172, 500–511. [Google Scholar] [CrossRef]
- Langenbruch, C.; Helfrich, M.; Flessa, H. Effects of beech (Fagus sylvatica), ash (Fraxinus excelsior) and lime (Tilia spec.) on soil chemical properties in a mixed deciduous forest. Plant Soil 2012, 352, 389–403. [Google Scholar] [CrossRef] [Green Version]
- Desie, E.; Vancampenhout, K.; Heyens, K.; Hlava, J.; Verheyen, K.; Muys, B. Forest conversion to conifers induces a regime shift in soil process domain affecting carbon stability. Soil Biol. Biochem. 2019, 136, 107540. [Google Scholar] [CrossRef]
- Haghverdia, K.; Kooch, Y. Effects of diversity of tree species on nutrient cycling and soil-related processes. Catena 2019, 178, 335–344. [Google Scholar] [CrossRef]
- Eisenhauer, N.; Bessler, H.; Engels, C.; Gleixner, G.; Habekost, M.; Milcu, A.; Partsch, S.; Sabais, A.C.W.; Scherber, C.; Steinbeiss, S.; et al. Plant diversity effects on soil microorganisms support the singular hypothesis. Ecology 2010, 91, 485–496. [Google Scholar] [CrossRef]
- Pfeiffer, B.; Fender, A.C.; Lasota, A.; Hertel, D.; Jungkunst, H.F.; Daniel, R. Leaf litter is the main driver for changes in bacterial community structures in the rhizosphere of ash and beech. Appl. Soil Ecol. 2013, 72, 150–160. [Google Scholar] [CrossRef]
- Chen, C.R.; Condron, L.M.; Xu, Z.H. Impacts of grassland afforestation with coniferous trees on soil phosphorus dynamics and associated microbial processes: A review. For. Ecol. Manag. 2008, 255, 396–409. [Google Scholar] [CrossRef]
- Chen, X.D.; Condron, L.M.; Dunfield, K.E.; Wakelin, S.A.; Chen, L.J. Impact of grassland afforestation with contrasting tree species on soil phosphorus fractions and alkaline phosphatase gene communities. Soil Biol. Biochem. 2021, 159, 108274. [Google Scholar] [CrossRef]
- Zeller, B.; Legout, A.; Bienaimé, S.; Gratia, B.; Santenoise, P.; Bonnaud, B.; Ranger, J. Douglas fir stimulates nitrifcation in French forest soils. Sci. Rep. 2019, 9, 10687. [Google Scholar] [CrossRef] [PubMed]
- Schmid, M.; Pautasso, M.; Holdenrieder, O. Ecological consequences of Douglas fir (Pseudotsuga menziesii) cultivation in Europe. Eur. J. Forest. Res. 2014, 133, 13–29. [Google Scholar] [CrossRef] [Green Version]




{kind=link}
{kind=link}
{kind=link}
| Physico-Chemical Properties | Horizon | P | Picea | Pseudotsuga | Fagus | Acer |
|---|---|---|---|---|---|---|
| pH/CaCl2 | O | <0.1 | 4.85 ± 0.40 a | 5.08 ± 0.35 a | 5.06 ± 0.21 a | 5.06 ± 0.35 a |
| A | <0.001 | 3.73 ± 0.09 b | 3.94 ± 0.28 b | 3.89 ± 0.23 b | 4.28 ± 0.26 a | |
| Soil moisture (%) | O | <0.01 | 29.71 ± 6.90 c | 61.62 ± 54.34 a | 32.47 ± 12.37 bc | 37.63 ± 13.76 b |
| A | <0.001 | 17.10 ± 3.12 b | 15.29 ± 5.52 b | 23.28 ± 7.22 ab | 36.80 ± 10.18 a | |
| C (%) | O | <0.001 | 32.37 ± 10.05 a | 32.53 ± 8.64 a | 22.61 ± 7.79 b | 21.42 ± 5.01 b |
| A | <0.001 | 4.60 ± 0.85 b | 4.15 ± 1.05 b | 3.73 ± 0.71 b | 7.19 ± 1.07 a | |
| N (%) | O | <0.001 | 1.65 ± 0.48 a | 1.70 ± 0.40 a | 1.22 ± 0.35 b | 1.48 ± 0.25 ab |
| A | <0.001 | 0.37 ± 0.06 b | 0.35 ± 0.08 b | 0.34 ± 0.05 b | 0.67 ± 0.09 a | |
| S (%) | O | 0.92 | 0.24 ± 0.06 a | 0.24 ± 0.05 a | 0.23 ± 0.23 a | 0.21 ± 0.05 a |
| A | <0.001 | 0.07 ± 0.03 b | 0.07 ± 0.02 b | 0.06 ± 0.01 b | 0.10 ± 0.02 a | |
| C:N ratio | O | <0.001 | 19.40 ± 1.41 a | 18.97 ± 2.07 a | 18.19 ± 1.46 a | 14.37 ± 1.05 b |
| A | <0.001 | 12.46 ± 0.92 a | 11.87 ± 1.02 a | 10.81 ± 0.67 b | 10.82 ± 0.70 b | |
| Litter weight (kg·m−2) | O | <0.001 | 1.46 ± 0.36 a | 1.24 ± 0.31 b | 0.85 ± 0.27 b | 1.45 ± 0.44 a |
| Ca2+ (mg·kg−1) | A | <0.001 | 475.1 ± 168.6 b | 742.8 ± 413.1 b | 692.7 ± 163.5 b | 1272.5 ± 379.5 a |
| Mg2+ (mg·kg−1) | A | <0.001 | 116.0 ± 33.9 b | 131.2 ± 21.9 b | 135.6 ± 13.3 b | 178.3 ± 29.8 a |
| K+ (mg·kg−1) | A | <0.001 | 49.13 ± 6.37 b | 56.11 ± 36.41 b | 67.23 ± 15.61 b | 86.99 ± 27.51 a |
| Microbial Properties | Horizon | P | Picea | Pseudotsuga | Fagus | Acer |
|---|---|---|---|---|---|---|
| Basal respiration (µg CO2·g−1·h−1) | O | <0.001 | 1.37 ± 0.49 b | 4.47 ± 2.89 a | 1.88 ± 1.51 b | 2.61 ± 2.38 ab |
| A | 0.11 | 0.18 ± 0.04 a | 0.16 ± 0.09 a | 0.20 ± 0.09 a | 0.42 ± 0.63 a | |
| Substrate-induced respiration (µg CO2·g−1·h−1) | O | <0.001 | 5.02 ± 1.97 b | 22.00 ± 23.62 a | 5.37 ± 2.42 b | 9.64 ± 6.67 b |
| A | <0.001 | 1.09 ± 0.61 b | 0.63 ± 0.29 b | 1.11 ± 0.46 b | 2.40 ± 0.82 a | |
| Catalase activity (ml O2·g−1·min−1) | O | <0.05 | 5.26 ± 0.93 b | 6.02 ± 2.26 ab | 6.62 ± 1.26 ab | 6.76 ± 1.39 a |
| A | <0.001 | 0.65 ± 0.20 a | 0.56 ± 0.17 a | 0.62 ± 0.21 a | 1.06 ± 0.26 a | |
| N mineralization (µg NH4+-N·g−1·d−1) | A | <0.001 | 0.74 ± 0.50 b | 1.41 ± 1.04 b | 4.00 ± 2.17 a | 5.38 ± 4.36 a |
| Microbial biomass carbon (µg·g−1) | O | <0.001 | 6440 ± 1250 ab | 7869 ± 3339 a | 4338 ± 1047 c | 4894 ± 1747 bc |
| A | <0.001 | 568.2 ± 169.7 b | 420.3 ± 99.2 c | 361.8 ± 105.3 c | 912.4 ± 185.4 a | |
| Richness of functional groups | O | <0.001 | 26.87 ± 2.13 c | 28.93 ± 1.79 a | 27.47 ± 1.99 ab | 28.40 ± 1.59 ab |
| A | <0.001 | 27.80 ± 1.01 a | 26.87 ± 2.13 a | 25.07 ± 1.79 b | 26.93 ± 1.75 a | |
| Diversity of functional groups | O | <0.001 | 10.48 ± 2.63 b | 17.60 ± 1.97 a | 16.55 ± 2.33 a | 13.53 ± 3.01 b |
| A | 0.01 | 11.45 ± 1.88 c | 15.18 ± 2.63 a | 12.99 ± 1.37 b | 11.18 ± 1.78 c |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gere, R.; Kočiš, M.; Židó, J.; Gömöry, D.; Gömöryová, E. Differential Effects of Tree Species on Soil Microbiota 45 Years after Afforestation of Former Pastures. Diversity 2022, 14, 515. https://doi.org/10.3390/d14070515
Gere R, Kočiš M, Židó J, Gömöry D, Gömöryová E. Differential Effects of Tree Species on Soil Microbiota 45 Years after Afforestation of Former Pastures. Diversity. 2022; 14(7):515. https://doi.org/10.3390/d14070515
Chicago/Turabian StyleGere, Richard, Mikuláš Kočiš, Ján Židó, Dušan Gömöry, and Erika Gömöryová. 2022. "Differential Effects of Tree Species on Soil Microbiota 45 Years after Afforestation of Former Pastures" Diversity 14, no. 7: 515. https://doi.org/10.3390/d14070515